Амебозоа - Amoebozoa
| Амебозоа | |
|---|---|
 | |
| Хаос каролиненсис | |
| Научная классификация | |
| Домен: | Эукариоты |
| (без рейтинга): | Unikonta |
| Тип: | Амебозоа Люэ, 1913 исправить. Кавалер-Смит, 1998 |
| Подтипы, инфрафилы и классы | |
| Синонимы | |
| |
Амебозоа является основным таксономический группа, содержащая около 2400 описанных видов амебовидный протисты,[2] часто обладая тупыми, похожими на пальцы, лобоз ложноножки и трубчатые митохондриальный кристы.[3][4] В большинстве классификационных схем амебозоа классифицируются как филюм в рамках Королевство Протиста[5] или королевство Простейшие.[6] В классификации, одобренной Международным обществом протистологов, он сохраняется как не имеющий рейтинга "супергруппа "внутри эукариот.[3] Молекулярно-генетический анализ поддерживает амебозоа как монофилетический клады. Наиболее филогенетические деревья идентифицировать его как сестринскую группу, чтобы Опистоконта, еще одна крупная клада, которая содержит оба грибы и животные а также около 300 видов одноклеточных протистов.[2][4] Amoebozoa и Opisthokonta иногда объединяются в группу высокого уровня. таксон, по-разному названные Unikonta,[6] Аморфея[3] или же Opimoda.[7]
Amoebozoa включает в себя многие из самых известных амебоидных организмов, такие как Хаос, Entamoeba, Пеломикс и род Амеба сам. Виды Amoebozoa могут иметь панцирь (семенник) или голые, а клетки могут иметь жгутики. Свободноживущие виды распространены как в соленой, так и в пресной воде, а также в почве, мхе и опавших листьях. Некоторые живут как паразиты или же симбиоты других организмов, а некоторые, как известно, вызывают заболевания у людей и других организмов.
Хотя большинство видов амебозойных являются одноклеточными, группа также включает несколько кладов слизевые формы, которые имеют макроскопическую, многоклеточную стадию жизни, в течение которой отдельные амебоидные клетки остаются вместе после многократного деления клеток, чтобы сформировать макроскопический плазмодий, или, в клеточных слизистых плесневых грибах, объединяются, чтобы сформировать один.
Амебозоа сильно различаются по размеру. Некоторые из них имеют диаметр всего 10–20 мкм, в то время как другие относятся к числу самых крупных простейших. Известный вид Амеба протей, который может достигать 800 мкм в длину, часто изучается в школах и лабораториях как репрезентативная клетка или модельный организм отчасти из-за удобного размера. Многоядерный амебы как Хаос и Пеломикс может быть несколько миллиметров в длину, а некоторые многоклеточные амебозоиды, такие как слизистая плесень "собачья рвота" Fuligo septica, может занимать площадь в несколько квадратных метров.[8]
Морфология
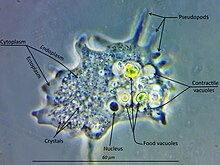
Амёбозоа - большая и разнообразная группа, но у многих ее членов есть общие черты. Клетка амебозойных обычно делится на гранулированную центральную массу, называемую эндоплазма и чистый внешний слой, называемый эктоплазмой. Во время передвижения эндоплазма течет вперед, а эктоплазма движется назад вдоль внешней стороны клетки. В движении многие амебозойные животные имеют четко выраженные переднюю и заднюю части и могут принимать «моноподиальную» форму, когда вся клетка функционирует как единый псевдопод. Большие псевдоподии могут давать многочисленные четкие выступы, называемые субпсевдоподиями (или детерминированными псевдоподиями), которые расширяются до определенной длины, а затем втягиваются либо для передвижения, либо для приема пищи. Клетка также может образовывать несколько неопределенных псевдоподий, через которые все содержимое клетки протекает в направлении движения. Они более или менее трубчатые и в основном заполнены зернистой эндоплазмой. Клеточная масса перетекает в ведущую ложноножку, а остальные в конечном итоге втягиваются, если организм не меняет направление.[9]
Хотя большинство амебозоидов «голые», как и знакомые Амеба и Хаос, или покрытые рыхлым слоем мелкой чешуи, например Cochliopodium и Коротневелла, члены ордена Арцеллинида образуют жесткие оболочки, или тесты, снабженный одним отверстием, через которое выходят ложноножки. Тесты на арцеллинид могут выделяться из органических материалов, как в Арчелла, или собранных из собранных частиц, склеенных вместе, как в Диффлугия.
