Хоанофлагеллята - Choanoflagellate
| Хоанофлагелляты Временной диапазон: 900–0 млн лет Окаменелости не известны, молекулярные часы свидетельствуют о происхождении 1050-800 млн лет назад.[1] | |
|---|---|
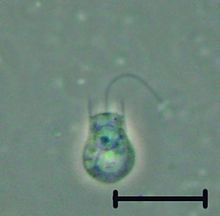 | |
| Codosiga sp. | |
| Научная классификация | |
| Домен: | Эукариоты |
| (без рейтинга): | Unikonta |
| (без рейтинга): | Obazoa |
| (без рейтинга): | Опистоконта |
| (без рейтинга): | Holozoa |
| (без рейтинга): | Филозоа |
| (без рейтинга): | Хоанозоа |
| Учебный класс: | Чоанофлагеллятея Кавалер-Смит, 1998 г.[2][3] |
| Типовой вид | |
| Monosiga brevicollis[4] | |
| Заказы и семьи | |
сестра: Metazoa | |
| Синонимы | |
| |
В хоанофлагелляты представляют собой группу свободноживущих одноклеточных и колониальных жгутик эукариоты считается ближайшими из ныне живущих родственников животные. Хоанофлагелляты - это жгутики с воротничком, имеющие воронкообразный воротник из соединенных между собой микроворсинки у основания жгутика. Хоанофлагелляты способны к бесполому и половому размножению.[7] У них отличительная ячейка морфология характеризуется яйцевидным или сферическим телом клеток 3–10мкм в диаметре с одним апикальным жгутиком, окруженным воротничком из 30-40 микроворсинок (см. рисунок). Движение жгутика создает водные потоки, которые могут продвигать свободно плавающие хоанофлагелляты через толщу воды и улавливать бактерии и детрит против воротника микроворсинок, в которые попадают эти продукты. Это кормление обеспечивает важную связь в глобальном цикл углерода, связывание трофические уровни. Помимо своей важной экологической роли, хоанофлагелляты представляют особый интерес для биологов-эволюционистов, изучающих происхождение многоклеточности у животных. Как ближайшие живущие родственники животных, хоанофлагелляты служат полезной моделью для реконструкций последнего одноклеточного предка животных.
Этимология
Хоанофлагеллята это гибридное слово от греческого χοάνη khoánē смысл "воронка"(из-за формы воротника) и латинского слова жгутик.
Внешность

Каждая хоанофлагеллята имеет один жгутик окруженный кольцом актин -заполненные выступы, называемые микроворсинками, образующие цилиндрический или конический воротник (хоано на греческом). Движение жгутика втягивает воду через воротник, а бактерии и детрит захватываются микроворсинками и проглатываются.[8] Водные потоки, создаваемые жгутик также толкайте свободно плавающие клетки, как в животное сперма. Напротив, большинство других жгутиконосцев являются потянул их жгутиками.
