Растение - Plant
| Растения Временной диапазон: Мезопротерозой – настоящее время | |
|---|---|
 | |
| Научная классификация | |
| Домен: | Эукариоты |
| (без рейтинга): | Потогонные |
| (без рейтинга): | Archaeplastida |
| Королевство: | Plantae Sensu Коупленд, 1956 год. |
| Суперотделения | |
| |
| Синонимы | |
| |
Растения в основном многоклеточный организмов, преимущественно фотосинтетический эукариоты из Королевство Plantae. Исторически растения рассматривались как одно из двух царств, включая все живые существа, которые не были животные, и все водоросли и грибы рассматривались как растения. Однако все современные определения Plantae исключают грибы и некоторые водоросли, а также прокариоты (в археи и бактерии ). Согласно одному определению, растения образуют клады Viridiplantae (Латинское название «зеленые растения»), группа, которая включает цветущие растения, хвойные породы и другие голосеменные, папоротники и их союзники, роголистник, печеночники, мхи, а зеленые водоросли, но исключает красный и бурые водоросли.
Зеленые растения получают большую часть энергии от Солнечный лучик через фотосинтез первичным хлоропласты которые получены из эндосимбиоз с цианобактерии. Их хлоропласты содержат хлорофиллы a и b, что придает им зеленый цвет. Некоторые растения паразитический или микотрофный и потеряли способность производить нормальное количество хлорофилла или фотосинтезировать, но все еще имеют цветы, фрукты и семена. Растения характеризуются половое размножение и смена поколений, несмотря на то что бесполое размножение также обычное дело.
Их около 320000 разновидность растений, из которых подавляющее большинство, около 260–290 тысяч, производить семена.[5] Зеленые растения обеспечивают значительную часть молекулярного кислорода в мире,[6] и являются основой большинства экосистем Земли. Растения, производящие зерно, фрукты, и овощи также образуют основные продукты питания человека и были одомашненный тысячелетиями. У растений много культурный и другое использование, как украшения, строительные материалы, письменный материал и в большом разнообразии они были источник лекарств и психоактивные препараты. Научное изучение растений известно как ботаника, филиал биология.
Определение
Все живые существа традиционно относили к одной из двух групп: растения и животные. Эта классификация может датироваться Аристотель (384 г. до н.э. - 322 г. до н.э.), которые проводили различие между растениями, которые, как правило, неподвижны, и животными, которые часто мобильны, чтобы ловить пищу. Намного позже, когда Линней (1707–1778) заложили основу современной системы научная классификация эти две группы стали королевства Vegetabilia (позже Metaphyta или Plantae) и Animalia (также называемые Metazoa). С тех пор стало ясно, что царство растений, как первоначально определено, включает несколько не связанных между собой групп, и грибы и несколько групп водоросли были перемещены в новые королевства. Однако эти организмы все еще часто считаются растениями, особенно в популярном контексте.[нужна цитата ]
Термин «растение» обычно подразумевает наличие следующих признаков: многоклеточность, наличие клеточных стенок, содержащих целлюлоза, и способность осуществлять фотосинтез с первичными хлоропластами.[7][8]
Текущие определения Plantae
Когда название Plantae или растение применяется к определенной группе организмов или таксон, это обычно относится к одному из четырех понятий. Эти четыре группы, от наименее до наиболее всеобъемлющих, таковы:
| Имя (а) | Объем | Описание |
|---|---|---|
| Наземные растения, также известные как Эмбриофита | Plantae Sensu strictissimo | Растения в самом строгом смысле слова включить печеночники, роголистник, мхи, и сосудистые растения, а также ископаемые растения, похожие на эти выжившие группы (например, Metaphyta Уиттакер, 1969 год.,[9] Plantae Маргулис, 1971[10]). |
| Зеленые растения, также известный как Viridiplantae, Виридифита, Хлоробионта или Хлоропластида | Plantae Sensu stricto | Растения в строгом смысле слова включить зеленые водоросли, и появившиеся в них наземные растения, в том числе каменная кладка. Взаимоотношения между группами растений все еще прорабатываются, и названия, данные им, значительно различаются. В клады Viridiplantae - это группа организмов, у которых есть целлюлоза в их клеточные стенки, владеть хлорофиллы а и б и имеют пластиды связаны только двумя мембранами, способными к фотосинтезу и хранению крахмала. Эта клада является основной темой данной статьи (например, Plantae Copeland, 1956[11]). |
| Archaeplastida, также известный как Plastida или Primoplantae | Plantae Sensu lato | Растения в широком смысле включают перечисленные выше зеленые растения плюс красные водоросли (Родофита ) и глаукофитные водоросли (Глаукофита ) этот магазин Флоридский крахмал вне пластид, в цитоплазме. В эту кладу входят все организмы, которые эоны назад приобрели свои первичные хлоропласты непосредственно поглощая цианобактерии (например, Plantae Кавалер-Смит, 1981[12]). |
| Старые определения растения (устарело) | Plantae Sensu amplo | Растения в самом широком смысле относится к более старым, устаревшим классификациям, которые помещали в Plantae различные водоросли, грибы или бактерии (например, Plantae или Vegetabilia Линней,[13] Plantae Геккель 1866 г.,[14] Метафита Геккель, 1894 г.,[15] Plantae Уиттакер, 1969 год.[9]). |
Другой способ взглянуть на отношения между различными группами, которые были названы «растениями», - через кладограмма, который показывает их эволюционные отношения. Они еще не окончательно решены, но одна принятая связь между тремя описанными выше группами показана ниже[требуется разъяснение ].[16][17][18][19][20][21][22] Те, что были названы «растениями», выделены жирным шрифтом (некоторые второстепенные группы опущены).
| группы традиционно называется зеленые водоросли |
Способы объединения и названий групп зеленых водорослей существенно различаются у разных авторов.
Водоросли

Водоросли включают несколько различных групп организмов, которые производят пищу путем фотосинтеза и, таким образом, традиционно были включены в царство растений. В водоросли варьируются от крупных многоклеточных водорослей до одноклеточных организмов и подразделяются на три группы: зеленые водоросли, красные водоросли и бурые водоросли. Есть веские доказательства того, что бурые водоросли произошли независимо от других, от нефотосинтетических предков, которые сформировали эндосимбиотические отношения с красными водорослями, а не с цианобактериями, и они больше не классифицируются как растения, как определено здесь.[23][24]
Viridiplantae, зеленые растения - зеленые водоросли и наземные растения - образуют клады, группа, состоящая из всех потомков одного общего предка. За некоторыми исключениями, зеленые растения имеют следующие общие черты: первичный хлоропласты получено из цианобактерий, содержащих хлорофиллы а и б, клеточные стенки, содержащие целлюлоза, и продуктовые магазины в виде крахмал содержится в пластидах. Они проходят закрытые митоз без центриоли, и обычно митохондрии с плоскими кристами. В хлоропласты зеленых растений окружены двумя мембранами, что позволяет предположить, что они произошли непосредственно от эндосимбиотических цианобактерии.
Две дополнительные группы, Родофита (красные водоросли) и Глаукофита (глаукофитные водоросли) также имеют первичные хлоропласты, которые, по-видимому, происходят непосредственно из эндосимбиотических цианобактерии, хотя они отличаются от Viridiplantae пигментами, которые используются в фотосинтезе, и поэтому различаются по цвету. Эти группы также отличаются от зеленых растений тем, что запасающий полисахарид флоридский крахмал и хранится в цитоплазме, а не в пластидах. Похоже, что они имели общее происхождение с Viridiplantae, и эти три группы образуют кладу Archaeplastida, название которой подразумевает, что их хлоропласты произошли от одного древнего эндосимбиотического события. Это самое широкое современное определение термина «растение».
Напротив, большинство других водорослей (например, бурые водоросли / диатомеи, гаптофиты, динофлагелляты, и эвглениды ) не только имеют разные пигменты, но также имеют хлоропласты с тремя или четырьмя окружающими мембранами. Они не являются близкими родственниками Archaeplastida, предположительно приобретая хлоропласты отдельно от проглоченных или симбиотических зеленых и красных водорослей. Таким образом, они не включены даже в самое широкое современное определение царства растений, хотя они были в прошлом.
Зеленые растения или Viridiplantae традиционно подразделялись на зеленые водоросли (в том числе каменные) и наземные растения. Однако теперь известно, что наземные растения произошли из группы зеленых водорослей, так что зеленые водоросли сами по себе являются парафилетический group, то есть группа, исключающая некоторых потомков общего предка. В современных классификациях обычно избегают парафилетических групп, так что в недавних обработках Viridiplantae были разделены на две клады: Хлорофита и Streptophyta (включая наземные растения и Charophyta).[25][26]
Chlorophyta (название, которое также использовалось для все зеленые водоросли) являются сестринской группой харофитов, от которых произошли наземные растения. Насчитывается около 4300 видов,[27] в основном одноклеточные или многоклеточные морские организмы, такие как морской салат, Ульва.
Другая группа Viridiplantae - это в основном пресноводные или наземные Streptophyta, которые состоят из наземных растений вместе с Charophyta, которые, в свою очередь, состоят из нескольких групп зеленых водорослей, таких как десмиды и каменная кладка. Водоросли-стрептофиты бывают одноклеточными или образуют многоклеточные нити, разветвленные или неразветвленные.[26] Род Спирогира это нитчатая водоросль стрептофита, знакомая многим, поскольку она часто используется в обучении, и является одним из организмов, ответственных за водорослевую «пену» на прудах. Пресноводные каменные породы очень напоминают наземные растения и считаются их ближайшими родственниками.[нужна цитата ] Растущие в пресной воде, они состоят из центрального стебля с мутовками веточек.
