Гемоглобин - Hemoglobin
| гемоглобин | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (гетеротетрамер, (αβ)2) | |||||||||||||
 Структура гемоглобина человека. α и β субъединицы выделены красным и синим цветом соответственно, а железосодержащие гем группы в зеленом цвете. Из PDB: 1GZX Протеопедия Гемоглобин | |||||||||||||
| Тип белка | металлопротеин, глобулин | ||||||||||||
| Функция | кислород -транспорт | ||||||||||||
| Кофактор (ы) | гем (4) | ||||||||||||
| |||||||||||||
Гемоглобин (Американский английский) или гемоглобин (Британский английский) (греч. Αἷμα (haîma, «кровь») + -in) + -o- + globulin (от латинского globus («шар, сфера») + -in) (/ˈчасямəˌɡлoʊбɪп,ˈчасɛ-,-мoʊ-/[1][2][3]), сокращенно Hb или же Hgb, это утюг -содержащий кислород -транспорт металлопротеин в красные кровяные тельца (эритроциты) почти всех позвоночные[4] (за исключением семейства рыб Каннихтииды[5]), а также ткани некоторых беспозвоночные. Гемоглобин в кровь переносит кислород из легкие или же жабры остальной части тела (то есть тканям). Там он выпускает кислород, чтобы позволить аэробного дыхания для обеспечения энергией функций организма в процессе, называемом метаболизм. У здорового человека на каждые 100 мл крови приходится от 12 до 20 граммов гемоглобина.
В млекопитающие белок составляет около 96% сухого содержания красных кровяных телец (по весу) и около 35% от общего содержания (включая воду).[6] Гемоглобин обладает способностью связывать кислород 1,34 мл O2 за грамм,[7] что увеличивает общую кислородная емкость крови в семьдесят раз по сравнению с растворенным в крови кислородом. Молекула гемоглобина млекопитающих может связывать (переносить) до четырех молекул кислорода.[8]
Гемоглобин участвует в транспортировке других газов: он переносит часть дыхательных путей организма. углекислый газ (около 20–25% от общего[9]) в качестве карбаминогемоглобин, в котором CO2 привязан к гемовый белок. Молекула также несет важную регуляторную молекулу оксид азота связаны с белком глобина тиол группа, высвобождая его одновременно с кислородом.[10]
Гемоглобин также находится вне красных кровяных телец и их линий-предшественников. Другие клетки, содержащие гемоглобин, включают A9 дофаминергические нейроны в черная субстанция, макрофаги, альвеолярные клетки, легкие, пигментный эпителий сетчатки, гепатоциты, мезангиальные клетки в почках, клетках эндометрия, цервикальных клетках и эпителиальных клетках влагалища.[11] В этих тканях гемоглобин не несет кислородную функцию в качестве антиоксидант и регулятор метаболизм железа.[12] Избыток глюкозы в крови может присоединяться к гемоглобину и повышать уровень гемоглобина A1c.[13]
Гемоглобин и гемоглобиноподобные молекулы также встречаются у многих беспозвоночных, грибов и растений.[14] У этих организмов гемоглобины могут переносить кислород или они могут действовать, чтобы транспортировать и регулировать другие небольшие молекулы и ионы, такие как диоксид углерода, оксид азота, сероводород и сульфид. Вариант молекулы, названный леггемоглобин, используется для удаления кислорода из анаэробный системы, такие как узелки фиксации азота бобовые растения, чтобы кислород не отравил (не отключил) систему.
Гемоглобинемия заболевание, при котором наблюдается избыток гемоглобина в плазма крови. Это эффект внутрисосудистый гемолиз, в котором гемоглобин отделяется от красные кровяные тельца, форма анемия.
История исследований

В 1825 г. Дж. Ф. Энгельхарт обнаружил, что соотношение железа к белку идентично в гемоглобинах нескольких видов.[16][17] По известной атомной массе железа он рассчитал молекулярную массу гемоглобина до п × 16000 (п = количество атомов железа на гемоглобин, теперь известное как 4), первое определение молекулярной массы белка. Этот «поспешный вывод» вызвал много насмешек со стороны ученых, которые не могли поверить, что какая-либо молекула может быть настолько большой. Гилберт Смитсон Адэр подтвердил результаты Энгельхарта в 1925 году, измерив осмотическое давление растворов гемоглобина.[18]
Свойство гемоглобина переносить кислород было открыто Хюнефельдом в 1840 году.[19] В 1851 году немецкий физиолог Отто Функе опубликовал серию статей, в которых описал выращивание кристаллов гемоглобина путем последовательного разбавления красных кровяных телец растворителем, таким как чистая вода, спирт или эфир, с последующим медленным испарением растворителя из полученного раствора белка.[20][21] Обратимая оксигенация гемоглобина была описана несколькими годами позже Феликс Хоппе-Зейлер.[22]
В 1959 г. Макс Перуц определил молекулярную структуру гемоглобина Рентгеновская кристаллография.[23][24] Результатом этой работы стал его обмен с Джон Кендрю 1962 г. Нобелевская премия по химии за исследования структуры глобулярных белков.
Роль гемоглобина в крови была выяснена французскими исследователями. физиолог Клод Бернард.Название гемоглобин происходит от слов гем и глобин, отражая тот факт, что каждый подразделение гемоглобина - это глобулярный белок со встроенным гем группа. Каждая группа гема содержит один атом железа, который может связывать одну молекулу кислорода посредством дипольных сил, индуцированных [ионами]. Наиболее распространенный тип гемоглобина у млекопитающих состоит из четырех таких субъединиц.
Генетика
Гемоглобин состоит из белковые субъединицы (молекулы «глобина»), а эти белки, в свою очередь, представляют собой свернутые цепи из большого количества различных аминокислот, называемых полипептиды. Аминокислотная последовательность любого полипептида, созданного клеткой, в свою очередь, определяется участками ДНК, называемыми генами. Во всех белках именно аминокислотная последовательность определяет химические свойства и функции белка.
Существует более одного гена гемоглобина: у людей гемоглобин А (основная форма присутствующего гемоглобина) кодируется генами, HBA1, HBA2, и HBB.[25] Аминокислотные последовательности белков глобина в гемоглобинах обычно различаются между видами. Эти различия растут с эволюционным расстоянием между видами. Например, наиболее распространенные последовательности гемоглобина у людей, бонобо и шимпанзе полностью идентичны, даже без единой аминокислотной разницы в альфа- или бета-белковых цепях глобина.[26][27][28] В то время как гемоглобин человека и гориллы различается одной аминокислотой как в альфа-, так и в бета-цепях, эти различия становятся больше между менее близкими видами.
Варианты гемоглобина существуют даже внутри одного вида, хотя одна последовательность обычно «наиболее распространена» у каждого вида. Мутации в гены для гемоглобина белок в виде результата варианты гемоглобина.[29][30] Многие из этих мутантных форм гемоглобина не вызывают болезней. Однако некоторые из этих мутантных форм гемоглобина вызывают группу наследственные болезни назвал гемоглобинопатии. Самая известная гемоглобинопатия - это серповидноклеточная анемия, которая была первой болезнью человека, механизм понималось на молекулярном уровне. (В основном) отдельный набор болезней, называемых талассемии включает недостаточное производство нормальных, а иногда и аномальных гемоглобинов из-за проблем и мутаций в глобине генная регуляция. Все эти болезни производят анемия.[31]

Вариации аминокислотных последовательностей гемоглобина, как и других белков, могут быть адаптивными. Например, было обнаружено, что гемоглобин по-разному приспосабливается к большой высоте. Организмы, живущие на большой высоте, испытывают более низкое парциальное давление кислорода по сравнению с живущими на уровне моря. Это представляет проблему для организмов, населяющих такие среды, потому что гемоглобин, который обычно связывает кислород при высоких парциальных давлениях кислорода, должен быть способен связывать кислород, когда он присутствует при более низком давлении. К такому вызову приспособились разные организмы. Например, недавние исследования предложили генетические варианты у мышей-оленей, которые помогают объяснить, как мыши-олени, живущие в горах, могут выжить в разреженном воздухе, который сопровождает большие высоты. Исследователь из Университета Небраски в Линкольне обнаружил мутации в четырех разных генах, которые могут объяснить различия между оленями, живущими в низинных прериях, и в горах. После изучения диких мышей, пойманных как в высокогорьях, так и в низинах, было обнаружено, что: гены этих двух пород «практически идентичны - за исключением тех, которые определяют способность их гемоглобина переносить кислород». «Генетические различия позволяют высокогорным мышам более эффективно использовать кислород», поскольку на больших высотах, например, в горах, доступно меньше кислорода.[32] Мамонт гемоглобин показал мутации, которые позволили доставить кислород при более низких температурах, что позволило мамонтам мигрировать в более высокие широты во время Плейстоцен.[33] Это также было обнаружено у колибри, населяющих Анды. Колибри уже расходуют много энергии и, таким образом, имеют высокие потребности в кислороде, и тем не менее было обнаружено, что андские колибри прекрасно себя чувствуют на больших высотах. Несинонимичные мутации в гене гемоглобина у нескольких видов, живущих на больших высотах (Oreotrochilus, A. castelnaudii, C. violifer, P. gigas, и A. viridicuada) вызвали меньшее сродство белка к гексафосфат инозита (IHP), молекула, обнаруженная у птиц, которая играет такую же роль, как 2,3-BPG у людей; это приводит к способности связывать кислород при более низких парциальных давлениях.[34]
Уникальные птицы кровообращение в легких также способствуют эффективному использованию кислорода при низких парциальных давлениях O2. Эти две адаптации усиливают друг друга и объясняют замечательные высотные способности птиц.
Адаптация к гемоглобину распространяется и на людей. Среди тибетских женщин с генотипами с высокой степенью насыщения кислородом, проживающих на высоте 4000 м, выше выживаемость потомства.[35] Естественный отбор, по-видимому, является основной силой, воздействующей на этот ген, потому что уровень смертности потомства значительно ниже для женщин с более высоким гемоглобин-кислородным сродством по сравнению с уровнем смертности потомства от женщин с низким сродством гемоглобин-кислород. Хотя точный генотип и механизм, с помощью которого это происходит, еще не ясны, отбор воздействует на способность этих женщин связывать кислород при низких парциальных давлениях, что в целом позволяет им лучше поддерживать важные метаболические процессы.
