Поведение динозавров - Dinosaur behavior
Поведение динозавров палеонтологам трудно изучать, поскольку большая часть палеонтологии зависит исключительно от физических остатков древней жизни. Однако следы окаменелостей и палеопатология могут дать представление о поведении динозавров. Интерпретации поведения динозавров обычно основаны на позе окаменелостей тела и их среда обитания, компьютерное моделирование от их биомеханика, и сравнения с современными животными в аналогичных экологические ниши. Таким образом, нынешнее понимание поведения динозавров основано на предположениях и, вероятно, останется спорным в обозримом будущем. Однако существует общее мнение, что некоторые виды поведения, которые характерны для крокодилов и птиц, ближайших живых родственников динозавров, также были распространены среди динозавров. Стачное поведение было обычным явлением для многих видов динозавров. Динозавры могли собираться в стада для защиты, для мигрирующий целей, или обеспечить защиту их молодых. Есть свидетельства того, что многие типы динозавров, включая различных теропод, зауроподов, анкилозавров, орнитопод и цератопсов, образовывали скопления незрелых особей. Гнезда и яйца были обнаружены у большинства основных групп динозавров, и кажется вероятным, что динозавры общались со своими детенышами так же, как современные птицы и крокодилы. Гребни и оборки некоторых динозавров, таких как маркоцефалы, тероподы и ламбеозаврины, возможно, были слишком хрупкими, чтобы их можно было использовать для активной защиты, и поэтому они, вероятно, использовались для сексуальных или агрессивных демонстраций, хотя мало что известно о спаривании динозавров и территориализм. Похоже, что большинство динозавров полагалось на наземное передвижение. Хорошее понимание того, как динозавры передвигались по земле, является ключом к модели поведения динозавров; наука о биомеханика, в частности, предоставил существенное понимание в этой области. Например, исследования сил, оказываемых мышцами и гравитацией на структуру скелета динозавров, показали, насколько быстро динозавры могут бегать.[1] ли диплодоциды мог создать звуковые удары через хлыст -как хвост,[2] и могут ли зауроподы плавать.[3]
Цератопсовое поведение

Родительская забота подразумевается окаменелыми останками группы Пситтакозавр состоит из одной взрослой особи и 34 молодых особей. В этом случае большое количество молоди может быть связано с совместным гнездованием.[4]
В отложениях окаменелостей преобладали большие количества цератопсидов отдельных видов, что позволяет предположить, что эти животные были, по крайней мере, в некоторой степени социальными.[5] Однако, точная природа ceratopsid социального поведения исторически спорная.[6] В 1997 году Леман утверждал, что скопления многих особей, сохранившихся в костях, возникли как местные «заражения», и сравнил их с аналогичными современными проявлениями у крокодилов и черепах.[6] Другие авторы, такие как Скотт Д. Сэмпсон, интерпретируют эти отложения как остатки больших «социально сложных» стад.[6]
Современные животные, у которых брачные сигналы столь же заметны, как рога и оборки цератопсов, имеют тенденцию образовывать такие большие и сложные ассоциации.[7] Сэмпсон обнаружил в предыдущей работе, что центрозаврин ceratopsids не достигали полностью развитых сигналов спаривания, пока почти полностью не выросли.[8] Он обнаруживает сходство между медленным ростом сигналов спаривания у центрозавринов и расширенными юность животных, чьи социальные структуры представляют собой ранжированные иерархии, основанные на возрастных различиях.[8] В таких группах молодые самцы обычно достигают половой зрелости за несколько лет до начала размножения, когда их брачные сигналы наиболее полно развиты.[9] У женщин же, напротив, нет такого длительного подросткового возраста.[9]