Грибы
Линнея Первоначальная классификация помещала грибы в состав Plantae, поскольку они, несомненно, не были ни животными, ни минералами, и это были единственные другие альтернативы. С развитием 19 века в микробиология, Эрнст Геккель представил новое царство Protista в дополнение к Plantae и Animalia, но вопрос о том, лучше ли поместить грибы в Plantae или их следует реклассифицировать как протистов, остается спорным. В 1969 г. Роберт Уиттакер предложил создание королевства Грибов. С тех пор молекулярные данные показали, что самый последний общий предок (сопредок) грибов, вероятно, был больше похож на животных из животных, чем на планты или любое другое царство.[28]
Первоначальная реклассификация Уиттакера была основана на фундаментальной разнице в питании грибов и растений. В отличие от растений, которые обычно получают углерод посредством фотосинтеза, и так называемые автотрофы грибы не обладают хлоропластами и обычно получают углерод, разрушая и поглощая окружающие материалы, и поэтому называются гетеротрофный сапротрофы. Кроме того, субструктура многоклеточных грибов отличается от субструктуры растений, принимая форму множества хитиновых микроскопических нитей, называемых гифы, которые могут быть далее подразделены на ячейки или могут образовывать синцитий содержащий много эукариотический ядра. Плодовые тела, из которых грибы являются наиболее знакомым примером, репродуктивными структурами грибов, и не похожи ни на какие структуры, производимые растениями.[нужна цитата ]
Разнообразие
В приведенной ниже таблице показаны некоторые оценки количества видов различных видов зеленых растений (Viridiplantae). Около 85–90% всех растений - цветковые. В настоящее время несколько проектов пытаются собрать все виды растений в онлайн-базах данных, например то Мировая Флора Онлайн и Мировые растения оба перечисляют около 350 000 видов.[29][30]
| Неформальная группа | Название подразделения | Распространенное имя | Кол-во живых видов | Примерное количество в неформальной группе |
|---|---|---|---|---|
| Зеленые водоросли | Хлорофита | зеленые водоросли (хлорофиты) | 3,800–4,300 [31][32] | 8,500 (6,600–10,300) |
| Чарофита | зеленые водоросли (например. десмиды & каменная кладка ) | 2,800–6,000 [33][34] | ||
| Мохообразные | Мархантиофита | печеночники | 6,000–8,000 [35] | 19,000 (18,100–20,200) |
| Anthocerotophyta | роголистник | 100–200 [36] | ||
| Мохообразные | мхи | 12,000 [37] | ||
| Птеридофиты | Lycopodiophyta | клубные мхи | 1,200 [24] | 12,000 (12,200) |
| Pteridophyta | папоротники, венчик папоротник и хвощ | 11,000 [24] | ||
| Семенные растения | Cycadophyta | саговники | 160 [38] | 260,000 (259,511) |
| Гинкгофита | гинкго | 1 [39] | ||
| Pinophyta | хвойные породы | 630 [24] | ||
| Гнетофита | гнетофиты | 70 [24] | ||
| Магнолиофита | цветущие растения | 258,650 [40] |
Наименование растений регулируется Международный кодекс номенклатуры водорослей, грибов и растений и Международный кодекс номенклатуры культурных растений (увидеть таксономия культурных растений ).
Эволюция
Эволюция растений привела к увеличению уровни сложности, с самого раннего маты из водорослей, через мохообразные, ликоподы, папоротники к комплексу голосеменные и покрытосеменные сегодня. Растения всех этих групп продолжают развиваться, особенно в той среде, в которой они развивались.
На земле образовалась водорослевая пена 1,200 миллион лет назад, но так было до Ордовикский период, около 450 миллион лет назад, что появились наземные растения.[41] Однако новые данные, полученные при изучении соотношений изотопов углерода в докембрийских породах, позволили предположить, что сложные фотосинтетические растения развивались на Земле более 1000 млн лет назад.[42] Более века считалось, что предки наземных растений эволюционировали в водной среде, а затем адаптировались к жизни на суше, идея, которую обычно приписывают ботаникам. Фредерик Орпен Бауэр в его книге 1908 года Происхождение наземной флоры. Недавняя альтернативная точка зрения, подтвержденная генетическими доказательствами, заключается в том, что они произошли от наземных одноклеточных водорослей,[43] и что даже общий предок красных и зеленых водорослей и одноклеточных пресноводных водорослей глаукофиты, возникшие в земной среде в пресноводных биопленках или микробных матах.[44] Примитивные наземные растения начали диверсифицироваться в конце Силурийский период, около 420 миллион лет назад, а результаты их диверсификации замечательно подробно показаны в Девонский скопление окаменелостей из Рини черт. В этом кремне сохранились клеточные детали ранних растений, окаменевших в вулканических источниках. К середине девонского периода большинство признаков, присущих растениям, присутствуют сегодня, включая корни, листья и вторичную древесину, а к позднему девонскому периоду появились семена.[45] Таким образом, позднедевонские растения достигли такой степени сложности, которая позволила им образовывать леса из высоких деревьев. Эволюционные инновации продолжались в каменноугольном и более позднем геологических периодах и продолжаются сегодня. Большинство групп растений не пострадали от Событие пермо-триасового вымирания, хотя структура сообществ изменилась. Это могло создать основу для эволюции цветковых растений в триасе (~200 миллион лет назад), взорвавшиеся в меловом и третичном периодах. Последней крупной группой растений, которые эволюционировали, были травы, которые стали важными в середине третичного периода, примерно со времен 40 миллион лет назад. Травы, как и многие другие группы, развили новые механизмы метаболизма, чтобы выжить в условиях низкой CO
2 и теплые, сухие условия тропиков за последние 10 миллион лет.
Предложенный 1997 г. филогенетическое дерево из Plantae, после Кенрика и Крейна,[46] выглядит следующим образом, с модификацией Pteridophyta от Smith и другие.[47] В Prasinophyceae площадь парафилетический совокупность ранних расходящихся линий зеленых водорослей, но рассматриваются как группа за пределами Chlorophyta:[48] более поздние авторы не последовали этому предложению.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Более новая предложенная классификация следует Leliaert et al. 2011 г.[49] и модифицирован с помощью Silar 2016[20][21][50][51] за клады зеленых водорослей и Новиков и Барабаш-Красни 2015[52] для клады наземных растений. Обратите внимание, что здесь Prasinophyceae помещены внутри Chlorophyta.
|
Позже была предложена филогения, основанная на геномах и транскриптомах 1153 видов растений.[53] Размещение групп водорослей подтверждается филогенезом, основанным на геномах Mesostigmatophyceae и Chlorokybophyceae, которые с тех пор были секвенированы.[54][55] Классификация Bryophyta поддерживается как Puttick и другие. 2018,[56] и филогенезом с участием геномов роголистника, которые с тех пор также были секвенированы.[57][58]
| сорт хлорофитных водорослей стрептофитные водоросли | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Эмбриофиты

Наиболее знакомые нам растения - это многоклеточный наземные растения, называемые эмбриофиты. Эмбриофиты включают сосудистые растения, например папоротники, хвойные и цветковые растения. Они также включают мохообразные, из которых мхи и печеночники являются наиболее распространенными.
Все эти растения имеют эукариотический клетки с клеточные стенки состоит из целлюлоза, и большинство из них получают энергию через фотосинтез, с помощью свет, вода и углекислый газ синтезировать пищу. Около трехсот видов растений не фотосинтезируют, но паразиты на другие виды фотосинтезирующих растений. Эмбриофиты отличаются от зеленые водоросли, которые представляют собой режим фотосинтетической жизни, подобный тому, из которого, как полагают, произошли современные растения, благодаря наличию специализированных репродуктивных органов, защищенных нерепродуктивными тканями.
Мохообразные впервые появились в период раннего Палеозой. В основном они живут в местах обитания, где влага доступна в течение значительных периодов времени, хотя некоторые виды, такие как Targionia, устойчивы к высыханию. Большинство видов мохообразных остаются мелкими на протяжении всего жизненного цикла. Это предполагает чередование двух поколений: гаплоидный этап, названный гаметофит, а диплоид этап, названный спорофит. У мохообразных спорофит всегда неразветвленный и остается питательно зависимым от своего родительского гаметофита. Эмбриофиты обладают способностью секретировать кутикула на их внешней поверхности - восковой слой, устойчивый к высыханию. в мхи и роголистник кутикула обычно образуется только на спорофите. Устьица отсутствуют у печеночников, но встречаются на спорангиях мхов и роголистников, обеспечивая газообмен.
Сосудистые растения впервые появились в Силурийский период, и Девонский были разнообразными и распространились во многих различных земных средах. Они разработали ряд приспособлений, которые позволили им распространиться во все более засушливых местах, особенно в тканях сосудов. ксилема и флоэма, которые транспортируют воду и пищу по всему организму. Корневые системы, способные получать почвенную воду и питательные вещества, также развивались в течение девона. У современных сосудистых растений спорофиты, как правило, крупные, разветвленные, питательно независимые и долгоживущие, но появляется все больше свидетельств того, что палеозойские гаметофиты были такими же сложными, как и спорофиты. Гаметофиты всех групп сосудистых растений в ходе эволюции уменьшились в размерах и стали заметными в жизненном цикле.
У семенных растений микрогаметофит превращается из многоклеточного свободно живого организма в несколько клеток в пыльцевом зерне и миниатюрный мегагаметофит остается внутри мегаспорангия, прикрепляется к родительскому растению и зависит от него. Мегаспорангий, заключенный в защитный слой, называемый покровом, известен как яйцеклетка. После оплодотворения спермой, произведенной пыльца зерен, внутри семяпочки развивается зародыш спорофит. Покровы становятся семенной оболочкой, а семяпочка превращается в семя. Семенные растения могут выживать и воспроизводиться в чрезвычайно засушливых условиях, потому что они не зависят от свободной воды для движения сперматозоидов или развития свободноживущих гаметофитов.
Первые семенные растения, птеридоспермия (семенные папоротники), ныне вымершие, появились в девоне и распространились по каменноугольному периоду. Они были предками современных голосеменные, из которых четыре выжившие группы широко распространены сегодня, особенно хвойные породы, которые являются доминирующими деревья в нескольких биомы. Название голосеменное происходит от Греческий составное слово γυμνόσπερμος (γυμνός gymnos, «голый» и σπέρμα sperma, «семя»), поскольку семяпочки и последующие семена не заключены в защитную структуру (плодолистики или плод), а переносятся обнаженными, обычно на чешуях шишек.
