Митоз - Mitosis




а. неделящиеся клетки
б. подготовка ядер к делению (стадия спиремы)
c. делящиеся клетки с митотическими фигурами
е. пара дочерних клеток вскоре после деления
В клеточная биология, митоз (/маɪˈтoʊsɪs/) является частью клеточный цикл, в котором воспроизведены хромосомы разделяются на два новых ядра. Деление клеток приводит к образованию генетически идентичных клеток, в которых сохраняется общее количество хромосом.[1] В целом митозу (делению ядра) предшествует S стадия межфазный (во время которого ДНК реплицируется) и часто сопровождается телофаза и цитокинез; что делит цитоплазма, органеллы и клеточная мембрана из одной ячейки в две новые клетки содержащие примерно равные доли этих клеточных компонентов.[2] Различные стадии митоза вместе определяют митотический (M) фаза цикла животных клеток - разделение материнской клетки на две дочерние клетки, генетически идентичные друг другу[3].
Процесс митоза делится на этапы, соответствующие завершению одного набора действий и началу следующего. Эти этапы профаза, прометафаза, метафаза, анафаза, и телофаза. Во время митоза хромосомы, которые уже удвоились, конденсируются и прикрепляются к веретену. волокна которые тянут по одной копии каждой хромосомы на противоположные стороны клетки.[4] В результате получается два генетически идентичных дочерних ядра. Оставшаяся часть клетки может продолжить деление путем цитокинеза с образованием двух дочерних клеток.[5] Различные фазы митоза можно визуализировать в реальном времени, используя визуализация живых клеток.[6] Производство трех или более дочерних клеток вместо нормальных двух - это митотическая ошибка, называемая триполярным митозом или мультиполярным митозом (прямое утроение / умножение клеток).[7] Другие ошибки во время митоза могут вызвать апоптоз (запрограммированная гибель клеток) или причина мутации. Определенные виды рак могут возникнуть в результате таких мутаций.[8]
Митоз возникает только в эукариотический клетки. Прокариотический клетки, лишенные ядра, делятся с помощью другого процесса, называемого двойное деление[нужна цитата ]. Митоз варьируется между организмами.[9] Например, животное клетки проходят «открытый» митоз, когда ядерная оболочка распадается до разделения хромосом, тогда как грибы проходят «закрытый» митоз, когда хромосомы делятся внутри неповрежденного ядра клетки.[10] Большинство клеток животных претерпевают изменение формы, известное как округление митотических клеток, чтобы принять почти сферическую морфологию в начале митоза. Большинство клеток человека продуцируются митотическим делением клеток. Важные исключения включают гаметы – сперма и яйцо клетки - которые производятся мейоз.
Открытие
Многочисленные описания деление клеток изготовлены в 18-19 веках с разной степенью точности.[11] В 1835 году немецкий ботаник Хьюго фон Моль, описал деление клеток в зеленая водоросль Cladophora glomerata, утверждая, что размножение клеток происходит посредством деления клеток.[12][13][14] В 1838 г. Маттиас Якоб Шлейден подтвердили, что образование новых клеток в их интерьере был общим законом для размножения клеток у растений, который позже был отвергнут в пользу модели Моля из-за вклада Роберт Ремак и другие.[15]
В клетках животных деление клеток с митозом было обнаружено у лягушки, кролика и кошки. роговица клетки в 1873 г. и впервые описанные польскими гистолог Вацлав Майзель в 1875 г.[16][17]
Бютчли, Шнайдер и Фол могли также заявить об открытии процесса, известного в настоящее время как «митоз».[11] В 1873 году немецкий зоолог Отто Бютчли опубликованные данные наблюдений за нематоды. Несколько лет спустя он открыл и описал митоз на основе этих наблюдений.[18][19][20]
Термин «митоз», введенный Вальтер Флемминг в 1882 г.,[21] происходит из Греческий слово μίτος (митос, «нить основы»).[22][23] Есть несколько альтернативных названий процесса,[24] например, «кариокинез» (деление ядра), термин, введенный Шлейхером в 1878 г.,[25][26] или "эквациональное деление", предложенное Август Вейсманн в 1887 г.[27] Однако термин «митоз» также используется некоторыми авторами в широком смысле для обозначения кариокинеза и цитокинез вместе.[28] В настоящее время «эквациональное деление» чаще используется для обозначения мейоз II, часть мейоза больше всего похожа на митоз [29].
Фазы
Обзор

Первичным результатом митоза и цитокинеза является перенос родительской клетки. геном на две дочерние клетки. Геном состоит из ряда хромосом - комплексов плотно свернутых ДНК которые содержат генетическая информация жизненно важен для правильного функционирования клеток [30]. Поскольку каждая полученная дочерняя клетка должна быть генетически идентичный для родительской клетки родительская клетка должна сделать копию каждой хромосомы перед митозом. Это происходит во время S фаза межфазного.[31] Дублирование хромосомы приводит к двум идентичным сестринские хроматиды связаны вместе когезин белки на центромера.
Когда начинается митоз, хромосомы уплотняются и становятся видимыми. У некоторых эукариот, например животных, ядерная оболочка, который отделяет ДНК от цитоплазмы, распадается на мелкие пузырьки. В ядрышко, который производит рибосомы в клетке, также исчезает. Микротрубочки выступают из противоположных концов клетки, прикрепляются к центромерам и выравнивают хромосомы по центру клетки. Затем микротрубочки сокращаются, чтобы разделить сестринские хроматиды каждой хромосомы.[32] Сестринские хроматиды на этом этапе называются дочерние хромосомы. По мере удлинения клетки соответствующие дочерние хромосомы притягиваются к противоположным концам клетки и максимально конденсируются в поздней анафазе. Новая ядерная оболочка образуется вокруг разделенных дочерних хромосом, которые деконденсируются с образованием интерфазных ядер.
Во время митотической прогрессии, обычно после начала анафазы, клетка может подвергаться цитокинезу. В клетки животных, а клеточная мембрана защипывается внутрь между двумя развивающимися ядрами, чтобы произвести две новые клетки. В клетки растений, а клеточная пластина образуется между двумя ядрами. Цитокинез возникает не всегда; ценоцитарные (тип многоядерных состояний) клетки проходят митоз без цитокинеза.