Синтез
Гемоглобин (Hb) синтезируется в сложной серии этапов. Гемовая часть синтезируется в несколько этапов в митохондрии и цитозоль незрелых эритроцитов, в то время как глобин белковые части синтезируются рибосомы в цитозоле.[36] Производство Hb продолжается в клетке на протяжении всего ее развития от проэритробласт к ретикулоцит в Костный мозг. На данный момент ядро теряется в эритроцитах млекопитающих, но не в птицы и многие другие виды. Даже после потери ядра у млекопитающих остаточные рибосомная РНК обеспечивает дальнейший синтез Hb до тех пор, пока ретикулоцит не потеряет свою РНК вскоре после попадания в сосудистая сеть (эта синтетическая гемоглобин-синтетическая РНК фактически придает ретикулоциту его сетчатый вид и название).[37]
Структура гема

Гемоглобин имеет четвертичная структура характерен для многих многосубъединичных глобулярных белков.[38] Большинство аминокислот в гемоглобине образуют альфа-спирали, и эти спирали соединены короткими неспиральными сегментами. Водородные связи стабилизируют спиральные участки внутри этого белка, вызывая притяжение внутри молекулы, что затем заставляет каждую полипептидную цепь складываться в определенную форму.[39] Четвертичная структура гемоглобина состоит из четырех его субъединиц, расположенных примерно в тетраэдрической форме.[38]
У большинства позвоночных гемоглобин молекула это сборка из четырех глобулярный белок субъединицы. Каждая субъединица состоит из белок цепь, прочно связанная с небелковым протез гем группа. Каждая белковая цепочка состоит из набора альфа-спираль структурные сегменты, соединенные вместе в глобиновая складка расположение. Такое название дано потому, что это расположение является тем же мотивом сворачивания, который используется в других белках гема / глобина, таких как миоглобин.[40][41] Этот шаблон складывания содержит карман, который прочно связывает гемовую группу.
Группа гема состоит из железа (Fe) ион проводился в гетероциклический кольцо, известное как порфирин. Это порфириновое кольцо состоит из четырех пиррол молекулы, циклически связанные вместе ( метин мосты) с связанным в центре ионом железа.[42] Ион железа, который является местом связывания кислорода, координируется с четырьмя азот атомы в центре кольца, которые все лежат в одной плоскости. Железо прочно (ковалентно) связано с глобулярным белком через атомы N имидазол кольцо F8 гистидин остаток (также известный как проксимальный гистидин) под порфириновым кольцом. Шестая позиция может обратимо связывать кислород посредством координировать ковалентную связь,[43] завершая октаэдрическую группу из шести лигандов. Эта обратимая связь с кислородом - вот почему гемоглобин так полезен для транспортировки кислорода по телу.[44] Кислород связывается по геометрии «с изогнутым концом», когда один атом кислорода связывается с Fe, а другой выступает под углом. Когда кислород не связан, очень слабосвязанная молекула воды заполняет участок, образуя искаженный октаэдр.
Несмотря на то, что диоксид углерода переносится гемоглобином, он не конкурирует с кислородом за позиции связывания железа, а связывается с аминогруппами белковых цепей, присоединенных к гемовым группам.
Ион железа может находиться либо в железо Fe2+ или в железо Fe3+ состояние, но ферригемоглобин (метгемоглобин ) (Fe3+) не может связывать кислород.[45] При связывании кислород временно и обратимо окисляется (Fe2+) в (Fe3+), а кислород временно превращается в супероксид ион, таким образом, железо должно существовать в степени окисления +2, чтобы связывать кислород. Если супероксид-ион связан с Fe3+ протонируется, железо гемоглобина остается окисленным и неспособным связывать кислород. В таких случаях фермент метгемоглобин редуктаза сможет в конечном итоге реактивировать метгемоглобин за счет восстановления центра железа.
У взрослых людей наиболее распространенным типом гемоглобина является гемоглобин. тетрамер (который содержит четыре субъединичных белка), называемый гемоглобин А, состоящий из двух нековалентно связанных α и двух β субъединиц, каждая из которых состоит из 141 и 146 аминокислотных остатков соответственно. Это обозначается как α2β2. Субъединицы структурно похожи и примерно одного размера. Каждая субъединица имеет молекулярную массу около 16000дальтон,[46] в общей сложности молекулярный вес тетрамера примерно 64000 дальтон (64458 г / моль).[47] Таким образом, 1 г / дл = 0,1551 ммоль / л. Гемоглобин А - наиболее изученная из молекул гемоглобина.
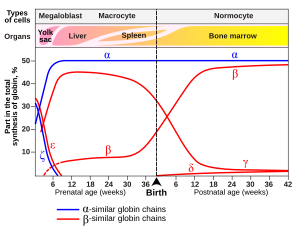
У младенцев человека молекула гемоглобина состоит из 2 цепей α и 2 цепей γ. Гамма-цепи постепенно заменяются β-цепями по мере роста ребенка.[48]
Четверка полипептидные цепи связаны друг с другом соляные мосты, водородные связи, а гидрофобный эффект.
Насыщение кислородом
Как правило, гемоглобин может быть насыщен молекулами кислорода (оксигемоглобин) или ненасыщен молекулами кислорода (дезоксигемоглобин).[49]
Оксигемоглобин
Оксигемоглобин формируется во время физиологическое дыхание когда кислород связывается с гемовым компонентом белка гемоглобина в красных кровяных тельцах. Этот процесс происходит в легочные капилляры рядом с альвеолы легких. Затем кислород проходит через кровоток и попадает в клетки, где он используется в качестве конечного акцептора электронов при производстве АТФ в процессе окислительного фосфорилирования. Однако это не помогает противодействовать снижению pH крови. Вентиляция, или дыхание, может изменить это состояние путем удаления углекислый газ, вызывая тем самым сдвиг pH.[50]
Гемоглобин существует в двух формах: натянутая (напряженная) форма (T) и a расслабленная форма (Р). Различные факторы, такие как низкий pH, высокий CO2 и высокий 2,3 BPG на уровне тканей предпочитают тугую форму, которая имеет низкое сродство к кислороду и выделяет кислород в тканях. И наоборот, высокий pH, низкий CO2, или низкий 2,3 BPG способствует расслабленной форме, которая может лучше связывать кислород.[51] Парциальное давление системы также влияет на O2 сродство, когда при высоких парциальных давлениях кислорода (таких как те, которые присутствуют в альвеолах) благоприятно расслабленное (высокое сродство, R) состояние. И наоборот, при низких парциальных давлениях (таких как те, которые присутствуют в дышащих тканях), напряженное состояние (низкое сродство, T) является предпочтительным.[52] Кроме того, связывание кислорода с гемом железа (II) притягивает железо к плоскости порфиринового кольца, вызывая небольшой конформационный сдвиг. Сдвиг побуждает кислород связываться с тремя оставшимися гемовыми единицами в гемоглобине (таким образом, связывание кислорода является кооперативным).
Деоксигенированный гемоглобин
Деоксигенированный гемоглобин - это форма гемоглобина без связанного кислорода. В спектры поглощения оксигемоглобина и дезоксигемоглобина различаются. Оксигемоглобин имеет значительно меньшее поглощение на длине волны 660 нм. длина волны чем дезоксигемоглобин, а при 940 нм его поглощение несколько выше. Эта разница используется для измерения количества кислорода в крови пациента с помощью прибора, называемого пульсоксиметр. Это различие также объясняет представление цианоз, от синего до пурпурного цвета, который ткани развиваются во время гипоксия.[53]
Деоксигенированный гемоглобин парамагнитный; его слабо привлекают магнитные поля.[54][55] Напротив, оксигенированный гемоглобин демонстрирует диамагнетизм, слабое отталкивание от магнитного поля.[55]
Эволюция гемоглобина позвоночных
Ученые согласны с тем, что событие, отделившее миоглобин от гемоглобина, произошло после миноги отклонился от челюстные позвоночные.[56] Это разделение миоглобина и гемоглобина позволило возникать и развиваться различным функциям двух молекул: миоглобин больше связан с хранением кислорода, а гемоглобин отвечает за перенос кислорода.[57] Гены α- и β-подобных глобинов кодируют отдельные субъединицы белка.[25] Предшественники этих генов возникли в результате другого события дупликации, также после того, как общий предок гнатосом произошел от бесчелюстных рыб, примерно 450–500 миллионов лет назад.[56] Исследования реконструкции предков предполагают, что преддупликационным предком α и β генов был димер, состоящий из идентичных субъединиц глобина, которые затем эволюционировали, чтобы собраться в тетрамерную архитектуру после дупликации.[58] Развитие генов α и β создало потенциал для того, чтобы гемоглобин состоял из нескольких отдельных субъединиц, физический состав которых имеет ключевое значение для способности гемоглобина переносить кислород. Наличие нескольких субъединиц способствует способности гемоглобина кооперативно связывать кислород, а также аллостерически регулироваться.[57][58] Впоследствии ген α также подвергся дупликации с образованием HBA1 и HBA2 гены.[59] Эти дальнейшие дупликации и дивергенции создали разнообразный диапазон α- и β-подобных глобиновых генов, которые регулируются таким образом, что определенные формы встречаются на разных стадиях развития.[57]
Самая ледяная рыба семейства Каннихтииды потеряли гены гемоглобина из-за адаптации к холодной воде.[5]
Степень окисления железа в оксигемоглобине
Определить степень окисления оксигенированного гемоглобина сложно, потому что оксигемоглобин (Hb-O2), согласно экспериментальным измерениям, является диамагнитным (нет чистых неспаренных электронов), однако электронная конфигурация с самой низкой энергией (в основном состоянии) как в кислороде, так и в железе - парамагнитный (что предполагает наличие в комплексе по крайней мере одного неспаренного электрона). Форма кислорода с наименьшей энергией и формы с наименьшей энергией соответствующих степеней окисления железа следующие:
- Триплетный кислород, молекулярный кислород с наименьшей энергией, имеет два неспаренных электрона на разрыхляющих π * молекулярных орбиталях.
- Железо (II) имеет тенденцию существовать в высокоспиновом 3D6 конфигурация с четырьмя неспаренными электронами.
- Железо (III) (3d5) имеет нечетное количество электронов и, следовательно, должен иметь один или несколько неспаренных электронов в любом энергетическом состоянии.
Все эти структуры парамагнитны (имеют неспаренные электроны), а не диамагнитны. Таким образом, неинтуитивное (например, более высокая энергия по крайней мере для одного вида) распределение электронов в комбинации железа и кислорода должно существовать, чтобы объяснить наблюдаемый диамагнетизм и отсутствие неспаренных электронов.
Две логические возможности производить диамагнитный (без чистого спина) Hb-O2 находятся:
- Низкоспиновый Fe2+ связывается с синглетный кислород. И низкоспиновое железо, и синглетный кислород диамагнитны. Однако единственная форма кислорода - это форма молекулы с более высокой энергией.
- Низкоспиновый Fe3+ связывается с O2•− (в супероксид ion), и два неспаренных электрона связываются антиферромагнитно, обеспечивая наблюдаемые диамагнитные свойства. Здесь железо окислилось (потеряло один электрон), а кислород восстановился (получил один электрон).
Другая возможная модель, в которой низкоспиновый Fe4+ связывается с пероксидом, O22−, можно исключить само по себе, потому что железо парамагнитно (хотя ион пероксида диамагнитен). Здесь железо окисляется двумя электронами, а кислород восстанавливается двумя электронами.
Прямые экспериментальные данные:
- Рентгеновская фотоэлектронная спектроскопия предполагает, что железо имеет степень окисления примерно 3,2.
- Инфракрасные колебательные частоты связи O-O предполагает соответствие длины связи с супероксидом (порядок связи примерно 1,6, с супероксидом 1,5).
