Курацин А - Curacin A
 | |
| Идентификаторы | |
|---|---|
| |
| Количество CAS | |
| PubChem CID | |
| ChemSpider | |
| КЕГГ | |
| ЧЭБИ | |
| ЧЭМБЛ | |
| Химические и физические данные | |
| Формула | C23ЧАС35NОS |
| Молярная масса | 373.60 г · моль−1 |
| 3D модель (JSmol ) | |
| |
| |
| (проверять) | |
Курацин А это гибрид поликетидсинтаза (ПКС) /негрибосомная пептидная синтаза (NRPS) полученный натуральный продукт производится изолированно от цианобактерии Lyngbya majuscula.[1] Курацин А принадлежит к семейству натуральных продуктов, включая ямайкамид, мупироцин и педерин у которых есть необычный терминал алкен. Кроме того, Curacin A содержит заметный тиазолин кольцо и уникальный циклопропил часть, которая важна для соединения биологическая активность.[1][2] Курацин А был охарактеризован как мощный антипролиферативный цитотоксический соединение с заметной противоопухолевой активностью для нескольких рак линии, включая рак почек, толстой кишки и груди.[2][3] Было показано, что курацин А взаимодействует с сайтами связывания колхицина на тубулине, что ингибирует полимеризацию микротрубочек, важный процесс деления и пролиферации клеток.[1][4]
Биосинтез
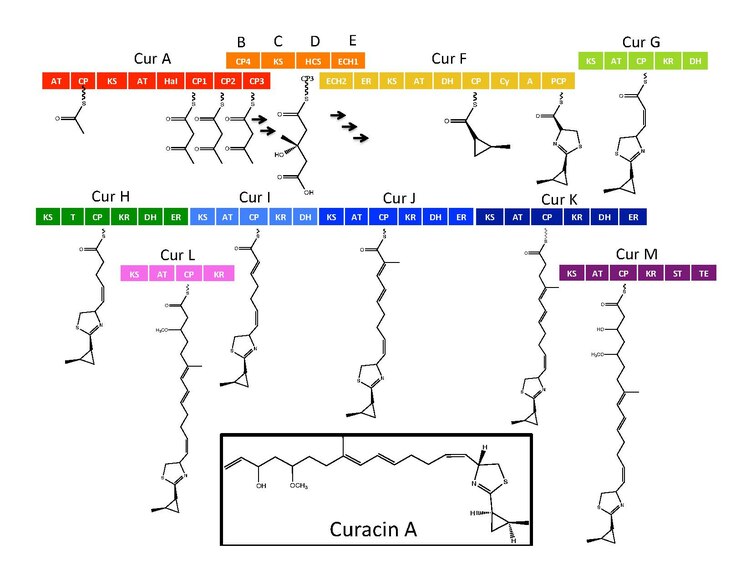
Синтетический ферменты для Curacin A обнаружены в кластере генов с 14 открытые рамки для чтения (ORF) с номенклатурой от CurA до CurN.[1] Анализ пути продемонстрировал наличие одного гибридного модуля NRPS / PKS, расположенного на CurF, одного гибридного модуля. HMG-CoA кассета синтазы, расположенная на CurD, и семь мономодульных модулей PKS.[1] CurA содержит уникальный, связанный с GCN5 N-ацетилтрансфераза (GNAT) загрузочный домен и связанный белок-носитель ацила (ACP).[2] Загрузочный модуль связывает ацетильную группу с ACP, которая затем конденсируется с одним из трех тандемных ACP, присутствующих в соседнем модуле CurA.[1][2][5] Кассета гидроксиметилглутарил-КоА-синтазы (мевалонатный путь) катализирует образование гидроксиметилглутариловой кислоты за счет добавления единицы малонил-КоА к концевому кетиду ацетоацетил-АСР фрагмента ACP1, ACP2 или ACP3.[5] последующие ферменты, включая уникальную гем-независимую галогеназу (HaI), катализируют образование циклопропильного кольца.[1][5][6] Специфичный для цистеина модуль NRPS, расположенный на CurF, следует после образования циклопропильного кольца и из-за активности домена циклизующей конденсации образует тиазольное кольцо, присоединенное к циклопропильному фрагменту в результате предыдущих реакций на этом пути.[1][5][6] Семь автономных модулей PKS следуют за расширением растущего поликетид цепь с S-аденозилметионин (SAM) зависимое метилирование, происходящее в положениях 10 и 13.[1] Редкая стратегия разгрузки, включающая сульфотрансферазу, используется конечным модулем курацин-синтазы. Сульфотрансфераза сульфатирует гидроксильную группу углерода 15, которая активирует молекулу для декарбоксилирования и образования концевого алкена.[7]
Образование циклопропилового кольца
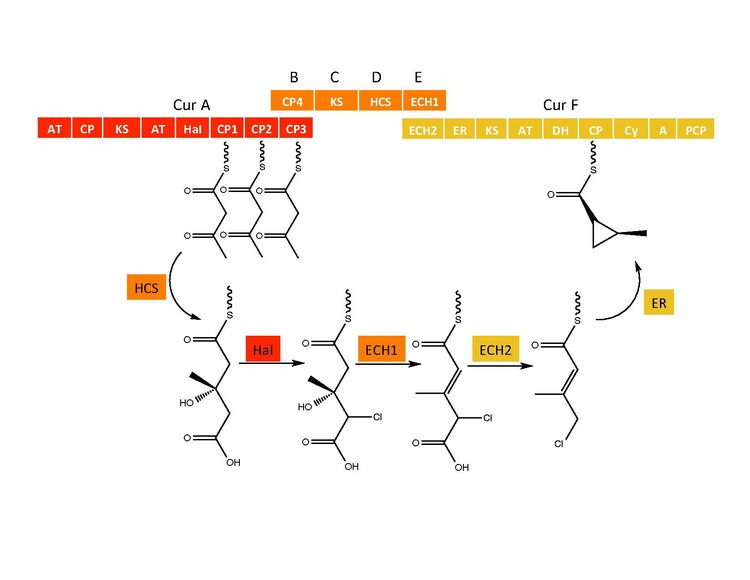