Межфазный
Митотическая фаза - это относительно короткий период клеточный цикл. Он чередуется с гораздо более длинными межфазный, где клетка готовится к процессу клеточного деления. Интерфаза разделена на три фазы: грамм1 (первый пробел), S (синтез), и грамм2 (второй пробел). Во время всех трех частей интерфазы клетка растет, продуцируя белки и цитоплазматические органеллы. Однако репликация хромосом происходит только во время S фаза. Таким образом, клетка растет (G1), продолжает расти, поскольку он дублирует свои хромосомы (S), растет больше и готовится к митозу (G2) и, наконец, делит (M) перед перезапуском цикла.[31] Все эти фазы клеточного цикла строго регулируются циклины, циклин-зависимые киназы и другие белки клеточного цикла. Фазы следуют одна за другой в строгом порядке, и есть "контрольно-пропускные пункты "которые дают клетке сигналы перейти от одной фазы к другой.[33] Клетки также могут временно или навсегда покинуть клеточный цикл и войти в грамм0 фаза прекратить деление. Это может произойти, когда клетки становятся переполненными (зависимое от плотности ингибирование ) или когда они различать выполнять определенные функции для организма, как в случае клетки сердечной мышцы человека и нейроны. Некоторые G0 клетки обладают способностью повторно входить в клеточный цикл.
Двухцепочечные разрывы ДНК могут быть отремонтирован во время интерфазы двумя основными процессами.[34] Первый процесс, негомологичное соединение концов (NHEJ), может соединить два разорванных конца ДНК в G1, S и G2 фазы интерфазы. Второй процесс, гомологичный рекомбинационный Repair (HRR), более точен, чем NHEJ в ремонте двухцепочечных разрывов. HRR активен во время фаз S и G2 интерфазы, когда Репликация ДНК выполняется либо частично, либо после завершения, так как HRR требует двух смежных гомологи.
Интерфаза помогает подготовить клетку к митотическому делению. Он определяет, произойдет ли деление митотических клеток. Он тщательно останавливает клетку от продолжения, когда ДНК клетки повреждена или не завершила важную фазу. Интерфаза очень важна, поскольку она определяет успешное завершение митоза. Это уменьшит количество продуцируемых поврежденных клеток и производство раковых клеток. Просчет ключевых интерфазных белков может иметь решающее значение, поскольку последние потенциально могут создавать раковые клетки. [35]. Сегодня проводится больше исследований, чтобы понять, как именно протекают указанные выше фазы.
Митоз

Препрофаза (растительные клетки)
Только в клетках растений профазе предшествует предпрофазная стадия. В очень вакуолизированный В клетках растений ядро должно мигрировать в центр клетки, прежде чем митоз может начаться. Это достигается за счет формирования фрагмосома, поперечный лист цитоплазмы, который рассекает клетку пополам по будущей плоскости деления клетки. Помимо образования фрагмосом, препрофаза характеризуется образованием кольца микротрубочек и актин волокна (называемые препрофазная полоса ) под плазматической мембраной вокруг экваториальной плоскости будущего митотического шпиндель. Эта полоса отмечает положение, в котором клетка в конечном итоге разделится. Клетки высших растений (например, цветущие растения ) недостаток центриоли; вместо этого микротрубочки образуют веретено на поверхности ядра и затем организуются в веретено самими хромосомами после разрушения ядерной оболочки.[36] При разрушении ядерной оболочки и формировании веретена в прометафазе полоса препрофазы исчезает.[37]:58–67
Профаза


Во время профазы, которая возникает после G2 интерфаза, клетка готовится к делению, плотно конденсируя свои хромосомы и инициируя формирование митотического веретена. Во время интерфазы генетический материал ядра состоит из неплотно упакованных хроматин. В начале профазы волокна хроматина конденсируются в дискретные хромосомы, которые обычно видны при большом увеличении через оптический микроскоп. На этой стадии хромосомы длинные, тонкие и нитевидные. Каждая хромосома имеет две хроматиды. Две хроматиды соединены центромеры.
Транскрипция генов прекращается во время профазы и не возобновляется до поздней анафазы до начала G1 фаза.[38][39][40] В ядрышко также исчезает на ранней профазе.[41]
Рядом с ядром клеток животных находятся структуры, называемые центросомы, состоящий из пары центриоли окруженный рыхлый сбор белков. Центросома является координирующим центром клеточного микротрубочки. Клетка наследует одну центросому при делении клетки, то есть дублируется ячейкой до того, как начнется новый раунд митоза, давая пару центросом. Две центросомы полимеризуются тубулин чтобы помочь сформировать аппарат веретена микротрубочек. Моторные белки затем протолкните центросомы вдоль этих микротрубочек к противоположным сторонам клетки. Хотя центросомы помогают организовать сборку микротрубочек, они не важны для формирования аппарата веретена, так как они отсутствуют у растений,[36] и не являются абсолютно необходимыми для митоза клеток животных.[42]
Прометафаза
В начале прометафазы в клетках животных фосфорилирование ядерные ламины вызывает ядерная оболочка распадаться на мелкую мембрану пузырьки. Когда это происходит, микротрубочки вторгаются в ядерное пространство. Это называется открытый митоз, и это происходит в некоторых многоклеточных организмах. Грибы и некоторые протисты, Такие как водоросли или же трихомонады, пройти вариацию, называемую закрытый митоз где веретено формируется внутри ядра, или микротрубочки проникают через неповрежденную ядерную оболочку.[43][44]
В поздней прометафазе кинетохорные микротрубочки начать искать и прикреплять к хромосомному кинетохоры.[45] А кинетохора это белковый структура, связывающая микротрубочки, которая формируется на центромере хромосомы во время поздней профазы.[45][46] Номер полярные микротрубочки находят и взаимодействуют с соответствующими полярными микротрубочками из противоположной центросомы, чтобы сформировать митотическое веретено.[47] Хотя структура и функция кинетохор до конца не изучены, известно, что они содержат некоторую форму молекулярный мотор.[48] Когда микротрубочка соединяется с кинетохорой, мотор активируется, используя энергию от АТФ чтобы «ползти» вверх по трубке к исходной центросоме. Эта двигательная активность в сочетании с полимеризацией и деполимеризацией микротрубочек обеспечивает тянущее усилие, необходимое для последующего разделения двух хроматид хромосомы.[48]
Метафаза


После того, как микротрубочки локализованы и прикреплены к кинетохорам в прометафазе, две центросомы начинают тянуть хромосомы к противоположным концам клетки. Возникающее напряжение заставляет хромосомы выравниваться по метафазная пластинка или же экваториальная плоскость, воображаемая линия, которая расположена в центре между двумя центросомами (примерно по средней линии клетки).[47] Чтобы обеспечить справедливое распределение хромосом в конце митоза, контрольная точка метафазы гарантирует, что кинетохоры правильно прикреплены к митотическому веретену и что хромосомы выровнены вдоль метафазной пластинки.[49] Если клетка успешно проходит через контрольную точку метафазы, она переходит в анафазу.
Анафаза

В течение анафаза А, то когезины которые связывают сестринские хроматиды вместе, расщепляются, образуя две идентичные дочерние хромосомы.[50] Укорочение микротрубочек кинетохор притягивает вновь образованные дочерние хромосомы к противоположным концам клетки. В течение анафаза Bполярные микротрубочки толкаются друг к другу, заставляя клетку удлиняться.[51] В поздней анафазе хромосомы также достигают максимального общего уровня конденсации, чтобы помочь сегрегация хромосом и переформирование ядра.[52] В большинстве клеток животных анафаза А предшествует анафазе В, но некоторые яйцеклетки позвоночных демонстрируют противоположный порядок событий.[50]
Телофаза