- Краевые структуры с поглощением рентгеновского излучения на железной К-кромке. Энергетический сдвиг на 5 эВ между дезоксигемоглобином и оксигемоглобином, как и для всех разновидностей метгемоглобина, убедительно свидетельствует о фактическом локальном заряде ближе к Fe3+ чем Fe2+.[60][61][62]
Таким образом, ближайшая формальная степень окисления железа в Hb-O2 состояние +3, с кислородом в состоянии -1 (как супероксид .О2−). Диамагнетизм в этой конфигурации возникает из-за того, что одиночный неспаренный электрон на супероксиде антиферромагнитно выравнивается с одиночным неспаренным электроном на железе (в низкоспиновом d5 состояние), чтобы не давать никакого чистого вращения всей конфигурации, в соответствии с диамагнитным оксигемоглобином из эксперимента.[63][64]
Второй выбор логических возможностей, приведенных выше для диамагнитного оксигемоглобина, который был найден правильным экспериментально, неудивителен: синглетный кислород (возможность № 1) является нереально высокоэнергетическим состоянием. Модель 3 приводит к неблагоприятному разделению зарядов (и не согласуется с магнитными данными), хотя она может внести незначительный вклад в качестве резонанс форма. Переход железа к более высокой степени окисления в Hb-O2 уменьшает размер атома и позволяет ему попасть в плоскость порфиринового кольца, притягивая координированный остаток гистидина и инициируя аллостерические изменения, наблюдаемые в глобулинах.
Ранние постулаты био-неорганических химиков утверждали, что возможность № 1 (выше) верна и что железо должно существовать в степени окисления II. Этот вывод казался правдоподобным, поскольку степень окисления железа III как метгемоглобин, когда нет в сопровождении супероксида .О2− "удерживать" электрон окисления, как было известно, делает гемоглобин неспособным связывать нормальный триплет O2 как это происходит в воздухе. Таким образом, предполагалось, что железо остается в виде Fe (II), когда газообразный кислород связывается в легких. Химия железа в этой предыдущей классической модели была элегантной, но необходимое присутствие диамагнитной высокоэнергетической молекулы синглетного кислорода так и не было объяснено. Классически утверждалось, что связывание молекулы кислорода помещает высокоспиновое железо (II) в октаэдрическое поле сильнопольных лигандов; это изменение поля увеличит энергия расщепления кристаллического поля, заставляя электроны железа спариваться в низкоспиновую конфигурацию, которая была бы диамагнитной в Fe (II). Считается, что это вынужденное низкоспиновое спаривание происходит в железе при связывании кислорода, но этого недостаточно, чтобы объяснить изменение размера железа. Извлечение дополнительного электрона из железа кислородом требуется для объяснения как меньшего размера железа, так и наблюдаемой повышенной степени окисления, а также более слабой связи кислорода.
Назначение целочисленной степени окисления - формализм, поскольку ковалентные связи не обязаны иметь совершенные порядки связи, включающие полный перенос электрона. Таким образом, все три модели парамагнитного Hb-O2 может вносить небольшой вклад (за счет резонанса) в реальную электронную конфигурацию Hb-O2. Однако модель железа в Hb-O2 быть Fe (III) более правильно, чем классическая идея, что он остается Fe (II).
Сотрудничество

Когда кислород связывается с комплексом железа, он заставляет атом железа двигаться назад к центру плоскости порфирин кольцо (см. подвижную диаграмму). В то же время имидазол боковая цепь остатка гистидина, взаимодействующего на другом полюсе железа, тянется к порфириновому кольцу. Это взаимодействие сдвигает плоскость кольца в сторону наружу от тетрамера, а также вызывает деформацию спирали белка, содержащей гистидин, по мере того, как она приближается к атому железа. Этот штамм передается оставшимся трем мономерам в тетрамере, где он вызывает аналогичные конформационные изменения в других сайтах гема, так что связывание кислорода с этими сайтами становится легче.
Когда кислород связывается с одним мономером гемоглобина, конформация тетрамера переходит из T (напряженного) состояния в R (расслабленное) состояние. Этот сдвиг способствует связыванию кислорода с остальными тремя группами гема мономера, тем самым насыщая молекулу гемоглобина кислородом.[65]
В тетрамерной форме нормального взрослого гемоглобина связывание кислорода, таким образом, является совместный процесс. Сродство связывания гемоглобина с кислородом увеличивается за счет насыщения молекулы кислородом, при этом первые молекулы, связанные с кислородом, влияют на форму участков связывания для следующих, что благоприятно для связывания. Эта положительная кооперативная привязка достигается за счет стерический конформационные изменения белкового комплекса гемоглобина, как обсуждалось выше; то есть, когда одна субъединица белка в гемоглобине становится оксигенированной, инициируется конформационное или структурное изменение всего комплекса, в результате чего другие субъединицы приобретают повышенное сродство к кислороду. Как следствие, кривая связывания кислорода гемоглобина имеет вид сигмовидный, или же S-образный, в отличие от нормального гиперболический кривая, связанная с некооперативным связыванием.
Динамический механизм кооперативности гемоглобина и его связь с низкочастотными резонанс обсуждалось.[66]
Связывание с лигандами, отличными от кислорода
Помимо кислорода лиганд, который кооперативно связывается с гемоглобином, лиганды гемоглобина также включают конкурентные ингибиторы Такие как монооксид углерода (CO) и аллостерические лиганды Такие как углекислый газ (CO2) и оксид азота (НЕТ). Двуокись углерода связывается с аминогруппами белков глобина с образованием карбаминогемоглобин; Считается, что этот механизм обеспечивает около 10% переноса углекислого газа у млекопитающих. Оксид азота также может переноситься гемоглобином; это связано с конкретными тиол группы в белке глобина с образованием S-нитрозотиола, который снова диссоциирует на свободный оксид азота и тиол, поскольку гемоглобин высвобождает кислород из своего гемового участка. Предполагается, что этот транспорт оксида азота к периферическим тканям способствует транспорту кислорода в тканях путем высвобождения сосудорасширяющий оксида азота к тканям с низким уровнем кислорода.[67]
Конкурентный
На связывание кислорода влияют такие молекулы, как оксид углерода (например, из курение табака, выхлопной газ, и неполное сгорание в печах). CO конкурирует с кислородом в сайте связывания гема. Сродство гемоглобина к связыванию CO в 250 раз превышает его сродство к кислороду,[68][69] Это означает, что небольшие количества CO резко снижают способность гемоглобина доставлять кислород к ткани-мишени.[70] Поскольку окись углерода представляет собой бесцветный газ без запаха и вкуса и представляет собой потенциально смертельную угрозу, детекторы окиси углерода стали коммерчески доступными для предупреждения об опасных уровнях в жилых домах. Когда гемоглобин соединяется с CO, он образует очень ярко-красное соединение, называемое карбоксигемоглобин, что может вызвать появление на коже Отравление CO жертвы после смерти выглядят розовыми, а не белыми или синими. Когда вдыхаемый воздух содержит уровень CO всего 0,02%, Головная боль и тошнота происходить; если концентрация CO увеличивается до 0,1%, наступает бессознательное состояние. У заядлых курильщиков углекислый газ может блокировать до 20% активных кислородных центров.
Аналогичным образом гемоглобин также обладает конкурентным сродством связывания с цианид (CN−), окись серы (SO), и сульфид (S2−), включая сероводород (ЧАС2S). Все они связываются с железом в геме, не меняя его степени окисления, но, тем не менее, они ингибируют связывание кислорода, вызывая серьезную токсичность.
Атом железа в гемовой группе должен изначально находиться в железо (Fe2+) степень окисления для поддержки связывания и транспорта кислорода и других газов (он временно переключается на трехвалентное железо, пока кислород связан, как объяснено выше). Первоначальное окисление до железо (Fe3+) состояние без кислорода превращает гемоглобин в «гемоглобин».яглобин »или метгемоглобин, который не может связывать кислород. Гемоглобин в нормальных эритроцитах защищен системой восстановления, чтобы этого не произошло. Оксид азота способен преобразовывать небольшую часть гемоглобина в метгемоглобин в красных кровяных тельцах. Последняя реакция - это остаток деятельности более древних диоксигеназа оксида азота функция глобинов.
Аллостерический
Углерод диоксид занимает другое место связывания в гемоглобине. В тканях, где концентрация углекислого газа выше, углекислый газ связывается с аллостерическим участком гемоглобина, облегчая разгрузку кислорода из гемоглобина и, в конечном итоге, его удаление из организма после того, как кислород высвобождается в ткани, подвергающиеся метаболизму. Это повышенное сродство венозной крови к углекислому газу известно как Эффект Бора. Через фермент карбоангидраза, углекислый газ реагирует с водой с образованием угольная кислота, который распадается на бикарбонат и протоны:
- CO2 + H2O → H2CO3 → HCO3− + H+

Следовательно, в крови с высоким уровнем углекислого газа также ниже pH (более кислый ). Гемоглобин может связывать протоны и углекислый газ, что вызывает изменение конформации белка и способствует высвобождению кислорода. Протоны связываются в разных местах белка, а диоксид углерода связывается с α-аминогруппой.[71] Двуокись углерода связывается с гемоглобином и образует карбаминогемоглобин.[72] Это снижение сродства гемоглобина к кислороду за счет связывания диоксида углерода и кислоты известно как Эффект Бора. Эффект Бора отдает предпочтение состоянию T, а не состоянию R. (сдвигает O2-кривая насыщения до верно). И наоборот, когда уровень углекислого газа в крови снижается (то есть в капиллярах легких), углекислый газ и протоны высвобождаются из гемоглобина, увеличивая сродство белка к кислороду. Снижение общей связывающей способности гемоглобина по отношению к кислороду (т. Е. Смещение кривой вниз, а не только вправо) из-за снижения pH называется корневой эффект. Это наблюдается у костистой рыбы.
Гемоглобину необходимо выделять кислород, который он связывает; в противном случае нет смысла его связывать. Сигмоидальная кривая гемоглобина делает его эффективным в связывании (поглощение O2 в легких) и эффективен при разгрузке (разгрузка O2 в тканях).[73]
У людей, акклиматизировавшихся на больших высотах, концентрация 2,3-бисфосфоглицерат (2,3-BPG) в крови повышается, что позволяет этим людям доставлять большее количество кислорода к тканям в условиях более низкого напряжение кислорода. Это явление, когда молекула Y влияет на связывание молекулы X с транспортной молекулой Z, называется гетеротропный аллостерический эффект. Гемоглобин у организмов на больших высотах также адаптировался так, что он имеет меньшее сродство к 2,3-BPG, и поэтому белок будет больше смещен в сторону своего R. В состоянии R гемоглобин будет легче связывать кислород, что позволяет организмам выполнять необходимые метаболические процессы, когда кислород присутствует при низких парциальных давлениях.[74]
Животные, кроме людей, используют другие молекулы для связывания с гемоглобином и изменения его O2 близость при неблагоприятных условиях. Рыба использует оба АТФ и GTP. Они связываются с фосфатным «карманом» на молекуле гемоглобина рыбы, который стабилизирует напряженное состояние и, следовательно, снижает сродство к кислороду.[75] ГТФ снижает сродство гемоглобина к кислороду намного больше, чем АТФ, что, как считается, связано с дополнительным водородная связь формируется, что дополнительно стабилизирует напряженное состояние.[76] В условиях гипоксии концентрация как АТФ, так и ГТФ в красных кровяных тельцах рыб снижается, что увеличивает сродство к кислороду.[77]
Вариант гемоглобина, называемый гемоглобин плода (HbF, α2γ2), находится в развивающихся плод, и связывает кислород с большим сродством, чем взрослый гемоглобин. Это означает, что кривая связывания кислорода для гемоглобина плода смещена влево (т. Е. Более высокий процент гемоглобина имеет кислород, связанный с ним при более низком напряжении кислорода), по сравнению с кривой гемоглобина взрослого человека. В результате кровь плода в плацента способен забирать кислород из материнской крови.
