Сотрудничество - Cooperativity
Эта статья нужны дополнительные цитаты для проверка. (Декабрь 2009 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |
Сотрудничество - это явление, отображаемое системами, включающими идентичные или почти идентичные элементы, которые действуют зависимо друг от друга, по сравнению с гипотетической стандартной невзаимодействующей системой, в которой отдельные элементы действуют независимо.[1] Одним из проявлений этого является ферменты или же рецепторы которые имеют несколько участок связывания где аффинность сайтов связывания лиганда равна по-видимому повысился, позитивное сотрудничество, или уменьшилось, отрицательное сотрудничествопри связывании лиганда с сайтом связывания.[2] Например, когда атом кислорода связывается с одним из четырех сайтов связывания гемоглобина, сродство к кислороду трех оставшихся доступных сайтов связывания увеличивается; то есть кислород с большей вероятностью связывается с гемоглобином, связанным с одним кислородом, чем с несвязанным гемоглобином. Это называется совместная привязка.[3]
Мы также видим кооперативность в молекулах с большой цепью, состоящих из множества идентичных (или почти идентичных) субъединиц (таких как ДНК, белки, и фосфолипиды ), когда такие молекулы претерпевают фазовые переходы, такие как плавление, разворачивание или раскручивание. Это называется кооперативностью субъединиц. Однако определение кооперативности, основанное на очевидном увеличении или уменьшении сродства к последовательным этапам связывания лиганда, является проблематичным, поскольку понятие «энергия» всегда должно определяться относительно стандартного состояния. Когда мы говорим, что аффинность увеличивается при связывании одного лиганда, эмпирически неясно, что мы имеем в виду, поскольку кривая некооперативного связывания необходима для точного определения энергии связывания и, следовательно, также аффинности. Гораздо более общее и полезное определение позитивной кооперативности: процесс, включающий несколько идентичных дополнительных шагов, в которых промежуточные состояния статистически недостаточно представлен относительно гипотетической стандартной системы (нулевая гипотеза), где шаги выполняются независимо друг от друга.
Точно так же определение отрицательной кооперативности было бы процессом, включающим несколько идентичных дополнительных шагов, в которых промежуточные состояния чрезмерно представлен относительно гипотетического стандартного состояния, в котором отдельные шаги происходят независимо.[4] Эти последние определения положительной и отрицательной кооперативности легко охватывают все процессы, которые мы называем «кооперативными», включая конформационные переходы в больших молекулах (таких как белки) и даже психологические явления большого числа людей (которые могут действовать независимо друг от друга или в кооперативная мода).
Кооперативная привязка
Когда субстрат связывается с одной субъединицей фермента, остальные субъединицы стимулируются и становятся активными. Лиганды может иметь положительную кооперативность, отрицательную кооперативность или отказ от сотрудничества.[2][1]
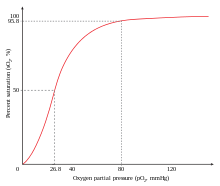
Примером позитивного сотрудничества является связывание кислород к гемоглобин. Один кислород молекула может связываться с двухвалентное железо молекулы гема в каждой из четырех цепей гемоглобин молекула. Дезоксигемоглобин имеет относительно низкое сродство к кислород, но когда одна молекула связывается с одним гемом, кислород сродство увеличивается, позволяя второй молекуле связываться более легко, а третьей и четвертой - еще легче. В кислород сродство 3-оксигемоглобина примерно в 300 раз больше, чем у дезоксигемоглобина. Такое поведение приводит к кривой сродства гемоглобин быть сигмовидный, скорее, чем гиперболический как с мономерным миоглобин. Таким же образом способность к гемоглобин терять кислород увеличивается по мере того, как связано меньше молекул кислорода.[3] Смотрите также Кривая диссоциации кислород-гемоглобин.
Отрицательная кооперативность означает, что верно обратное; в качестве лиганды привязать к белок, то белок Сродство к лиганду будет уменьшаться, то есть становится менее вероятным связывание лиганда с белком. Примером этого является взаимосвязь между глицеральдегид-3-фосфат и фермент глицеральдегид-3-фосфатдегидрогеназа.
Гомотропный Кооперативность относится к тому факту, что молекула, вызывающая кооперативность, будет ею затронута. Гетеротропный Кооперативность - это когда стороннее вещество вызывает изменение аффинности. Гомотропная или гетеротропная кооперативность может быть как положительной, так и отрицательной, в зависимости от того, поддерживает она или препятствует дальнейшему связыванию молекул лиганда с ферментами.[5]
Подразделение кооперативности
Кооперативность - это не только явление связывания лиганда, но также применяется в любое время, когда энергетические взаимодействия облегчают или затрудняют что-то, что происходит с участием нескольких единиц, а не с отдельными единицами. (То есть проще или сложнее по сравнению с тем, что ожидается при учете только добавления нескольких единиц). Например, раскрутка ДНК предполагает взаимодействие: части ДНК должны раскручиваться, чтобы ДНК могла выполнять репликация, транскрипция и рекомбинация. Положительное взаимодействие между соседними ДНК нуклеотиды позволяет размотать целую группу соседних нуклеотидов легче, чем разматывать такое же количество нуклеотидов, распределенных по цепи ДНК. В размер кооперативного подразделения - это количество смежных баз, которые имеют тенденцию раскручиваться как единое целое из-за эффектов положительного взаимодействия. Это явление применимо и к другим типам цепных молекул, таким как сворачивание и разворачивание белков и «плавление» фосфолипид цепи, которые составляют мембраны клеток. Кооперативность субъединиц измеряется по относительной шкале, известной как константа Хилла.
Уравнение Хилла
Простая и широко используемая модель молекулярных взаимодействий - это модель Уравнение Хилла, который обеспечивает способ количественной оценки кооперативного связывания путем описания доли сайтов связывания насыщенного лиганда как функции от концентрации лиганда.
Коэффициент Хилла
Коэффициент Хилла является мерой сверхчувствительность (т.е. насколько крута кривая отклика).
С эксплуатационной точки зрения коэффициент Хилла можно рассчитать как:
- .