Телофаза (от Греческий слово τελος означает «конец») - это обращение профазных и прометафазных событий. В телофазе полярные микротрубочки продолжают удлиняться, удлиняя клетку еще больше. Если ядерная оболочка разрушена, новая ядерная оболочка образуется с использованием мембранных пузырьков старой ядерной оболочки родительской клетки. Новая оболочка формируется вокруг каждого набора разделенных дочерних хромосом (хотя мембрана не охватывает центросомы), и ядрышко появляется снова. Оба набора хромосом, теперь окруженные новой ядерной мембраной, начинают «расслабляться» или деконденсироваться. Митоз полный. Каждое дочернее ядро имеет идентичный набор хромосом. Деление клеток может происходить или не происходить в это время в зависимости от организма.
Цитокинез


Цитокинез - это не фаза митоза, а скорее отдельный процесс, необходимый для завершения деления клетки. В клетках животных борозда декольте (защемление), содержащее сократительное кольцо, развивается там, где раньше была метафазная пластинка, отщипывая разделенные ядра.[53] Как в животных, так и в растительных клетках деление клеток также осуществляется везикулами, происходящими из аппарат Гольджи, которые движутся по микротрубочкам к середине клетки.[54] У растений эта структура сливается в клеточную пластину в центре фрагмопласт и развивается в клеточную стенку, разделяющую два ядра. Фрагмопласт представляет собой структуру микротрубочек, типичную для высших растений, тогда как некоторые зеленые водоросли используют фикопласт Массив микротрубочек во время цитокинеза.[37]:64–7, 328–9 Каждая дочерняя клетка имеет полную копию генома своей родительской клетки. Конец цитокинеза отмечает конец М-фазы.
Есть много клеток, в которых митоз и цитокинез происходят отдельно, образуя отдельные клетки с множеством ядер. Наиболее заметное проявление этого - среди грибы, слизевые формы, и ценоцитарные водоросли, но это явление встречается и у различных других организмов. Даже у животных цитокинез и митоз могут происходить независимо, например, на определенных этапах плодовая муха эмбриональное развитие.[55]
Функция
Митоз "функция "или значимость зависит от сохранения хромосомного набора; каждая сформированная клетка получает хромосомы, которые по составу и количеству равны хромосомам родительской клетки.
Митоз возникает при следующих обстоятельствах:
- Развитие и рост: Количество клеток в организме увеличивается за счет митоза. Это основа развития многоклеточного тела из одной клетки, т.е. зигота а также основа роста многоклеточный тело.
- Замена ячейки: В некоторых частях тела, например Клетки кожи и пищеварительного тракта постоянно отшелушиваются и заменяются новыми. Новые клетки образуются в результате митоза, как и точные копии заменяемых клеток. Таким же образом красные кровяные тельца имеют короткую продолжительность жизни (всего около 4 месяцев), а новые эритроциты образуются путем митоза.
- Регенерация: Некоторые организмы могут регенерировать части тела. Производство новых клеток в таких случаях достигается митозом. Например, морская звезда регенерировать потерянные руки через митоз.
- Бесполое размножение: Некоторые организмы производят генетически похожее потомство за счет бесполое размножение. Например, гидра размножается бесполым почкованием. Клетки на поверхности гидры подвергаются митозу и образуют массу, называемую почкой. Митоз продолжается в клетках почки, и это перерастает в новую особь. Такое же деление происходит при бесполом размножении или вегетативное размножение в растениях.
Вариации
Формы митоза
Процесс митоза в клетках эукариотических организмов происходит по аналогичной схеме, но с вариациями в трех основных деталях. «Закрытый» и «открытый» митоз можно различить на основе ядерная оболочка остаются неповрежденными или ломаются. Промежуточная форма с частичной деградацией ядерной оболочки называется «полуоткрытым» митозом. С учетом симметрии шпиндельный аппарат во время метафазы приблизительно аксиально-симметричная (центрированная) форма называется «ортомитозом», в отличие от эксцентрических веретен «плевромитоза», при котором митотический аппарат имеет двустороннюю симметрию. Наконец, третий критерий - расположение центральный шпиндель при закрытом плевромитозе: «внеядерный» (веретено в цитоплазме) или «внутриядерный» (в ядре).[9]

закрыто
внутриядерный
плевромитоз
закрыто
внеядерный
плевромитоз
закрыто
ортомитоз
полуоткрытый
плевромитоз
полуоткрытый
ортомитоз
открыто
ортомитоз




Ядерное деление происходит только в клетках организмов эукариотический домен, как бактерии и археи не имеют ядра. Бактерии и археи делятся по другому типу.[нужна цитата ]Внутри каждого эукариотического супергруппы встречается митоз открытой формы, а также митоз закрытый, за исключением Экскавата, которые демонстрируют исключительно закрытый митоз.[56] Следующее, возникновение форм митоза у эукариот:[9][57]
- Закрытый внутриядерный плевромитоз типично для Фораминиферы, немного Празиномонадида, немного Кинетопластида, то Оксимонадида, то Гаплоспоридии, много грибов (хитриды, оомицеты, зигомицеты, аскомицеты ), и немного Радиолярии (Спумеллария и Акантария ); вроде бы самый примитивный тип.
- Закрытый экстраядерный плевромитоз происходит в Трихомонадиды и Динофлагеллата.
- Закрытый ортомитоз находится среди диатомеи, инфузории, немного Микроспоридии, одноклеточный дрожжи и некоторые многоклеточные грибы.
- Полуоткрытый плевромитоз типично для большинства Apicomplexa.
- Полуоткрытый ортомитоз встречается с разными вариантами у некоторых амеб (Лобоса ) и некоторые зеленые жгутиковые (например, Raphidophyta или же Volvox ).
- Открытый ортомитоз типично в млекопитающие и другие Metazoa, И в наземные растения; но это также встречается у некоторых протистов.
Ошибки и другие вариации
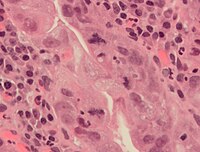
Ошибки могут возникать во время митоза, особенно на ранней стадии. эмбриональный развитие у человека.[58] На каждом этапе митоза обычно есть контрольные точки, которые контролируют нормальный результат митоза. [59]. Но иногда и почти редко ошибки случаются. Митотические ошибки могут создавать анеуплоидный клетки, у которых слишком мало или слишком много одной или нескольких хромосом, состояние, связанное с рак.[60][61] Ранние человеческие эмбрионы, раковые клетки, инфицированные или отравленные клетки также могут страдать от патологического деления на три или более дочерних клетки (триполярный или многополярный митоз), что приводит к серьезным ошибкам в их хромосомных дополнениях.[7]
В нерасхождение сестринские хроматиды не разделяются во время анафазы.[62] Одна дочерняя клетка получает обе сестринские хроматиды от неразрывной хромосомы, а другая клетка не получает ни одной. В результате первая клетка получает три копии хромосомы, состояние, известное как трисомия, и последний будет иметь только одну копию, условие, известное как моносомия. Иногда, когда клетки испытывают нерасхождение, они не могут завершить цитокинез и удерживать оба ядра в одной клетке, что приводит к двухъядерные клетки.[63]
Запаздывание анафазы возникает, когда движение одной хроматиды затруднено во время анафазы.[62] Это может быть вызвано неспособностью митотического веретена должным образом прикрепиться к хромосоме. Отстающая хроматида исключена из обоих ядер и утрачена. Следовательно, одна из дочерних клеток будет моносомной для этой хромосомы.
Эндоредупликация (или эндорепликация) происходит, когда хромосомы дублируются, но клетка впоследствии не делится. Это приводит к полиплоид клетки или, если хромосомы многократно дублируются, политенные хромосомы.[62][64] Эндоредупликация встречается у многих видов и, по-видимому, является нормальной частью разработка.[64] Эндомитоз - это вариант эндоредупликации, при котором клетки реплицируют свои хромосомы во время S-фазы и вступают в митоз, но преждевременно завершают его. Вместо разделения на два новых дочерних ядра реплицированные хромосомы сохраняются в исходном ядре.[55][65] Затем клетки снова входят в G1 и S-фаза и снова реплицируют свои хромосомы.[65] Это может происходить несколько раз, увеличивая число хромосом с каждым раундом репликации и эндомитоза. Тромбоцит -производство мегакариоциты проходят эндомитоз во время клеточной дифференцировки.[66][67]
Амитоз у инфузорий и в тканях плаценты животных приводит к случайному распределению родительских аллелей.
Кариокинез без цитокинеза происходит многоядерный ячейки называются ценоциты.
Диагностический маркер