Гемоглобин также несет оксид азота (NO) в глобиновой части молекулы. Это улучшает доставку кислорода на периферию и способствует контролю дыхания. NO обратимо связывается со специфическим остатком цистеина в глобине; связывание зависит от состояния (R или T) гемоглобина. Образующийся в результате S-нитрозилированный гемоглобин влияет на различные активности, связанные с NO, такие как контроль сосудистого сопротивления, артериального давления и дыхания. NO не выделяется в цитоплазме эритроцитов, а транспортируется из них с помощью анионообменника, называемого AE1.[78]
Типы у людей
Варианты гемоглобина являются частью нормального эмбриональный и плод разработка. Они также могут быть патологическими мутантными формами гемоглобина в численность населения, вызванные вариациями в генетике. Некоторые известные варианты гемоглобина, такие как серповидноклеточная анемия, несут ответственность за заболевания и считаются гемоглобинопатии. Другие варианты не обнаруживаются патология, и поэтому считаются непатологическими вариантами.[79][80]
в эмбрион:
У плода:
- Гемоглобин F (α2γ2) (PDB: 1FDH).
После рождения:
- Гемоглобин А (взрослый гемоглобин) (α2β2) (PDB: 1BZ0) - самая обычная с нормальным количеством более 95%
- Гемоглобин А2 (α2δ2) - синтез δ-цепи начинается в конце третьего триместра, и у взрослых он имеет нормальный диапазон 1,5–3,5%.
- Гемоглобин F (гемоглобин плода) (α2γ2) - У взрослых гемоглобин F ограничен ограниченной популяцией эритроцитов, называемых F-клетками. Однако уровень Hb F может быть повышен у людей с серповидно-клеточной анемией и бета-талассемия.
Вариантные формы, вызывающие заболевание:
- Гемоглобин D-Пенджаб - (α2βD2) - вариантная форма гемоглобина.
- Гемоглобин H (β4) - Вариант формы гемоглобина, образованный тетрамером β-цепей, который может присутствовать в вариантах α талассемия.
- Гемоглобин Бартс (γ4) - вариантная форма гемоглобина, образованная тетрамером γ-цепей, которая может присутствовать при вариантах α-талассемии.
- Гемоглобин S (α2βS2) - вариантная форма гемоглобина, обнаруженная у людей с серповидно-клеточной анемией. Существует разновидность гена β-цепи, вызывающая изменение свойств гемоглобина, что приводит к серповидному выбросу красных кровяных телец.
- Гемоглобин С (α2βC2) - Другой вариант из-за вариации гена β-цепи. Этот вариант вызывает легкое хроническое гемолитическая анемия.
- Гемоглобин E (α2βE2) - Другой вариант из-за вариации гена β-цепи. Этот вариант вызывает легкую хроническую гемолитическую анемию.
- Гемоглобин AS - гетерозиготная форма, вызывающая серповидноклеточная черта с одним взрослым геном и одним геном серповидно-клеточной анемии
- Болезнь гемоглобина SC - сложная гетерозиготная форма с одним серповидным геном и другим кодирующим Гемоглобин С.
- Гемоглобин Хопкинса-2 - Вариант формы гемоглобина, который иногда рассматривается в сочетании с Гемоглобин S вызвать серповидно-клеточную анемию.
Деградация у позвоночных животных
Когда красные кровяные тельца достигают конца своей жизни из-за старения или дефектов, они удаляются из кровотока в результате фагоцитарной активности макрофагов в селезенке или печени или гемолизируются в кровотоке. Свободный гемоглобин затем выводится из кровотока через транспортер гемоглобина CD163, который экспрессируется исключительно на моноцитах или макрофагах. Внутри этих клеток молекула гемоглобина расщепляется, а железо перерабатывается. Этот процесс также производит одну молекулу монооксида углерода на каждую молекулу разложенного гема.[81] Разложение гема является одним из немногих естественных источников окиси углерода в организме человека и отвечает за нормальный уровень окиси углерода в крови даже у людей, дышащих воздухом.[нужна цитата ] Другим основным конечным продуктом разложения гема является билирубин. Повышенный уровень этого химического вещества обнаруживается в крови, если красные кровяные тельца разрушаются быстрее, чем обычно. Неправильно расщепленный белок гемоглобина или гемоглобин, который слишком быстро высвобождается из клеток крови, может закупорить мелкие кровеносные сосуды, особенно тонкие сосуды, фильтрующие кровь. почки, вызывая повреждение почек. Железо удаляется из гема и используется для дальнейшего использования, оно хранится в виде гемосидерина или ферритин в тканях и переносится в плазме бета-глобулинами в виде трансферрины. Когда порфириновое кольцо разрывается, фрагменты обычно секретируются в виде желтого пигмента, называемого билирубином, который секретируется в кишечник в виде желчи. В кишечнике билирубин превращается в уробилиноген. Уробилиноген покидает организм с фекалиями в виде пигмента, называемого стеркобилин. Глобулин метаболизируется в аминокислоты, которые затем попадают в кровоток.
Роль в болезни
Дефицит гемоглобина может быть вызван либо пониженным количеством молекул гемоглобина, как в анемия или снижением способности каждой молекулы связывать кислород при том же парциальном давлении кислорода. Гемоглобинопатии (генетические дефекты, приводящие к аномальной структуре молекулы гемоглобина)[82] может вызвать оба. В любом случае дефицит гемоглобина уменьшается. способность крови переносить кислород. Дефицит гемоглобина, как правило, строго отличается от гипоксемия, определяемое как уменьшенное частичное давление кислорода в крови,[83][84][85][86] хотя оба являются причинами гипоксия (недостаточное снабжение тканей кислородом).
Другие частые причины низкого гемоглобина включают потерю крови, дефицит питания, проблемы с костным мозгом, химиотерапию, почечную недостаточность или аномальный гемоглобин (например, при серповидно-клеточной анемии).
Способность каждой молекулы гемоглобина переносить кислород обычно изменяется изменением pH крови или CO2, вызывая измененный кривая диссоциации кислород-гемоглобин. Однако он также может быть патологически изменен, например, в отравление угарным газом.
Снижение гемоглобина с абсолютным уменьшением количества эритроцитов или без него приводит к появлению симптомов анемии. Анемия имеет много разных причин, хотя недостаток железа и его результат железодефицитная анемия являются наиболее частыми причинами в западном мире. Поскольку отсутствие железа снижает синтез гема, эритроциты при железодефицитной анемии гипохромный (без красного пигмента гемоглобина) и микроцитарный (меньше обычного). Другие анемии встречаются реже. В гемолиз (ускоренный распад красных кровяных телец), связанный желтуха вызывается метаболитом гемоглобина билирубином, а циркулирующий гемоглобин может вызывать почечная недостаточность.
Некоторые мутации в цепи глобина связаны с гемоглобинопатии, таких как серповидно-клеточная анемия и талассемия. Другие мутации, обсуждаемые в начале статьи, являются доброкачественными и обозначаются просто как варианты гемоглобина.
Существует группа генетических нарушений, известных как порфирии которые характеризуются ошибками в метаболических путях синтеза гема. король Георг III Соединенного Королевства был, вероятно, самым известным больным порфирией.
В небольшой степени гемоглобин А медленно соединяется с глюкоза на конце валина (альфа-аминокислота) каждой β-цепи. Полученную молекулу часто называют Hb A1c, а гликозилированный гемоглобин. Связывание глюкозы с аминокислотами в гемоглобине происходит спонтанно (без помощи фермента) во многих белках и, как известно, не служит полезной цели. Однако по мере увеличения концентрации глюкозы в крови процент Hb A, который превращается в Hb A1c увеличивается. В диабетики чья глюкоза обычно высока, процент Hb A1c также работает высоко. Из-за низкой скорости комбинации Hb A с глюкозой Hb A1c процент отражает средневзвешенное значение уровня глюкозы в крови за время жизни эритроцитов, которое составляет примерно 120 дней.[87] Поэтому уровни гликозилированного гемоглобина измеряются для мониторинга долгосрочного контроля над хроническим заболеванием сахарным диабетом 2 типа (СД2). Плохой контроль СД2 приводит к высокому уровню гликозилированного гемоглобина в эритроцитах. Нормальный эталонный диапазон составляет приблизительно 4,0–5,9%. Несмотря на то, что получить данные трудно, для людей с СД2 рекомендуются значения менее 7%. Уровни выше 9% связаны с плохим контролем гликозилированного гемоглобина, а уровни выше 12% связаны с очень плохим контролем. У диабетиков, у которых уровень гликозилированного гемоглобина близок к 7%, гораздо больше шансов избежать осложнений, которые могут сопровождать диабет (чем у тех, у кого уровень гликозилированного гемоглобина составляет 8% или выше).[88] Кроме того, повышенное гликозилирование гемоглобина увеличивает его сродство к кислороду, тем самым предотвращая его высвобождение в ткани и в крайних случаях вызывая гипоксию.[89]
Повышенный уровень гемоглобина связан с увеличением количества или размера эритроцитов, называемых полицитемия. Это повышение может быть вызвано врожденный порок сердца, легочное сердце, легочный фиброз, слишком много эритропоэтин, или же истинная полицитемия.[90] Высокий уровень гемоглобина также может быть вызван пребыванием на большой высоте, курением, обезвоживанием (искусственно путем концентрации гемоглобина), тяжелыми заболеваниями легких и некоторыми опухолями.[48]
Недавнее исследование, проведенное в Пондичерри, Индия, показывает его важность при ишемической болезни сердца.[91]
Диагностическое использование

Измерение концентрации гемоглобина - одно из наиболее часто выполняемых анализы крови, обычно как часть полный анализ крови. Например, его обычно тестируют до или после донорство крови. Результаты представлены в грамм /L, грамм/дл или же моль / L. 1 г / дл равен примерно 0,6206 ммоль / л, хотя последние единицы не используются так часто из-за неопределенности относительно полимерного состояния молекулы.[92] Этот коэффициент пересчета с использованием единой молекулярной массы глобина 16000 Да, более характерен для концентрации гемоглобина в крови. Для MCHC (средней концентрации корпускулярного гемоглобина) более распространен коэффициент преобразования 0,155, который использует массу тетрамера 64 500 Да.[93] Нормальные уровни:
- Мужчины: от 13,8 до 18,0 г / дл (от 138 до 180 г / л или от 8,56 до 11,17 ммоль / л)
- Женщины: от 12,1 до 15,1 г / дл (от 121 до 151 г / л или от 7,51 до 9,37 ммоль / л)
- Дети: от 11 до 16 г / дл (от 110 до 160 г / л или от 6,83 до 9,93 ммоль / л)
- Беременные женщины: от 11 до 14 г / дл (от 110 до 140 г / л, или от 6,83 до 8,69 ммоль / л) (от 9,5 до 15 обычных значений во время беременности)[94][95]
Нормальные значения гемоглобина в 1-м и 3-м триместрах беременных должны быть не менее 11 г / дл и не менее 10,5 г / дл во 2-м триместре.[96]
Обезвоживание или гипергидратация могут сильно повлиять на измеряемый уровень гемоглобина. Альбумин может указывать на состояние гидратации.
