Трехцепочечная ДНК - Triple-stranded DNA
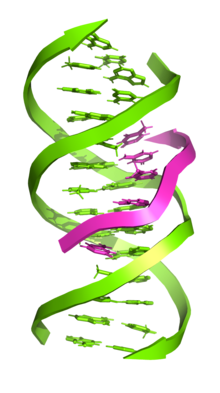
Трехцепочечная ДНК (также известен как H-ДНК или Триплекс-ДНК) это Структура ДНК в котором три олигонуклеотиды обвивают друг друга и образуют тройная спираль. В трехцепочечной ДНК третья цепь связывается с B-форма ДНК (через Спаривание оснований Уотсона – Крика ) двойная спираль, образуя Hoogsteen пар оснований или обратные водородные связи Хугстина.
Структуры

Спаривание оснований Хугстина
Тимин (т) азотистое основание может привязать к Спаривание оснований Уотсона – Крика T-A путем формирования Водородная связь Хугстина. Водород тимина связывается с аденозином (A) исходной двухцепочечной ДНК с образованием триплета оснований T-A * T.[1] В кислых условиях протонированный цитозин, представленный как C +, может образовывать базовый триплет с парой C-G через Спаривание оснований Хугстина, образуя C-G * C +. Пары оснований TA * T и CG * C + являются наиболее стабильными парами триплет-основание, которые могут образовываться, в то время как TA * G и CG * G являются наиболее дестабилизированными парами триплет-основание.[2]
Межмолекулярные и внутримолекулярные образования
Существует два класса триплексной ДНК: межмолекулярные и внутримолекулярные образования. Межмолекулярный триплекс относится к образованию триплекса между дуплексом и другой (третьей) цепью ДНК. Третья цепь может происходить либо из соседней хромосомы, либо из триплексообразующего олигонуклеотида (TFO). Внутримолекулярная триплексная ДНК образуется из дуплекса с гомопурин и гомопиримидин пряди с зеркальной симметрией повторения.[3] Степень суперспирализация в ДНК влияет на количество происходящего внутримолекулярного образования триплекса.[4] Существует два различных типа внутримолекулярной триплексной ДНК: H-ДНК и H * -ДНК. Образование H-ДНК стабилизируется в кислых условиях и в присутствии двухвалентные катионы такие как Mg2+. В этой конформации гомопиримидиновая цепь в дуплексе изгибается назад, чтобы связываться с пуриновой цепью параллельным образом. Базовые триады, используемые для стабилизации этой конформации, - это T-A * T и C-G * C.+. Цитозин этой триады оснований должен быть протонирован, чтобы образовать эту внутримолекулярную тройную спираль, поэтому эта конформация стабилизируется в кислых условиях.[5] H * -ДНК имеет благоприятные условия образования при нейтральном pH и в присутствии двухвалентных катионов.[4] Эта внутримолекулярная конформация образуется в результате связывания гомопуриновой и пуриновой цепей дуплекса антипараллельным образом. Он стабилизируется базовыми триплетами T-A * A и C-G * G.[3][5]
Функция
Трехцепочечная ДНК участвует в регуляции нескольких генов. Например, c-мой с Ген был подвергнут обширным мутациям, чтобы изучить роль, которую триплексная ДНК, по сравнению с линейной последовательностью, играет в регуляции гена. А с-мой с Промоторный элемент, называемый чувствительным к нуклеазе элементом или NSE, может образовывать тандемные внутримолекулярные триплексы типа H-ДНК и имеет мотив повторяющейся последовательности (ACCCTCCCC)4. Мутировавший NSE исследовали на транскрипционную активность и на его способность формировать внутри- и межмолекулярный триплекс. Транскрипционная активность мутантных NSE может быть предсказана по способности элемента образовывать H-ДНК, а не по количеству повторов, положению или количеству мутантных пар оснований. Следовательно, ДНК может быть динамическим участником транскрипции гена c-myc.[6]
Триплекс-образующие олигонуклеотиды (TFO)
TFO представляют собой короткие (≈15-25 нуклеотидов) цепи нуклеиновых кислот, которые связываются в большой бороздке двухцепочечной ДНК с образованием внутримолекулярных триплексных структур ДНК. Есть некоторые свидетельства того, что они также могут модулировать активность генов. in vivo. В пептидная нуклеиновая кислота (PNA) сахарно-фосфатный остов ДНК заменяется протеиноподобным остовом. ПНК образуют P-петли, взаимодействуя с дуплексной ДНК, образуя триплекс с одной цепью ДНК, вытесняя другую. Предполагается, что очень необычная рекомбинация или параллельные триплексы, или R-ДНК, образуются под белком RecA в ходе гомологичной рекомбинации.[7]