У всех амебозоидов основной способ питания - фагоцитоз, в котором клетка окружает потенциальные частицы пищи своими псевдоножками, запечатывая их в вакуоли в котором они могут перевариваться и всасываться. У некоторых амебозоидов есть задняя луковица, называемая уроидом, которая может служить для накопления отходов, периодически отделяясь от остальной части клетки.[нужна цитата ] Когда пищи не хватает, большинство видов могут образовывать кисты, которые можно переносить по воздуху и знакомить с новыми условиями.[нужна цитата ] В плесневых грибах эти структуры называются спорами и образуются на стеблях, называемых плодовыми телами или спорангии.
У большинства амебозоа отсутствуют жгутики и вообще не образуют микротрубочка -опорные конструкции, кроме митоз. Однако жгутики действительно встречаются среди Архамебы, и многие формы слизи производят бифлагеллят гаметы[нужна цитата ]. Жгутик обычно закреплен конусом микротрубочек, что указывает на тесную связь с опистоконцы.[нужна цитата ] В митохондрии в клетках амебозойных животных характерно наличие ветвящихся трубчатых кристы. Однако среди Архамебы, которые адаптированы к бескислородной или микроаэрофильной среде обитания, митохондрии были потеряны.
Классификация
Место амебозоа в эукариотическом дереве
Похоже (на основании молекулярной генетики), что члены Amoebozoa образуют сестринская группа животным и грибам, отходящим от этой линии после того, как она отделилась от других групп,[10]как показано ниже на упрощенной схеме:
| Opimoda |
| ||||||||||||||||||||||||||||||||||||

Сильное сходство между Amoebozoa и Опистоконц приводят к гипотезе о том, что они образуют отдельную кладу.[11] Томас Кавалье-Смит предложил название «unikonts» (формально Unikonta) для этой ветви, члены которой, как полагали, произошли от общего предка, обладающего единым эмерджентным жгутиком, укорененным в одном. базальное тело.[1][2] Однако, хотя тесная связь между Amoebozoa и Opisthokonta убедительно подтверждается, недавняя работа показала, что гипотеза об одноцилиндровом предке, вероятно, неверна. В своей пересмотренной классификации эукариот (2012) Адл и др. предложил Amorphea как более подходящее название для клады примерно того же состава, сестринской группы Потогонные.[3] Более поздняя работа объединяет членов Amorphea с малавимониды и коллодиктиониды в предлагаемой кладе под названием Opimoda, которая включает одну из двух основных ветвей, расходящихся у корня эукариотического древа жизни.[7]
Подтипы амебозоа: лобоза и коноза
Традиционно все амебозои с лопастными ложноножками объединяли в класс. Lobosea, помещенный с другими амебоидами в тип Sarcodina или Ризоподы, но эти группы считались неестественными. Структурные и генетические исследования выявили перколозои и несколько архамеб как самостоятельные группы. В филогении на основе рРНК их представители были отделены от других амеб и, казалось, расходились у основания эукариотический эволюция, как и большинство слизистых форм.