В гистопатология скорость митоза является важным параметром в различных типах образцов тканей, как для диагностики, так и для дальнейшего определения агрессивности опухолей. Например, обычно проводится количественная оценка митотический счет в классификации рака груди.[68] Митозы необходимо подсчитывать в области с наибольшей митотической активностью. Визуально идентифицировать эти области сложно в опухолях с очень высокой митотической активностью.[69] Также обнаружение атипичных форм митоза может использоваться как диагностический, так и прогностический маркер.[нужна цитата ] Например, лаг-митоз (неприкрепленное сжатое хроматин в области митотической фигуры) указывает на высокий риск папилломавирусная инфекция -связанные с Рак шейки матки.[нужна цитата ]

Нормальные и атипичные формы митоза в раковых клетках. А - нормальный митоз; B, хроматиновый мостик; C - мультиполярный митоз; D - кольцевой митоз; E - диспергированный митоз; F - асимметричный митоз; G - митоз лаг-типа; и H, микроядра. Пятно H&E.

Связанные клеточные процессы
Округление ячеек

В животной ткани большинство клеток во время митоза округляются до почти сферической формы.[70][71][72] В эпителий и эпидермис, эффективный процесс округления коррелирует с правильным митотическое веретено выравнивание и последующее правильное расположение дочерних клеток.[71][72][73][74] Более того, исследователи обнаружили, что сильное подавление округления может привести к дефектам шпинделя, в первую очередь к расщеплению полюсов и невозможности эффективного захвата. хромосомы.[75] Следовательно, округление митотических клеток считается, что он играет защитную роль в обеспечении точного митоза.[74][76]
Силы округления вызваны реорганизацией F-актин и миозин (актомиозин) в сократительную однородную клеточная кора что 1) укрепляет периферию клетки[76][77][78] и 2) способствует образованию внутриклеточных гидростатическое давление (до 10 раз выше, чем межфазный ).[79][80][81] Создание внутриклеточного давления особенно важно в условиях ограничения, которое было бы важно в сценарии с тканями, где должны создаваться внешние силы, направленные против окружающих клеток и / или внеклеточный матрикс. Создание давления зависит от формин -опосредованный F-актин зарождение[81] и Ро-киназа (РОК) -опосредованный миозин II сокращение[77][79][81] оба из которых регулируются сигнальными путями. RhoA и ECT2[77][78] через деятельность Cdk1.[81] Благодаря его важности в митозе, молекулярные компоненты и динамика митотического актомиозиновая кора это область активных исследований.
Митотическая рекомбинация
Митотические клетки, облученные Рентгеновские лучи в Фаза G1 из клеточный цикл ремонт рекомбиногенный Повреждения ДНК в первую очередь рекомбинация между гомологичные хромосомы.[82] Митотические клетки, облученные в Фаза G2 устранять такие повреждения преимущественно путем рекомбинация сестринских хроматид.[82] Мутации в гены кодирующие ферменты, используемые в рекомбинации, вызывают повышенную чувствительность клеток к уничтожению различными повреждающими ДНК агентами.[83][84][85] Эти результаты предполагают, что митотическая рекомбинация является адаптацией для восстановления повреждений ДНК, в том числе потенциально летальных.
Эволюция
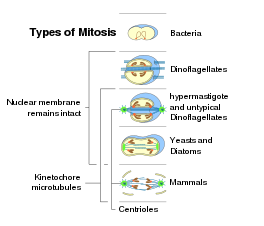
Есть прокариотические гомологи всех ключевых молекул митоза эукариот (например, актинов, тубулинов). Являясь универсальным эукариотическим свойством, митоз, вероятно, возник в основе эукариотического древа. Поскольку митоз менее сложен, чем мейоз, мейоз мог возникнуть после митоза.[86] Однако половое размножение с участием мейоза также является примитивной характеристикой эукариот.[87] Таким образом, мейоз и митоз могли развиться параллельно из наследственных прокариотических процессов.
Пока в деление бактериальных клеток, после дублирование ДНК, две кольцевые хромосомы прикреплены к особому участку клеточной мембраны, митоз эукариот обычно характеризуется наличием множества линейных хромосом, кинетохоры которых прикрепляются к микротрубочкам веретена. В отношении форм митоза закрытый внутриядерный плевромитоз представляется наиболее примитивным типом, поскольку он больше похож на деление бактерий.[9]
Галерея
Митотические клетки можно визуализировать под микроскопом с помощью окрашивание их с флуоресцентный антитела и красители.

Ранняя профаза: Полярные микротрубочки, показанные как зеленые нити, создали матрицу вокруг в настоящее время неповрежденного ядра с конденсирующимися хромосомами синего цвета. Красные узелки - центромеры.

Ранняя прометафаза: Ядерная мембрана только что разобрана, что позволяет микротрубочкам быстро взаимодействовать с кинетохорами, которые собираются на центромерах конденсирующихся хромосом.