Помимо единственного апикального жгутика, окруженного актин -заполненный микроворсинки что характерно для хоанофлагеллят, внутренняя организация органеллы в цитоплазма постоянно.[9] Жгутик базальное тело сидит в основании апикальный жгутик и второй, не жгутиковый базальное тело лежит под прямым углом к основанию жгутика. В ядро занимает апикально-центральное положение в клетке, и пищевые вакуоли расположены в базальный регион цитоплазма.[9][10] Кроме того, клеточное тело многих хоанофлагеллят окружено характерным внеклеточный матрикс или же перипласт. Эти клеточные покрытия сильно различаются по структуре и составу и используются систематиками для целей классификации. Многие хоанофлагелляты строят сложные «домики» в форме корзины, называемые лорика из нескольких склеенных вместе полос диоксида кремния.[9] Функциональное значение перипласта неизвестно, но считается, что у сидячих организмов он способствует прикреплению к субстрату. У планктонных организмов есть предположение, что перипласт увеличивает сопротивление, тем самым противодействуя силе, создаваемой жгутиком, и повышая эффективность питания.[11]
Хоанофлагелляты либо свободно плавают в толще воды, либо сидячий, прилипая к субстрату напрямую или через перипласт или тонкую ножку.[12] Хотя считается, что хоанофлагелляты строго свободноживущие и гетеротрофный, ряд родственников хоанофлагеллат, таких как представители Ихтиоспорея или мезомицетозоа, следуйте паразитический или же патогенный Стиль жизни.[13] Истории жизни хоанофлагеллят изучены недостаточно. Многие виды считаются одиночными; однако колониальность, кажется, возникала независимо несколько раз внутри группы, и колониальные виды сохраняют одиночную стадию.[12]
Экология

Более 125 современных видов хоанофлагеллят[8] известны, распространены по всему миру в морской, солоноватый и пресная вода среды от Арктики до тропиков, занимая оба пелагический и бентосный зоны. Хотя большая часть проб хоанофлагеллят происходила между 0 м и 25 м, они были собраны с глубины 300 м в открытой воде.[14] и 100 м под ледяным покровом Антарктики.[15] Предполагается, что многие виды космополитичный в глобальном масштабе [например, Diaphanoeca grandis поступило сообщение из Северная Америка, Европа и Австралия (OBIS)], тогда как другие виды имеют ограниченное региональное распространение.[16] Совместно распространенные виды хоанофлагеллят могут занимать совершенно разные микроокружения, но в целом факторы, влияющие на распределение и дисперсию хоанофлагеллят, еще предстоит выяснить.
Номер разновидность, например, в род Протероспонгия, форма простая колонии,[8] планктонный Грозди, напоминающие миниатюрную гроздь винограда, в которой каждая клетка в колонии жгутиковата, или скопления клеток на одном стебле.[9][17] В октябре 2019 года ученые обнаружили новую полосу поведения хоанофлагеллят: они, по-видимому, могут координироваться, чтобы реагировать на свет.[18]
Питаются хоанофлагелляты бактерии и связать недоступные иным образом формы углерод к организмам, находящимся выше в трофической цепи.[19] Даже сегодня они важны в цикл углерода и микробный пищевой сети.[8] Есть некоторые свидетельства того, что хоанофлагелляты также питаются вирусами.[20]
Жизненный цикл

Хоанофлагелляты растут вегетативно, причем несколько видов подвергаются продольному делению;[10] однако репродуктивный жизненный цикл хоанофлагеллят еще предстоит выяснить. В документе, опубликованном в августе 2017 года, показано, что изменения окружающей среды, включая присутствие определенных бактерий, вызывают роение и последующее половое размножение хоанофлагеллят.[7] В плоидность уровень неизвестен;[22] однако открытие как ретротранспозонов, так и ключевых генов, участвующих в мейозе[23] ранее предполагалось, что они использовали половое размножение как часть их жизненного цикла. Некоторые хоанофлагелляты могут подвергаться инцистментации, которая включает в себя втягивание жгутика и воротника и заключение в плотную электронно-фибриллярную стенку. При переносе на свежую среду происходит эксцистмент; хотя это еще предстоит наблюдать.[24] Дальнейшее обследование хоанофлагеллат жизненный цикл будет информативно о механизмах образования колоний и признаках, присутствующих до эволюция многоклеточности животных.