Если концентрация ниже нормы, это называется анемией. Анемии классифицируются по размеру красных кровяных телец, клеток, которые содержат гемоглобин у позвоночных. Анемия называется «микроцитарной», если эритроциты маленькие, «макроцитарной», если они большие, и «нормоцитарной» в противном случае.
Гематокрит доля объема крови, занятая эритроцитами, обычно примерно в три раза превышает концентрацию гемоглобина, измеренную в г / дл. Например, если уровень гемоглобина составляет 17 г / дл, это сопоставимо с гематокритом 51%.[97]
Лабораторные методы определения гемоглобина требуют взятия пробы крови (артериальной, венозной или капиллярной) и анализа на гематологическом анализаторе и СО-оксиметре. Кроме того, доступен новый неинвазивный метод определения гемоглобина (SpHb), называемый пульсовой CO-оксиметрией, с точностью, сравнимой с инвазивными методами.[98]
Концентрации оксигемоглобина и дезоксигемоглобина можно измерять непрерывно, регионально и неинвазивно, используя NIRS.[99][100][101][102][103] NIRS можно применять как на голове, так и на мышцах. Этот метод часто используется для исследований, например, элитная спортивная подготовка, эргономика, реабилитация, наблюдение за пациентами, неонатальные исследования, функциональный мониторинг мозга, интерфейс мозг-компьютер, урология (сокращение мочевого пузыря), неврология (нервно-сосудистая связь) и многое другое.
Долгосрочный контроль содержание сахара в крови концентрацию можно измерить по концентрации Hb A1c. Для его прямого измерения потребуется много образцов, потому что уровень сахара в крови сильно варьируется в течение дня. Hb A1c продукт необратимая реакция гемоглобина А с глюкозой. Более высокий уровень глюкозы концентрация приводит к увеличению Hb A1c. Поскольку реакция медленная, Hb A1c Пропорция представляет собой средний уровень глюкозы в крови за период полураспада эритроцитов, обычно составляет 50–55 дней. Hb A1c доля 6,0% или менее показывает хороший долгосрочный контроль уровня глюкозы, тогда как значения выше 7,0% являются повышенными. Этот тест особенно полезен для диабетиков.[104]
В функциональная магнитно-резонансная томография (ФМРТ) аппарат использует сигнал от дезоксигемоглобина, который чувствителен к магнитным полям, поскольку является парамагнитным. Комбинированное измерение с NIRS показывает хорошую корреляцию как с сигналом оксигемоглобина, так и с сигналом дезоксигемоглобина по сравнению с сигналом Жирный сигнал.[105]
Использование спортивного и самотрекинга
Гемоглобин можно отслеживать неинвазивно, чтобы создать индивидуальный набор данных, отслеживающих эффекты гемоконцентрации и гемодилюции при повседневной активности, для лучшего понимания спортивных результатов и тренировок. Спортсменов часто беспокоят выносливость и интенсивность упражнений. В датчике используются светоизлучающие диоды, которые излучают красный и инфракрасный свет через ткань на световой датчик, который затем отправляет сигнал процессору для расчета поглощения света белком гемоглобина.[106]Этот датчик похож на пульсоксиметр, который состоит из небольшого чувствительного устройства, которое крепится к пальцу.
Аналоги у беспозвоночных организмов
Разнообразные белки, переносящие и связывающие кислород, существуют в организмах во всех царствах животных и растений. Организмы, включая бактерии, простейшие, и грибы все они содержат гемоглобиноподобные белки, чьи известные и предсказанные роли включают обратимое связывание газообразных лиганды. Поскольку многие из этих белков содержат глобины и гем часть (железо в плоской порфириновой подложке), их часто называют гемоглобинами, даже если их общая третичная структура сильно отличается от структуры гемоглобина позвоночных. В частности, различение «миоглобина» и гемоглобина у низших животных часто невозможно, поскольку некоторые из этих организмов не содержат мышцы. Или у них может быть узнаваемый отдельный сердечно-сосудистая система но не тот, который занимается транспортом кислорода (например, многие насекомые и другие членистоногие ). Во всех этих группах молекулы, содержащие гем / глобин (даже мономерные глобиновые), которые имеют дело со связыванием газа, называются оксигемоглобинами. Помимо транспортировки и определения кислорода, они также могут иметь дело с NO, CO.2, сульфидные соединения и даже O2 уборка мусора в среде, которая должна быть анаэробной.[107] Они могут даже заниматься детоксикацией хлорированных материалов аналогично гемсодержащим ферментам P450 и пероксидазам.

Структура гемоглобинов различается у разных видов. Гемоглобин встречается во всех царствах организмов, но не во всех организмах. Примитивные виды, такие как бактерии, простейшие, водоросли, и растения часто имеют моноглобиновые гемоглобины. Много нематода черви, моллюски, и ракообразные содержат очень большие мультисубъединичные молекулы, намного больше, чем у позвоночных. В частности, химерные гемоглобины, обнаруженные в грибы и гигант кольчатые червя может содержать как глобин, так и другие типы белков.[14]
Одним из наиболее ярких примеров использования гемоглобина в организмах является гигантский трубчатый червь (Рифтия пахиптила, также называемый Вестиментифера), который может достигать 2,4 метра в длину и населяет океан. вулканические жерла. Вместо пищеварительный тракт эти черви содержат популяцию бактерий, составляющую половину веса организма. Бактерии окисляют H2S из вентиляционного отверстия с O2 из воды для производства энергии для приготовления пищи из H2O и CO2. Верхний конец червей представляет собой веерообразную структуру темно-красного цвета («шлейф»), которая простирается в воду и поглощает H2S и O2 для бактерий и CO2 для использования в качестве синтетического сырья, аналогичного фотосинтетическим растениям. Структуры имеют ярко-красный цвет из-за содержания в них нескольких чрезвычайно сложных гемоглобинов, которые имеют до 144 цепей глобина, каждая из которых включает связанные гемовые структуры. Эти гемоглобины примечательны тем, что способны переносить кислород в присутствии сульфида и даже нести сульфид, не будучи полностью им «отравленным» или подавленным им, как гемоглобины у большинства других видов.[108][109]
Другие связывающие кислород белки
- Миоглобин
- Обнаруженный в мышечной ткани многих позвоночных животных, включая человека, он придает мышечной ткани отчетливый красный или темно-серый цвет. Он очень похож на гемоглобин по структуре и последовательности, но не является тетрамером; вместо этого это мономер, у которого отсутствует кооперативное связывание. Он используется для хранения кислорода, а не для его транспортировки.
- Гемоцианин
- Второй наиболее распространенный белок, переносящий кислород в природе, содержится в крови многих членистоногих и моллюсков. Использует простетические группы меди вместо гемовых групп железа и имеет синий цвет при насыщении кислородом.
- Гемеритрин
- Некоторые морские беспозвоночные и несколько видов кольчатых червей используют этот железосодержащий негемовый белок для переноса кислорода в кровь. При насыщении кислородом выглядит розово-фиолетовым, в противном случае - прозрачным.
- Хлорокруорин
- Обнаруженный у многих кольчатых червей, он очень похож на эритрокруорин, но группа гема значительно отличается по структуре. Зеленый при обескислороженном состоянии и красный при насыщении кислородом.
- Ванабинс
- Также известный как ванадий хромагены, они находятся в крови морские брызги. Однажды они выдвинули гипотезу об использовании редкого металла ванадия в качестве простетической группы, связывающей кислород. Однако, хотя они предпочтительно содержат ванадий, они, по-видимому, связывают мало кислорода и, таким образом, выполняют некоторые другие функции, которые не были выяснены (морские брызги также содержат некоторое количество гемоглобина). Они могут действовать как токсины.
- Эритрокруорин
- Встречается у многих кольчатых червей, в том числе у дождевые черви, это гигантский свободно плавающий белок крови, содержащий многие десятки, а возможно, и сотни субъединиц белков, несущих железо и гем, связанных вместе в единый белковый комплекс с молекулярной массой более 3,5 миллионов дальтон.
- Пиннаглобин
- Видно только у моллюска Pinna nobilis. Коричневый порфириновый белок на основе марганца.
- Леггемоглобин
- У зернобобовых растений, таких как люцерна или соя, азотфиксирующие бактерии в корнях защищены от кислорода этим железным гемом, содержащим кислородсвязывающий белок. Защищенный специфический фермент нитрогеназа, который не способен восстанавливать газообразный азот в присутствии свободного кислорода.
- Кобоглобин
- Синтетический порфирин на основе кобальта. Кобопротеин будет бесцветным при насыщении кислородом и желтым при попадании в вены.
Наличие в неэритроидных клетках
Некоторые неэритроидные клетки (то есть клетки, отличные от линии красных кровяных телец) содержат гемоглобин. В мозгу к ним относится A9 дофаминергический нейроны в черная субстанция, астроциты в кора головного мозга и гиппокамп, и во всем зрелом олигодендроциты.[12] Было высказано предположение, что гемоглобин головного мозга в этих клетках может обеспечивать «хранение кислорода для обеспечения гомеостатического механизма в аноксических условиях, что особенно важно для нейронов A9 DA, которые имеют повышенный метаболизм с высокой потребностью в выработке энергии».[12] Далее было отмечено, что "A9 дофаминергический нейроны могут подвергаться особому риску, поскольку в дополнение к своей высокой митохондриальной активности они подвергаются интенсивному окислительному стрессу, вызванному выработкой перекиси водорода посредством автоокисления и / или опосредованного моноаминоксидазой (МАО) дезаминирования дофамина и последующей реакции доступного двухвалентного железа. генерировать высокотоксичные гидроксильные радикалы ».[12] Это может объяснить риск дегенерации этих клеток в болезнь Паркинсона.[12] Полученное из гемоглобина железо в этих клетках не является причиной посмертной темноты этих клеток (происхождение латинского названия, субстанция негр), а скорее связано с нейромеланин.
Вне мозга гемоглобин не несет кислородных функций как антиоксидант и регулятор метаболизм железа в макрофаги,[110] альвеолярные клетки,[111] и мезангиальные клетки в почках.[112]
В истории, искусстве и музыке

Исторически ассоциация между цветом крови и ржавчиной возникает в ассоциации планеты Марс, с римским богом войны, так как планета оранжево-красная, что напомнило древним кровь. Хотя цвет планеты обусловлен соединениями железа в сочетании с кислородом в марсианской почве, распространено заблуждение, что железо в гемоглобине и его оксидах придает крови красный цвет. Цвет на самом деле из-за порфирин часть гемоглобина, с которым связано железо, а не самого железа,[113] хотя лигирование и окислительно-восстановительное состояние железа может влиять на электронные переходы порфирина с пи на пи * или от n до пи * и, следовательно, на его оптические характеристики.