Однако исправленные деревья Кавалер-Смит и Чао в 1996 г.[12] предположили, что оставшиеся лобозаны действительно образуют монофилетическую группу, к которой были тесно связаны архамебы и микетозоа, хотя перколозои не были. Впоследствии они изменили тип Amoebozoa, включив в него как подтип Lobosa, так и новый подтип. Коноса, включающий архамеб и Mycetozoa.[13]
Недавние молекулярно-генетические данные, по-видимому, подтверждают это первичное разделение амебозоа на лобоза и коноза.[4] Первые, по определению Кавалье-Смита и его сотрудников, в основном состоят из классических Lobosea: не жгутиковых амеб с тупыми лопастными псевдоножками (Амеба, Acanthamoeba, Arcella, Difflugia и др.). Последний состоит как из амебоидных, так и из жгутиковых клеток, для которых характерны более заостренные или слегка ветвящиеся субпсевдоподии (Archamoebae и микетозойские слизистые формы).
Филогения и таксономия амебозоа
Из более ранних исследований Кавальер-Смита, Чао и Льюиса, 2016 г.[14] и Silar 2016.[15] Также недавняя филогения указывает на то, что Lobosa парафилетичны: Conosa - сестра Cutosea.[16][17][18]
Тип Amoebozoa Lühe 1913 исправить. Кавалер-Смит 1998 [Амебобиота; Eumycetozoa Zopf 1884 исправить Olive 1975]
- Clade Дискоса Кавальер-Смит 2004 стат. ноя Adl et al. 2018 г.
- Заказ ?Стереомиксида Грелль 1971
- Заказ ?Стигамебида Смирнов и Кавальер-Смит 2011
- Учебный класс Центрамёбия Кавальер-Смит и др. 2016 г.
- Заказ Centramoebida Роджерсон и Паттерсон 2002 г. Кавалер-Смит 2004
- Заказ Himatismenida Стр. 1987 [Cochliopodiida]
- Заказ Пеллитида Стр. 1987 [Cochliopodiida]
- Учебный класс Flabellinia Смирнов и Кавальер-Смит 2011 г. Кудрявцев и др. 2014 г.
- Заказ Thecamoebida Schaeffer 1926 em. Смирнов и Кавальер-Смит 2011
- Заказ Dermamoebida Кавальер-Смит 2004 г. Смирнов и Кавальер-Смит 2011
- Заказ Ваннеллида Смирнов и др. 2005 г.
- Заказ Dactylopodida Смирнов и др. 2005 г.
- Clade Тевоса Канг и др. 2017 г.
- Clade Tubulinea Смирнов и др. Стат. За 2005 г. ноя Adl et al. 2018 г.
- Учебный класс Корицидия Канг и др. Стат. За 2017 г. ноя Adl et al. 2018 г.
- Заказ Трихозида Мебиус 1889
- Семья Microcoryciidae де Saedeleer 1934
- Учебный класс Эхинамобия Cavalier-Smith 2016 стат. ноя Adl et al. 2018 г.
- Заказ Эхинамебида Кавальер-Смит 2004 г. 2011 г.
- Учебный класс Элардия Канг и др. Стат. За 2017 г. ноя Adl et al. 2018 г.
- Подкласс Лептомиксия Кавалер-Смит 2016
- Заказ Лептомиксида Pussard & Pons 1976 г. Стр. 1987
- Подкласс Евлобозия Кавалер-Смит 2016
- Заказ Euamoebida Лепши 1960 г. Кавалер-Смит 2016
- Заказ Арцеллинида Кент 1880
- Подкласс Лептомиксия Кавалер-Смит 2016
- Учебный класс Корицидия Канг и др. Стат. За 2017 г. ноя Adl et al. 2018 г.
- Clade Evosea Канг и др. Стат. За 2017 г. ноя Adl et al. 2018 г.
- Clade Cutosa Cavalier-Smith 2016 стат. ноя
- Учебный класс Cutosea Кавалер-Смит 2016
- Заказ Сквамокутида Кавалер-Смит 2016
- Учебный класс Cutosea Кавалер-Смит 2016
- Подтип Коноса Кавальер-Смит 1998 стат. ноя
- Инфрафилум Архамебы Кавалер-Смит 1993 стат. п. 1998 г.