Доказательства полового размножения были получены у видов хоанофлагеллят. Salpingoeca rosetta.[25][26] Также сообщалось о наличии консервативных мейотические гены в хоанофлагеллятах Monosiga brevicollis и Monosiga ovata.[27]
Биоминерализация кремния
Хоанофлагелляты акантоцидов образуют внеклеточную структуру корзины, известную как лорика. Лорика состоит из отдельных реберных полосок, изготовленных из кремнеземно-белкового биокомпозита. Каждая реберная полоска формируется внутри клетки хоанофлагеллат и затем секретируется на поверхность клетки. У нудиформных хоанофлагеллят сборка лорика происходит с использованием ряда щупалец после того, как будет произведено достаточное количество реберных полосок, чтобы составить полную лорику. У тектиформных хоанофлагеллят реберные полоски скапливаются в определенном порядке под воротником. Во время деления клетки новая клетка принимает эти реберные полоски как часть цитокинез и собирает свою собственную лорику, используя только эти ранее изготовленные ленты.[28]
Биосилицификация хоанофлагеллят требует концентрации кремниевая кислота внутри клетки. Это осуществляется белками Silicon Transporter (SIT). Анализ СИТ хоанофлагеллат показывает, что они аналогичны транспортерам кремния типа СИТ из диатомеи и другие кремнеземообразующие страменопилы. Семейство генов SIT демонстрирует небольшую или нулевую гомологию с любыми другими генами, даже с генами некремнистых хоанофлагеллят или страменопилов. Это предполагает, что семейство генов SIT эволюционировало посредством латерального переноса генов между акантоцидами и страменопилами. Это замечательный случай горизонтальный перенос генов между двумя отдаленно родственными эукариотическими группами и предоставил ключ к разгадке биохимии и кремний-белковых взаимодействий уникального семейства генов SIT.[29]
Классификация
Отношение к многоклеточным животным
Dujardin, французский биолог, интересующийся эволюцией простейших, зафиксировал морфологическое сходство хоанофлагеллят и хоаноцитов губок и предположил возможность близкого родства еще в 1841 году.[11] За последнее десятилетие эта предполагаемая связь между хоанофлагеллятами и животными была подтверждена независимым анализом нескольких несвязанных последовательностей: 18S рДНК, ядерных генов, кодирующих белок, и митохондриальных геномов (Steenkamp, et al., 2006; Burger, et al., 2003;[13] Уэйнрайт и др., 1993). Важно отметить, что сравнения последовательностей митохондриального генома хоанофлагелляты и трех губок подтверждают размещение хоанофлагеллят в качестве внешней группы для Metazoa и отрицают возможность того, что хоанофлагелляты произошли от многоклеточных (Lavrov, et al., 2005). Наконец, недавние исследования генов, экспрессируемых в хоанофлагеллятах, показали, что хоанофлагелляты синтезируют гомологи сигнальных и адгезионных генов клеток многоклеточных.[30] (King, 2003) Секвенирование генома показывает, что среди живых организмов хоанофлагелляты наиболее близки к животным.[8]Поскольку хоанофлагелляты и многоклеточные животные тесно связаны между собой, сравнения между двумя группами обещают дать представление о биологии их последнего общего предка и самых ранних событиях в мире. многоклеточный эволюция. В хоаноциты (также известные как "клетки с ошейником") губки (считаются одними из самых базальных многоклеточных) имеют такое же основное строение, что и хоанофлагелляты. Ошейниковые клетки встречаются у других групп животных, таких как ленточные черви,[31] предполагая, что это было морфология их последнего общего предка. Последний общий предок животных и хоанофлагеллят был одноклеточным, возможно, образующим простые колонии; Напротив, последний общий предок всех эуметазоа животные был многоклеточным организмом с дифференцированными тканями, определенным «планом тела» и эмбриональным развитием (включая гаструляцию).[8] Время разделения этих линий трудно определить, но, вероятно, это произошло в позднем докембрии,>600 миллион лет назад.[8]
Некоторые недавние статьи не подтверждают родственные отношения хоанофлагеллят с животными. Хоанофлагелляты могут быть связаны с Ихтиоспорея.[32]
Внешние связи Choanoflagellatea.[33]
| Опистоконта |
| ||||||||||||||||||||||||||||||||||||
Филогенетические отношения
Хоанофлагелляты включены в Chrysophyceae до Хибберда, 1975.[34] Недавние молекулярные филогенетический Реконструкция внутренних взаимоотношений хоанофлагеллят позволяет поляризовать эволюцию характера внутри клады. Крупные фрагменты ядерной СумГУ и LSU рибосомная РНК, альфа-тубулин, и белок теплового шока 90 кодирующих генов были использованы для выяснения внутренних отношений и полярности характера внутри хоанофлагеллят.[17] Каждый из четырех генов независимо показал сходные результаты, и анализ объединенного набора данных (соединенный ) вместе с последовательностями других близкородственных видов (животные и грибы ) демонстрируют, что хоанофлагелляты имеют сильную поддержку как монофилетический и подтверждают свою позицию как ближайшего из известных одноклеточных живых родственников животных.