Художник Джулиан Восс-Андреэ создал скульптура называется Сердце из стали (гемоглобин) в 2005 году на основе основы белка. Скульптура изготовлена из стекла и атмосферостойкая сталь. Намеренное ржавление первоначально блестящего произведения искусства отражает фундаментальную химическую реакцию гемоглобина - связывание кислорода с железом.[114][115]
Монреальский художник Николас Байер создал Блеск (Гемоглобин), скульптура из нержавеющей стали, демонстрирующая структуру молекулы гемоглобина. Он отображается в атриуме Центр здоровья Университета Макгилла исследовательский центр в Монреале. Скульптура имеет размеры около 10 метров × 10 метров × 10 метров.[116][117]
Смотрите также
Варианты гемоглобина: Белковые субъединицы (гены) гемоглобина: Соединения гемоглобина:
|
|
Рекомендации
- ^ Джонс, Дэниел (2003) [1917], Питер Роуч; Джеймс Хартманн; Джейн Сеттер (ред.), Словарь английского произношения, Кембридж: Издательство Кембриджского университета, ISBN 978-3125396838
- ^ «Гемоглобин». Dictionary.com Несокращенный. Случайный дом.
- ^ «Гемоглобин». Словарь Merriam-Webster.
- ^ Матон, Антея; Жан Хопкинс; Чарльз Уильям Маклафлин; Сьюзан Джонсон; Марианна Куон Уорнер; Дэвид ЛаХарт; Джилл Д. Райт (1993). Биология человека и здоровье. Энглвуд Клиффс, Нью-Джерси, США: Прентис Холл. ISBN 978-0139811760.
- ^ а б Сиделл, Брюс; Кристин О'Брайен (2006). «Когда плохие вещи случаются с хорошими рыбами: потеря гемоглобина и экспрессии миоглобина у антарктических ледяных рыб». Журнал экспериментальной биологии. 209 (Pt 10): 1791–802. Дои:10.1242 / jeb.02091. PMID 16651546.
- ^ Сорняк, Роберт I .; Рид, Клод Ф .; Берг, Джордж (1963). «Является ли гемоглобин важным структурным компонентом мембран эритроцитов человека?». J Clin Invest. 42 (4): 581–88. Дои:10.1172 / JCI104747. ЧВК 289318. PMID 13999462.
- ^ Домингес де Вильота ED, Руис Кармона MT, Рубио JJ, де Андрес S (1981). «Равенство кислородсвязывающей способности гемоглобина in vivo и in vitro у пациентов с тяжелым респираторным заболеванием». Br J Anaesth. 53 (12): 1325–28. Дои:10.1093 / bja / 53.12.1325. PMID 7317251. S2CID 10029560.
- ^ Костанцо, Линда С. (2007). Физиология. Хагерствон, доктор медицины: Липпинкотт Уильямс и Уилкинс. ISBN 978-0781773119.
- ^ Паттон, Кевин Т. (10 февраля 2015 г.). Анатомия и психология. Elsevier Health Sciences. ISBN 9780323316873. В архиве из оригинала от 26.04.2016. Получено 2016-01-09.
- ^ Эпштейн, Ф. Х .; Ся, К. С. У. (1998). «Дыхательная функция гемоглобина». Медицинский журнал Новой Англии. 338 (4): 239–47. Дои:10.1056 / NEJM199801223380407. PMID 9435331.
- ^ Саха Д., Редди К.В. и др. (2014). "Экспрессия гемоглобина в неэритроидных клетках: новинка или повсеместное распространение?". Int J Воспламенение. 2014 (803237): 1–8. Дои:10.1155/2014/803237. ЧВК 4241286. PMID 25431740.
- ^ а б c d е Biagioli M, Pinto M, Cesselli D и др. (2009). «Неожиданная экспрессия альфа- и бета-глобина в мезэнцефальных дофаминергических нейронах и глиальных клетках». Proc. Natl. Акад. Наука. 106 (36): 15454–59. Bibcode:2009PNAS..10615454B. Дои:10.1073 / pnas.0813216106. ЧВК 2732704. PMID 19717439.
- ^ «Анализы крови». Национальный институт сердца, легких и крови (NHLBI). В архиве из оригинала на 2019-04-09. Получено 2019-04-27.
- ^ а б Вебер Р.Е., Виноградов С.Н. (2001). «Гемоглобины беспозвоночных: функции и молекулярные адаптации». Physiol. Rev. 81 (2): 569–628. Дои:10.1152 / Physrev.2001.81.2.569. PMID 11274340. S2CID 10863037.
- ^ "Макс Перуц, отец молекулярной биологии, умер в возрасте 87 лет В архиве 2016-04-23 в Wayback Machine ". Нью-Йорк Таймс. 8 февраля 2002 г.
- ^ Энгельхарт, Иоганн Фридрих (1825). Commentatio de vera materia sanguini purpureum colorem impertientis natura (на латыни). Геттинген: Дитрих.
- ^ "Энгельхард и Роуз о окраске крови". Эдинбургский медико-хирургический журнал. 27 (90): 95–102. 1827. ЧВК 5763191. PMID 30330061.
- ^ Адэр, Гилберт Смитсон (1925). «Критическое исследование прямого метода измерения осмотического давления гемоглобина». Proc. R. Soc. Лондон. А 108 (750): 292–300. Bibcode:1925RSPSA.109..292A. Дои:10.1098 / rspa.1925.0126.
- ^ Хюнефельд Ф. (1840). "Die Chemismus in der thierischen Organization". Лейпциг. Цитировать журнал требует
| журнал =(помощь) - ^ Функе О. (1851). "Über das milzvenenblut". Z Rat Med. 1: 172–218.
- ^ «Рецепт НАСА для кристаллографии белков» (PDF). Обучающая записка. Национальное управление по аэронавтике и исследованию космического пространства. Архивировано из оригинал (PDF) на 2008-04-10. Получено 2008-10-12.
- ^ Хоппе-Зейлер F (1866 г.). "Uber die oxydation in lebendem blute". Лаборатория Med-Chem Untersuch. 1: 133–40.
- ^ Perutz, M.F .; Россманн, М.Г .; Cullis, A.F .; Muirhead, H .; Will, G .; Север, A.C.T. (1960). «Структура гемоглобина: трехмерный синтез Фурье с разрешением 5,5 А, полученный с помощью рентгеновского анализа». Природа. 185 (4711): 416–22. Bibcode:1960Натура.185..416П. Дои:10.1038 / 185416a0. PMID 18990801. S2CID 4208282.
- ^ Перуц М.Ф. (1960). «Структура гемоглобина». Брукхейвенские симпозиумы по биологии. 13: 165–83. PMID 13734651.
- ^ а б Хардисон, Росс К. (2012-12-01). «Эволюция гемоглобина и его генов». Перспективы Колд-Спринг-Харбор в медицине. 2 (12): a011627. Дои:10.1101 / cshperspect.a011627. ISSN 2157-1422. ЧВК 3543078. PMID 23209182.
- ^ Оффнер, Сьюзан (01.04.2010). «Использование баз данных генома NCBI для сравнения генов бета-гемоглобина человека и шимпанзе». Американский учитель биологии. 72 (4): 252–256. Дои:10.1525 / abt.2010.72.4.10. ISSN 0002-7685. S2CID 84499907.
- ^ «HBB - субъединица гемоглобина бета - Pan paniscus (карликовый шимпанзе) - ген и белок HBB». www.uniprot.org. Получено 2020-03-10.
- ^ «HBA1 - субъединица гемоглобина альфа - пантроглодиты (шимпанзе) - ген и белок HBA1». www.uniprot.org. Получено 2020-03-10.
- ^ Программа вариантов человеческого гемоглобина (1996) В архиве 2006-09-01 на Wayback Machine. Globin.cse.psu.edu. Проверено 5 сентября 2013.
- ^ Варианты гемоглобина В архиве 2006-11-05 на Wayback Machine. Labtestsonline.org. Проверено 5 сентября 2013.
- ^ Усман, доктор медицины, изд. «Гемоглобинопатии и талассемии». Архивировано из оригинал на 2007-12-15. Получено 2007-12-26.
- ^ Рид, Лесли. «Адаптация обнаружена в генах мышей». Omaha World-Herald 11 августа 2009 г .: EBSCO. Интернет. 30 октября 2009 г.
- ^ «У мамонтов была антифризная кровь». BBC. 2010-05-02. В архиве из оригинала 2010-05-04. Получено 2010-05-02.
- ^ Проекто-Гарсия, Хоана; Натараджан, Чандрасекар; Морияма, Хидеаки; Вебер, Рой Э .; Фаго, Анджела; Cheviron, Zachary A .; Дадли, Роберт; Макгуайр, Джимми А .; Витт, Кристофер К. (17 декабря 2013 г.). «Повторяющиеся повышенные изменения функции гемоглобина в ходе эволюции андских колибри». Труды Национальной академии наук. 110 (51): 20669–74. Bibcode:2013ПНАС..11020669П. Дои:10.1073 / pnas.1315456110. ISSN 0027-8424. ЧВК 3870697. PMID 24297909.
- ^ Beall, Cynthia M .; Сонг, Киджунг; Элстон, Роберт С .; Гольдштейн, Мелвин С. (28 сентября 2004 г.). «Более высокая выживаемость потомства среди тибетских женщин с генотипами с высокой насыщенностью кислородом, проживающими на высоте 4000 м». Труды Национальной академии наук Соединенных Штатов Америки. 101 (39): 14300–04. Bibcode:2004ПНАС..10114300Б. Дои:10.1073 / pnas.0405949101. ISSN 0027-8424. ЧВК 521103. PMID 15353580.
- ^ «Синтез гемоглобина». 14 апреля 2002 г. В архиве с оригинала 26 декабря 2007 г.. Получено 2007-12-26.
- ^ Бурка, Эдвард (1969). «Характеристики деградации РНК в эритроидной клетке». Журнал клинических исследований. 48 (7): 1266–72. Дои:10.1172 / jci106092. ЧВК 322349. PMID 5794250. В архиве с оригинала 9 августа 2018 г.. Получено 8 октября 2014.
- ^ а б van Kessel et al. (2003) «2.4 Белки - Натуральные полиамиды». Химия 12. Торонто: Нельсон, с. 122.
- ^ «Учебник по гемоглобину». В архиве 2009-11-26 на Wayback Machine Массачусетский университет в Амхерсте. Интернет. 23 октября 2009 г.
- ^ Стейнберг, MH (2001). Нарушения гемоглобина: генетика, патофизиология и клиническое лечение. Издательство Кембриджского университета. п. 95. ISBN 978-0521632669. В архиве из оригинала на 17.11.2016. Получено 2016-02-18.
- ^ Hardison, RC (1996). «Краткая история гемоглобинов: растений, животных, простейших и бактерий». Proc Natl Acad Sci USA. 93 (12): 5675–79. Bibcode:1996PNAS ... 93.5675H. Дои:10.1073 / пнас.93.12.5675. ЧВК 39118. PMID 8650150.
- ^ «Гемоглобин». В архиве 2009-11-13 на Wayback Machine Школа химии - Бристольский университет - Великобритания. Интернет. 12 октября 2009 г.
- ^ WikiPremed> Координационная химия В архиве 2009-08-23 на Wayback Machine. Проверено 2 июля 2009 г.
- ^ Основы биологии (2015). "Кровяные клетки".