TFO специфически связываются с гомопурин-гомопиримидиновыми участками, которые часто встречаются в промоторных и интронных последовательностях генов, влияющих на передачу сигналов в клетке.[8] TFO могут ингибировать транскрипцию путем связывания с высокой специфичностью со спиралью ДНК, тем самым блокируя связывание и функцию факторов транскрипции для определенных последовательностей. Путем введения TFO в клетку (посредством трансфекции или другими способами) можно контролировать экспрессию определенных генов.[9] Это приложение имеет новые возможности для сайт-специфического мутагенеза и генной терапии. В клетках рака простаты человека фактор транскрипции Ets2 сверхэкспрессирован и, как считается, способствует росту и выживанию клеток в таком избытке. Carbone et al. разработали специфичный для последовательности TFO для последовательности промотора Ets2, который подавлял экспрессию гена и приводил к замедлению роста и гибели клеток.[10] Changxian et al. также представили TFO, нацеленный на промоторную последовательность bcl-2, гена, ингибирующего апоптоз.[11]
Наблюдаемое ингибирование транскрипции также может иметь негативные последствия для здоровья, например, его роль в рецессивном аутосомном гене Атаксия Фридрейха.[12] При атаксии Фредрика образование триплексной ДНК нарушает экспрессию интрон 1 из Ген FXN. Это приводит к дегенерации нервной системы и спинного мозга, нарушая подвижность конечностей.[13] Было показано, что для борьбы с этой триплексной нестабильностью белки эксцизионной репарации нуклеотидов (NER) распознают и восстанавливают трехцепочечные структуры ДНК, восстанавливая полную доступность ранее ингибированного и нестабильного гена.[14]
Генетическая нестабильность
Были проведены значительные исследования биологических последствий, связанных с присутствием триплексной ДНК, а точнее H-ДНК, в областях основных точек разрыва (Mbr) и точек разрыва в двух цепях (DBS) определенных генов.
Например, в дополнение к другим последовательностям ДНК, отличным от B, обнаруженным рядом с промотором P1 c-MYC были обнаружены последовательности, образующие H-ДНК полипуринового зеркального повтора, которые связаны с основными горячими точками в этой области. Случаи генетической нестабильности также наблюдались у потомства F1 трансгенных мышей после включения человеческих H-ДНК-образующих последовательностей в паре с Z-ДНК последовательности в их геномы, о нестабильности которых ранее не сообщалось.[15] Кроме того, формирование Р.R.Y. триплексные конформации наблюдались на Mbr bcl-2 ген. Было высказано предположение, что образование этих структур вызывает транслокацию t (14; 18), наблюдаемую при многих раковых заболеваниях и большинстве фолликулярных лимфом. Это наблюдение привело к исследованиям, которые показали, что после блокирования образования H-ДНК можно наблюдать существенное уменьшение количества транслокаций путем небольшого изменения последовательности этой области.[15][16] Также наблюдали, что длинные участки GAA · TTC образуют очень стабильные триплексные структуры. Было показано, что взаимодействия между этими двумя триплексными структурами, называемыми липкой ДНК, прерывают транскрипцию X25, или ген фратаксина. Поскольку снижение уровня белка фратаксина связано с атаксией Фридрейха, предполагается, что формирование этой нестабильности является основой этого генетического заболевания.[17][18]
История

Трехцепочечные структуры ДНК были распространенными гипотезами в 1950-х годах, когда ученые изо всех сил пытались открыть истинную структурную форму ДНК. Уотсон и Крик (которые позже получили Нобелевскую премию за свою модель двойной спирали) изначально рассматривали модель тройной спирали, как и Полинг и Кори, которые опубликовали предложение о своей модели тройной спирали в 1953 году,[19][20] а также научный сотрудник Фрейзер.[21] Однако вскоре Уотсон и Крик выявили несколько проблем с этими моделями:
- Отрицательно заряженные фосфаты около оси отталкиваются друг от друга, оставляя вопрос, как трехцепочечная структура остается вместе.
- В модели тройной спирали (в частности, модели Полинга и Кори) некоторые из расстояния Ван-дер-Ваальса кажутся слишком маленькими.
Модель Фрейзера отличалась от модели Полинга и Кори тем, что в его модели фосфаты находятся снаружи, а основания - внутри, связанные водородными связями. Однако Уотсон и Крик сочли модель Фрейзера слишком неопределенной, чтобы специально комментировать ее недостатки.