Ранее Choanoflagellida были разделены на эти три семейства в зависимости от состава и структуры их перипласта: Codonosigidae, Salpingoecidae и Acanthoecidae. Представители семейства Codonosigidae при исследовании с помощью световой микроскопии не имеют перипласта, но могут иметь тонкую внешнюю оболочку, видимую только электронная микроскопия. Семейство Salpingoecidae состоит из видов, клетки которых заключены в твердую теку, которая видна как при световой, так и при электронной микроскопии. В CA секретное покрытие, состоящее преимущественно из целлюлоза или другой полисахариды.[35] Эти подразделения теперь известны как парафилетический, с широко распространенной конвергентной эволюцией этих форм. Третье семейство хоанофлагеллят, Acanthoecidae, считается монофилетической группой. Эта клада обладает синапоморфия клеток, обнаруженных в корзинообразной лорике, что дает альтернативное название «Loricate Choanoflagellates». Acanthoecid lorica состоит из ряда кремнистый реберные полосы расположены в виде узора лорики, характерного для конкретного вида ".[9][11]
Дерево хоанофлагеллат, основанное на молекулярной филогенетике, делится на три хорошо обоснованных клады.[17] Каждая клада 1 и клад 2 состоят из комбинации видов, традиционно относимых к Codonosigidae и Salpingoecidae, тогда как клад 3 включает виды из группы, таксономически классифицированной как Acanthoecidae.[17] Отображение черт характера на этой филогении указывает на то, что последний общий предок хоанофлагеллят был морским организмом с дифференцированным жизненный цикл с сидячий и подвижный этапы.[17]
Таксономия
Хоанофлагелляты[3]
- Заказ Craspedida Кавальер-Смит 1997 г. Nitsche et al. 2011 г.
- Семья Сальпингоэциды Кент 1880-1882 гг.
- ?Дикраспеделла Эллис 1930
- ?Diploeca Эллис 1930
- ?Диплозигопсис Франсе 1897
- ?Pachysoeca Эллис 1930
- ?Пиропсис Менье 1910
- ?Salpingorhiza Клуг 1936
- ?Сфаэродендрон Жуков, Мыльников и Моисеев 1976 non Seemann 1865
- ?Стексомонады Лэки 1942
- Астросига Кент 1880-1882 гг.
- Ауломонады Лэки 1942
- Choanoeca Эллис 1930
- Кладоспонгия Айенгар и Раматан 1940
- Кодоносигопсис Сенн 1900
- Codosiga Джеймс-Кларк 1866
- Диплосига Френцель 1891
- Desmarella Кент 1880-1882 гг.
- Hartaetosiga Карр, Рихтер и Ницше 2017
- Кентросига Шиллер 1953
- Моносига Кент 1880-1882 гг.
- Мыльносига Карр, Рихтер и Ницше 2017
- Lagenoeca Кент 1881
- Microstomoeca Карр, Рихтер и Ницше 2017
- Парамоносига Jeuck, Arndt & Nitsche 2014
- Протероспонгия Кент 1882
- Salpingoeca Джеймс-Кларк 1868 не Эллис 1933
- Sphaeroeca Лаутерборн 1894 не Мейрик 1895
- Stagondoeca Карр, Рихтер и Ницше 2017
- Стилохромонада Лэки 1940
- Семья Сальпингоэциды Кент 1880-1882 гг.