- Учебный класс Archamoebea Кавалер-Смит 1983 стат. п. 2004 г.
- Семья Tricholimacidae Кавалер-Смит 2013
- Семья Endamoebidae Калкинс 1926
- Заказ Entamoebida Кавалер-Смит 1993
- Заказ Пелобионтида Страница 1976 исправление. Кавалер Смит 1987
- Учебный класс Archamoebea Кавалер-Смит 1983 стат. п. 2004 г.
- Инфрафилум Семиконосия Кавалер-Смит 2013
- Учебный класс Variosea Кавальер-Смит и др. 2004 г.
- Заказ ?Flamellidae Кавалер-Смит 2016
- Заказ ?Holomastigida Лаутерборн 1895 [Artodiscida Кавалер-Смит 2013]
- Заказ Phalansteriida Хибберд 1983
- Заказ Рамамоебида Кавалер-Смит 2016
- Заказ Profiliida Канг и др. 2017 г. [Protosteliida Олив и Стоянович 1966 г. Shadwick & Spiegel 2012]
- Заказ Fractovitellida Lahr et al. 2011 г. Канг и др. 2017 г.
- Суперкласс Mycetozoa де Бари, 1859 г., бывший Ростафинский, 1873 г.
- Заказ Эхиностелиопсида
- Учебный класс Диктиостелея Хоксворт и др. 1983 г.
- Заказ Ацитоостелиальные Балдауф, Шейх и Тулин 2017
- Заказ Dictyosteliales Листер 1909 г. Оливковое 1970
- Учебный класс Цератиомиксомицеты Хоксворт, Саттон и Эйнсворт, 1983 г.
- Заказ Протоспорангииды Shadwick & Spiegel, 2012 г.
- Заказ Ceratiomyxida Мартин 1961 бывший Фарр и Алексопулос
- Учебный класс Миксомицеты Ссылка 1833 эм. Геккель 1866 г.
- Подкласс Lucisporomycetidae Леонтьев и др. 2019 г.
- Суперзаказ Cribrarianae Леонтьев 2015
- Заказ Cribrariales Macbr. 1922 г.
- Суперзаказ Trichianae Леонтьев 2015
- Заказ Ретикулярные Леонтьев 2015
- Заказ Liceales Ян 1928
- Заказ Трихиалес Макбрайд 1922
- Суперзаказ Cribrarianae Леонтьев 2015
- Подкласс Columellomycetidae Леонтьев и др. 2019 г.
- Суперзаказ Echinostelianae Леонтьев 2015
- Заказ Echinosteliales Мартин 1961
- Суперзаказ Stemonitanae Леонтьев 2015 [Fuscisporida Кавалер-Смит 2012]
- Заказ Clastodermatales Леонтьев 2015
- Заказ Meridermatales Леонтьев 2015
- Заказ Stemonitales Макбрайд 1922
- Заказ Physarales Макбрайд 1922
- Суперзаказ Echinostelianae Леонтьев 2015
- Подкласс Lucisporomycetidae Леонтьев и др. 2019 г.
- Учебный класс Variosea Кавальер-Смит и др. 2004 г.
- Инфрафилум Архамебы Кавалер-Смит 1993 стат. п. 1998 г.
- Clade Cutosa Cavalier-Smith 2016 стат. ноя
- Clade Tubulinea Смирнов и др. Стат. За 2005 г. ноя Adl et al. 2018 г.
Окаменелости
В форме вазы микрофоссилий (VSM), обнаруженные по всему миру, показывают, что амебозойные животные существовали с Неопротерозойский Эра. Ископаемые виды Меланоцириллий гексодиадема, Палеоарцелла атаната, и Hemisphaeriella ornata происходят из скал возрастом 750 миллионов лет. Все три VSM имеют полусферическую форму, инвагинированную апертуру и регулярные углубления, которые сильно напоминают современные арцеллиниды, которые раковинные амебоиды. P. athanata в частности выглядит так же, как и существующий род Арчелла.[19][20]
Список простейших амебозойных, патогенных для человека
Мейоз
Недавно доступные Акантамеба геном последовательность выявила несколько ортологи генов, используемых в мейоз сексуального эукариоты. Эти гены включали Spo11, Mre11, Rad50, Rad51, Rad52, Mnd1, Dmc1, Мш и Mlh.[21] Это открытие предполагает, что Акантамеба способен к некоторой форме мейоза и может подвергаться половому размножению.
