Функциональный ответ - Functional response
А функциональный ответ в экология скорость приема потребитель как функция из плотность пищи (количество еды, доступное в данном экотоп ). Это связано с числовой ответ, какой воспроизведение показатель потребителя в зависимости от плотности пищи. Следующий К. С. Холлинг Функциональные реакции обычно делятся на три типа, которые называются типом Холлинга I, II и III.[1]
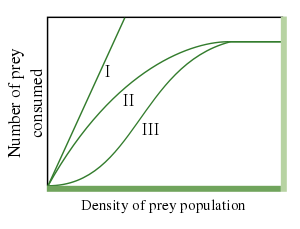
Тип I
Функциональная реакция типа I предполагает линейное увеличение скорости приема пищи в зависимости от плотности пищи либо для всех плотностей пищи, либо только для плотностей пищи вплоть до максимума, за пределами которого скорость приема является постоянной. Линейное увеличение предполагает, что время, необходимое потребителю для обработки пищевого продукта, незначительно или что потребление пищи не мешает поиску пищи. Функциональный ответ типа I используется в Лотка – Вольтерра модель хищник – жертва. Это был первый описанный тип функционального ответа, а также самый простой из трех детализированных в настоящее время функциональных ответов.
Тип II
Функциональная реакция типа II характеризуется замедлением скорости приема пищи, что следует из предположения, что потребитель ограничен своей способностью обрабатывать пищу. Функциональный ответ типа II часто моделируется прямоугольная гипербола, например, как по уравнению диска Холлинга,[2] который предполагает, что обработка пищи и поиск пищи являются взаимоисключающими действиями. Уравнение


куда ж обозначает скорость поступления и р обозначает плотность пищи (или ресурсов). Скорость, с которой потребитель встречает продукты питания на единицу плотности пищи, называется скоростью атаки. а. Среднее время, затрачиваемое на обработку продукта, называется временем обработки, час. Подобные уравнения представляют собой Уравнение Моно для роста микроорганизмы и Уравнение Михаэлиса – Ментен на скорость ферментативных реакций.
В примере с волки и карибу, поскольку количество карибу увеличивается при постоянном сохранении волков, количество убитых карибу увеличивается, а затем стабилизируется. Это связано с тем, что доля убитых карибу на одного волка уменьшается с увеличением плотности карибу. Чем выше плотность карибу, тем меньше доля убитых карибу на одного волка. Если объяснить несколько иначе, при очень высокой плотности карибу волкам нужно очень мало времени, чтобы найти добычу, и они тратят почти все свое время на ловлю добычи и очень мало времени на поиски. Затем волки насыщаются, и общее количество убитых карибу достигает плато.[3]
Тип III
Функциональная реакция типа III аналогична типу II в том, что при высокой плотности добычи происходит насыщение. Но теперь, при низких уровнях плотности добычи, графическая зависимость количества съеденной жертвы и плотности популяции жертвы является более чем линейно возрастающей функцией добычи, потребляемой хищниками. Эта ускоряющая функция носит в основном описательный характер и часто оправдывается временем обучения, переключением добычи или комбинацией обоих явлений, но функциональная реакция типа III не имеет строгого теоретического обоснования функциональной реакции типа II.
Время обучения определяется как естественное улучшение эффективности поиска и нападения хищников или естественное улучшение их эффективности в обращении с увеличением плотности добычи. Представьте себе, что плотность добычи настолько мала, что вероятность встречи хищника с этой добычей крайне мала. Поскольку хищник находит добычу так редко, у него не было достаточно опыта, чтобы разработать лучшие способы поимки и подчинения этого вида добычи. Холлинг идентифицировал этот механизм в землеройки и олени мыши питаясь пилильщики. При небольшом количестве коконов пилильщика на акр у мышей-оленей особенно наблюдался экспоненциальный рост количества коконов, потребляемых на одну особь, по мере увеличения плотности коконов. Характерная точка насыщения функционального ответа III типа наблюдалась и у мышей-оленей. При определенной плотности коконов на акр скорость потребления мышей-оленями достигла величины насыщения, поскольку плотность коконов продолжала увеличиваться.[2]
Смена добычи включает два или более видов добычи и один вид хищников. Когда все виды добычи имеют одинаковую плотность, хищник будет без разбора выбирать между видами добычи. Однако, если плотность одного из видов добычи уменьшается, хищник начинает выбирать другой, более распространенный вид добычи с большей частотой. Мердок продемонстрировал этот эффект с гуппи охотятся на тубифицидов и плодовые мошки. По мере того, как численность плодовых мух уменьшалась, гуппи переключились с кормления плодовых мух на поверхности воды на питание более многочисленными тубифицидами вдоль дна.[4]
Смотрите также
Рекомендации
- ^ Холлинг, С. С. (май 1959 г.). «Компоненты хищничества, выявленные в результате изучения хищничества европейского соснового пилильщика на мелких млекопитающих». Канадский энтомолог. 91 (5): 293–320. Дои:10.4039 / Ent91293-5.
- ^ а б Холлинг, С. С. (июль 1959 г.). «Некоторые характеристики простых видов хищничества и паразитизма». Канадский энтомолог. 91 (7): 385–98. Дои:10.4039 / Ent91385-7.
- ^ а б Дейл, Брюс В .; Adams, Layne G .; Бойер, Р. Терри (1994). «Функциональный ответ волков, охотящихся на бесплодных карибу в экосистеме с множеством жертв». Журнал экологии животных. 63: 644–652.
- ^ Мердок, Уильям У. (1977). «Стабилизирующие эффекты пространственной неоднородности в системах хищник-жертва». Теоретическая популяционная биология. 11: 252–73.