- Заказ Акантоцида Норрис 1965 г. Nitsche et al. 2011 г. (Лориатные хоанофлагелляты)
- Conioeca Томсен и Остергард 2019
- Семья Acanthoecidae Норрис 1965 г. Nitsche et al. 2011 г. (Нудиформные хоанофлагелляты)
- Acanthoeca Эллис 1930
- Энибас Schiwitza, Arndt & Nitsche 2019
- Helgoeca Ледбитер 2008
- Polyoeca Кент 1880
- Savillea Лёблих III 1967
- Семья Stephanoecidae Ледбитер 2011 (Тектиформные хоанофлагелляты)
- ?Conion Томсен 1982
- ?Spiraloecion Маршан и Перрин 1986
- Акантокорбис Хара и Такахаши 1984
- Амоэноскопа Хара и Такахаши 1987
- Афелоцион Томсен 1983
- Бикоста Ледбитер 1978
- Каллиаканта Ледбитер 1978
- Калотека Thomsen & Moestrup 1983 non Desv. 1810 non Spreng. 1817 не Гейден 1887
- Cosmoeca Томсен 1984
- Кринолина Томсен 1976 не Сметана 1982
- Crucispina Эспеланд и Трондсен 1986
- Diaphanoeca Эллис 1930
- Didymoeca Дауэлд 2003
- Kakoeca Бак и Марчант 1991
- Монокоста Томсен 1979 non Monocostus Schumann 1904
- Nannoeca Томсен 1988
- Parvicorbicula Дефландр 1960
- Pleurasiga Шиллер 1925
- Полифибула Мантон 1981
- Saepicula Ледбитер 1980
- Saroeca Томсен 1979
- Spinoeca Томсен, Остергард и Хансен, 1995 г., не Поульсен, 1973 г.
- Стефанаканта Томсен 1983
- Стефаноэка Эллис 1930
- Синдетофиллум Томсен и Моэструп, 1983 г.
- Томсенелла Ёздикмен 2009
Геномы и транскриптомы
Геномы двух видов хоанофлагеллят полностью секвенированы, а еще два вида имеют транскриптом данные опубликованы.
Monosiga brevicollis геном
Геном Monosiga brevicollis, с 41,6 миллионами пар оснований,[8] по размеру похож на мицелиальные грибы и другие свободноживущие одноклеточные эукариоты, но намного меньше, чем у типичных животных.[8] В 2010 году филогеномное исследование показало, что несколько генов водорослей присутствуют в геноме Monosiga brevicollis. Это могло быть связано с тем, что на раннем этапе эволюции хоанофлагелляты потребляли водоросли в пищу через фагоцитоз.[36] Carr et al. (2010)[27] просмотрел М. brevicollis геном для известных эукариотический гены мейоза. Из 19 известных тестированных эукариотических мейотических генов (включая 8, которые не функционируют ни в каком другом процессе, кроме мейоза), 18 были идентифицированы в М. brevicollis. Присутствие мейотических генов, включая гены, специфичные для мейоза, указывает на то, что мейоз и косвенно секс присутствует в составе хоанофлагеллят.
Salpingoeca rosetta геном
Геном Salpingoeca rosetta имеет размер 55 мегабаз.[37] В геноме присутствуют гомологи генов клеточной адгезии, метаболизма нейропептидов и гликосфинголипидов.С. розетта имеет сексуальный жизненный цикл и переходы между гаплоидная и диплоидная стадии.[26] В ответ на ограничение питательных веществ гаплоидные культуры С. розетта стать диплоидным. Этот сдвиг плоидности совпадает со спариванием, во время которого маленькие флагеллированные клетки сливаются с более крупными флагеллированными клетками. Есть также свидетельства исторического спаривания и рекомбинация в С. розетта.
S. rosetta побуждается к половому размножению морской бактерией Вибрио фишери.[25] Один В. фишери протеина, EroS полностью воспроизводит афродизиакоподобную активность живых V. fisheri.
Monosiga ovata транскриптом
Набор данных EST из Monosiga ovata вышла в 2006 году.[38] Основным открытием этого транскриптома стал домен Хоглета хоанофлагелляты, который пролил свет на роль перетасовки доменов в эволюции Сигнальный путь ежа. М. овата имеет по крайней мере четыре эукариотических мейотических гена.[27]
Стефаноэка диплокостата транскриптом
Первый транскриптом лориатной хоанофлагелляты[29] привело к открытию хоанофлагеллатных переносчиков кремния. Впоследствии подобные гены были идентифицированы у второго вида лориатников, Diaphanoeca grandis. Анализ этих генов показал, что SIT хоанофлагеллят проявляют гомологию с кремниевыми транспортерами SIT-типа диатомеи и развились через горизонтальный перенос генов.
Галерея

Salpingoeca под ПКМ

Salpingoeca sp. раздел под ТЕМ

Desmarella moniliformis колония под ПКМ

Codosiga колония под световая микроскопия

Sphaeroeca колония (около 230 особей) под световой микроскопией.