Окаменелости

Эта секция нужны дополнительные цитаты для проверка. (Ноябрь 2018) (Узнайте, как и когда удалить этот шаблон сообщения) |
Растение окаменелости включают корни, древесину, листья, семена, фрукты, пыльца, споры, фитолиты, и янтарь (окаменелая смола, производимая некоторыми заводами). Ископаемые наземные растения встречаются в наземных, озерных, речных и прибрежных морских отложениях. Пыльца, споры и водоросли (динофлагелляты и акритархи ) используются для датировки толщ осадочных пород. Остатки ископаемых растений не так распространены, как ископаемые животные, хотя ископаемые остатки растений локально встречаются в изобилии во многих регионах мира.
Самые ранние окаменелости, которые явно можно отнести к Kingdom Plantae, - это ископаемые зеленые водоросли из Кембрийский. Эти окаменелости напоминают кальцинированный многоклеточный члены Dasycladales. Ранее Докембрийский Известны окаменелости, напоминающие одноклеточные зеленые водоросли, но окончательная идентичность с этой группой водорослей неясна.
Самые ранние окаменелости, приписываемые зеленым водорослям, датируются Докембрийский (около 1200 млн лет назад).[59][60] Прочные внешние стены празинофит цисты (известные как фикоматы) хорошо сохранились в ископаемых отложениях Палеозой (около 250–540 млн лет назад). Нитевидное ископаемое (Протероклад) из отложений среднего неопротерозоя (около 750 млн лет назад) относят к Кладофоралес, а самые старые достоверные записи Bryopsidales, Dasycladales ) и каменная кладка из Палеозой.[48][61]
Самые старые известные окаменелости эмбриофитов датируются Ордовик, хотя такие окаменелости фрагментарны. Посредством Силурийский, сохранились окаменелости целых растений, в том числе простого сосудистого растения Cooksonia в среднем силурии и в гораздо более крупных и сложных ликофит Барагванатия длиннолистная в позднем силуре. С раннего девона Рини черт, подробные окаменелости ликофитов и риниофиты были обнаружены детали отдельных клеток в органах растений и симбиотическая ассоциация этих растений с грибами порядка Glomales. В Девонский период также увидел эволюцию листьев и корней, и первое современное дерево, Археоптерис. Это дерево с папоротниковой листвой и стволом с хвойным деревом было гетероспористый производство спор двух разных размеров - ранний шаг в эволюции семян.[62]
В Угольные меры являются основным источником Палеозой окаменелости растений, и в настоящее время существует множество групп растений. Отвалы угольных шахт - лучшее место для сбора; каменный уголь сам по себе является остатками окаменелых растений, хотя структурные детали окаменелостей растений редко видны в угле. в Fossil Grove в Виктория Парк в Глазго, Шотландия, пни Лепидодендрон деревья находятся в исходных положениях роста.
Окаменелые останки хвойное дерево и покрытосеменные корни, стебли и ветви местами может быть много в озере и на берегу осадочные породы от Мезозойский и Кайнозойский эпох. Секвойя и его союзники, магнолия, дуб, и ладони встречаются часто.
Окаменелое дерево распространен в некоторых частях мира и чаще всего встречается в засушливых или пустынных районах, где он более подвержен воздействию эрозия. Окаменелая древесина часто сильно окремненный (в органический материал заменен на диоксид кремния ), а пропитанная ткань часто сохраняется в мельчайших деталях. Такие образцы можно вырезать и полировать с помощью гранильный оборудование. Ископаемые леса из окаменелого дерева были найдены на всех континентах.
Окаменелости семенных папоротников, таких как Глоссоптерис широко распространены на нескольких континентах Южное полушарие, факт, который поддержал Альфред Вегенер ранние идеи относительно Континентальный дрифт теория.
Структура, рост и развитие

Большая часть твердого материала в растении поступает из атмосферы. В процессе фотосинтез, большинство растений используют энергию в Солнечный лучик преобразовать углекислый газ от атмосферы, плюс воды, в простой сахара. Затем эти сахара используются в качестве строительных блоков и образуют основной структурный компонент растения. Хлорофилл, зеленого цвета, магний -содержащий пигмент имеет важное значение для этого процесса; он обычно присутствует в растении уходит, а часто и в других частях растения. Паразитические растения, с другой стороны, используют ресурсы своего хозяина для обеспечения материалов, необходимых для обмена веществ и роста.
Растения обычно полагаются на почву в первую очередь как на опору и воду (в количественном выражении), но они также получают соединения из азот, фосфор, калий, магний и другие элементарные питательные вещества из почвы. Эпифитный и литофит растения зависят от воздуха и ближайшего мусора в качестве питательных веществ, и хищные растения дополняют свои потребности в питательных веществах, особенно в азоте и фосфоре, добычей насекомых, которых они ловят. Для успешного роста большинства растений им также требуется кислород в атмосфере и вокруг своих корней (почвенный газ ) за дыхание. Растения используют кислород и глюкоза (которые могут быть произведены из хранящихся крахмал ) для обеспечения энергией.[63] Некоторые растения растут как подводные водные растения, используя кислород, растворенный в окружающей воде, и несколько специализированных сосудистых растений, таких как мангровые заросли и тростник (Phragmites australis ),[64] могут расти своими корнями в аноксический условия.
Факторы, влияющие на рост
Геном растения контролирует его рост. Например, отобранные сорта или генотипы пшеницы растут быстро, созревая в течение 110 дней, тогда как другие, в тех же условиях окружающей среды, растут медленнее и созревают в течение 155 дней.[65]
Рост также определяется факторами окружающей среды, такими как температура, имеется в наличии воды, имеется в наличии свет, углекислый газ и доступно питательные вещества в почве. Любое изменение доступности этих внешних условий отразится на росте растения и сроках его развития.[нужна цитата ]
Биотические факторы также влияют на рост растений. Растения могут быть настолько переполнены, что ни одна особь не дает нормального роста, вызывая этиоляция и хлороз. Оптимальному росту растений могут препятствовать пасущиеся животные, неоптимальный состав почвы, отсутствие микоризный грибки и нападения насекомых или болезни растений, в том числе вызванные бактериями, грибами, вирусами и нематодами.[65]

Простые растения, такие как водоросли, могут иметь короткую продолжительность жизни по отдельности, но их популяция обычно носит сезонный характер. Однолетние растения расти и воспроизводиться в одном сезон созревания, двулетние растения растут в течение двух вегетационных сезонов и обычно размножаются на второй год, и многолетние растения живут в течение многих вегетационных сезонов и после созревания часто воспроизводят ежегодно. Эти обозначения часто зависят от климата и других факторов окружающей среды. Растения однолетние в альпийский или умеренный регионы могут быть двухлетними или многолетними в более теплом климате. Среди сосудистых растений к многолетникам относятся как вечнозеленые растения которые хранят свои листья круглый год, и лиственный растения, которые частично теряют листья. В умеренном и бореальный климат, они обычно теряют листья зимой; много тропический растения теряют листья во время сухой сезон.[нужна цитата ]
Скорость роста растений крайне изменчива. Некоторые виды мхов растут менее 0,001 миллиметра в час (мм / ч), тогда как большинство деревьев вырастают 0,025–0,250 мм / ч. Некоторые виды альпинистов, например кудзу, которые не требуют образования толстой поддерживающей ткани, могут расти до 12,5 мм / ч.[нужна цитата ]
Растения защищаются от мороз и обезвоживание стресс с антифризы протеины, белки теплового шока и сахара (сахароза обычное дело). LEA (Обильный поздний эмбриогенез ) экспрессия белка индуцируется стрессом и защищает другие белки от агрегации в результате высыхание и замораживание.[66]
Эффекты замораживания
Последствия замерзания воды для растений во многом зависят от того, происходит ли замерзание внутри клеток (внутриклеточно) или вне клеток в межклеточных пространствах.[67] Внутриклеточное замораживание, которое обычно приводит к гибели клетки[68] независимо от морозостойкости растения и его тканей, редко встречается в природе, потому что скорость охлаждения редко бывает достаточно высокой, чтобы поддерживать его. Чтобы вызвать внутриклеточное образование льда, обычно требуется скорость охлаждения в несколько градусов Цельсия в минуту.[69] При скорости охлаждения несколько градусов Цельсия в час в межклеточных пространствах происходит расслоение льда.[70] Это может привести или не привести к летальному исходу, в зависимости от прочности ткани. При отрицательных температурах вода в межклеточных пространствах растительной ткани сначала замерзает, хотя вода может оставаться незамерзшей до тех пор, пока температура не упадет ниже -7 ° C (19 ° F).[67] После начального образования межклеточного льда клетки сжимаются, поскольку вода теряется в сегрегированном льду, и клетки подвергаются сублимационной сушке. Это обезвоживание считается основной причиной обморожения.