Альтернативная трехцепочечная структура ДНК была описана в 1957 году.[22] Считалось, что это произошло только в одном in vivo биологический процесс: как промежуточный продукт при действии Кишечная палочка рекомбинационный фермент RecA. Его роль в этом процессе не изучена.[22]
Рекомендации
- ^ Ри С., Хан З., Лю К., Майлз Х. Т., Дэвис Д. Р. (декабрь 1999 г.). «Структура тройной спиральной ДНК с тройным дуплексным переходом». Биохимия. 38 (51): 16810–5. Дои:10.1021 / bi991811m. PMID 10606513.
- ^ Mergny JL, Sun JS, Rougée M, Montenay-Garestier T, Barcelo F, Chomilier J, Hélène C (октябрь 1991 г.). «Специфичность последовательности в формировании тройной спирали: экспериментальные и теоретические исследования влияния несовпадений на стабильность триплекса». Биохимия. 30 (40): 9791–8. Дои:10.1021 / bi00104a031. PMID 1911764.
- ^ а б Ussery DW, Sinden RR (июнь 1993 г.). «Влияние окружающей среды на уровень in vivo внутримолекулярной триплексной ДНК в Escherichia coli». Биохимия. 32 (24): 6206–13. Дои:10.1021 / bi00075a013. PMID 8512930.
- ^ а б Дайн А., Самадашвили Г.М., Миркин С.М. (декабрь 1992 г.). «Внутримолекулярные триплексы ДНК: необычные требования к последовательности и влияние на полимеризацию ДНК». Труды Национальной академии наук Соединенных Штатов Америки. 89 (23): 11406–10. Bibcode:1992PNAS ... 8911406D. Дои:10.1073 / пнас.89.23.11406. ЧВК 50559. PMID 1454828.
- ^ а б Лямичев В.И., Миркин С.М., Франк-Каменецкий М.Д. (февраль 1986 г.). «Структуры гомопурин-гомопиримидинового тракта в сверхспиральной ДНК». Журнал биомолекулярной структуры и динамики. 3 (4): 667–9. Дои:10.1080/07391102.1986.10508454. PMID 3271043.
- ^ Firulli AB, Maibenco DC, Kinniburgh AJ (апрель 1994 г.). «Способность промотора c-myc формировать триплекс определяет силу промотора». Архивы биохимии и биофизики. 310 (1): 236–42. Дои:10.1006 / abbi.1994.1162. PMID 8161210.
- ^ Франк-Каменецкий М.Д., Миркин С.М. (01.01.1995). «Триплексные структуры ДНК». Ежегодный обзор биохимии. 64: 65–95. Дои:10.1146 / annurev.bi.64.070195.000433. PMID 7574496.
- ^ Браздова М., Тихи В., Хельма Р., Бажантова П., Полашкова А., Крейчи А. и др. (2016). «p53 специфически связывает триплексную ДНК in vitro и в клетках». PLOS One. 11 (12): e0167439. Дои:10.1371 / journal.pone.0167439. ЧВК 5131957. PMID 27907175.
- ^ Грэм МК, Браун Т.Р., Миллер П.С. (апрель 2015 г.). «Нацеливание на ген рецептора андрогена человека с платинированными триплекс-образующими олигонуклеотидами». Биохимия. 54 (13): 2270–82. Дои:10.1021 / bi501565n. PMID 25768916.
- ^ Carbone GM, Napoli S, Valentini A, Cavalli F, Watson DK, Catapano CV (2004-08-03). «Триплексное ДНК-опосредованное подавление экспрессии Ets2 приводит к подавлению роста и апоптозу клеток рака простаты человека». Исследования нуклеиновых кислот. 32 (14): 4358–67. Дои:10.1093 / нар / гх744. ЧВК 514370. PMID 15314206.
- ^ Шен С., Раттат Д., Бак А., Мехрке Г., Полат Б., Рибберт Х., Ширрмейстер Х., Марен Б., Матушек С., Реске С. Н. (февраль 2003 г.). «Нацеливание bcl-2 на триплекс-образующий олигонуклеотид - перспективный носитель для генной лучевой терапии». Биотерапия рака и радиофармпрепараты. 18 (1): 17–26. Дои:10.1089/108497803321269296. PMID 12667305.
- ^ Сакамото Н., Честейн П.Д., Парневски П., Охима К., Пандольфо М., Гриффит Д.Д., Уэллс Р.Д. (апрель 1999 г.). «Липкая ДНК: свойства самоассоциации длинных повторов GAA.TTC в триплексных структурах R.R.Y от атаксии Фридрейха». Молекулярная клетка. 3 (4): 465–75. Дои:10.1016 / с1097-2765 (00) 80474-8. PMID 10230399.