Рекомендации
- ^ Парфри Л.В., Лар DJ, Knoll AH, Кац Л.А. (август 2011 г.). «Оценка сроков ранней диверсификации эукариот с помощью мультигенных молекулярных часов». Труды Национальной академии наук Соединенных Штатов Америки. 108 (33): 13624–9. Bibcode:2011ПНАС..10813624П. Дои:10.1073 / pnas.1110633108. ЧВК 3158185. PMID 21810989.
- ^ Кавальер-Смит Т. (1998). «Неомонада и происхождение животных и грибов». В Coombs GH, Vickerman K, Sleigh MA, Warren A (ред.). Эволюционные отношения между простейшими. Лондон: Клувер. С. 375–407.
- ^ а б Ледбитер Б.С. (2015). Хоанофлагелляты: эволюция, биология и экология. Бирмингемский университет. ISBN 978-0-521-88444-0.
- ^ Кинг Н., Уэстбрук М.Дж., Янг С.Л., Куо А., Абедин М., Чепмен Дж. И др. (Февраль 2008 г.). "Геном хоанофлагелляты Monosiga brevicollis и происхождение многоклеточных". Природа. 451 (7180): 783–8. Bibcode:2008Натура.451..783K. Дои:10.1038 / природа06617. ЧВК 2562698. PMID 18273011.
- ^ Сэвилл-Кент, В. (1880). Руководство по инфузории. Лондон, т. 1, стр. 324, г. [1].
- ^ Ницше Ф, Карр М, Арндт Х, Ледбитер Б.С. (2011). «Таксономия высокого уровня и молекулярная филогенетика Choanoflagellatea». Журнал эукариотической микробиологии. 58 (5): 452–62. Дои:10.1111 / j.1550-7408.2011.00572.x. PMID 21895836.
- ^ а б «Бактериальный белок действует как афродизиак для хоанофлагеллят».
- ^ а б c d е ж грамм час я Король N, Уэстбрук М.Дж., Янг С.Л., Куо А., Абедин М., Чепмен Дж. И др. (Февраль 2008 г.). «Геном хоанофлагеллаты Monosiga brevicollis и происхождение многоклеточных животных». Природа. 451 (7180): 783–8. Bibcode:2008Натура.451..783K. Дои:10.1038 / природа06617. ЧВК 2562698. PMID 18273011.
- ^ а б c d е Ледбитер Б.С., Томсен Х. (2000). «Отряд Чоанофлагеллиды». Иллюстрированное руководство по простейшим, второе издание. Лоуренс: Общество протозоологов. 451: 14–38.
- ^ а б Карпов С., Ледбитер Б.С. (май 1998 г.). «Строение и состав цитоскелета хоанофлагеллят». Журнал эукариотической микробиологии. 45 (3): 361–367. Дои:10.1111 / j.1550-7408.1998.tb04550.x.
- ^ а б c Ледбитер Б.С., Келли М. (2001). «Эволюция животных хоанофлагеллят и губок». Вода и атмосфера онлайн. 9 (2): 9–11.
- ^ а б Ледбитер Б.С. (февраль 1983 г.). «История жизни и ультраструктура нового морского вида Протероспонгия (Choanoflagellida) ". J. Mar. Biol. Доц. ВЕЛИКОБРИТАНИЯ. 63 (1): 135–160. Дои:10.1017 / S0025315400049857.
- ^ а б Мендоса Л., Тейлор Дж. В., Аджелло Л. (2002). «Класс mesomycetozoea: гетерогенная группа микроорганизмов на границе между животными и грибами». Ежегодный обзор микробиологии. 56: 315–44. Дои:10.1146 / annurev.micro.56.012302.160950. PMID 12142489. S2CID 14764188.
- ^ Томсен Х (1982). Планктонные хоанофлагелляты из Диско-Бугт, Западная Гренландия, с обзором морского нанопланктона в этом районе. Meddelelser om Gronland, Bioscience. 8. С. 3–63. ISBN 978-87-635-1149-0.