Другие исследователи, поддерживающие идею выпаса цератопсидов, предполагают, что эти ассоциации были сезонными.[10] Согласно этой гипотезе, цератопсиды живут небольшими группами у побережья во время сезона дождей и внутри страны с наступлением сухого сезона.[10] Поддержка идеи о том, что цератопсиды образовывали стада внутри страны, исходит из большего обилия костных пластов во внутренних отложениях, чем на прибрежных. Миграция цератопсидов от берегов могла быть связана с переездом в места их гнездования.[10] Сегодня многие африканские пастушьи животные занимаются этим сезонным выпасом.[10] Стада также обеспечивали некоторый уровень защиты от главных хищников цератопсид, тираннозавриды.[11]
Поведение орнитоподов и парксозавров
Следы также подтвердили родительское поведение среди орнитопод из Остров Скай на северо-западе Шотландия.[12]
Oryctodromeus
Основываясь на современных свидетельствах окаменелостей динозавров, таких как Oryctodromeus, некоторые травоядные виды, похоже, частично роющий (роющий) образ жизни.[13]
Игуанодон
Первое возможное свидетельство стадо поведение было обнаружено в 1878 году 31Игуанодон динозавры, которые тогда считались погибшими вместе в Берниссар, Бельгия, после того, как они упали в глубокую затопленную воронка и утонул.[14]
Hadrosauridae
Следы сотен или даже тысяч травоядных животных указывают на то, что утиные клювы (гадрозавриды), возможно, передвигались большими стадами, как Американский бизон или африканский Газель. Джека Хорнера 1978 открытие Майасаура ("хорошая мать-динозавр") гнездование земля в Монтана продемонстрировали, что родительская забота продолжалась долгое время после рождения среди орнитоподы.[15]
Поведение зауроподоморфа
Следы зауроподов документально подтверждают, что эти животные путешествовали группами, состоящими из нескольких разных видов, по крайней мере, в Оксфордшир, Англия,[16] хотя нет данных о конкретных структурах стада.[17] Есть свидетельства того, что Патагонский титанозавр зауроподы (открытие 1997 г.) гнездились большими группами.[18] Эмбрион динозавра (относящийся к прозауропод Массоспондил ) был обнаружен без зубов, что указывает на необходимость родительской заботы, чтобы прокормить молодого динозавра.[19]
Поведение теропод
Интерпретация динозавров как стадных также распространилась на изображение хищных теропод в виде стаи охотников работая вместе, чтобы сбить крупную добычу.[20][21] Однако такой образ жизни необычен среди современных родственников динозавров (крокодилы и другие рептилии, и птицы – Ястреб Харриса является хорошо задокументированным исключением), а тафономический свидетельства, предполагающие стайную охоту на таких теропод, как Дейноних и Аллозавр также можно интерпретировать как результат фатальных споров между кормящимися животными, как это видно во многих современных диапсид хищники.[22] Раны на голове от укусов говорят о том, что тероподы, по крайней мере, вели активные агрессивные столкновения.[23]
В 2001, Брюс Ротшильд и другие опубликовали исследование, посвященное изучению доказательств стрессовые переломы в теропод динозавры и последствия для их поведения.[24] Поскольку стрессовые переломы вызваны повторной травмой, они с большей вероятностью являются результатом поведения животного, чем переломы, полученные во время единичного травматического события.[25] Распределение стрессовых переломов также имеет поведенческое значение.[26] Стресс-переломы руки с большей вероятностью являются результатом хищнического поведения, поскольку травмы ног могут быть получены во время бега или миграции.[26] Чтобы идентифицировать стрессовые переломы, возникающие в ногах именно из-за хищного поведения, исследователи проверили, имеют ли пальцы ног, на которые приходится наибольшая часть веса животного во время движения, также наибольший процент стрессовых переломов.[27] Поскольку нижний конец третьего плюсневой сначала соприкоснется с землей, пока теропод бежит, он будет испытывать наибольший стресс и должен быть наиболее предрасположен к более частому воздействию факторов стресса.