Метафаза: Центросомы переместились к полюсам клетки и установили митотическое веретено. Хромосомы собрались на метафазной пластинке.

Анафаза: Микротрубочки кинетохор раздвигают два набора хромосом, а удлинение полярных микротрубочек раздвигает половинки делящейся клетки дальше друг от друга, в то время как хромосомы максимально конденсируются.

Телофаза: Обращение профазных и прометафазных событий и, таким образом, завершение клеточный цикл.





Смотрите также
Рекомендации
- ^ «Деление и рост клеток». britannica.com. ENCYCLOPDIA BRITANNICA. В архиве с оригинала на 2018-10-28. Получено 2018-11-04.
- ^ Картер Дж. С. (14 января 2014 г.). "Митоз". biology.clc.uc.edu. Архивировано из оригинал на 2012-10-27. Получено 2019-11-12.
- ^ «Митоз - обзор | Темы ScienceDirect». www.sciencedirect.com. Получено 2020-11-24.
- ^ «Разделение клеток: этапы митоза | Изучение науки в Scitable». www.nature.com. В архиве из оригинала от 14.11.2015. Получено 2015-11-16.
- ^ Матон А., Хопкинс Дж. Дж., ЛаХарт С., Куон Уорнер Д., Райт М., Джилл Д. (1997). Клетки: строительные блоки жизни. Нью-Джерси: Прентис-Холл. стр.70–4. ISBN 978-0-13-423476-2.
- ^ Сандоз ПА (декабрь 2019 г.). «Анализ изображений живых клеток млекопитающих с использованием трехмерных карт показателя преломления без меток позволяет выявить новую динамику органелл и поток сухой массы». PLOS Биология. 17 (12): e3000553. Дои:10.1371 / journal.pbio.3000553. ЧВК 6922317. PMID 31856161.
- ^ а б Калатова Б., Есенская Р., Глинка Д., Дудас М. (январь 2015 г.). «Триполярный митоз в клетках и эмбрионах человека: возникновение, патофизиология и медицинские последствия». Acta Histochemica. 117 (1): 111–25. Дои:10.1016 / j.acthis.2014.11.009. PMID 25554607.
- ^ Копс Г.Дж., Уивер Б.А., Кливленд Д.В. (октябрь 2005 г.). «На пути к раку: анеуплоидия и митотическая контрольная точка». Обзоры природы. Рак. 5 (10): 773–85. Дои:10.1038 / nrc1714. PMID 16195750. S2CID 2515388.
- ^ а б c d Райков И.Б. (1994). «Разнообразие форм митоза у простейших: сравнительный обзор». Европейский журнал протистологии. 30 (3): 253–69. Дои:10.1016 / S0932-4739 (11) 80072-6.
- ^ Де Соуза С.П., Османи С.А. (сентябрь 2007 г.). «Митоз, не только открытый или закрытый». Эукариотическая клетка. 6 (9): 1521–7. Дои:10.1128 / EC.00178-07. ЧВК 2043359. PMID 17660363.
- ^ а б Росс, Анна Э. "Анатомия и физиология человека I: хронология описания митоза". Университет христианских братьев. Дата обращения 2 мая 2018. связь В архиве 2016-05-12 в Wayback Machine.
- ^ фон Моль H (1835). Ueber die Vermehrung der Pflanzenzellen durch Theilung. Инаугурационная диссертация (Тезис). Тюбинген.
- ^ Карл Мэгдефрау (1994), "Моль, Хьюго фон", Neue Deutsche Biographie (NDB) (на немецком), 17, Берлин: Duncker & Humblot, стр. 690–691.; (полный текст онлайн )
- ^ «Заметки и меморандумы: покойный профессор фон Моль». Ежеквартальный журнал микроскопической науки, т. XV, Новая серия, стр. 178-181, 1875. связь.
- ^ Вейерс, Вольфганг (2002). 150 лет деления клеток. Дерматопатология: практическая и концептуальная, Vol. 8, №2. связь В архиве 2019-04-02 в Wayback Machine
- ^ Комендер Дж. (2008). "Kilka słów o doktorze Wacławie Mayzlu i jego odkryciu" [О Вацлаве Майзеле и его наблюдении за митотическим делением] (PDF). Postępy Biologii Komórki (по польски). 35 (3): 405–407. В архиве (PDF) из оригинала от 27.10.2012.
- ^ Иловецкий М (1981). Dzieje nauki polskiej. Варшава: Wydawnictwo Interpress. п. 187. ISBN 978-83-223-1876-8.
- ^ Бючли, О. (1873). Beiträge zur Kenntnis der freilebenden Nematoden. Nova Acta der Kaiserlich Leopoldinisch-Carolinischen Deutschen Akademie der Naturforscher 36, 1-144. связь В архиве 2018-08-11 в Wayback Machine.
- ^ Бючли, О. (1876). Studien über die ersten Entwicklungsvorgänge der Eizelle, die Zelleilung und die Conjugation der Infusorien. Abh.d. Senckenb. Натурф. Ges. Франкфурт а. М. 10, 213-452. связь В архиве 2018-08-09 в Wayback Machine.
- ^ Фокин С.И. (2013). "Отто Бютчли (1848–1920) Где мы преклоним колени?" (PDF). Протистология. 8 (1): 22–35. В архиве (PDF) из оригинала на 2014-08-08. Получено 2014-08-06.
- ^ Sharp LW (1921). Введение в цитологию. Нью-Йорк: McGraw Hill Book Company Inc., стр. 143.
- ^ "митоз". Интернет-словарь этимологии. В архиве из оригинала от 28.09.2017. Получено 2019-11-12.
- ^ μίτος. Лидделл, Генри Джордж; Скотт, Роберт; Греко-английский лексикон на Проект Персей
- ^ Батталья Э (2009). «Карионема, альтернатива хромосоме и новая кариологическая номенклатура» (PDF). Кариология. 62 (4): 1–80. Архивировано из оригинал (PDF) на 2016-03-04.
- ^ Шлейхер W (1878). "Die Knorpelzelltheilung". Arch. Миркроскоп. Анат. 16: 248–300. Архивировано из оригинал на 2018-08-11.
- ^ Топфер Г. «Кариокинез». БиоКонцепты. Архивировано из оригинал на 2018-05-03. Получено 2 мая 2018.
- ^ Батталья Э (1987). «Эмбриологические вопросы: 12. Имейте Полигонум и Allium типы были правильно установлены? ». Энн Бот. Рим. 45: 81–117.
п. 85: Уже в 1887 году Вейсманн дал названия Aequationstheilung к обычному делению клеток, и Reduktionstheilungen двум подразделениям, участвующим в процессе сокращения вдвое количества Kernsegmente