Повреждение и восстановление ДНК
Растения постоянно подвергаются целому ряду биотических и абиотических стрессов. Эти стрессы часто вызывают Повреждение ДНК прямо или косвенно через создание активные формы кислорода.[71] Растения способны к ответу на повреждение ДНК, что является критическим механизмом для поддержания стабильности генома.[72] Ответ на повреждение ДНК особенно важен во время семя прорастание, поскольку качество семян имеет тенденцию к ухудшению с возрастом в связи с накоплением повреждений ДНК.[73] Во время прорастания активируются процессы восстановления, чтобы справиться с этим накопленным повреждением ДНК.[74] В частности, одно- и двухцепочечные разрывы в ДНК могут быть отремонтирован.[75] Киназа контрольной точки ДНК Банкомат играет ключевую роль в интеграции прорастания и репарации на повреждения ДНК, накопленные старыми семенами.[76]
Клетки растений
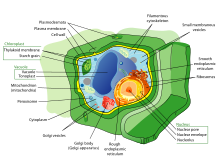
Растительные клетки обычно отличаются большими заполненными водой центральными ячейками. вакуоль, хлоропласты, и жесткий клеточные стенки которые состоят из целлюлоза, гемицеллюлоза, и пектин. Деление клеток также характеризуется развитием фрагмопласт для строительства клеточная пластина на поздних стадиях цитокинез. Так же, как и у животных, растительные клетки дифференцируются и развиваются в разные типы клеток. Тотипотент меристематический клетки могут дифференцироваться в сосудистый, хранение, защитное (например, эпидермальный слой ), или же репродуктивный ткани, у более примитивных растений отсутствуют некоторые типы тканей.[77]
Физиология
Фотосинтез
Растения фотосинтетический, что означает, что они производят собственные молекулы пищи, используя энергию, полученную из свет. Основным механизмом улавливания световой энергии у растений является пигмент хлорофилл. Все зеленые растения содержат две формы хлорофилла, хлорофилл а и хлорофилл б. Последний из этих пигментов не содержится в красных или коричневых водорослях. Простое уравнение фотосинтеза выглядит следующим образом:
![{displaystyle {ce {6CO2 {} + 6H2O {} -> [{ext {light}}] C6H12O6 {} + 6O2 {}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/100302228047a00799cc68db892940dd5e3adc9e)
Иммунная система
С помощью клеток, которые ведут себя как нервы, растения получают и распределяют в своих системах информацию об интенсивности и качестве падающего света. Падающий свет, который стимулирует химическую реакцию в одном листе, вызовет цепную реакцию сигналов для всего растения через тип клетки, называемый клеткой. связка оболочки клетки. Исследователи из Варшавский университет естественных наук в Польше обнаружили, что растения обладают особой памятью на различные условия освещения, что подготавливает их иммунную систему к сезонным патогенам.[78] Растения используют рецепторы распознавания образов для распознавания консервативных микробных сигнатур. Это распознавание вызывает иммунный ответ. Первые растительные рецепторы консервативных микробных сигнатур были идентифицированы у риса (XA21, 1995).[79] И в Arabidopsis thaliana (FLS2, 2000).[80] Растения также несут иммунные рецепторы, распознающие очень вариабельные эффекторы патогенов. К ним относятся белки класса NBS-LRR.
Внутреннее распространение
Сосудистые растения отличаются от других растений тем, что питательные вещества переносятся между своими частями через специализированные структуры, называемые ксилема и флоэма. У них также есть корни для впитывания воды и минералов. Ксилема перемещает воду и минералы от корня к остальным частям растения, а флоэма обеспечивает корни сахаром и другими питательными веществами, производимыми листьями.[77]
Геномика
Растения имеют одни из самых больших геномы среди всех организмов.[81] Самый крупный геном растения (по количеству генов) - это геном пшеница (Triticum asestivum), который, по прогнозам, кодирует ≈94000 генов[82] и, таким образом, почти в 5 раз больше, чем человеческий геном. Первым секвенированным геномом растения был геном Arabidopsis thaliana который кодирует около 25 500 генов.[83] С точки зрения чистой последовательности ДНК, наименьший из опубликованных геномов - это геном плотоядных животных. пузырчатка (Utricularia gibba) на 82 Мб (хотя кодирует еще 28500 генов)[84] а самый крупный из Ель обыкновенная (Picea abies), занимает 19 600 мегабайт (кодирует около 28 300 генов).[85]
Экология
Фотосинтез, осуществляемый наземными растениями и водорослями, является основным источником энергии и органического материала почти во всех экосистемах. Фотосинтез, сначала цианобактериями, а затем фотосинтезирующими эукариотами, радикально изменил состав бескислородной атмосферы ранней Земли, которая в результате теперь составляет 21%. кислород. Животные и большинство других организмов аэробный, полагаясь на кислород; те, которые этого не делают, ограничиваются относительно редкими анаэробная среда. Растения - это первичные производители в большинстве наземных экосистем и составляют основу пищевой сети в этих экосистемах. Многие животные полагаются на растения как на укрытие, а также как кислород и пищу.[нужна цитата ] Растения составляют около 80% мира биомасса около 450 гигатонн (4,4×1011 длинные тонны; 5.0×1011 короткие тонны) углерода.[86]
Наземные растения - ключевые компоненты круговорот воды и несколько других биогеохимические циклы. Некоторые растения имеют совместно развитый с фиксация азота бактерии, что делает растения важной частью азотный цикл. Корни растений играют важную роль в почва развитие и предотвращение эрозия почвы.[нужна цитата ]
Распределение
Растения распространены практически по всему миру. Пока они населяют множество биомы и экорегионы, немногие можно найти за пределами тундры в самых северных регионах континентальные шельфы. На южных крайностях растения Антарктическая флора Наличие стойко адаптированы к условиям.[нужна цитата ]
Растения часто являются доминирующим физическим и структурным компонентом местообитаний, в которых они встречаются. Многие из земных биомы названы в честь типа растительности, потому что растения являются доминирующими организмами в этих биомах, например луга, тайга и тропический лес.[нужна цитата ]
Экологические отношения

Эта секция нужны дополнительные цитаты для проверка. (Ноябрь 2018) (Узнайте, как и когда удалить этот шаблон сообщения) |
Многие животные эволюционировали одновременно с растениями. Многие животные опылять цветы в обмен на пищу в виде пыльцы или нектар. Многие животные рассыпать семена, часто во время еды фрукты и передавая семена в их кал. Мирмекофиты растения, которые эволюционировали вместе с муравьи. Растение обеспечивает дом, а иногда и пищу для муравьев. Взамен муравьи защищают растение от травоядные животные а иногда и конкурирующие растения. Муравьиные отходы обеспечивают органическое удобрение.
Большинство видов растений имеют различные виды грибов, связанных с их корневой системой в виде мутуалистический симбиоз известный как микориза. Грибы помогают растениям получать воду и минеральные питательные вещества из почвы, в то время как растение дает грибам углеводы, вырабатываемые в процессе фотосинтеза. Некоторые растения служат домом для эндофитный грибы, которые защищают растение от травоядных, вырабатывая токсины. Грибковый эндофит, Neotyphodium coenophialum, в овсяница высокая (Festuca arundinacea) наносит огромный экономический ущерб животноводству в США.
Среди растений также довольно распространены различные формы паразитизма, начиная с полупаразитарных. омела который просто забирает некоторые питательные вещества от своего хозяина, но все еще имеет фотосинтезирующие листья, полностью паразитирующим заразиха и зубочистка которые получают все свои питательные вещества через связи с корнями других растений и поэтому не имеют хлорофилл. Некоторые растения, известные как микогетеротрофы, паразитируют на микоризных грибах и, следовательно, действуют как эпипаразиты на других растениях.
Многие растения эпифиты, то есть они растут на других растениях, обычно деревьях, не паразитируя на них. Эпифиты могут косвенно нанести вред своему растению-хозяину, улавливая минеральные питательные вещества и свет, которые иначе получил бы хозяин. Вес большого количества эпифитов может сломать ветви деревьев. Гемиепифиты словно душитель фиг начинаются как эпифиты, но в конечном итоге пустили корни, одолели и убили своего хозяина. Много орхидеи, бромелии, папоротники и мхи часто растут как эпифиты. Эпифиты бромелий накапливают воду в пазухах листьев, образуя фитотельматы которые могут содержать сложные водные пищевые сети.[87]
Около 630 заводов плотоядный, такой как Венерина мухоловка (Dionaea muscipula) и росянка (Drosera виды). Они ловят мелких животных и переваривают их, чтобы получить минеральные питательные вещества, особенно азот и фосфор.[88]
Важность
Изучение использования растений людьми называется экономической ботаникой или этноботаника.[89] Выращивание растений человеком является частью сельское хозяйство, которая является основой человеческой цивилизации.[90] Растениеводство подразделяется на: агрономия, садоводство и лесное хозяйство.[91]
Еда

Люди зависят от растений в еда, либо напрямую, либо в качестве корма для домашние животные. сельское хозяйство занимается производством продовольственных культур и имеет сыграли ключевую роль в истории мировых цивилизаций. Сельское хозяйство включает агрономия для сельскохозяйственных культур, садоводство для овощей и фруктов, и лесное хозяйство для пиломатериалов.[92] В пищу использовалось около 7000 видов растений, хотя большая часть сегодняшней пищи получена только из 30 видов. Основным скобы включают хлопья такие как рис и пшеница, крахмалистые корни и клубни, такие как маниока и картофель, и бобовые такие как горох и бобы. Растительные масла такие как оливковое масло предоставлять липиды, в то время как фрукты и овощи делать вклад витамины и минералы к диете.[93]
Лекарства
Лекарственные растения являются основным источником органические соединения, как по их лечебным и физиологическим эффектам, так и по промышленным синтез из огромного количества органических химикатов.[94] Многие сотни лекарств получают из растений, причем оба традиционных лекарства используются в травничество[95][96] и химические вещества, очищенные от растений или впервые идентифицированные в них, иногда этноботанический поиск, а затем синтезированный для использования в современной медицине. Современные лекарства, полученные из растений, включают: аспирин, таксол, морфий, хинин, Резерпин, колхицин, дигиталис и винкристин. Растения, используемые в траволечении включают гинкго, эхинацея, пиретрум, и Зверобой. В фармакопея из Диоскорид, De Materia Medica, описывающий около 600 лекарственных растений, был написан между 50 и 70 годами нашей эры и использовался в Европе и на Ближнем Востоке примерно до 1600 года нашей эры; он был предшественником всех современных фармакопей.[97][98][99]