[27] Отсутствие такой предвзятости в изученных окаменелостях указывает на происхождение стрессовых трещин от источника, отличного от бега, например, взаимодействия с добычей.[27] Они предположили, что такие травмы могли произойти в результате того, что теропод пытался удержать сопротивляющуюся добычу ногами.[27] Контакт с сопротивляющейся добычей также является вероятной причиной отрыва сухожилий на передних конечностях Аллозавр и Тиранозавр.[27] Авторы пришли к выводу, что наличие стрессовых переломов свидетельствует об «очень активной» диете, основанной на хищничестве, а не об обязательном питании.[24]
Исследования, основанные на энергетическом моделировании, также подтвердили роль активного хищного поведения у теропод и фактически исключили обязательную уборку мусора как вероятную стратегию.[28] Однако уборка мусора по-прежнему была важным ресурсом для многих теропод, и размер тела играл ключевую роль в его важности. В целом исследователи обнаружили, что мелкие виды, такие как целурозавры, и виды весом более 1000 кг, например взрослые особи Тираннозавр рекс, были бы плохими мусорщиками. В отличие от видов среднего размера, таких как Дилофозавр, было обнаружено, что они способны получать значительную энергию через мусор.[28]
Маджунгазавр
Каннибализм среди некоторых видов динозавров были подтверждены следами зубов, обнаруженными на Мадагаскаре в 2003 году, с участием теропод Маджунгазавр.[29]
Поведение орнитомимидов
Синорнитомим
Один пример незрелых динозавров, образующих социальные группы, взят из Внутренняя Монголия который дал останки более двадцати Синорнитомим, от одного года до семи лет. Это собрание интерпретируется как социальная группа, попавшая в грязь.[30]
Tyrannosauridae
Характер износа зубов намекает на то, что при кормлении тиранозавров могло быть связано сложное покачивание головой.[31] Частично заживший хвост Эдмонтозавр поврежден таким образом, что показывает, что животное было укушено тираннозавром, но выжило.[32]
Oviraptoridae
В Монгольский овирапторид Читипати был обнаружен в курица -подобно задумчивый положение в 1993 году, что может означать, что он был покрыт изоляционным слоем из перьев, который сохранял яйца теплый.[33]
Troodontidae
Недавно обнаруженный Troodont Ископаемое свидетельствует о том, что некоторые динозавры спали, закрыв голову руками.[34] Такое поведение, которое могло помочь сохранить голову в тепле, также характерно для современных птиц.
Dromaeosauridae
Примитивный дромеозавриды Такие как Микрораптор могло быть древесный (лазание по деревьям).[35]
Велоцираптор
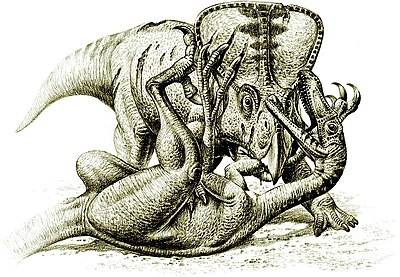
С точки зрения поведения одна из самых ценных окаменелостей динозавров была обнаружена в пустыня Гоби в 1971 году. Велоцираптор нападая на Протоцератопс,[36] предоставление доказательств того, что динозавры действительно нападали друг на друга.[32]
Scansoriopterygidae
Загадочный скансориоптеригиды могло быть древесный (лазание по деревьям).[37]
Смотрите также
Сноски
- ^ Александр Р.М. (2006). «Биомеханика динозавров». Труды Королевского общества B. 273 (1596): 1849–1855. Дои:10.1098 / rspb.2006.3532. ЧВК 1634776. PMID 16822743.
- ^ Горили А, МакМиллен Т (2002). «Форма хрустящей плети». Письма с физическими проверками. 88 (24): 244301. Bibcode:2002ПхРвЛ..88х4301Г. Дои:10.1103 / PhysRevLett.88.244301. PMID 12059302.
- ^ Хендерсон, Д. (2003). «Влияние камней в желудке на плавучесть и равновесие плавающего крокодила: вычислительный анализ». Канадский журнал зоологии. 81 (8): 1346–1357. Дои:10.1139 / z03-122.
- ^ Мэн Цинцзинь; Лю Цзиньюань; Варриккио, Дэвид Дж .; Хуанг, Тимоти; Гао Чуньлин (2004). «Родительская забота у орнитисхий динозавров». Природа. 431 (7005): 145–146. Bibcode:2004Натура 431..145М. Дои:10.1038 / 431145a. PMID 15356619. S2CID 4413450.
- ^ «Абстракция», Сэмпсон (2001); стр.263.