У эукариот, размножающихся половым путем, гомологичная рекомбинация (HR) обычно возникает во время мейоза. Специфический для мейоза рекомбиназа, Dmc1, необходим для эффективного сердечного ритма мейоза, а Dmc1 выражается в Entamoeba histolytica.[22] Очищенный Dmc1 от E. histolytica формы пресинаптический нити и катализаторы АТФ -зависимый гомологичное спаривание ДНК и обмен цепями ДНК более чем на несколько тысяч пар оснований.[22] Реакции спаривания ДНК и обмена цепей усиливаются эукариотическим мейоз-специфическим дополнительным фактором рекомбинации (гетеродимером) Hop2-Mnd1.[22] Эти процессы являются центральными для мейотической рекомбинации, предполагая, что E. histolytica претерпевает мейоз.[22]
Исследования Entamoeba invadens обнаружил, что при преобразовании из тетраплоид безъядерный трофозоит к четырехъядерной кисте, гомологичная рекомбинация усиливается.[23] Экспрессия генов с функциями, связанными с основными этапами мейотической рекомбинации, также увеличивалась во время энцистаций.[23] Эти выводы в E. invadensв сочетании с данными исследований E. histolytica указывают на наличие мейоза в Entamoeba. Сравнительный генетический анализ показал, что мейотические процессы присутствуют во всех основных линиях амеб.[24]
С Амебозоа эти результаты также предполагают, что мейоз присутствовал на ранних этапах эволюции эукариот.
Человеческое здоровье
Амебиаз, также известный как амебиаз или энтамебиаз,[25][26] это инфекция, вызванная любым из амебозоидов Entamoeba группа. Симптомы чаще всего возникают при заражении Entamoeba histolytica. Амебиаз может протекать бесследно, в легкой или тяжелой форме. симптомы. Симптомы могут включать: боль в животе, незначительный понос, кровавый понос или же тяжелый колит с смерть ткани и перфорация. Это последнее осложнение может вызвать перитонит. У пострадавших может развиться анемия из-за потери крови.[27]
Причины поражения слизистой оболочки кишечника амебная кровавая диарея или же амебный колит. Если паразит попадает в кровоток, он может распространиться по телу, чаще всего в печени, где вызывает амебные абсцессы печени. Печень абсцессы могут возникать без предшествующей диареи. Кисты Entamoeba может выжить до месяца в почве или до 45 минут под ногтями. Важно различать амебиаз и бактериальный колит. Предпочтительный метод диагностики - исследование кала под микроскопом, но требует наличия опытного микроскописта и может быть ненадежным при исключении инфекции. Однако этот метод не может разделить определенные типы. Повышенное количество лейкоцитов присутствует в тяжелых случаях, но не в легких. Самый точный тест для антитела в крови, но он может оставаться положительным после лечения.[27]
Профилактика амебиаза заключается в отделении пищи и воды от фекалий и санитария меры. Вакцины нет. В зависимости от локализации инфекции существует два варианта лечения. Амебиаз в тканях лечится либо метронидазол, тинидазол, нитазоксанид, дегидроэметин или же хлорохин, в то время как люминальная инфекция лечится дилоксанид фуроат или же йодохинолин. Чтобы лечение было эффективным против всех стадий амебы, может потребоваться комбинация лекарств. Инфекции без симптомов не требуют лечения, но инфицированные люди могут передавать паразита другим, и можно рассмотреть возможность лечения. Лечение других Entamoeba инфекции помимо E. histolytica не нужен.[27]