Непродовольственные товары

Растения, выращенные как технические культуры являются источником широкого спектра продуктов, используемых в производстве, иногда столь интенсивно, что это может нанести вред окружающей среде.[100] Непищевые продукты включают эфирные масла, натуральные красители, пигменты, воски, смолы, дубильные вещества, алкалоиды, янтарь и пробка. Продукты, полученные из растений, включают мыло, шампуни, парфюмерию, косметику, краски, лаки, скипидар, каучук, латекс, смазочные материалы, линолеум, пластмассы, чернила и десны. Возобновляемые виды топлива на заводах включают: дрова, торф и другие биотопливо.[101][102] В ископаемое топливо каменный уголь, нефть и натуральный газ происходят из останков водных организмов, в том числе фитопланктон в геологическое время.[103]
Структурные ресурсы и волокна растений используются для строительства жилищ и производства одежды. Дерево используется не только для зданий, лодок и мебели, но и для небольших предметов, таких как музыкальные инструменты и спортивный инвентарь. Дерево измельченный сделать бумагу и картон.[104] Ткань часто делают из хлопок, лен, рами или синтетические волокна, такие как район и ацетат полученный из растений целлюлоза. Нить также используется для шитья ткани, в основном из хлопка.[105]
Эстетическое использование

Тысячи видов растений выращиваются в эстетических целях, а также для создания тени, изменения температуры, уменьшения ветра, уменьшения шума, обеспечения уединения и предотвращения эрозии почвы. Растения составляют основу индустрии туризма с доходом в несколько миллиардов долларов в год, включая поездки в исторические сады, национальные парки, тропические леса, леса с красочными осенними листьями и фестивалями, такими как Японии[106] и Фестивали цветения сакуры в Америке.[107]

Хотя некоторые сады засажены продовольственными культурами, многие из них высажены в эстетических, декоративных или природоохранных целях. Дендрарии и ботанические сады являются общедоступными коллекциями живых растений. В частных открытых садах используются газонные травы, тенистые деревья, декоративные деревья, кустарники, виноградные лозы, многолетние травянистые растения и клумбы. Сады могут выращивать растения в естественном состоянии или могут скульптурировать их рост, как с топиарий или шпалер. Садоводство это самый популярный вид досуга в США, работа с растениями или садоводческая терапия полезен для реабилитации людей с ограниченными возможностями.[нужна цитата ]
Растения также можно выращивать или держать в помещении как комнатные растения, или в специализированных зданиях, таких как теплицы которые предназначены для ухода за живыми растениями и их выращивания. Венерина мухоловка, чувствительное растение и воскрешение примеры растений, проданных как новинки. Существуют также формы искусства, специализирующиеся на расположении срезанных или живых растений, такие как бонсай, икебана, а также композиция из срезанных или засушенных цветов. Декоративные растения иногда меняли ход истории, как в тюльпомания.[108]
Архитектурные конструкции, напоминающие растения, появляются в столицах Древнеегипетский колонны, которые были вырезаны, чтобы напоминать Египетский белый лотос или папирус.[109] Изображения растений часто используются в живописи и фотографии, а также на текстиле, деньгах, марках, флагах и гербах.[нужна цитата ]
Научное и культурное использование

Основные биологические исследования часто проводились с растениями. В генетика, разведение растений гороха разрешено Грегор Мендель вывести основные законы, регулирующие наследование,[110] и изучение хромосомы в кукурузе разрешено Барбара МакКлинток чтобы продемонстрировать их связь с унаследованными чертами.[111] Завод Arabidopsis thaliana используется в лабораториях как модельный организм чтобы понять как гены контролировать рост и развитие структур растений.[112] НАСА предсказывает, что однажды космические станции или космические колонии будут полагаться на растения для жизнеобеспечение.[113]
Древние деревья почитаются, и многие из них известный. Годовые кольца сами по себе являются важным методом датировки в археологии и служат для записи климата прошлого.[нужна цитата ]
Растения занимают видное место в мифология, религия и литература.[114][115][116] Они используются как национальный и государственные гербы, в том числе государственные деревья и государственные цветы. Растения часто используются в качестве памятников, подарков и для обозначения особых случаев, таких как рождение, смерть, свадьбы и праздники. Композицию цветов можно использовать для отправки скрытых Сообщения.[нужна цитата ]
Отрицательные эффекты
Сорняки нежелательные растения, растущие в управляемых средах, таких как фермы, городские районы, сады, газоны, и парки. Люди распространили растения за пределы родных ареалов, и некоторые из этих завезенных растений стали инвазивный, нанося ущерб существующим экосистемам, вытесняя местные виды, и иногда становясь серьезными сорняками культивирования.[нужна цитата ]
Растения могут причинить вред животным, в том числе людям. Растения, производящие ветром пыльца вызывать аллергические реакции у людей, страдающих Сенная лихорадка. Большое разнообразие растений ядовитый. Токсальбумины яды растений смертельны для большинства млекопитающих и служат серьезным сдерживающим фактором для потребления. Некоторые растения вызывают раздражение кожи при прикосновении, например Ядовитый плющ. Некоторые растения содержат психотропный химикаты, которые извлекаются, проглатываются или копчены, в том числе никотин из табак, каннабиноиды из Каннабис сатива, кокаин из Erythroxylon coca и опиум из опийный мак. Курение наносит вред здоровью или даже смерть, в то время как некоторые лекарства также могут быть вредными или смертельными для людей.[117][118] Как нелегальные, так и легальные наркотики, полученные из растений, могут оказывать негативное влияние на экономику, влияя на производительность труда и затраты правоохранительных органов.[119][120]
Смотрите также
Рекомендации
- ^ Кавальер-Смит, Т. (1981). «Царства эукариотов: семь или девять?». Биосистемы. 14 (3–4): 461–481. Дои:10.1016/0303-2647(81)90050-2. PMID 7337818.
- ^ Lewis, L.A .; Маккорт, Р. (2004). «Зеленые водоросли и происхождение наземных растений». Американский журнал ботаники. 91 (10): 1535–1556. Дои:10.3732 / ajb.91.10.1535. PMID 21652308.
- ^ Кенрик, Пол; Крейн, Питер Р. (1997). Происхождение и ранняя диверсификация наземных растений: кладистическое исследование. Вашингтон, округ Колумбия: Smithsonian Institution Press. ISBN 978-1-56098-730-7.
- ^ Адл, С. и другие. (2005). «Новая классификация эукариот более высокого уровня с упором на таксономию простейших». Журнал микробиологии эукариот. 52 (5): 399–451. Дои:10.1111 / j.1550-7408.2005.00053.x. PMID 16248873. S2CID 8060916.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ «Количество видов, находящихся под угрозой исчезновения, по основным группам организмов (1996–2010 годы)» (PDF). Международный союз охраны природы. 11 марта 2010 г. В архиве (PDF) из оригинала 21 июля 2011 г.. Получено 27 апреля 2011.
- ^ Филд, C.B .; Behrenfeld, M.J .; Randerson, J.T .; Фальковски, П. (1998). «Первичная продукция биосферы: интеграция наземных и океанических компонентов». Наука. 281 (5374): 237–240. Bibcode:1998Научный ... 281..237F. Дои:10.1126 / science.281.5374.237. PMID 9657713. В архиве из оригинала 25 сентября 2018 г.. Получено 10 сентября 2018.
- ^ «растение [2] - определение из онлайн-словаря Merriam-Webster». В архиве из оригинала 19 сентября 2011 г.. Получено 25 марта 2009.
- ^ «растение (форма жизни) - Британская онлайн-энциклопедия». В архиве из оригинала 13 марта 2009 г.. Получено 25 марта 2009.
- ^ а б Уиттакер, Р. Х. (1969). «Новые концепции царств или организмов» (PDF). Наука. 163 (3863): 150–160. Bibcode:1969Sci ... 163..150W. CiteSeerX 10.1.1.403.5430. Дои:10.1126 / science.163.3863.150. PMID 5762760. Архивировано из оригинал (PDF) 17 ноября 2017 г.. Получено 4 ноября 2014.
- ^ Маргулис, L (1971). «Пять царств организмов Уиттекера: незначительные исправления, предложенные соображениями происхождения митоза». Эволюция. 25 (1): 242–245. Дои:10.2307/2406516. JSTOR 2406516. PMID 28562945.
- ^ Коупленд, Х.Ф. (1956). Классификация низших организмов. Пало-Альто: Pacific Books, стр. 6, [1] В архиве 14 октября 2014 г. Wayback Machine.
- ^ Кавальер-Смит, Т. (1981). «Царства эукариотов: семь или девять?». Биосистемы. 14 (3–4): 461–481. Дои:10.1016/0303-2647(81)90050-2. PMID 7337818.
- ^ Линней, К. (1751). Философия ботаника В архиве 23 июня 2016 г. Wayback Machine, 1-е изд., С. 37.
- ^ Геккель, Э. (1866). Generale Morphologie der Organismen. Берлин: Verlag von Georg Reimer. стр. т. 1: i – xxxii, 1–574, табл. I – II, т. 2: i – clx, 1–462, табл. I – VIII.
- ^ Геккель, Э. (1894). Die systematische Phylogenie В архиве 27 апреля 2016 г. Wayback Machine.
- ^ На основе Рогозин, И.Б .; Basu, M.K .; Чюрёш М. и Кунин Е.В.(2009), «Анализ редких геномных изменений не поддерживает филогению Униконт – Биконт и предполагает, что цианобактериальный симбиоз является точкой первичного облучения эукариот», Геномная биология и эволюция, 1: 99–113, Дои:10.1093 / gbe / evp011, ЧВК 2817406, PMID 20333181 и Беккер, Б. и Марин, Б. (2009), «Стрептофитные водоросли и происхождение эмбриофитов», Анналы ботаники, 103 (7): 999–1004, Дои:10.1093 / aob / mcp044, ЧВК 2707909, PMID 19273476; см. также немного другую кладограмму в Льюис, Луиза А. и Маккорт, Р.М. (2004), «Зеленые водоросли и происхождение наземных растений», Am. J. Bot., 91 (10): 1535–1556, Дои:10.3732 / ajb.91.10.1535, PMID 21652308
- ^ Парфри, Лаура Вегенер; Lahr, Daniel J.G .; Knoll, Andrew H .; Кац, Лаура А. (16 августа 2011 г.). «Оценка сроков ранней диверсификации эукариот с помощью мультигенных молекулярных часов». Труды Национальной академии наук. 108 (33): 13624–13629. Bibcode:2011ПНАС..10813624П. Дои:10.1073 / pnas.1110633108. ЧВК 3158185. PMID 21810989.