- ^ а б c «Введение», Сэмпсон (2001); стр.264.
- ^ «Социоэкология цератопсов», Sampson (2001); страницы 267-268.
- ^ а б «Замедленный рост сигналов спаривания», Сэмпсон (2001); стр.270.
- ^ а б «Социологические корреляты у современных позвоночных», Сэмпсон (2001); стр.265.
- ^ а б c d «Эксплуатация ресурсов и среда обитания», Сэмпсон (2001); стр.269.
- ^ «Давление хищников», Сэмпсон (2001); стр.272.
- ^ Семейные следы динозавров Следы показывают материнский инстинкт после выхода из гнезда.
- ^ Varricchio DJ; Мартин AJ и Кацура Y (2007). «Первые следы и ископаемые останки трупа роющего норы динозавра». Труды Королевского общества B. 274 (1616): 1361–1368. Дои:10.1098 / rspb.2006.0443. ЧВК 2176205. PMID 17374596.
- ^ Янс Дж; Dejax J; Pons D; Дюпюи С; Taquet P (2005). "Палеонтологические и геодинамические последствия палинологических данных о сборах на фасадах Берниссара (бассен де Монс, Бельгия)". Comptes Rendus Palevol (На французском). 4 (1–2): 135–150. Дои:10.1016 / j.crpv.2004.12.003.
- ^ Хорнер, J.R .; Макела, Роберт (1979). «Гнездо молоди свидетельствует о семейной структуре динозавров». Природа. 282 (5736): 296–298. Bibcode:1979Натура.282..296H. Дои:10.1038 / 282296a0. S2CID 4370793.
- ^ Day, J.J .; Upchurch, P; Норман, ДБ; Гейл, А.С.; Пауэлл, HP (2002). «Следы зауроподов, эволюция и поведение». Наука. 296 (5573): 1659. Дои:10.1126 / science.1070167. PMID 12040187. S2CID 36530770.
- ^ Райт, Джоанна Л. (2005). «Шаги в понимании биологии зауроподов». В Карри Роджерс, Кристина А .; Уилсон, Джеффри А. (ред.). Зауроподы: эволюция и палеобиология. Беркли: Калифорнийский университет Press. стр.252 –284. ISBN 0-520-24623-3.
- ^ Chiappe, Луис М .; Джексон, Фрэнки; Coria, Rodolfo A .; Дингус, Лоуэлл (2005). «Гнездование титанозавров из Аука Махуево и прилегающих территорий». В Карри Роджерс, Кристина А .; Уилсон, Джеффри А. (ред.). Зауроподы: эволюция и палеобиология. Беркли: Калифорнийский университет Press. стр.285 –302. ISBN 0-520-24623-3.
- ^ Рейс Р.Р., Скотт, Д. Сьюз, H-D, Эванс, округ Колумбия и Раат, Массачусетс (2005). «Эмбрионы раннеюрского прозауроподного динозавра и их эволюционное значение». Наука. 309 (5735): 761–764. Bibcode:2005Наука ... 309..761R. Дои:10.1126 / science.1114942. PMID 16051793. S2CID 37548361.CS1 maint: несколько имен: список авторов (связь)
- ^ Лессем, Дон; Glut, Дональд Ф. (1993). "Аллозавр". Энциклопедия динозавров Общества динозавров. Случайный дом. стр.19–20. ISBN 0-679-41770-2.
- ^ Maxwell, W. D .; Остром, Джон (1995). "Тафономия и палеобиологические последствия Тенонтозавр-Дейноних ассоциации ». Журнал палеонтологии позвоночных. 15 (4): 707–712. Дои:10.1080/02724634.1995.10011256.(Абстрактные В архиве 2007-09-27 на Wayback Machine )
- ^ Роуч, Брайан Т .; Бринкман, Дэниел Л. (2007). "Переоценка совместной охоты стаей и общительности в Deinonychus antirrhopus и другие динозавры-тероподы, не являющиеся морскими животными ". Бюллетень музея естественной истории Пибоди. 48 (1): 103–138. Дои:10.3374 / 0079-032X (2007) 48 [103: AROCPH] 2.0.CO; 2.