- ^ Маузет Дж. Д. (1991). Ботаника: введение в биологию растений. Филадельфия: Издательство Saunders College.
п. 102: деление клетки - это цитокинез, а деление ядра - это кариокинез. Слова «митоз» и «мейоз» технически относятся только к кариокинезу, но также часто используются для описания цитокинеза.
- ^ Купер, Джеффри М. (2000). «Мейоз и оплодотворение». Клетка: молекулярный подход. 2-е издание.
- ^ Браун, Теренс А. (2002). Геном человека. Wiley-Liss.
- ^ а б Блоу Джей Джей, Танака Т.У. (ноябрь 2005 г.). «Хромосомный цикл: координация репликации и сегрегации. Второй в серии обзоров циклов». EMBO отчеты. 6 (11): 1028–34. Дои:10.1038 / sj.embor.7400557. ЧВК 1371039. PMID 16264427.
- ^ Чжоу Дж, Яо Дж, Джоши ХК (сентябрь 2002 г.). «Крепление и натяжение в КПП шпиндельного узла». Журнал клеточной науки. 115 (Pt 18): 3547–55. Дои:10.1242 / jcs.00029. PMID 12186941.
- ^ Биология онлайн. "Митоз". Биология онлайн.
- ^ Шибата А. (2017). «Регуляция выбора пути репарации при двухцепочечных разрывах ДНК». Mutat Res. 803-805: 51–55. Дои:10.1016 / j.mrfmmm.2017.07.011. PMID 28781144.
- ^ «Инъекция антицентромерных антител в интерфазе нарушает события, необходимые для движения хромосом при митозе». Журнал клеточной биологии. 111 (4): 1519–1533. 1990-10-01. ISSN 0021-9525. ЧВК 2116233. PMID 2211824.
- ^ а б Ллойд С., Чан Дж. (Февраль 2006 г.). «Не так поделено: общая основа деления клеток растений и животных». Обзоры природы. Молекулярная клеточная биология. 7 (2): 147–52. Дои:10.1038 / nrm1831. PMID 16493420. S2CID 7895964.
- ^ а б Ворон PH, Evert RF, Eichhorn SE (2005). Биология растений (7-е изд.). Нью-Йорк: W.H. Freeman and Co. ISBN 978-0716710073.
- ^ Прасант К.В., Сакко-Бубуля ПА, Прасант С.Г., Спектор Д.Л. (март 2003 г.). «Последовательное вхождение компонентов машины экспрессии генов в дочерние ядра». Молекулярная биология клетки. 14 (3): 1043–57. Дои:10.1091 / mbc.E02-10-0669. ЧВК 151578. PMID 12631722.
- ^ Кадауке С., Блобель Г.А. (апрель 2013 г.). «Митотическая закладка по факторам транскрипции». Эпигенетика и хроматин. 6 (1): 6. Дои:10.1186/1756-8935-6-6. ЧВК 3621617. PMID 23547918.
- ^ Прескотт Д.М., Бендер М.А. (март 1962 г.). «Синтез РНК и белка во время митоза в клетках культуры ткани млекопитающих». Экспериментальные исследования клеток. 26 (2): 260–8. Дои:10.1016/0014-4827(62)90176-3. PMID 14488623.
- ^ Олсон МО (2011). Ядрышко. Том 15 Обзоров Белка. Берлин: Springer Science & Business Media. п. 15. ISBN 9781461405146.
- ^ Басто Р., Лау Дж., Виноградова Т., Гардиол А., Вудс К. Г., Ходжаков А., Рафф Дж. В. (июнь 2006 г.). «Летает без центриолей». Клетка. 125 (7): 1375–86. Дои:10.1016 / j.cell.2006.05.025. PMID 16814722. S2CID 2080684.
- ^ Хейвуд П. (июнь 1978 г.). «Ультраструктура митоза в водоросли хлоромонадофид Vacuolaria virescens». Журнал клеточной науки. 31: 37–51. PMID 670329.
- ^ Рибейро К.С., Перейра-Невес А., Бенчимол М. (июнь 2002 г.). «Митотическое веретено и связанные мембраны в закрытом митозе трихомонад». Биология клетки. 94 (3): 157–72. Дои:10.1016 / S0248-4900 (02) 01191-7. PMID 12206655.
- ^ а б Чан Г.К., Лю С.Т., Йен Т.Дж. (ноябрь 2005 г.). «Строение и функции кинетохор». Тенденции в клеточной биологии. 15 (11): 589–98. Дои:10.1016 / j.tcb.2005.09.010. PMID 16214339.
- ^ Чизмен И.М., Десаи А. (январь 2008 г.). «Молекулярная архитектура интерфейса кинетохора-микротрубочка». Обзоры природы. Молекулярная клеточная биология. 9 (1): 33–46. Дои:10.1038 / nrm2310. PMID 18097444. S2CID 34121605.
- ^ а б Winey M, Mamay CL, O'Toole ET, Mastronarde DN, Giddings TH, McDonald KL, McIntosh JR (июнь 1995 г.). «Трехмерный ультраструктурный анализ митотического веретена Saccharomyces cerevisiae». Журнал клеточной биологии. 129 (6): 1601–15. Дои:10.1083 / jcb.129.6.1601. ЧВК 2291174. PMID 7790357.
- ^ а б Maiato H, DeLuca J, Salmon ED, Earnshaw WC (ноябрь 2004 г.). «Динамический интерфейс кинетохора-микротрубочка» (PDF). Журнал клеточной науки. 117 (Pt 23): 5461–77. Дои:10.1242 / jcs.01536. PMID 15509863. S2CID 13939431. В архиве (PDF) из оригинала от 18.08.2017. Получено 2018-04-20.
- ^ Чан Г.К., Йен Т.Дж. (2003). «Контрольная точка митоза: сигнальный путь, который позволяет одиночной неприсоединенной кинетохоре ингибировать выход из митоза». Прогресс в исследованиях клеточного цикла. 5: 431–9. PMID 14593737.
- ^ а б ФитцХаррис Дж. (Март 2012 г.). «Анафаза B в яйце мыши предшествует анафазе A» (PDF). Текущая биология. 22 (5): 437–44. Дои:10.1016 / j.cub.2012.01.041. PMID 22342753. В архиве (PDF) из оригинала на 2018-07-24. Получено 2019-09-17.
- ^ Миллер К. Р., Левин Дж. (2000). «Анафаза». Биология (5-е изд.). Пирсон Прентис Холл. стр.169 –70. ISBN 978-0-13-436265-6.
- ^ Европейская лаборатория молекулярной биологии (12 июня 2007 г.). «Конденсация хромосом через митоз». Science Daily. В архиве из оригинала 13 июня 2007 г.. Получено 4 октября 2020.
- ^ Глотцер М (март 2005 г.). «Молекулярные требования для цитокинеза». Наука. 307 (5716): 1735–9. Bibcode:2005Наука ... 307.1735G. Дои:10.1126 / science.1096896. PMID 15774750. S2CID 34537906.