- ^ Линберг Р., Коновер С.Д., Шум К.Л., Шорр Р.Г. (1998). «Переносчики кислорода на основе гемоглобина: сколько метгемоглобина - это слишком много?». Artif Cells Blood Substit Immobil Biotechnol. 26 (2): 133–48. Дои:10.3109/10731199809119772. PMID 9564432.
- ^ Гемоглобин В архиве 2017-03-15 в Wayback Machine. Worthington-biochem.com. Проверено 5 сентября 2013.
- ^ Ван Биквельт MC, Colier WN, Wevers RA, Van Engelen BG (2001). «Проведение спектроскопии в ближнем инфракрасном диапазоне при измерении местного потребления O2 и кровотока в скелетных мышцах». J Appl Physiol. 90 (2): 511–19. Дои:10.1152 / jappl.2001.90.2.511. PMID 11160049.
- ^ а б «Гемоглобин». В архиве 2012-01-24 в Wayback Machine MedicineNet. Интернет. 12 октября 2009 г.
- ^ «Гемоглобин домашний». В архиве 2009-12-01 в Wayback Machine Биология @ Дэвидсон. Интернет. 12 октября 2009 г.
- ^ «График насыщения гемоглобина». altitude.org. Архивировано из оригинал на 31.08.2010. Получено 2010-07-06.
- ^ Кинг, Майкл В. "Страница медицинской биохимии - Гемоглобин". В архиве из оригинала от 04.03.2012. Получено 2012-03-20.
- ^ Воет, Д. (2008) Основы биохимии, 3-й. изд., рис. 07_06, John Wiley & Sons. ISBN 0470129301
- ^ Аренс; Кимберли, Бэшем (1993). Основы оксигенации: значение для клинической практики. Джонс и Бартлетт Обучение. п. 194. ISBN 978-0867203325.
- ^ Огава, S; Menon, R. S .; Tank, D.W .; Kim, S. G .; Merkle, H; Ellermann, J.M .; Угурбил, К (1993). «Функциональное картирование мозга с помощью контрастной магнитно-резонансной томографии в зависимости от уровня оксигенации крови. Сравнение характеристик сигнала с биофизической моделью». Биофизический журнал. 64 (3): 803–12. Bibcode:1993BpJ .... 64..803O. Дои:10.1016 / S0006-3495 (93) 81441-3. ЧВК 1262394. PMID 8386018.
- ^ а б Брен К.Л., Айзенберг Р., Грей HB (2015). «Открытие магнитного поведения гемоглобина: начало биоинорганической химии». Proc Natl Acad Sci U S A. 112 (43): 13123–27. Bibcode:2015ПНАС..11213123Б. Дои:10.1073 / pnas.1515704112. ЧВК 4629386. PMID 26508205.
- ^ а б Гудман, Моррис; Мур, Г. Уильям; Мацуда, Гэндзи (1975-02-20). «Дарвиновская эволюция в генеалогии гемоглобина». Природа. 253 (5493): 603–08. Bibcode:1975Натура.253..603Г. Дои:10.1038 / 253603a0. PMID 1089897. S2CID 2979887.
- ^ а б c Сторц, Джей Ф .; Opazo, Juan C .; Хоффманн, Федерико Г. (01.02.2013). «Дублирование генов, дупликация генома и функциональная диверсификация глобинов позвоночных». Молекулярная филогенетика и эволюция. 66 (2): 469–78. Дои:10.1016 / j.ympev.2012.07.013. ISSN 1095-9513. ЧВК 4306229. PMID 22846683.
- ^ а б Pillai, Arvind S .; Чендлер, Шейн А .; Лю, Ян; Синьор, Энтони В .; Кортез-Ромеро, Карлос Р.; Benesch, Justin L.P .; Лагановский, Артур; Сторц, Джей Ф .; Хохберг, Георг К. А .; Торнтон, Джозеф В. (май 2020 г.). «Происхождение сложности в эволюции гемоглобина». Природа. 581 (7809): 480–485. Дои:10.1038 / с41586-020-2292-у. ISSN 1476-4687.
- ^ Циммер, Э. А .; Martin, S.L .; Beverley, S.M .; Kan, Y. W .; Уилсон, А.С. (1980-04-01). «Быстрая дупликация и потеря генов, кодирующих альфа-цепи гемоглобина». Труды Национальной академии наук Соединенных Штатов Америки. 77 (4): 2158–62. Bibcode:1980PNAS ... 77.2158Z. Дои:10.1073 / pnas.77.4.2158. ISSN 0027-8424. ЧВК 348671. PMID 6929543.
- ^ Пин С., Альперт Б., Михалович А. (1982). «Кислородная связь в человеческом гемоглобине и его изолированных субъединицах: исследование XANES». FEBS Lett. 147 (1): 106–10. Дои:10.1016/0014-5793(82)81021-1. PMID 7140986. S2CID 5920899.
- ^ Pin, S .; Valat, P .; Cortes, R .; Michalowicz, A .; Альперт, Б. (1985). «Процессы связывания лигандов в гемоглобине. Химическая реакционная способность железа изучена с помощью спектроскопии XANES». Биофизический журнал. 48 (6): 997–1001. Bibcode:1985BpJ .... 48..997P. Дои:10.1016 / S0006-3495 (85) 83862-5. ЧВК 1329432. PMID 4092074.
- ^ Бьянкони А., Конгиу-Кастеллано А., Делль'Аричча М., Джованнелли А., Бураттини Е., Дарем П.Дж. (1985). «Повышение эффективного заряда Fe в гемопротеинах в процессе оксигенации». Сообщения о биохимических и биофизических исследованиях. 131 (1): 98–102. Дои:10.1016 / 0006-291X (85) 91775-9. PMID 4038310.
- ^ Чайлдс ЧП (2001). «Гемоглобин - молекулярное легкое: 2». Химия в действии (65). ISSN 0332-2637. Архивировано из оригинал на 16 января 2009 г.
- ^ Чен Х, Икеда-Сайто М, Шайк С (2008). «Природа связывания Fe-O2 в окси-миоглобине: эффект белка». Журнал Американского химического общества. 130 (44): 14778–90. Дои:10.1021 / ja805434m. PMID 18847206.
- ^ Михайлеску, Михаэла-Рита; Руссу, Ирина М. (27.03.2001). «Признак перехода T → R в гемоглобине человека». Труды Национальной академии наук Соединенных Штатов Америки. 98 (7): 3773–77. Bibcode:2001ПНАС ... 98.3773М. Дои:10.1073 / pnas.071493598. ISSN 0027-8424. ЧВК 31128. PMID 11259676.
- ^ Чжоу К.С. (1989). «Низкочастотный резонанс и кооперативность гемоглобина». Trends Biochem. Наука. 14 (6): 212–13. Дои:10.1016/0968-0004(89)90026-1. PMID 2763333.
- ^ Дженсен, Фрэнк Б. (2009). «Двойная роль красных кровяных телец в доставке кислорода тканям: переносчики кислорода и регуляторы местного кровотока». Журнал экспериментальной биологии. 212 (Пт 21): 3387–93. Дои:10.1242 / jeb.023697. PMID 19837879.
- ^ Холл, Джон Э. (2010). Гайтон и Холл учебник медицинской физиологии (12-е изд.). Филадельфия, Пенсильвания: Saunders / Elsevier. п. 502. ISBN 9781416045748.
- ^ Забудьте, Б.Г .; Банн, Х. Ф. (1 февраля 2013 г.). «Классификация нарушений гемоглобина». Перспективы Колд-Спринг-Харбор в медицине. Лаборатория Колд-Спринг-Харбор. 3 (2): a011684. Дои:10.1101 / cshperspect.a011684. ISSN 2157-1422. ЧВК 3552344. PMID 23378597.
- ^ Родос, Карл Э .; Варакалло, Мэтью (04.03.2019). «Физиология, кислородный транспорт». Книжная полка NCBI. PMID 30855920. Получено 2019-05-04.
Важно отметить, что в условиях карбоксигемоглобинемии патологией является не снижение способности переносить кислород, а нарушение доставки связанного кислорода к тканям-мишеням.
- ^ Nelson, D. L .; Кокс, М. М. (2000). Принципы биохимии Ленингера, 3-е изд. Нью-Йорк: Издательство Worth. п. 217, ISBN 1572599316.
- ^ Guyton, Arthur C .; Джон Э. Холл (2006). Учебник медицинской физиологии (11-е изд.). Филадельфия: Эльзевьер Сондерс. п. 511. ISBN 978-0721602400.
- ^ Лекция - 12 Миоглобин и гемоглобин на YouTube
- ^ Биохимия (Восьмое изд.). Нью-Йорк: У. Х. Фриман. 2015-04-08. ISBN 9781464126109.
- ^ Rutjes, H.A .; Nieveen, M.C .; Weber, R.E .; Витте, Ф .; Ван ден Тиллар, Г. Э. Дж. М. (20 июня 2007 г.). «Множественные стратегии цихлид озера Виктория, чтобы справиться с пожизненной гипоксией, включают переключение гемоглобина». AJP: регуляторная, интегративная и сравнительная физиология. 293 (3): R1376–83. Дои:10.1152 / ajpregu.00536.2006. PMID 17626121.
- ^ Gronenborn, Angela M .; Clore, G.Marius; Брунори, Маурицио; Джардина, Бруно; Фальчони, Джанкарло; Перуц, Макс Ф. (1984). «Стереохимия АТФ и ГТФ, связанных с гемоглобинами рыб». Журнал молекулярной биологии. 178 (3): 731–42. Дои:10.1016/0022-2836(84)90249-3. PMID 6492161.
- ^ Вебер, Рой Э .; Фрэнк Б. Дженсен (1988). «Функциональные адаптации гемоглобинов экзотермических позвоночных». Ежегодный обзор физиологии. 50: 161–79. Дои:10.1146 / annurev.ph.50.030188.001113. PMID 3288089.
- ^ Rang, H.P .; Dale M.M .; Ritter J.M .; Мур П.К. (2003). Фармакология, пятое издание. Эльзевир. ISBN 978-0443072024.
- ^ «Варианты гемоглобина». Лабораторные тесты онлайн. Американская ассоциация клинической химии. 2007-11-10. В архиве из оригинала от 20.09.2008. Получено 2008-10-12.
- ^ Huisman THJ (1996). "Программа вариантов человеческого гемоглобина". Генный сервер Globin. Государственный университет Пенсильвании. В архиве из оригинала 11.12.2008. Получено 2008-10-12.
- ^ Kikuchi, G .; Yoshida, T .; Ногучи, М. (2005). «Гемоксигеназа и деградация гема». Сообщения о биохимических и биофизических исследованиях. 338 (1): 558–67. Дои:10.1016 / j.bbrc.2005.08.020. PMID 16115609.
- ^ "гемоглобинопатия " в Медицинский словарь Дорланда
- ^ гипоксемия В архиве 2009-02-02 в Wayback Machine. Британская энциклопедия, заявляя гипоксемия (снижение напряжения кислорода в крови).
- ^ Biology-Online.org -> Словарь »H» Гипоксемия В архиве 2009-11-21 на Wayback Machine последнее изменение 29 декабря 2008 г.
