Ретрон - Retron - Wikipedia
| Ретрон msr РНК | |
|---|---|
 Предсказанный вторичная структура и сохранение последовательности MSR | |
| Идентификаторы | |
| Символ | MSR |
| Рфам | RF00170 |
| Прочие данные | |
| РНК тип | Ген |
| Домен (ы) | Бактерии |
| ТАК | ТАК: 0000233 |
| PDB структуры | PDBe |
А ретрон это отдельный ДНК последовательность найдена в геном из многих бактерии виды, которые кодируют обратная транскриптаза и уникальный одноцепочечный гибрид ДНК / РНК, названный многокопия одноцепочечной ДНК (мсДНК). Ретрон msr РНК это некодирующая РНК продуцируется ретронными элементами и является непосредственным предшественником синтеза мсДНК. РНК msr ретрона складывается в характерную вторичную структуру, содержащую консервативную гуанозин остаток на конце петли стебля. Синтез ДНК кодируемой ретроном обратной транскриптазой (RT) приводит к образованию химеры ДНК / РНК, которая состоит из небольшой одноцепочечной ДНК, связанной с небольшой одноцепочечной РНК. Нить РНК присоединяется к 5′ конец цепи ДНК через 2'-5'-фосфодиэфирную связь, которая начинается в положении 2 'консервативного внутреннего остатка гуанозина.
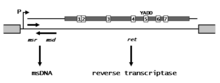
Элементы ретрона имеют длину около 2 кбайт. Они содержат один оперон контроль синтеза транскрипта РНК, несущего три локуса, MSR, MSD, и Ret, которые участвуют в синтезе мсДНК. Часть ДНК мсДНК кодируется MSD ген, часть РНК кодируется MSR ген, а продукт Ret ген представляет собой обратную транскриптазу, аналогичную RT, продуцируемым ретровирусы и другие виды ретроэлементов.[1] Как и другие обратные транскриптазы, ретрон RT содержит семь областей консервативных аминокислот (обозначенных 1-7 на рисунке), включая высококонсервативную последовательность tyr-ala-asp-asp (YADD), связанную с каталитическим ядром. В Ret генный продукт отвечает за обработку msd / msr часть транскрипта РНК в мсДНК.
В течение многих лет после их открытия у вирусов животных считалось, что обратная транскриптаза отсутствует у прокариот. Однако в настоящее время элементы RT-кодирования, т.е. ретроэлементы, были обнаружены в большом количестве различных бактерий. Ретроны были первым семейством ретроэлементов, обнаруженных у бактерий; два других семейства известных бактериальных ретроэлементов: интроны группы II и создающие разнообразие ретроэлементы (DGR).[2] Интроны группы II представляют собой наиболее хорошо охарактеризованный бактериальный ретроэлемент и единственный тип, который, как известно, проявляет автономную подвижность; они состоят из RT, закодированной в каталитической самосплайсинговой структуре РНК. Подвижность интронов группы II опосредуется рибонуклеопротеин состоящий из интронного лариата, связанного с двумя кодируемыми интронами белками. Второе семейство бактериальных ретроэлементов, DGR, не является мобильным, но функционирует, чтобы разнообразить последовательности ДНК.[3] Например, DGR опосредуют переключение между патогенной и свободноживущей фазами Bordetella.[4]
Поскольку ретроны не подвижны, их появление у различных видов бактерий не означает "эгоистичная ДНК "феномен. Скорее, ретроны должны давать некоторое селективное преимущество организму-хозяину. Какое это преимущество может быть неизвестно. За исключением продуцирования мсДНК, с ними не был связан четкий фенотип. Несмотря на обширные исследования, очень мало известно о функции мсДНК, подвижность ретронных элементов или их влияние на хозяйскую клетку.[5][6] Однако недавние данные свидетельствуют о том, что они играют роль в предотвращении распространения бактериофагов в популяции.[7]
Ретроны развиваются в инструменты редактирования генома.[8]
использованная литература
- ^ Лэмпсон BC, Иноуе М., Иноуе С. (2005). «Ретроны, мсДНК и бактериальный геном» (PDF). Cytogenet Genome Res. 110 (1–4): 491–499. Дои:10.1159/000084982. PMID 16093702.
- ^ Саймон Д.М., Циммерли С. (2008). «Разнообразие не охарактеризованных обратных транскриптаз в бактериях». Нуклеиновые кислоты Res. 36 (22): 7219–7229. CiteSeerX 10.1.1.358.8390. Дои:10.1093 / nar / gkn867. ЧВК 2602772. PMID 19004871.
- ^ Медхекар Б., Милле Дж. Ф. (2007). "Разнообразные ретроэлементы". Текущее мнение в микробиологии. 10 (4): 388–395. Дои:10.1016 / j.mib.2007.06.004. ЧВК 2703298. PMID 17703991.
- ^ Лю М., Джинджери М., Доулатов С.Р., Лю Ю., Ходес А., Бейкер С., Дэвис П., Симмондс М., Черчер С., Мунгалл К., Квейл М.А., Престон А., Харвилл Е.Т., Маскелл Д.Д., Эйзерлинг Ф.А., Пархилл Дж., Миллер Д. (2004). «Геномно-генетический анализ Bordetella Бактериофаги, кодирующие кассеты с переключением тропизма, опосредованным обратной транскриптазой ». J. Bacteriol. 186 (5): 1503–1517. Дои:10.1128 / JB.186.5.1503-1517.2004. ЧВК 344406. PMID 14973019.
- ^ Ахмед, AM; Симамото Т. (2003). "msDNA-St85, многокопийная одноцепочечная ДНК, выделенная из Salmonella enterica серовар Typhimurium LT2 с геномным анализом его ретрона ». FEMS Microbiol Lett. 224 (2): 291–297. Дои:10.1016 / S0378-1097 (03) 00450-6. PMID 12892895.
- ^ Лэмпсон, Британская Колумбия; Сюй С; Рис SA; Иноуе С (2002). "Частичная копия мсДНК из нового ретронного элемента, вероятно, представляет собой ретротранспозицию ДНК, обнаруженную в миксобактериях. Nannocystis exedens". Ген. 299 (1–2): 251–261. Дои:10.1016 / S0378-1119 (02) 00977-0. PMID 12459273.
- ^ Миллман А., Бернхейм А., Стокар-Авихаил А., Федоренко Т., Войчек М., Ливитт А., Оппенгеймер-Шаанан Ю., Сорек Р. (2020). «Бактериальные ретроны в защите от фагов». Ячейка. 183 (6): 1551–1561. Дои:10.1016 / j.cell.2020.09.065.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ Саймон А.Дж., Эллингтон А.Д., Финкельштейн И.Дж. (2019). «Ретроны и их применение в геномной инженерии». Исследования нуклеиновых кислот. 47 (21): 11007–11019. Дои:10.1093 / нар / gkz865.CS1 maint: использует параметр авторов (ссылка на сайт)
внешние ссылки
- Страница для РНК Retron msr в Рфам
- Загадочная молекула бактерий - охранник, on: EurekAlert !, 5 ноября 2020 г. Источник: WEIZMANN INSTITUTE OF SCIENCE