- ^ Альбертсон Р., Риггс Б., Салливан В. (февраль 2005 г.). «Мембранный трафик: движущая сила цитокинеза». Тенденции в клеточной биологии. 15 (2): 92–101. Дои:10.1016 / j.tcb.2004.12.008. PMID 15695096.
- ^ а б Лилли М.А., Дуронио Р.Дж. (апрель 2005 г.). «Новые данные о контроле клеточного цикла с помощью эндоцикла дрозофилы». Онкоген. 24 (17): 2765–75. Дои:10.1038 / sj.onc.1208610. PMID 15838513.
- ^ Бетчер Б., Баррал И. (2013). «Клеточная биология открытого и закрытого митоза». Ядро. 4 (3): 160–5. Дои:10.4161 / nucl.24676. ЧВК 3720745. PMID 23644379.
- ^ Р. Десаль, Б. Шируотер: Ключевые переходы в эволюции животных. CRC Press, 2010, стр. 12, связь В архиве 2019-01-02 в Wayback Machine.
- ^ Мантику Э., Вонг К.М., Реппинг С., Мастенбрук С. (декабрь 2012 г.). «Молекулярное происхождение митотических анеуплоидий у преимплантационных эмбрионов». Biochimica et Biophysica Acta (BBA) - Молекулярная основа болезни. 1822 (12): 1921–30. Дои:10.1016 / j.bbadis.2012.06.013. PMID 22771499.
- ^ Вассманн, Катя; Бенезра, Роберт (2001-02-01). «Контрольные точки митоза: от дрожжей до рака». Текущее мнение в области генетики и развития. 11 (1): 83–90. Дои:10.1016 / S0959-437X (00) 00161-1. ISSN 0959-437X.
- ^ Дравиам В.М., Се С., Соргер П.К. (апрель 2004 г.). «Хромосомная сегрегация и геномная стабильность». Текущее мнение в области генетики и развития. 14 (2): 120–5. Дои:10.1016 / j.gde.2004.02.007. PMID 15196457.
- ^ Сантагуида С., Амон А. (август 2015 г.). «Краткосрочные и долгосрочные последствия неправильной сегрегации хромосом и анеуплоидии». Обзоры природы. Молекулярная клеточная биология. 16 (8): 473–85. Дои:10.1038 / nrm4025. HDL:1721.1/117201. PMID 26204159. S2CID 205495880.
- ^ а б c Юров И.Ю., Ворсанова С.Г., Юров Ю.Б. (2006). «Хромосомные вариации в нейронных клетках млекопитающих: известные факты и привлекательные гипотезы». В Jeon KJ (ред.). Международный обзор цитологии: обзор клеточной биологии. 249. Уолтем, Массачусетс: Academic Press. п. 146. ISBN 9780080463506.
- ^ Ши Кью, король Р.В. (октябрь 2005 г.). «Нерасхождение хромосом дает тетраплоидные, а не анеуплоидные клетки в линиях клеток человека». Природа. 437 (7061): 1038–42. Bibcode:2005Натура.437.1038S. Дои:10.1038 / природа03958. PMID 16222248. S2CID 1093265.
- ^ а б Эдгар Б.А., Орр-Уивер Т.Л. (май 2001 г.). «Циклы эндорепликационных клеток: больше за меньшие деньги». Клетка. 105 (3): 297–306. Дои:10.1016 / S0092-8674 (01) 00334-8. PMID 11348589. S2CID 14368177.
- ^ а б Ли Хо, Дэвидсон Дж. М., Дуронио Р. Дж. (Ноябрь 2009 г.). «Эндорепликация: полиплоидия с целью». Гены и развитие. 23 (21): 2461–77. Дои:10.1101 / gad.1829209. ЧВК 2779750. PMID 19884253.
- ^ Итальяно Дж. Э., Шивдасани Р. А. (июнь 2003 г.). «Мегакариоциты и не только: рождение тромбоцитов». Журнал тромбоза и гемостаза. 1 (6): 1174–82. Дои:10.1046 / j.1538-7836.2003.00290.x. PMID 12871316.
- ^ Витрат Н., Коэн-Солаль К., Пике С., Ле Кудик Дж. П., Норол Ф., Ларсен А. К., Кац А., Вайнченкер В., Дебили Н. (май 1998 г.). «Эндомитоз мегакариоцитов человека происходит из-за абортивного митоза». Кровь. 91 (10): 3711–23. Дои:10.1182 / blood.V91.10.3711. PMID 9573008.
- ^ «Инфильтрирующая протоковая карцинома груди (карцинома без особого типа)». Медицинский факультет Стэнфордского университета. В архиве из оригинала на 2019-09-11. Получено 2019-10-02.
- ^ Bertram CA, Aubreville M, Gurtner C, Bartel A, Corner SM, Dettwiler M и др. (Март 2020 г.). «Компьютеризированный расчет распределения количества митозов в срезах опухоли тучных клеток кожи собаки: количество митозов зависит от площади» (PDF). Ветеринарная патология. 57 (2): 214–226. Дои:10.1177/0300985819890686. PMID 31808382. S2CID 208767801.
- ^ Зауэр ФК (1935). «Митоз в нервной трубке». Журнал сравнительной неврологии. 62 (2): 377–405. Дои:10.1002 / cne.900620207.
- ^ а б Мейер Э.Дж., Икми А., Гибсон М.С. (март 2011 г.). «Межкинетическая миграция ядер - широко консервативный признак деления клеток в псевдостратифицированном эпителии». Текущая биология. 21 (6): 485–91. Дои:10.1016 / j.cub.2011.02.002. PMID 21376598. В архиве из оригинала на 2013-10-08. Получено 2015-05-10.
- ^ а б Люксенбург C, Пазолли HA, Williams SE, Fuchs E (март 2011 г.). «Роли развития Srf, кортикального цитоскелета и формы клеток в ориентации веретена эпидермиса». Природа клеточной биологии. 13 (3): 203–14. Дои:10.1038 / Ncb2163. ЧВК 3278337. PMID 21336301.
- ^ Накадзима Ю., Мейер Э.Д., Крузен А., МакКинни С.А., Гибсон М.С. (август 2013 г.). «Эпителиальные соединения поддерживают архитектуру ткани, направляя планарную ориентацию веретена». Природа. 500 (7462): 359–62. Bibcode:2013Натура.500..359Н. Дои:10.1038 / природа12335. PMID 23873041. S2CID 4418619.
- ^ а б Cadart C, Zlotek-Zlotkiewicz E, Le Berre M, Piel M, Matthews HK (апрель 2014 г.). «Изучение функции формы и размера клеток во время митоза». Клетка развития. 29 (2): 159–69. Дои:10.1016 / j.devcel.2014.04.009. PMID 24780736.
- ^ Ланкастер О.М., Ле Берр М., Димитракопулос А., Бонацци Д., Злотек-Злоткевич Э., Пиконе Р., Дюк Т., Пиль М., Баум Б. (май 2013 г.). «Митотическое округление изменяет геометрию клетки, чтобы обеспечить эффективное формирование биполярного веретена». Клетка развития. 25 (3): 270–83. Дои:10.1016 / j.devcel.2013.03.014. PMID 23623611.