куда и - входные значения, необходимые для получения 10% и 90% максимального отклика соответственно.


Коэффициент отклика
Показатели глобальной чувствительности, такие как коэффициент Хилла, не характеризуют локальное поведение s-образных кривых. Вместо этого эти особенности хорошо отражаются с помощью меры коэффициента отклика. [6] определяется как:

Связь между коэффициентом Хилла и коэффициентом отклика
Altszyler et al. (2017) показали, что эти меры сверхчувствительности могут быть связаны следующим уравнением:[7]

куда обозначает среднее значение переменной x в диапазоне [a, b].

Сверхчувствительность в функциональном составе
Рассмотрим два связанных сверхчувствительных модуля, не принимая во внимание эффекты связывания молекулярных компонентов между слоями. В этом случае выражение для кривой доза-ответ системы, F, является результатом математической композиции функций, , которые описывают отношения ввода / вывода изолированных модулей :



Brown et al. (1997) [8][7] показали, что локальная сверхчувствительность различных слоев мультипликативно сочетается:
- .

В связи с этим результатом Ferrell et al. (1997) [9] показал для модулей типа Хилла, что общая глобальная сверхчувствительность каскада должна быть меньше или равна произведению оценок глобальной сверхчувствительности каждого слоя каскада,[7]
- ,

куда и - коэффициент Хилла модулей 1 и 2 соответственно.


Altszyler et al. (2017) [7] показали, что глобальную сверхчувствительность каскада можно рассчитать аналитически:

куда и ограничил рабочий диапазон входных данных Хилла составной системы, то есть входные значения для i-слоя так, чтобы последний слой (соответствующий в данном случае) достиг 10% и 90% от максимального уровня выпуска. Из этого уравнения следовало, что коэффициент Хилла системы п может быть записано как произведение двух факторов, и , который характеризует среднюю локальную чувствительность по соответствующей входной области для каждого слоя: , с в этом случае.





![{ displaystyle [X10_ {i}, X90_ {i}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/2a6a9f701cc2d5bd5754f6206af5ce8e2b644e8f)
Для более общего случая каскада N модулей коэффициент Хилла можно выразить как:
- ,