- ^ Баколла А., Уэллс Р.Д. (апрель 2009 г.). «Конформации ДНК, отличные от B, как детерминанты мутагенеза и болезней человека». Молекулярный канцерогенез. 48 (4): 273–85. Дои:10.1002 / mc.20507. PMID 19306308.
- ^ Каушик Тивари М., Адаку Н., Пирт Н., Роджерс Ф.А. (сентябрь 2016 г.). «Триплексные структуры индуцируют двухцепочечные разрывы ДНК через коллапс репликационной вилки в клетках с дефицитом NER». Исследования нуклеиновых кислот. 44 (16): 7742–54. Дои:10.1093 / нар / gkw515. ЧВК 5027492. PMID 27298253.
- ^ а б Ван Г., Васкес К.М. (июль 2014 г.). «Влияние альтернативных структур ДНК на повреждение ДНК, восстановление ДНК и генетическую нестабильность». Ремонт ДНК. 19: 143–51. Дои:10.1016 / j.dnarep.2014.03.017. ЧВК 4216180. PMID 24767258.
- ^ Raghavan SC, Chastain P, Lee JS, Hegde BG, Houston S, Langen R, Hsieh CL, Haworth IS, Lieber MR (июнь 2005 г.). «Доказательства конформации триплексной ДНК в области основной точки разрыва bcl-2 транслокации t (14; 18)». Журнал биологической химии. 280 (24): 22749–60. Дои:10.1074 / jbc.M502952200. PMID 15840562.
- ^ Ван Г., Васкес К.М. (сентябрь 2004 г.). «Встречающиеся в природе последовательности, образующие H-ДНК, являются мутагенными в клетках млекопитающих». Труды Национальной академии наук Соединенных Штатов Америки. 101 (37): 13448–53. Дои:10.1073 / pnas.0405116101. ЧВК 518777. PMID 15342911.
- ^ Ветчер А.А., Напьерала М., Айер Р.Р., Честейн П.Д., Гриффит Д.Д., Уэллс Р.Д. (октябрь 2002 г.). «Липкая ДНК, длинный триплекс GAA.GAA.TTC, который образуется внутримолекулярно, в последовательности интрона 1 гена фратаксина». Журнал биологической химии. 277 (42): 39217–27. Дои:10.1074 / jbc.M205209200. PMID 12161437.
- ^ Полинг Л., Кори Р. Б. (февраль 1953 г.). «Предлагаемая структура нуклеиновых кислот». Труды Национальной академии наук Соединенных Штатов Америки. 39 (2): 84–97. Дои:10.1073 / pnas.39.2.84. ЧВК 1063734. PMID 16578429.
- ^ Полинг Л., Кори Р. Б. (февраль 1953 г.). «Структура нуклеиновых кислот». Природа. 171 (4347): 346. Дои:10.1038 / 171346a0. PMID 13036888.
- ^ Фрейзер Б. (1953). «Структура нуклеиновой кислоты дезоксирибозы». Журнал структурной биологии. 145 (3): 184–5. Дои:10.1016 / j.jsb.2004.01.001. PMID 14997898.
- ^ а б Фельзенфельд Г., Дэвис Д. Р., Рич А. (1957). «Формирование трехцепочечной полинуклеотидной молекулы». Журнал Американского химического общества. 79 (8): 2023–2024. Дои:10.1021 / ja01565a074.
дальнейшее чтение
- Рич А. (декабрь 1993 г.). «ДНК бывает разных форм». Ген. 135 (1–2): 99–109. Дои:10.1016/0378-1119(93)90054-7. PMID 8276285.
- Сойфер В.Н., Потаман В.Н. (1995). Тройные спиральные нуклеиновые кислоты. Нью-Йорк: Спрингер. ISBN 978-0-387-94495-1.
- Миллс М., Аримондо П.Б., Лакруа Л., Гарстье Т., Элен С., Кламп Н., Мергни Д.Л. (сентябрь 1999 г.). «Энергетика реакций вытеснения цепей в тройных спиралях: спектроскопическое исследование». Журнал молекулярной биологии. 291 (5): 1035–54. Дои:10.1006 / jmbi.1999.3014. PMID 10518941.
- Уотсон Дж. Д., Крик Ф. Х. (апрель 1953 г.). «Молекулярная структура нуклеиновых кислот; структура нуклеиновой кислоты дезоксирибозы». Природа. 171 (4356): 737–8. Bibcode:1953 г., природа. 171..737 Вт. Дои:10.1038 / 171737a0. PMID 13054692.