- ^ Бак К.Р., Гарнизон DL (июнь 1988 г.). «Распространение и численность хоанофлагеллят (Acanthoecidae) в зоне кромки льда в море Уэдделла, Антарктида». Mar. Biol. 98 (2): 263–269. Дои:10.1007 / BF00391204.
- ^ Томсен Х, Бак К., Чавес Ф (1991). «Хоанофлагелляты центральных вод Калифорнии: систематика, морфология и видовые сообщества». Офелия. 33 (2): 131–164. Дои:10.1080/00785326.1991.10429736.
- ^ а б c d е Карр М., Ледбитер Б.С., Хассан Р., Нельсон М., Балдауф С.Л. (октябрь 2008 г.). «Молекулярная филогения хоанофлагеллят, сестринская группа Metazoa». Труды Национальной академии наук Соединенных Штатов Америки. 105 (43): 16641–6. Bibcode:2008PNAS..10516641C. Дои:10.1073 / pnas.0801667105. ЧВК 2575473. PMID 18922774.
- ^ «Недавно обнаруженные микроорганизмы объединяются», Flip Out'". HHMI.org. Получено 2019-10-29.
- ^ Баттерфилд, штат Нью-Джерси (1 апреля 1997 г.). «Экология планктона и переход от протерозоя к фанерозою». Палеобиология. 23 (2): 247–262. Дои:10.1017 / S009483730001681X.
- ^ Ву, Кэтрин Дж. (24 сентября 2020 г.). «Ничто не ест вирусы, верно? Познакомьтесь с голодными протистами: новые генетические данные подтверждают, что одноклеточные морские микробы могут поедать вирусы». Нью-Йорк Таймс. п. D3 (29 сентября 2020 г., распечатанная ред.). Получено 1 декабря, 2020.
- ^ Де Луф, А. и Скуфс, Л. (2019) «Механизм действия фарнезола,« благородного », неизвестного, в частности, в гомеостазе Са (2+), и его эфиры ювенильных гормонов в эволюционной ретроспективе». Границы неврологии, 13: 141. Дои:10.3389 / fnins.2019.00141.
- ^ Клаус Нильсен. Эволюция животных: взаимоотношения живых существ. 3-е изд. Клаус Нильсен. Оксфорд, Великобритания: Oxford University Press, 2012, стр. 14.
- ^ Карр М., Ледбитер Б.С., Балдауф С.Л. (2002). «Консервированные мейотические гены указывают на пол у хоанофлагеллят». Журнал эукариотической микробиологии. 57 (1): 56–62. Дои:10.1111 / j.1550-7408.2009.00450.x. PMID 20015185.
- ^ Ледбитер Б.С., Карпов С.А. (сентябрь – октябрь 2000 г.). «Формирование кист в пресноводном штамме хоанофлагелляты Desmarella moniliformis Kent». Журнал эукариотической микробиологии. 47 (5): 433–9. Дои:10.1111 / j.1550-7408.2000.tb00071.x. PMID 11001139.
- ^ а б Возница А., Гердт Дж. П., Хьюлетт Р. Э., Кларди Дж., Король Н. (сентябрь 2017 г.). «Спаривание у ближайших живых родственников животных индуцируется бактериальной хондроитиназой». Клетка. 170 (6): 1175–1183.e11. Дои:10.1016 / j.cell.2017.08.005. ЧВК 5599222. PMID 28867285.
- ^ а б Левин Т.С., Король Н. (ноябрь 2013 г.). «Доказательства пола и рекомбинации у хоанофлагелляты Salpingoeca rosetta». Текущая биология. 23 (21): 2176–80. Дои:10.1016 / j.cub.2013.08.061. ЧВК 3909816. PMID 24139741.
- ^ а б c Карр М., Ледбитер Б.С., Балдауф С.Л. (2010). «Консервированные мейотические гены указывают на пол у хоанофлагеллят». Журнал эукариотической микробиологии. 57 (1): 56–62. Дои:10.1111 / j.1550-7408.2009.00450.x. PMID 20015185.