Супрамультипликативность
Несколько авторов сообщили о существовании супрамультипликативного поведения в сигнальных каскадах. [10][11](т.е. сверхчувствительность комбинации слоев выше, чем результат индивидуальных сверхчувствительности), но во многих случаях окончательное происхождение супрамультипликативности оставалось неуловимым. Altszyler et al. (2017)[7] Структура естественно предложила общий сценарий, в котором может иметь место супрамультипликативное поведение. Это могло произойти, когда для данного модуля соответствующий входной рабочий диапазон Хилла был расположен во входной области с локальной сверхчувствительностью выше, чем общая сверхчувствительность соответствующей кривой доза-реакция.
Рекомендации
- ^ а б Шринивасан, Бхарат; Фоухар, Фархад; Шукла, Арпит; Сампанги, Четана; Кулькарни, Соня; Абашидзе, Мариам; Ситхараман, Джаяраман; Лью, Скотт; Мао, Лэй; Актон, Томас Б .; Сяо, Жун (март 2014 г.). «Аллостерическая регуляция и активация субстрата в цитозольной нуклеотидазе II из Legionella pneumophila». Журнал FEBS. 281 (6): 1613–1628. Дои:10.1111 / фев.12727. ЧВК 3982195. PMID 24456211.
- ^ а б Шринивасан, Бхарат (08.10.2020). «Явное лечение не Михаэлиса-Ментена и атипичной кинетики при раннем открытии лекарств». Дои:10.20944 / препринты202010.0179.v1. Цитировать журнал требует
| журнал =(помощь) - ^ а б Уитфорд Д. (2005). Белки: структура и функции. Джон Вили и сыновья. С. 66–74.
- ^ Абелиович Х (июль 2005 г.). «Эмпирический принцип экстремума для коэффициента Хилла во взаимодействиях лиганд-белок, показывающий отрицательную кооперативность». Биофизический журнал. 89 (1): 76–9. Bibcode:2005BpJ .... 89 ... 76A. Дои:10.1529 / biophysj.105.060194. ЧВК 1366580. PMID 15834004.
- ^ Хуссейн Р., Кумари И., Шарма С., Ахмед М., Хан Т.А., Ахтер Ю. (декабрь 2017 г.). «Каталитическое разнообразие и гомотропная аллостерия двух монооксигеназных белков цитохрома P450 из Trichoderma brevicompactum». Журнал биологической неорганической химии. 22 (8): 1197–1209. Дои:10.1007 / s00775-017-1496-6. PMID 29018974. S2CID 25685603.
- ^ Холоденко Б.Н., Хук Дж.Б., Вестерхофф Х.В., Браун Г.К. (сентябрь 1997 г.). «Количественная оценка передачи информации через пути передачи клеточного сигнала». Письма FEBS. 414 (2): 430–4. Дои:10.1016 / S0014-5793 (97) 01018-1. PMID 9315734. S2CID 19466336.
- ^ а б c d е Альцзилер Э., Вентура А.С., Колман-Лернер А., Черноморец А. (29 июня 2017 г.). «Пересмотр сверхчувствительности в сигнальных каскадах: увязка оценок локальной и глобальной сверхчувствительности». PLOS ONE. 12 (6): e0180083. arXiv:1608.08007. Bibcode:2017PLoSO..1280083A. Дои:10.1371 / journal.pone.0180083. ЧВК 5491127. PMID 28662096.
 Эта статья содержит цитаты из этого источника, который доступен под Creative Commons Attribution 4.0 International (CC BY 4.0) лицензия.
Эта статья содержит цитаты из этого источника, который доступен под Creative Commons Attribution 4.0 International (CC BY 4.0) лицензия. - ^ Браун Г.К., Хук Дж. Б., Холоденко Б. Н. (август 1997 г.). «Почему каскады протеинкиназ имеют более одного уровня?». Тенденции в биохимических науках. 22 (8): 288. Дои:10.1016 / s0968-0004 (97) 82216-5. PMID 9270298.
- ^ Феррелл Дж. Э. (август 1997 г.). «Как ответы становятся более похожими на переключатели при движении вниз по каскаду протеинкиназ». Тенденции в биохимических науках. 22 (8): 288–9. Дои:10.1016 / s0968-0004 (97) 82217-7. PMID 9270299.
- ^ Альцзилер Э., Вентура А., Колман-Лернер А., Черноморец А. (октябрь 2014 г.). «Влияние ограничений восходящего и нисходящего потоков на сверхчувствительность сигнального модуля». Физическая биология. 11 (6): 066003. Bibcode:2014PhBio..11f6003A. Дои:10.1088/1478-3975/11/6/066003. ЧВК 4233326. PMID 25313165.
- ^ Рац Э., Слепченко Б.М. (июль 2008 г.). «Об усилении чувствительности во внутриклеточных сигнальных каскадах». Физическая биология. 5 (3): 036004. Bibcode:2008ФБио ... 5c6004R. Дои:10.1088/1478-3975/5/3/036004. ЧВК 2675913. PMID 18663279.