- ^ Дерелль, Ромен; Торруэлла, Гифре; Климеш, Владимир; Бринкманн, Хеннер; Ким, Ынсу; Влчек, Честмир; Ланг, Б. Франц; Элиаш, Марек (17 февраля 2015 г.). «Бактериальные белки определяют единственный корень эукариот». Труды Национальной академии наук. 112 (7): E693 – E699. Bibcode:2015PNAS..112E.693D. Дои:10.1073 / pnas.1420657112. ЧВК 4343179. PMID 25646484.
- ^ Джексон, Кристофер; Клейден, Сьюзен; Рейес-Прието, Адриан (1 января 2015 г.). "Glaucophyta: сине-зеленые растения в двух словах". Acta Societatis Botanicorum Poloniae. 84 (2): 149–165. Дои:10.5586 / asbp.2015.020.
- ^ а б Санчес-Баракальдо, Патрисия; Рэйвен, Джон А .; Пизани, Давиде; Knoll, Эндрю Х. (12 сентября 2017 г.). «Ранние фотосинтезирующие эукариоты населяли среды обитания с низкой соленостью». Труды Национальной академии наук. 114 (37): E7737 – E7745. Дои:10.1073 / pnas.1620089114. ЧВК 5603991. PMID 28808007.
- ^ а б Gitzendanner, Matthew A .; Soltis, Pamela S .; Wong, Gane K.-S .; Ruhfel, Brad R .; Солтис, Дуглас Э. (2018). «Пластидный филогеномный анализ зеленых растений: миллиард лет эволюционной истории». Американский журнал ботаники. 105 (3): 291–301. Дои:10.1002 / ajb2.1048. PMID 29603143.
- ^ Гаврилюк, Райан М. Р .; Тихоненков, Денис В .; Хехенбергер, Элизабет; Хусник, Филип; Мыльников, Александр П .; Килинг, Патрик Дж. (8 августа 2019 г.). «Нефотосинтезирующие хищники - сестра красных водорослей». Природа. 572 (7768): 240–243. Дои:10.1038 / s41586-019-1398-6. ISSN 1476-4687. PMID 31316212. S2CID 197542583.
- ^ Маргулис, Л. (1974). «Классификация пяти царств, происхождение и эволюция клеток». Возможности филогенетических подходов к обнаружению горизонтально переносимых генов. Эволюционная биология. 7. С. 45–78. Дои:10.1007/978-1-4615-6944-2_2. ISBN 978-1-4615-6946-6. ЧВК 1847511. PMID 17376230.
- ^ а б c d е Рэйвен, Питер Х .; Эверт, Рэй Ф .; Эйххорн, Сьюзен Э. (2005). Биология растений (7-е изд.). Нью-Йорк: W.H. Фримен и компания. ISBN 978-0-7167-1007-3.
- ^ Льюис, Луиза А. и Маккорт, Р.М. (2004), «Зеленые водоросли и происхождение наземных растений», Am. J. Bot., 91 (10): 1535–1556, Дои:10.3732 / ajb.91.10.1535, PMID 21652308
- ^ а б Беккер, Б. и Марин, Б. (2009), «Стрептофитные водоросли и происхождение эмбриофитов», Анналы ботаники, 103 (7): 999–1004, Дои:10.1093 / aob / mcp044, ЧВК 2707909, PMID 19273476
- ^ Гири, доктор медицины; Гири, Г. (2007). "Тип: браузер таксономии Chlorophyta". AlgaeBase версии 4.2 Всемирное электронное издание, Национальный университет Ирландии, Голуэй. В архиве из оригинала 18 июня 2009 г.. Получено 23 сентября 2007.
- ^ Дьякон, Дж. (2005). Грибковая биология. Вайли. ISBN 978-1-4051-3066-0. В архиве из оригинала от 3 июня 2016 г.. Получено 8 января 2016.
- ^ «Интернет-флора всех известных растений». Мировая Флора Онлайн. Получено 26 октября 2020.
- ^ Хасслер, Майкл. «Общее количество видов». Мировые растения. Контрольный список синонимов и распространение мировой флоры. Получено 26 октября 2020.
- ^ Van den Hoek, C .; Mann, D.G .; И Янс, Х. 1995 г. Водоросли: введение в психологию. С. 343, 350, 392, 413, 425, 439 и 448 (Кембридж: Издательство Кембриджского университета). ISBN 0-521-30419-9
- ^ Гайри, доктор медицины и Гири, Г. (2011), Основа водорослей: Chlorophyta, Всемирное электронное издание, Национальный университет Ирландии, Голуэй, в архиве из оригинала 13 сентября 2019 г., получено 26 июля 2011
- ^ Гайри, доктор медицины и Гири, Г. (2011), Основа водорослей: Charophyta, Всемирное электронное издание, Национальный университет Ирландии, Голуэй, в архиве из оригинала 13 сентября 2019 г., получено 26 июля 2011
- ^ Van den Hoek, C .; Mann, D.G .; И Янс, Х. 1995 г. Водоросли: введение в психологию. С. 457, 463 и 476. (Кембридж: Издательство Кембриджского университета). ISBN 0-521-30419-9
- ^ Crandall-Stotler, Barbara & Stotler, Raymond E., 2000. «Морфология и классификация Marchantiophyta». п. 21 год в А. Джонатан Шоу и Бернард Гоффине (редакторы), Биология мохообразных. (Кембридж: издательство Кембриджского университета). ISBN 0-521-66097-1
- ^ Шустер, Рудольф М., Hepaticae и Anthocerotae Северной Америки, том VI, стр. 712–713. (Чикаго: Филдский музей естественной истории, 1992). ISBN 0-914868-21-7.
- ^ Гоффине, Бернар; Уильям Р. Бак (2004). «Систематика Bryophyta (мхов): от молекул к пересмотренной классификации». Монографии по систематической ботанике. 98: 205–239.
- ^ Гиффорд, Эрнест М .; Фостер, Эдрианс С. (1988). Морфология и эволюция сосудистых растений (3-е изд.). Нью-Йорк: W.H. Фримен и компания. п. 358. ISBN 978-0-7167-1946-5.
- ^ Тейлор, Томас Н .; Тейлор, Эдит Л. (1993). Биология и эволюция ископаемых растений. Нью-Джерси: Прентис-Холл. п. 636. ISBN 978-0-13-651589-0.
- ^ Международный союз охраны природы и природных ресурсов, 2006 г. Красный список видов, находящихся под угрозой исчезновения МСОП: сводная статистика В архиве 27 июня 2014 г. Wayback Machine
- ^ «Самые древние окаменелости показывают эволюцию несосудистых растений в период от среднего до позднего ордовика (≈450–440 млн лет назад) на основе ископаемых спор». Переход растений на сушу В архиве 2 марта 2008 г. Wayback Machine
- ^ Strother, Paul K .; Баттисон, Лейла; Brasier, Martin D .; Веллман, Чарльз Х. (26 мая 2011 г.). «Самые ранние неморские эукариоты на Земле». Природа. 473 (7348): 505–509. Bibcode:2011Натура.473..505S. Дои:10.1038 / природа09943. PMID 21490597. S2CID 4418860.
- ^ Хархольт, Джеспер; Моэструп, Эйвинд; Ульвсков, Петр (1 февраля 2016 г.). "Почему растения с самого начала были наземными". Тенденции в растениеводстве. 21 (2): 96–101. Дои:10.1016 / j.tplants.2015.11.010. PMID 26706443.
- ^ Понсе-Толедо, Р. И .; Deschamps, P .; López-García, P .; Zivanovic, Y .; Benzerara, K .; Морейра, Д. (2017). «Пресноводная цианобактерия с ранним ветвлением, лежащая в основе пластид». Текущая биология. 27 (3): 386–391. Дои:10.1016 / j.cub.2016.11.056. ЧВК 5650054. PMID 28132810.
- ^ Rothwell, G.W .; Scheckler, S.E .; Гиллеспи, W.H. (1989). "Элькинсия ген. nov., позднедевонское голосеменное растение с куполообразными семяпочками ». Ботанический вестник. 150 (2): 170–189. Дои:10.1086/337763. JSTOR 2995234. S2CID 84303226.
- ^ Кенрик, Пол и Питер Р. Крейн. 1997 г. Происхождение и ранняя диверсификация наземных растений: кладистическое исследование. (Вашингтон, округ Колумбия, Smithsonian Institution Press.) ISBN 1-56098-730-8.
- ^ Smith Alan R .; Прайер, Кэтлин М .; Schuettpelz, E .; Korall, P .; Schneider, H .; Вольф, Пол Г. (2006). «Классификация сохранившихся папоротников» (PDF). Таксон. 55 (3): 705–731. Дои:10.2307/25065646. JSTOR 25065646. Архивировано из оригинал (PDF) 26 февраля 2008 г.
- ^ а б Leliaert, F .; Smith, D.R .; Moreau, H .; Herron, M.D .; Verbruggen, H .; Delwiche, C.F .; Де Клерк, О. (2012). «Филогения и молекулярная эволюция зеленых водорослей» (PDF). Критические обзоры в науках о растениях. 31: 1–46. Дои:10.1080/07352689.2011.615705. S2CID 17603352. Архивировано из оригинал (PDF) 26 июня 2015 г.
- ^ Лелиаерт, Фредерик; Verbruggen, Heroen; Зехман, Фредерик В. (2011). «В глубину: новые открытия в основе филогении зеленых растений». BioEssays. 33 (9): 683–692. Дои:10.1002 / bies.201100035. PMID 21744372. S2CID 40459076.
- ^ Силар, Филипп (2016), "Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes", HAL Архивы-обзоры: 1–462, в архиве из оригинала 13 мая 2016 г., получено 21 июля 2016
- ^ Михайлюк, Татьяна; Лукешова, Алена; Глейзер, Карин; Хольцингер, Андреас; Обвегезер, Сабрина; Ныпорко, Светлана; Фридл, Томас; Карстен, Ульф (2018). «Новые таксоны стрептофитных водорослей (Streptophyta) из наземных местообитаний выявлены с использованием интегративного подхода». Протист. 169 (3): 406–431. Дои:10.1016 / j.protis.2018.03.002. ISSN 1434-4610. ЧВК 6071840. PMID 29860113.