Амебиаз присутствует во всем мире.[28] Около 480 миллионов человек инфицированы, как представляется, E. histolytica и это приводит к смерти от 40 000 до 110 000 человек ежегодно. Большинство инфекций теперь приписывают E. dispar. E. dispar чаще встречается в определенных областях, и симптоматических случаев может быть меньше, чем сообщалось ранее. Первый случай амебиаза был задокументирован в 1875 году, а в 1891 году болезнь была подробно описана, в результате чего были сформулированы следующие термины: амебной дизентерией и амебный абсцесс печени. Дальнейшие данные, полученные на Филиппинах в 1913 г., показали, что при проглатывании цист E. histolytica добровольцы заболели. С 1897 года известно, что по крайней мере один не вызывающий болезни вид Entamoeba существовал (Entamoeba coli), но впервые он был официально признан ВОЗ в 1997 г. E. histolytica было два вида, несмотря на то, что это было впервые предложено в 1925 году. В дополнение к признанным ныне E. dispar данные показывают, что существует по крайней мере два других вида Entamoeba которые выглядят одинаково у людей - Е. мошковский и Entamoeba бангладешский. Причина, по которой эти виды не были дифференцированы до недавнего времени, заключается в зависимости от внешнего вида.[27]
Галерея

Арчелла sp. тест (Lobosa: Tubulinea)

Акантамеба sp. (Лобоса: Дискоса )

Thecamoeba sp. (Лобоса: Дискоса)

Pelomyxa palustris (Conosa: Archamoebae)

Стемонит sp. (Коноса: Миксогастрия )

Dictyostelium discoideum (Коноса: Диктиостелия )








Рекомендации
- ^ Парфри Л.В., Лар DJ, Knoll AH, Кац Л.А. (август 2011 г.). «Оценка сроков ранней диверсификации эукариот с помощью мультигенных молекулярных часов». Труды Национальной академии наук Соединенных Штатов Америки. 108 (33): 13624–9. Дои:10.1073 / pnas.1110633108. ЧВК 3158185. PMID 21810989.
- ^ а б Pawlowski J, Audic S, Adl S, Bass D, Belbahri L, Berney C и др. (6 ноября 2012 г.). «Рабочая группа протистов CBOL: штрих-кодирование богатства эукариот за пределами царства животных, растений и грибов». PLOS Биология. 10 (11): e1001419. Дои:10.1371 / journal.pbio.1001419. ЧВК 3491025. PMID 23139639.
- ^ а б c d Адл С.М., Симпсон А.Г., Лейн С.Е., Лукеш Дж., Басс Д., Баузер С.С. и др. (Сентябрь 2012 г.). «Пересмотренная классификация эукариот». Журнал эукариотической микробиологии. 59 (5): 429–93. Дои:10.1111 / j.1550-7408.2012.00644.x. ЧВК 3483872. PMID 23020233.
- ^ а б c Кавальер-Смит Т., Фиоре-Донно А.М., Чао Э., Кудрявцев А., Берни С., Снелл Е.А., Льюис Р. (февраль 2015 г.). «Мультигенная филогения разрешает глубокое ветвление амебозоа». Молекулярная филогенетика и эволюция. 83: 293–304. Дои:10.1016 / j.ympev.2014.08.011. PMID 25150787.
- ^ Корлисс Дж. О. (1984). «Королевство Протиста и его 45 Фила». Биосистемы. 17 (2): 87–126. Дои:10.1016/0303-2647(84)90003-0. PMID 6395918.
- ^ а б Кавальер-Смит Т (2003). «Филогения простейших и классификация простейших высокого уровня». Европейский журнал протистологии. 39 (4): 338–348. Дои:10.1078/0932-4739-00002.