- ^ Leadbeater BS, Yu Q, Kent J, Stekel DJ (январь 2009 г.). «Объемные изображения хоанофлагелляты loricae». Ход работы. Биологические науки. 276 (1654): 3–11. Дои:10.1098 / rspb.2008.0844. ЧВК 2581655. PMID 18755674.
- ^ а б Маррон А.О., Олстон М.Дж., Небеса Д., Акам М., Каккамо М., Голландия PW, Уокер G (апрель 2013 г.). «Семейство диатомоподобных переносчиков кремния в кремнистых лориатных хоанофлагеллятах». Ход работы. Биологические науки. 280 (1756): 20122543. Дои:10.1098 / rspb.2012.2543. ЧВК 3574361. PMID 23407828.
- ^ Король N, Кэрролл С.Б. (Декабрь 2001 г.). «Рецепторная тирозинкиназа из хоанофлагеллят: молекулярное понимание ранней эволюции животных». Труды Национальной академии наук Соединенных Штатов Америки. 98 (26): 15032–7. Bibcode:2001ПНАС ... 9815032К. Дои:10.1073 / pnas.261477698. ЧВК 64978. PMID 11752452.
- ^ Кантелл С., Францен А., Сенсенбо Т. (октябрь 1982 г.). «Ультраструктура многослойных воротниковых клеток личинки пилидиума Линия билинеатус (Немертини) ". Зооморфология. 101 (1): 1–15. Дои:10.1007 / BF00312027.
- ^ Корсаро Д., Валочник Дж., Вендитти Д., Мюллер К.Д., Хаурёдер Б., Мишель Р. (декабрь 2014 г.). «Повторное открытие Nucleophaga amoebae, нового члена Rozellomycota». Паразитологические исследования. 113 (12): 4491–8. Дои:10.1007 / s00436-014-4138-8. PMID 25258042.
- ^ Торруэлла, Гифре; Мендоса, Алекс де; Грау-Бове, Ксавье; Antó, Meritxell; Чаплин, Марк А .; Кампо, Хавьер дель; Эме, Лаура; Перес-Кордон, Грегорио; Уиппс, Кристофер М. (2015). «Филогеномика раскрывает конвергентную эволюцию образа жизни у близких родственников животных и грибов». Текущая биология. 25 (18): 2404–2410. Дои:10.1016 / j.cub.2015.07.053. PMID 26365255.
- ^ Ревьер, Б. де. (2006). Biologia e Filogenia das Algas. Editora Artmed, Порту-Алегри, стр. 156.
- ^ (Адл и др., 2005)
- ^ Сунь Г, Ян З, Ишвар А, Хуан Дж (декабрь 2010 г.). «Гены водорослей у ближайших родственников животных». Молекулярная биология и эволюция. 27 (12): 2879–89. Дои:10.1093 / molbev / msq175. PMID 20627874.
- ^ Fairclough SR, Chen Z, Kramer E, Zeng Q, Young S, Robertson HM, Begovic E, Richter DJ, Russ C, Westbrook MJ, Manning G, Lang BF, Haas B, Nusbaum C, King N (февраль 2013 г.). «Эволюция генома преметазоана и регуляция дифференцировки клеток хоанофлагелляты Salpingoeca rosetta». Геномная биология. 14 (2): R15. Дои:10.1186 / gb-2013-14-2-r15. ЧВК 4054682. PMID 23419129.
- ^ Снелл Е.А., Брук Н.М., Тейлор В.Р., Казан Д., Филипп Х., Голландия PW (Февраль 2006 г.). «Необычный белок хоанофлагеллят, выпущенный автокаталитической обработкой Hedgehog». Ход работы. Биологические науки. 273 (1585): 401–7. Дои:10.1098 / rspb.2005.3263. ЧВК 1560198. PMID 16615205.
внешняя ссылка
- ChoanoWiki совместный ресурс, поддерживаемый исследовательским сообществом Choanoflagellate
- Веб-страница "Древо жизни" для хоанофлагеллят
- Monosiga brevicollis браузер генома
- Химия вашего мозга существовала до того, как появились животные
- Choanobase, генетическая библиотека Choanoflagellate, разработанная и поддерживаемая Николь Кинг лаборатория в Калифорнийский университет в Беркли