- ^ Новиков и Барабаш-Красни (2015). Современная систематика растений. Лига-Прес. п. 685. Дои:10.13140 / RG.2.1.4745.6164. ISBN 978-966-397-276-3.
- ^ Leebens-Mack, M .; Баркер, М .; Карпентер, Э .; Дейхолос, М.К .; Gitzendammer, M.A .; Graham, S.W .; Grosse, I .; Ли, Чжэн (2019). «Тысяча транскриптомов растений и филогеномика зеленых растений». Природа. 574 (7780): 679–685. Дои:10.1038 / s41586-019-1693-2. ЧВК 6872490. PMID 31645766.
- ^ Лян, Чжэ; и другие. (2019). «Геном и транскриптом Mesostigma viride обеспечивают понимание происхождения и эволюции Streptophyta». Передовая наука. 7 (1): 1901850. Дои:10.1002 / advs.201901850. ЧВК 6947507. PMID 31921561.
- ^ Ван, Сибо; и другие. (2020). «Геномы ранних расхождений стрептофитных водорослей проливают свет на процесс наземной реализации растений». Природа Растения. 6 (2): 95–106. Дои:10.1038 / s41477-019-0560-3. ЧВК 7027972. PMID 31844283.
- ^ Паттик, Марк; и другие. (2018). «Взаимосвязь наземных растений и природа эмбриофитов-предков». Текущая биология. 28 (5): 733–745. Дои:10.1016 / j.cub.2018.01.063. PMID 29456145.
- ^ Чжан, Цзянь; и другие. (2020). «Геном роголистника и ранняя эволюция наземных растений». Природа Растения. 6 (2): 107–118. Дои:10.1038 / s41477-019-0588-4. ЧВК 7027989. PMID 32042158.
- ^ Ли, Фэй Вэй; и другие. (2020). «Геномы Anthoceros проливают свет на происхождение наземных растений и уникальную биологию роголистников». Природа Растения. 6 (3): 259–272. Дои:10.1038 / s41477-020-0618-2. PMID 32170292.
- ^ Нолл, Эндрю H (2003). Жизнь на молодой планете: первые три миллиарда лет эволюции на Земле. Издательство Принстонского университета.
- ^ Таппан, H (1980). Палеобиология протистов растений. Фриман, Сан-Франциско.
- ^ Баттерфилд, Николас Дж .; Knoll, Andrew H .; Светт, Кин (1994). "Палеобиология неопротерозойской свиты Сванбергфьеллет, Шпицберген". Lethaia. 27 (1): 76. Дои:10.1111 / j.1502-3931.1994.tb01558.x.
- ^ Стюарт, Уилсон А .; Ротвелл, Гар В. (1993). Палеоботаника и эволюция растений (2-е изд.). Издательство Кембриджского университета. ISBN 978-0521382946.
- ^ Уилсон, Эдвард О.; и другие. (1973). Жизнь на Земле (Первое изд.). Стэмфорд, Коннектикут, Sinauer Associates. п.145. ISBN 978-0-87893-934-3.
- ^ Р.М.М., Кроуфорд (1982). «Физиологические реакции при наводнении». Энциклопедия физиологии растений. 12B: 453–477.
- ^ а б Роббинс, W.W .; Weier, T.E .; и другие., Ботаника: растениеводство, 3-е издание, Wiley International, Нью-Йорк, 1965.
- ^ Goyal, K .; Walton, L.J .; Туннаклифф, А. (2005). «Белки LEA предотвращают агрегацию белков из-за водного стресса». Биохимический журнал. 388 (Часть 1): 151–157. Дои:10.1042 / BJ20041931. ЧВК 1186703. PMID 15631617.
- ^ а б Glerum, C. 1985. Морозостойкость саженцев хвойных пород: принципы и применение. стр. 107–123 в Дурья, М. (Ред.). Ход работы: Оценка качества рассады: принципы, процедуры и прогностические возможности основных тестов. Workshop, октябрь 1984 г., Университет штата Орегон, For. Res. Lab., Corvallis OR.
- ^ Lyons, J.M .; Raison, J.K .; Степонкус, П. 1979. Растительная мембрана в ответ на низкую температуру: обзор. стр. 1–24 в Lyons, J.M .; Graham, D .; Raison, J.K. (Ред.). Низкотемпературный стресс у сельскохозяйственных культур. Academic Press, Нью-Йорк, штат Нью-Йорк.
- ^ Мазур, П. 1977. Роль внутриклеточного замораживания в гибели клеток, охлажденных со сверхоптимальной скоростью. Криобиология 14: 251–272.
- ^ Sakai, A .; Ларчер, В. (ред.) 1987. Морозостойкость растений. Спрингер-Верлаг, Нью-Йорк. 321 с.
- ^ Roldán-Arjona, T .; Ариза, Р.Р. (2009). «Ремонт и толерантность к окислительным повреждениям ДНК у растений». Мутационные исследования. 681 (2–3): 169–179. Дои:10.1016 / j.mrrev.2008.07.003. PMID 18707020.
- ^ Ёсияма, К. (2016). «SOG1: главный регулятор реакции на повреждение ДНК у растений». Гены и генетические системы. 90 (4): 209–216. Дои:10.1266 / ggs.15-00011. PMID 26617076.
- ^ Waterworth, W.M .; Bray, C.M .; Запад, CE (2015). «Важность защиты целостности генома для прорастания и долголетия семян». Журнал экспериментальной ботаники. 66 (12): 3549–3558. Дои:10.1093 / jxb / erv080. PMID 25750428.
- ^ Koppen, G .; Вершев, Л. (2001). «Щелочной одноклеточный гель-электрофорез / анализ комет: способ изучения восстановления ДНК в корешковых клетках прорастающих Vicia faba». Folia Biologica (Прага). 47 (2): 50–54. PMID 11321247.
- ^ Waterworth, W.M .; Маснави, G .; Bhardwaj, R.M .; Jiang, Q .; Bray, C.M .; Запад, C.E. (2010). «ДНК-лигаза растений - важный фактор, определяющий долголетие семян». Журнал растений. 63 (5): 848–860. Дои:10.1111 / j.1365-313X.2010.04285.x. PMID 20584150.
- ^ Waterworth, W.M .; Footitt, S .; Bray, C.M .; Finch-Savage, W.E .; Запад, CE (2016). «Киназа контрольной точки повреждения ДНК ATM регулирует прорастание и поддерживает стабильность генома в семенах». PNAS. 113 (34): 9647–9652. Дои:10.1073 / pnas.1608829113. ЧВК 5003248. PMID 27503884.
- ^ а б Кэмпбелл, Нил А .; Рис, Джейн Б. (2005). Биология (7-е изд.). Пирсон / Бенджамин Каммингс. ISBN 978-0-8053-7146-8.
- ^ Гилл, Виктория (14 июля 2010 г.). "Растения умеют думать и помнить'". Новости BBC. BBC. В архиве из оригинала 5 июня 2018 г.. Получено 20 июн 2018.
- ^ Песня, W.Y .; и другие. (1995). «Рецепторный киназоподобный белок, кодируемый геном устойчивости к болезни риса, XA21». Наука. 270 (5243): 1804–1806. Bibcode:1995Научный ... 270.1804S. Дои:10.1126 / science.270.5243.1804. PMID 8525370. S2CID 10548988. В архиве из оригинала 7 ноября 2018 г.. Получено 10 сентября 2018.
- ^ Gomez-Gomez, L .; и другие. (2000). «FLS2: киназа, подобная рецептору LRR, участвующая в восприятии бактериального элиситора флагеллина в Арабидопсис". Молекулярная клетка. 5 (6): 1003–1011. Дои:10.1016 / S1097-2765 (00) 80265-8. PMID 10911994.
- ^ Майкл, Тодд П .; Джексон, Скотт (1 июля 2013 г.). «Первые 50 геномов растений». Геном растений. 6 (2): 0. Дои:10.3835 / plantgenome2013.03.0001in.
- ^ Бренчли, Рэйчел; Спаннагл, Мануэль; Пфайфер, Матиас; Barker, Gary L.A .; Д'Амор, Розалинда; Аллен, Александра М .; Маккензи, Нил; Крамер, Мелисса; Керхорну, Арнау (29 ноября 2012 г.). «Анализ генома мягкой пшеницы с использованием полногеномного секвенирования дробовика». Природа. 491 (7426): 705–710. Bibcode:2012Натура 491..705Б. Дои:10.1038 / природа11650. ЧВК 3510651. PMID 23192148.
- ^ Инициатива по геному арабидопсиса (14 декабря 2000 г.). «Анализ последовательности генома цветкового растения Arabidopsis thaliana». Природа. 408 (6814): 796–815. Bibcode:2000Натура 408..796Т. Дои:10.1038/35048692. PMID 11130711.
- ^ Ибарра-Лаклетт, Энрике; Лайонс, Эрик; Эрнандес-Гусман, Густаво; Перес-Торрес, Клаудиа Анахи; Карретеро-Поле, Лоренцо; Чанг, Тянь-Хао; Лань, Тяньин; Уэлч, Андреанна Дж .; Хуарес, Мария Джасмин Абрахам (6 июня 2013 г.). «Архитектура и эволюция миниатюрного генома растения». Природа. 498 (7452): 94–98. Bibcode:2013Натура 498 ... 94I. Дои:10.1038 / природа12132. ЧВК 4972453. PMID 23665961.
- ^ Nystedt, Björn; Street, Nathaniel R .; Веттербом, Анна; Цукколо, Андреа; Линь, Яо-Чэн; Скофилд, Дуглас Дж .; Вецци, Франческо; Деломм, Николас; Джакомелло, Стефания (30 мая 2013 г.). «Последовательность генома европейской ели и эволюция генома хвойных». Природа. 497 (7451): 579–584. Bibcode:2013Натура.497..579N. Дои:10.1038 / природа12211. PMID 23698360.