- ^ а б Дерелле Р., Торруэлла Г., Климеш В., Бринкманн Х., Ким Е., Влчек Ч., Ланг Б. Ф., Элиаш М. (февраль 2015 г.). «Бактериальные белки определяют единственный корень эукариот». Труды Национальной академии наук Соединенных Штатов Америки. 112 (7): E693–9. Дои:10.1073 / pnas.1420657112. ЧВК 4343179. PMID 25646484.
- ^ Жулидов Д.А., Робартс Р.Д., Жулидов А.В., Жулидова О.В., Маркелов Д.А., Русанов В.А., Хедли СП (2002). «Накопление цинка слизистой плесенью Fuligo septica (L.) Wiggers в бывшем Советском Союзе и Северной Корее». Журнал качества окружающей среды. 31 (3): 1038–42. Дои:10.2134 / jeq2002.1038. PMID 12026071.
- ^ Чон К.В. (1973). Биология амебы. Нью-Йорк: Academic Press. стр.100. ISBN 978-0-323-14404-9.
- ^ Eichinger L, Pachebat JA, Glöckner G, Rajandream MA, Sucgang R, Berriman M, et al. (Май 2005 г.). «Геном социальной амебы Dictyostelium discoideum». Природа. 435 (7038): 43–57. Дои:10.1038 / природа03481. ЧВК 1352341. PMID 15875012.
- ^ Докинз, Ричард; Вонг, Ян (2016). Рассказ предков. ISBN 978-0544859937.
- ^ Кавальер-Смит Т., Чао Э. (декабрь 1996 г.). «Молекулярная филогения свободноживущего архезоана Trepomonas agilis и природа первого эукариота». Журнал молекулярной эволюции. 43 (6): 551–62. Дои:10.1007 / BF02202103. PMID 8995052. S2CID 28992966.
- ^ Кавальер-Смит Т. (август 1998 г.). «Пересмотренная система жизни шести царств». Биологические обзоры Кембриджского философского общества. 73 (3): 203–66. Дои:10.1111 / j.1469-185X.1998.tb00030.x. PMID 9809012. S2CID 6557779.
- ^ Кавальер-Смит Т., Чао Э., Льюис Р. (июнь 2016 г.). «Филогения из 187 генов простейших типа Amoebozoa раскрывает новый класс (Cutosea) глубоко разветвленных, ультраструктурно уникальных, обволакивающих морских лобоза и проясняет эволюцию амеб». Молекулярная филогенетика и эволюция. 99: 275–296. Дои:10.1016 / j.ympev.2016.03.023. PMID 27001604.
- ^ Силар П. (2016). "Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes" (PDF). HAL archives-ouvertes. С. 1–462. ISBN 978-2-9555841-0-1.
- ^ Канг С., Тайс А.К., Шпигель Ф.В., Зильберман Дж.Д., Панек Т., Цепицка И. и др. (Сентябрь 2017 г.). «Между стручком и тяжелым испытанием: глубокая эволюция амеб». Молекулярная биология и эволюция. 34 (9): 2258–2270. Дои:10.1093 / молбев / msx162. ЧВК 5850466. PMID 28505375.
- ^ Панек Т., Задробилкова Э., Уокер Дж., Браун М.В., Гентекаки Э., Градова М. и др. (Май 2016). «Первый мультигенный анализ Archamoebae (Amoebozoa: Conosa) убедительно раскрывает его филогению и показывает, что Entamoebidae представляет собой глубокую ветвь этой группы». Молекулярная филогенетика и эволюция. 98: 41–51. Дои:10.1016 / j.ympev.2016.01.011. PMID 26826602.
- ^ Леонтьев, Дмитрий В .; Шнитлер, Мартин; Стивенсон, Стивен Л .; Новожилов Юрий К .; Щепин, Олег Н. (март 2019). «К филогенетической классификации миксомицетов». Фитотакса. 399 (3): 209–238. Дои:10.11646 / phytotaxa.399.3.5.