- ^ а б Ланкастер О.М., Баум Б. (октябрь 2014 г.). «Формирование для разделения: координация актина и ремоделирования цитоскелета микротрубочек во время митоза». Семинары по клеточной биологии и биологии развития. 34: 109–15. Дои:10.1016 / j.semcdb.2014.02.015. PMID 24607328.
- ^ а б c Мэддокс А.С., Берридж К. (январь 2003 г.). «RhoA необходим для ретракции коры и жесткости во время округления митотических клеток». Журнал клеточной биологии. 160 (2): 255–65. Дои:10.1083 / jcb.200207130. ЧВК 2172639. PMID 12538643.
- ^ а б Мэтьюз Х. К., Делабре Ю., Рон Дж. Л., Гук Дж., Кунда П., Баум Б. (август 2012 г.). "Изменения в локализации Ect2 пара актомиозин-зависимых клеточных изменений в митотической прогрессии". Клетка развития. 23 (2): 371–83. Дои:10.1016 / j.devcel.2012.06.003. ЧВК 3763371. PMID 22898780.
- ^ а б Стюарт М.П., Хелениус Дж., Тойода Ю., Раманатан С.П., Мюллер Д.Д., Хайман А.А. (январь 2011 г.). «Гидростатическое давление и актомиозиновая кора приводят к округлению митотических клеток». Природа. 469 (7329): 226–30. Bibcode:2011Натура.469..226S. Дои:10.1038 / природа09642. PMID 21196934. S2CID 4425308.
- ^ Фишер-Фридрих Э., Хайман А.А., Юлихер Ф., Мюллер Д.Д., Хелениус Дж. (Август 2014 г.). «Количественная оценка поверхностного натяжения и внутреннего давления, создаваемого отдельными митотическими клетками». Научные отчеты. 4 (6213): 6213. Bibcode:2014НатСР ... 4Э6213Ф. Дои:10.1038 / srep06213. ЧВК 4148660. PMID 25169063.
- ^ а б c d Раманатан С.П., Хелениус Дж., Стюарт М.П., Кэттин С.Дж., Хайман А.А., Мюллер Д.Д. (февраль 2015 г.). «Cdk1-зависимое митотическое обогащение кортикального миозина II способствует округлению клеток против ограничения». Природа клеточной биологии. 17 (2): 148–59. Дои:10.1038 / ncb3098. PMID 25621953. S2CID 5208968.
- ^ а б Кадык Л.С., Хартвелл Л.Х. (октябрь 1992 г.). «Сестринские хроматиды предпочтительнее гомологов в качестве субстратов для рекомбинационной репарации у Saccharomyces cerevisiae». Генетика. 132 (2): 387–402. ЧВК 1205144. PMID 1427035.
- ^ Botthof JG, Bielczyk-Maczyńska E, Ferreira L, Cvejic A (май 2017 г.). "rad51 приводит к симптомам анемии Фанкони у рыбок данио". Труды Национальной академии наук Соединенных Штатов Америки. 114 (22): E4452 – E4461. Дои:10.1073 / pnas.1620631114. ЧВК 5465903. PMID 28512217.
Здесь мы приводим доказательства in vivo того, что уменьшение количества HSPC у взрослых рыб действительно происходит из-за комбинации снижения пролиферации и увеличения апоптоза во время эмбрионального развития. Этот дефект, по-видимому, опосредуется через p53 (10), поскольку наши двойные мутанты p53 / rad51 не обнаруживают каких-либо наблюдаемых гематологических дефектов у эмбрионов или взрослых.
- ^ Stürzbecher HW, Donzelmann B, Henning W., Knippschild U, Buchhop S (апрель 1996 г.). «p53 напрямую связан с процессами гомологичной рекомбинации через взаимодействие белков RAD51 / RecA». Журнал EMBO. 15 (8): 1992–2002. Дои:10.1002 / j.1460-2075.1996.tb00550.x. ЧВК 450118. PMID 8617246.
- ^ Сонода Э., Сасаки М.С., Бюрстедде Дж. М., Беззубова О., Шинохара А., Огава Х. и др. (Январь 1998 г.). «Клетки позвоночных с дефицитом Rad51 накапливают хромосомные разрывы до гибели клеток». Журнал EMBO. 17 (2): 598–608. Дои:10.1093 / emboj / 17.2.598. ЧВК 1170409. PMID 9430650.
- ^ Уилкинс А.С., Холлидей Р. (январь 2009 г.). «Эволюция мейоза из митоза». Генетика. 181 (1): 3–12. Дои:10.1534 / генетика.108.099762. ЧВК 2621177. PMID 19139151.
- ^ Бернштейн, Х., Бернштейн, С. Эволюционное происхождение и адаптивная функция мейоза. В «Meiosis», Intech Publ (редакторы Кэрол Бернстайн и Харрис Бернстайн), Глава 3: 41-75 (2013).
дальнейшее чтение
- Морган DL (2007). Клеточный цикл: принципы контроля. Лондон: Издается New Science Press совместно с Oxford University Press. ISBN 978-0-9539181-2-6.
- Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). "Митоз". Молекулярная биология клетки (4-е изд.). Наука о гирляндах. Получено 2006-01-22.
- Кэмпбелл Н., Рис Дж. (Декабрь 2001 г.). «Клеточный цикл». Биология (6-е изд.). Сан-Франциско: Бенджамин Каммингс / Аддисон-Уэсли. стр.217–224. ISBN 978-0-8053-6624-2.
- Купер Дж. (2000). «События фазы М». Клетка: молекулярный подход (2-е изд.). Sinaeur Associates, Inc. Получено 2006-01-22.
- Фримен С (2002). «Подразделение клеток». Биологические науки. Река Аппер Сэдл, Нью-Джерси: Prentice Hall. стр.155–174. ISBN 978-0-13-081923-9.
- Лодиш Х, Берк А., Зипурски Л., Мацудаира П., Балтимор Д., Дарнелл Дж. (2000). «Обзор клеточного цикла и его контроля». Молекулярная клеточная биология (4-е изд.). В. Х. Фриман. Получено 2006-01-22.
внешняя ссылка
- Flash-анимация, сравнивающая митоз и мейоз
- Ханская академия, лекция
- Изучение митоза в культивируемых клетках млекопитающих
- Общие ресурсы класса K-12 для лечения митоза
- Онтология клеточного цикла
- WormWeb.org: интерактивная визуализация C. elegans Клеточная линия - Визуализируйте все дерево происхождения клеток и все деления клеток нематоды C. elegans