- ^ Бар-Он Ю.М., Филлипс Р., Майло Р. (июнь 2018 г.). «Распределение биомассы на Земле» (PDF). Труды Национальной академии наук Соединенных Штатов Америки. 115 (25): 6506–6511. Дои:10.1073 / pnas.1711842115. ЧВК 6016768. PMID 29784790.
- ^ Фрэнк, Ховард, Бромелиевые фитотельматы В архиве 20 августа 2009 г. Wayback Machine, Октябрь 2000 г.
- ^ Barthlott, W .; Порембски, С .; Seine, R .; Тайзен, И. 2007. Любопытный мир хищных растений: полное руководство по их биологии и выращиванию. Timber Press: Портленд, Орегон.
- ^ Кочхар, С. (31 мая 2016 г.). Экономическая ботаника: комплексное исследование. Издательство Кембриджского университета. ISBN 9781316675397.
- ^ Гаечный ключ, Джейсон С. (9 января 2013 г.). Коммуникация на рабочем месте в 21 веке: инструменты и стратегии, влияющие на чистую прибыль [2 тома]: инструменты и стратегии, оказывающие влияние на чистую прибыль. ABC-CLIO. ISBN 9780313396328.
- ^ Служба сельскохозяйственных исследований США (1903 г.). Отчет о сельскохозяйственных опытных станциях. Типография правительства США.
- ^ «Развитие сельского хозяйства». Национальная география. 2016. Архивировано с оригинал 14 апреля 2016 г.. Получено 1 октября 2017.
- ^ "Еда и напитки". Kew Gardens. Архивировано из оригинал 28 марта 2014 г.. Получено 1 октября 2017.
- ^ «Химические вещества из растений». Ботанический сад Кембриджского университета. Архивировано из оригинал 9 декабря 2017 г.. Получено 9 декабря 2017. Обратите внимание, что подробные сведения о каждом растении и производимых им химических веществах описаны на связанных подстраницах.
- ^ Tapsell, L.C .; Hemphill, I .; Cobiac, L. (август 2006 г.). «Польза трав и специй для здоровья: прошлое, настоящее, будущее». Med. Дж. Ост. 185 (4 Прил.): S4–24. Дои:10.5694 / j.1326-5377.2006.tb00548.x. PMID 17022438. S2CID 9769230.
- ^ Lai, P.K .; Рой, Дж .; Рой (июнь 2004 г.). «Противомикробные и химиопрофилактические свойства трав и специй». Curr. Med. Chem. 11 (11): 1451–1460. Дои:10.2174/0929867043365107. PMID 15180577.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ «Греческая медицина». Национальные институты здоровья США. 16 сентября 2002 г. В архиве из оригинала от 9 ноября 2013 г.. Получено 22 мая 2014.
- ^ Хефферон, Кэтлин (2012). Да будет пища твоя лекарством. Издательство Оксфордского университета. п. 46. ISBN 978-0199873982.
- ^ Руни, Энн (2009). История медицины. Издательство "Арктур". п. 143. ISBN 978-1848580398.
- ^ «Промышленное растениеводство». Фонд Благодати Коммуникаций. 2016 г. В архиве из оригинала 10 июня 2016 г.. Получено 20 июн 2016.
- ^ "Промышленные культуры и продукты Международный журнал ". Эльзевир. В архиве из оригинала 2 октября 2017 г.. Получено 20 июн 2016.
- ^ Cruz, Von Mark V .; Дириг, Дэвид А. (2014). Промышленные культуры: селекция для получения биоэнергии и биопродуктов. Springer. стр. 9 и пасс. ISBN 978-1-4939-1447-0. В архиве из оригинала 22 апреля 2017 г.. Получено 1 октября 2017.
- ^ Сато, Мотоаки (1990). Термохимия образования ископаемого топлива (PDF). Взаимодействие жидкости и минералов: дань уважения Х. П. Ойгстеру, специальная публикация № 2. Геохимическое общество. В архиве (PDF) из оригинала 20 сентября 2015 г.. Получено 1 октября 2017.
- ^ Сикста, Герберт, изд. (2006). Справочник по целлюлозе. 1. Винхейм, Германия: Wiley-VCH. п. 9. ISBN 978-3-527-30997-9.
- ^ «Натуральные волокна». Откройте для себя натуральные волокна. Архивировано из оригинал 20 июля 2016 г.. Получено 20 июн 2016.
- ^ Сосноски, Даниэль (1996). Введение в японскую культуру. Таттл. п.12. ISBN 978-0-8048-2056-1. Получено 13 декабря 2017.
- ^ "История сакуры и фестиваля". Национальный фестиваль цветения сакуры: О нас. Национальный фестиваль цветения сакуры. Архивировано из оригинал 14 марта 2016 г.. Получено 22 марта 2016.
- ^ Ламберт, Тим (2014). «Краткая история садоводства». BBC. В архиве из оригинала 9 июня 2016 г.. Получено 21 июн 2016.
- ^ Уилкинсон, Ричард Х. (2000). Полные храмы Древнего Египта. Темза и Гудзон. стр.65–66. ISBN 978-0-500-05100-9.
- ^ Блумберг, Роджер Б. «Бумага Менделя на английском языке». В архиве из оригинала 13 января 2016 г.. Получено 9 декабря 2017.
- ^ "Барбара МакКлинток: Краткий биографический очерк". WebCite. Архивировано из оригинал 21 августа 2011 г.. Получено 21 июн 2016.
- ^ «Об арабидопсисе». ТАИР. В архиве из оригинала 22 октября 2016 г.. Получено 21 июн 2016.
- ^ «Инженерная жизнь». НАСА. В архиве из оригинала от 8 июня 2016 г.. Получено 21 июн 2016.
- ^ Лейттен, Ребекка Роуз. «Мифы и легенды растений». Корнельский университет Либерти Консерватория Гайд-Бейли. Архивировано из оригинал 7 августа 2016 г.. Получено 20 июн 2016.
- ^ «Семь самых священных растений в мире». BBC. Получено 12 октября 2020.
- ^ «Литературные растения». Природа Растения. 1 (11): 15181. 2015. Дои:10.1038 / nplants.2015.181. PMID 27251545.
- ^ «кокаин / крэк». В архиве из оригинала 20 мая 2007 г.. Получено 25 мая 2007.
- ^ «Смерти, связанные с кокаином». В архиве из оригинала 17 июля 2006 г.. Получено 25 мая 2007.
- ^ «Незаконные наркотики отводят 160 миллиардов долларов в год из американской экономики». Архивировано из оригинал 15 февраля 2008 г.
- ^ «Социальная цена незаконного употребления наркотиков в Испании». Сентябрь 2002 г. В архиве из оригинала 2 октября 2007 г.. Получено 25 мая 2007.
дальнейшее чтение
- Общее
- Эванс, Л. (1998). Накормить десять миллиардов - растения и численность населения Рост. Издательство Кембриджского университета. Мягкая обложка, 247 стр. ISBN 0-521-64685-5.
- Кенрик, Пол и Крейн, Питер Р. (1997). Происхождение и ранняя диверсификация наземных растений: кладистическое исследование. Вашингтон, округ Колумбия: Smithsonian Institution Press. ISBN 1-56098-730-8.
- Рэйвен, Питер Х .; Эверт, Рэй Ф .; И Эйххорн, Сьюзан Э. (2005). Биология растений (7-е изд.). Нью-Йорк: W.H. Фримен и компания. ISBN 0-7167-1007-2.
- Тейлор, Томас Н. и Тейлор, Эдит Л. (1993). Биология и эволюция ископаемых растений. Энглвуд Клиффс, Нью-Джерси: Prentice Hall. ISBN 0-13-651589-4.
- Тревавас А (2003). «Аспекты интеллекта растений». Анналы ботаники. 92 (1): 1–20. Дои:10.1093 / aob / mcg101. ЧВК 4243628. PMID 12740212.
- Оценка и подсчет видов
- Комиссия по выживанию видов Международного союза охраны природы и природных ресурсов (МСОП) (2004 г.). Красный список МСОП [2].
- Пренс Г. (2001). «Открывая мир растений». Таксон. 50 (2, Золотой юбилей, часть 4): 345–359. Дои:10.2307/1223885. JSTOR 1223885.
внешняя ссылка
- Jones, T.M .; Reid, C.S .; Урбач, Л. «Визуальное исследование дивизионных растений». Цитировать журнал требует
| журнал =(Помогите) (требуется Microsoft Silverlight ) - Chaw, S.-M .; и другие. (1997). «Молекулярная филогения современных голосеменных растений и эволюция семенных растений: анализ ядерных последовательностей 18s рРНК» (PDF). Мол. Биол. Evol. 14 (1): 56–68. Дои:10.1093 / oxfordjournals.molbev.a025702. PMID 9000754. Архивировано из оригинал (PDF) 24 января 2005 г.
- Index Nominum Algarum
- Интерактивная классификация Кронквиста
- Растительные ресурсы Тропической Африки
- Древо жизни
- Ботанические и растительные базы данных
- База данных Африканской инициативы по растениям
- Австралия
- Чилийские заводы в Чилебоск
- e-Floras (Флора Китая, Флора Северной Америки и др.)
- Флора Europaea
- Флора Центральной Европы (на немецком)
- Флора Северной Америки
- Список японских диких растений онлайн
- Познакомьтесь с растениями - национальный тропический ботанический сад
- Центр полевых цветов леди Берд Джонсон - Информационная сеть по местным растениям Техасского университета в Остине
- Список растений
- Министерство сельского хозяйства США не ограничивается видами континентальной части США
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Растение группы | |||||||||||
| |||||||||||
| |||||||||||
| Рост и привычка растений | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Практика | |||||||||||
| |||||||||||
| |||||||||||
Элементы природа | |||||
|---|---|---|---|---|---|
| Вселенная | |||||
| земной шар | |||||
| Погода | |||||
| Окружающая среда | |||||
| Жизнь | |||||
| |||||