- ^ Портер С.Х., Мейстерфельд Р., Нолл А.Х. (2003). «Вазообразные микрофоссилии из неопротерозойской группы Chuar, Гранд-Каньон: классификация, проводимая современными раковинными амебами» (PDF). Журнал палеонтологии. 77 (3): 409–429. Дои:10.1666 / 0022-3360 (2003) 077 <0409: VMFTNC> 2.0.CO; 2.
- ^ Портер С.М. (2006). "Протерозойские ископаемые летописи гетеротрофных эукариот". В Xiao S, Kaufman AJ (ред.). Неопротерозойская геолобиология и палеобиология. Разделы геобиологии. 27. Дордрехт, Нидерланды: Springer. С. 1–21. Дои:10.1007/1-4020-5202-2. ISBN 978-1-4020-5201-9.
- ^ Хан Н.А., Сиддики Р. (июнь 2015 г.). «Есть ли свидетельства полового размножения (мейоза) у акантамебы?». Патогены и глобальное здоровье. 109 (4): 193–5. Дои:10.1179 / 2047773215Y.0000000009. ЧВК 4530557. PMID 25800982.
- ^ а б c d Келсо А.А., Сэй А.Ф., Шарма Д., Ледфорд Л.Л., Турчик А., Саски К.А., Кинг А.В., Аттавей С.К., Темесвари Л.А., Сехорн М.Г. (2015). «Entamoeba histolytica Dmc1 катализирует гомологичное спаривание ДНК и обмен цепей, который стимулируется кальцием и Hop2-Mnd1». PLOS ONE. 10 (9): e0139399. Дои:10.1371 / journal.pone.0139399. ЧВК 4589404. PMID 26422142.
- ^ а б Сингх Н., Бхаттачарья А., Бхаттачарья С. (2013). «Гомологичная рекомбинация происходит у Entamoeba и усиливается во время стресса роста и конверсии стадий». PLOS ONE. 8 (9): e74465. Дои:10.1371 / journal.pone.0074465. ЧВК 3787063. PMID 24098652.
- ^ Hofstatter PG, Brown MW, Lahr DJG (ноябрь 2018 г.). «Сравнительная геномика поддерживает пол и мейоз у различных амебозоа». Геном Биол Эвол. 10 (11): 3118–3128. Дои:10.1093 / gbe / evy241. ЧВК 6263441. PMID 30380054.
- ^ «Энтамебиаз - MeSH - NCBI». www.ncbi.nlm.nih.gov. Получено 2015-07-21.
- ^ «Энтамебиаз». mesh.kib.ki.se. Получено 2015-07-21.
- ^ а б c d Фаррар Дж., Хотез П., Юнгханс Т., Канг Дж., Лаллоо Д., Белый Нью-Джерси (2013-10-26). Тропические болезни Мэнсона. Elsevier Health Sciences. С. 664–671. ISBN 978-0-7020-5306-1.
- ^ Бичинг Н, Гилл Г (17 апреля 2014 г.). «19». Конспект лекций: тропическая медицина. Джон Вили и сыновья. С. 177–182. ISBN 978-1-118-73456-8.
дальнейшее чтение
- Шильде С., Шаап П. (2013). «Амебозоа» (PDF). Dictyostelium discoideum Протоколы. Методы молекулярной биологии. 983. С. 1–15. Дои:10.1007/978-1-62703-302-2_1. ISBN 978-1-62703-301-5. ЧВК 4037983. PMID 23494299.
внешняя ссылка
- Павловски Я. «Молекулярная филогения амебоидных протистов - древо амебозоа». Группа молекулярной систематики (MSG). Кафедра зоологии и биологии животных Женевского университета. Архивировано из оригинал 2 января 2005 г.
- Килинг П., Леандер Б.С., Симпсон А. «Древо жизни эукариотов». Проект «Древо жизни».
- Лейди Дж (1879). «Тарелки амебы». Пресноводные корневища Северной Америки. Вашингтон, округ Колумбия: Государственная типография.
- «Амебозоа». Браузер таксономии NCBI. 554915.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Дискоса |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tubulinea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Evosea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||