CurB (ACP), CurC (кетосинтаза) и CurD (HMG-CoA редуктаза) ответственны за образование (S) HMG-ACP3.[6] HaI из гена CurA представляет собой уникальную негемовую галогеназу, которая проходит через предполагаемый промежуточный продукт Fe (IV) = O, чтобы присоединить атом хлора к неактивированному атому углерода.[6] После хлорирования ECH1, действуя как дегидратирующий HMG-ACP3, превращается в 3-метилгултаконил-ACP3, и ECH2 выполняет необходимое декарбоксилирование.[6] Наконец, необычная реакция циклизации, катализируемая ER, предположительно проходящая по механизму замещения, образует циклопропановое кольцо.[6] Добавленный атом хлора способствует стадии декарбоксилирования и, вероятно, служит уходящей группой во время образования циклопропанового кольца.[6]
Рекомендации
- ^ а б c d е ж грамм час я Чанг З., Ситачитта Н., Росси СП, Робертс М.А., Флатт П.М., Цзя Дж. И др. (Август 2004 г.). «Биосинтетический путь и кластерный анализ генов курацина А, антитубулинового природного продукта из тропической морской цианобактерии Lyngbya majuscula». Журнал натуральных продуктов. 67 (8): 1356–67. Дои:10.1021 / np0499261. PMID 15332855.
- ^ а б c d Гу Л., Гедерс Т.В., Ван Б., Гервик У.Х., Хоканссон К., Смит Д.Л., Шерман Д.Х. (ноябрь 2007 г.). «GNAT-подобная стратегия инициации поликетидной цепи». Наука. 318 (5852): 970–4. Bibcode:2007Наука ... 318..970Г. Дои:10.1126 / science.1148790. PMID 17991863.
- ^ Verdier-Pinard P, Lai JY, Yoo HD, Yu J, Marquez B, Nagle DG и др. (Январь 1998 г.). «Анализ структуры-активности взаимодействия курацина А, мощного антимитотического агента сайта колхицина, с тубулином и эффектов аналогов на рост клеток рака молочной железы MCF-7». Молекулярная фармакология. 53 (1): 62–76. Дои:10,1124 / моль 53.1.62. PMID 9443933.
- ^ Блохин А.В., Yoo HD, Джеральдс Р.С., Нэгл Д.Г., Гервик У.Х., Хамель Э. (сентябрь 1995 г.). «Характеристика взаимодействия морского цианобактериального природного продукта курацина А с колхициновым сайтом тубулина и первоначальные исследования структуры-активности с аналогами». Молекулярная фармакология. 48 (3): 523–31. PMID 7565634.
- ^ а б c d Gu L, Eisman EB, Dutta S, Franzmann TM, Walter S, Gerwick WH и др. (Март 2011 г.). «Тандемные белки-носители ацила в пути биосинтеза курацина способствуют последовательным мультиферментным реакциям с синергетическим эффектом». Angewandte Chemie. 50 (12): 2795–8. Дои:10.1002 / anie.201005280. ЧВК 3081611. PMID 21387490.
- ^ а б c d е ж грамм Гу Л., Ван Б., Кулкарни А., Гедерс Т. В., Гриндберг Р. В., Гервик Л. и др. (Июнь 2009 г.). «Сборка метаморфических ферментов в диверсификации поликетидов». Природа. 459 (7247): 731–5. Bibcode:2009Натура.459..731Г. Дои:10.1038 / природа07870. ЧВК 2918389. PMID 19494914.
- ^ Маккарти Дж. Г., Эйсман Э. Б., Кулкарни С., Гервик Л., Гервик У. С., Випф П. и др. (Декабрь 2012 г.). «Структурные основы активации функциональных групп сульфотрансферазами в сложных метаболических путях». ACS Химическая биология. 7 (12): 1994–2003. Дои:10.1021 / cb300385m. ЧВК 3528841. PMID 22991895.