- ^ Уильям, К. Уилсон; Гранде, Кристофер М .; Хойт, Дэвид Б. (2007). «Патофизиология острой дыхательной недостаточности». Травма, том II: интенсивная терапия. Тейлор и Фрэнсис. п. 430. ISBN 9781420016840. В архиве из оригинала на 17.11.2016. Получено 2016-02-18.
- ^ Макгаффиган, П. А. (1996). «Опасности гипоксемии: как уберечь пациента от низкого уровня кислорода». Уход. 26 (5): 41–46, тест 46. Дои:10.1097/00152193-199626050-00013. PMID 8710285.
- ^ «NGSP: HbA1c и eAG». www.ngsp.org. В архиве с оригинала на 2015-10-15. Получено 2015-10-28.
- ^ «Определение гликозилированного гемоглобина». В архиве 2014-01-23 на Wayback Machine Сеть медицины. Интернет. 12 октября 2009 г.
- ^ Madsen, H; Дицель, Дж (1984). «Транспорт кислорода в крови в первом триместре диабетической беременности». Acta Obstetricia et Gynecologica Scandinavica. 63 (4): 317–20. Дои:10.3109/00016348409155523. PMID 6741458. S2CID 12771673.
- ^ Гемоглобин В архиве 2016-06-10 в Wayback Machine в Medline Plus
- ^ Padmanaban, P .; Тура, Б. (2011). «Гемоглобин: новый маркер стабильной ишемической болезни сердца». Хроники молодых ученых. 2 (2): 109. Дои:10.4103/2229-5186.82971.
- ^ Общество биомедицинских исследований диабета. Калькулятор преобразования единиц СИ В архиве 2013-03-09 в Wayback Machine.
- ^ Handin, Роберт I .; Люкс, Сэмюэл Э. и Стоссель Блад, Томас П. (2003). Кровь: принципы и практика гематологии. Липпинкотт Уильямс и Уилкинс, ISBN 0781719933
- ^ Тест на уровень гемоглобина В архиве 2007-01-29 на Wayback Machine. Ibdcrohns.about.com (16 августа 2013 г.). Проверено 5 сентября 2013.
- ^ Хотя другие источники могут иметь немного разные значения, например гемоглобин (контрольный диапазон) В архиве 2009-09-25 на Wayback Machine. gpnotebook.co.uk
- ^ Мюррей С.С. и МакКинни Е.С. (2006). Основы ухода за матерями и новорожденными. 4-е изд., С. 919. Филадельфия: Сондерс Эльзевьер. ISBN 1416001417.
- ^ «Гематокрит (HCT) или объем упакованных клеток (PCV)». DoctorsLounge.com. В архиве из оригинала от 02.01.2008. Получено 2007-12-26.
- ^ Frasca, D .; Dahyot-Fizelier, C .; Екатерина, К .; Levrat, Q .; Debaene, B .; Мимоз, О. (2011). «Точность постоянного неинвазивного монитора гемоглобина у пациентов отделения интенсивной терапии *». Реанимационная медицина. 39 (10): 2277–82. Дои:10.1097 / CCM.0b013e3182227e2d. PMID 21666449. S2CID 205541592.
- ^ Феррари, М .; Binzoni, T .; Куаресима, В. (1997). «Окислительный метаболизм в мышцах». Философские труды Королевского общества B: биологические науки. 352 (1354): 677–83. Bibcode:1997RSPTB.352..677F. Дои:10.1098 / рстб.1997.0049. ЧВК 1691965. PMID 9232855.
- ^ Madsen, P. L .; Секер, Н. Х. (1999). «Ближняя инфракрасная оксиметрия головного мозга». Прогресс в нейробиологии. 58 (6): 541–60. Дои:10.1016 / S0301-0082 (98) 00093-8. PMID 10408656. S2CID 1092056.
- ^ McCully, K. K .; Хамаока, Т. (2000). «Спектроскопия в ближнем инфракрасном диапазоне: что она может сказать нам о насыщении кислородом скелетных мышц?». Обзоры упражнений и спортивных наук. 28 (3): 123–27. PMID 10916704.
- ^ Перри, С. П. (2008). «Неинвазивная NIR-спектроскопия функции мозга человека во время физических упражнений». Методы. 45 (4): 289–99. Дои:10.1016 / j.ymeth.2008.04.005. PMID 18539160.
- ^ Рольф, П. (2000). «Инвивонар-Инфракрасная спектроскопия». Ежегодный обзор биомедицинской инженерии. 2: 715–54. Дои:10.1146 / annurev.bioeng.2.1.715. PMID 11701529.
- ^ Это Hb A1c Уровень полезен только для людей, у которых есть красные кровяные тельца (эритроциты) с нормальной выживаемостью (то есть с нормальным периодом полураспада). У людей с патологическими эритроцитами, вызванными аномальными молекулами гемоглобина (такими как гемоглобин S при серповидноклеточной анемии) или дефектами мембран эритроцитов, или другими проблемами, период полужизни эритроцитов часто укорачивается. У этих людей может быть использован альтернативный тест, называемый «уровень фруктозамина». Он измеряет степень гликирования (связывания глюкозы) с альбумином, наиболее распространенным белком крови, и отражает средний уровень глюкозы в крови за предыдущие 18–21 день, который представляет собой период полураспада молекул альбумина в кровотоке.
- ^ Mehagnoul-Schipper DJ, van der Kallen BF, Colier WN, van der Sluijs MC, van Erning LJ, Thijssen HO, Oeseburg B, Hoefnagels WH, Jansen RW (2002). «Одновременное измерение изменений церебральной оксигенации во время активации мозга с помощью спектроскопии в ближнем инфракрасном диапазоне и функциональной магнитно-резонансной томографии у здоровых молодых и пожилых людей». Hum Brain Mapp. 16 (1): 14–23. Дои:10.1002 / hbm.10026. ЧВК 6871837. PMID 11870923.
- ^ «Cercacor - Как работает неинвазивная технология гемоглобина Ember». Cercacor - Как работает неинвазивная технология гемоглобина Ember. В архиве из оригинала от 04.11.2016. Получено 2016-11-03.
- ^ Л. Инт Панис; Б. Годдирис; Р. Верхейен (1995). «Концентрация гемоглобина у личинок Chironomus cf.Plumosus L. (Diptera: Chironomidae) из двух лентиных местообитаний». Нидерландский журнал водной экологии. 29 (1): 1–4. Дои:10.1007 / BF02061785. S2CID 34214741. В архиве из оригинала на 2018-09-05. Получено 2013-11-10.
- ^ Зал Ф, Лаллье Ф.Х., Грин Б.Н., Виноградов С.Н., Тулмонд А (1996). «Мультигемоглобиновая система червя гидротермальной вентиляции Riftia pachyptila. II. Полный состав полипептидной цепи исследован методом максимальной энтропии масс-спектров». J. Biol. Chem. 271 (15): 8875–81. Дои:10.1074 / jbc.271.15.8875. PMID 8621529.
- ^ Миник З., Эрве Г. (2004). «Биохимические и энзимологические аспекты симбиоза глубоководного трубчатого червя Riftia pachyptila и его бактериального эндосимбионта». Евро. J. Biochem. 271 (15): 3093–102. Дои:10.1111 / j.1432-1033.2004.04248.x. PMID 15265029.
- ^ Лю Л., Цзэн М., Стамлер Дж. С. (1999). «Индукция гемоглобина в макрофагах мыши». Труды Национальной академии наук Соединенных Штатов Америки. 96 (12): 6643–47. Bibcode:1999PNAS ... 96.6643L. Дои:10.1073 / pnas.96.12.6643. ЧВК 21968. PMID 10359765.
- ^ Ньютон Д.А., Рао К.М., Длухи Р.А., Баатц Дж. Э. (2006). «Гемоглобин экспрессируется альвеолярными эпителиальными клетками». Журнал биологической химии. 281 (9): 5668–76. Дои:10.1074 / jbc.M509314200. PMID 16407281.
- ^ Nishi, H .; Inagi, R .; Kato, H .; Tanemoto, M .; Кодзима, I .; Сын, Д .; Fujita, T .; Нангаку, М. (2008). «Гемоглобин экспрессируется мезангиальными клетками и снижает окислительный стресс». Журнал Американского общества нефрологов. 19 (8): 1500–08. Дои:10.1681 / ASN.2007101085. ЧВК 2488266. PMID 18448584.
- ^ Бох, Ларри (2001). Руководство по фармацевтической практике: Руководство по клиническому опыту. Липпинкотт Уильямс и Уилкинс. ISBN 978-0781725415.
- ^ Холден, Констанс (2005). «Кровь и сталь». Наука. 309 (5744): 2160. Дои:10.1126 / science.309.5744.2160d. S2CID 190178048.
- ^ Моран Л., Хортон Р.А., Скримджер Дж., Перри М. (2011). Принципы биохимии. Бостон, Массачусетс: Пирсон. п. 127. ISBN 978-0321707338.
- ^ Генри, Шон (7 августа 2014 г.). "Взгляните на коллекцию произведений искусства MUHC". CBC News. В архиве из оригинала 5 февраля 2016 г.. Получено 1 февраля, 2016.
- ^ «Блеск (Гемоглобин) 2014». Art Public Montréal. Монреаль. В архиве с оригинала 1 февраля 2016 г.. Получено 1 февраля, 2016.
дальнейшее чтение
| Библиотечные ресурсы о Гемоглобин |
- Кэмпбелл, МК (1999). Биохимия (третье изд.). Харкорт. ISBN 978-0030244261.
- Эшагян, С; Хорвич, ТБ; Фонаров, GC (2006). «Неожиданная обратная зависимость между уровнем HbA1c и смертностью у пациентов с диабетом и тяжелой систолической сердечной недостаточностью». Я сердце J. 151 (1): 91.e1–91.e6. Дои:10.1016 / j.ahj.2005.10.008. PMID 16368297.
- Ганонг, ВФ (2003). Обзор медицинской физиологии (21-е изд.). Ланге. ISBN 978-0071402361.
- Хагер, Т. (1995). Сила природы: жизнь Лайнуса Полинга. Саймон и Шустер. ISBN 978-0684809090.
- Кнейп Дж., Балакришнан Дж., Чен Р., Шен Т. Дж., Саху С. К., Хо Н. Т., Джованнелли Дж. Л., Симпласану В., Хо С., Спиро Т. (2005). «Динамика аллостерии в гемоглобине: роль предпоследних тирозиновых Н-связей». Дж Мол Биол. 356 (2): 335–53. Дои:10.1016 / j.jmb.2005.11.006. PMID 16368110.
Хардисон, Росс К. (2012). «Эволюция гемоглобина и его генов». Перспективы Колд-Спринг-Харбор в медицине. 2 (12): a011627. Дои:10.1101 / cshperspect.a011627. ISSN 2157-1422. ЧВК 3543078. PMID 23209182.
внешняя ссылка
- Протеопедия Гемоглобин
- Национальный совет действий при анемии - anemia.org
- Новый тип гемоглобина вызывает ложную диагностику с помощью пульсовых оксиметров
- Анимация гемоглобина: от дезокси к окси форме
Связанные вопросы:
- Идеальный уровень гемоглобина при беременности
- Как низко может упасть гемоглобин перед смертью
- Уровень гемоглобина