- ^ Танке, Даррен Х. (1998). "Клевание головы у тероподных динозавров: палеопатологические данные" (PDF). Гайя (15): 167–184. ISSN 0871-5424. Архивировано из оригинал (PDF) 27 февраля 2008 г.
- ^ а б «Аннотация», Ротшильд и др., И др. (2001); стр. 331.
- ^ «Введение» в Rothschild, et al. (2001); страницы 331-332.
- ^ а б «Введение», Ротшильд и др. (2001); стр. 332.
- ^ а б c d е «Обсуждение», Ротшильд и др. (2001); стр. 334.
- ^ а б Кейн, А .; Хили, К; Ruxton, GD; Джексон, А.Л. (2016). "Размер тела как движущая сила мусора у динозавров теропод". Американский натуралист. 187 (6): 706–716. Дои:10.1086/686094. HDL:10023/10617. PMID 27172591. S2CID 3840870.
- ^ Роджерс, Раймонд Р .; Краузе, DW; Карри Роджерс, К. (2003). "Каннибализм у мадагаскарских динозавров Маджунгатолус атопус". Природа. 422 (6931): 515–518. Bibcode:2003Натура.422..515R. Дои:10.1038 / природа01532. PMID 12673249. S2CID 4389583.
- ^ Varricchio, D.J .; Серено, Пол С .; Сицзинь, Чжао; Линь, Тан; Wilson, Jeffery A .; Лион, Габриель Х. (2008). «Стадо в грязевой ловушке - свидетельство самобытной социальной жизни динозавров». Acta Palaeontologica Polonica. 53 (4): 567–578. Дои:10.4202 / приложение.2008.0402.
- ^ «Введение», Аблер (2001); стр.84.
- ^ а б Карпентер, К. (1998). «Свидетельства хищного поведения тероподных динозавров». Гайя. 15: 135–144. Архивировано из оригинал на 2007-11-17. Получено 2007-12-05.
- ^ Гнездование овирапторов Овираптор гнезда или Протоцератопс ?
- ^ Сюй, X .; Норелл, М.А. (2004). «Новый динозавр троодонтид из Китая со спящей позой, напоминающей птицу». Природа. 431 (7010): 838–841. Bibcode:2004Натура.431..838X. Дои:10.1038 / природа02898. PMID 15483610. S2CID 4362745.
- ^ Chatterjee, S .; Темплин, Р. Дж. (2007). «Форма крыла биплана в плане и летные характеристики пернатого динозавра. Microraptor gui" (PDF). Труды Национальной академии наук. 104 (5): 1576–1580. Bibcode:2007PNAS..104.1576C. Дои:10.1073 / pnas.0609975104. ЧВК 1780066. PMID 17242354.
- ^ "Боевые динозавры". Американский музей естественной истории. Архивировано из оригинал 9 марта 2012 г.. Получено 2007-12-05.
- ^ Zhang, F .; Чжоу, З .; Сюй, X .; Ван, X. (2002). «Молодой теропод целурозавра из Китая указывает на древесные привычки». Naturwissenschaften. 89 (9): 394–398. Bibcode:2002NW ..... 89..394Z. Дои:10.1007 / s00114-002-0353-8. PMID 12435090. S2CID 556221.
Рекомендации
- Abler, W.L. 2001. Модель зубцов тираннозавра в виде пропилов и сверлений. п. 84-89. В: Мезозойская жизнь позвоночных. Под ред. Танке, Д. Х., Карпентер, К., Скрепник, М. В. Издательство Индианского университета.
- Ротшильд, Б., Танке, Д.Х., и Форд, Т.Л., 2001, Стрессовые переломы теропод и разрывы сухожилий как ключ к активности: В: Мезозойская жизнь позвоночных, под редакцией Танке, Д.Х., и Карпентер, К., Indiana University Press, п. 331-336.
- Сэмпсон, С. Д., 2001, Размышления о социоэкологии динозавров цератопсидов (Orinthischia: Neoceratopsia): In: Mesozoic Vertebrate Life, под редакцией Танке Д. Х. и Карпентера К., Indiana University Press, стр. 263–276.