Деление клеток - Cell division

Деление клеток это процесс, с помощью которого родитель клетка делится на две и более дочерних клетки.[1] Деление клеток обычно происходит как часть более крупного клеточный цикл. В эукариоты, есть два различных типа клеточного деления; вегетативное деление, при котором каждая дочерняя клетка генетически идентична родительской клетке (митоз ) и деление репродуктивной клетки, в результате чего количество хромосомы в дочерних клетках уменьшается вдвое, чтобы произвести гаплоид гаметы (мейоз ).[2] В клеточная биология, митоз (/ maɪˈtoʊsɪs /) является частью клеточный цикл, в котором воспроизведены хромосомы разделяются на два новых ядра. Деление клеток приводит к образованию генетически идентичных клеток, в которых сохраняется общее количество хромосом. В целом митозу (делению ядра) предшествует S стадия межфазный (во время которого ДНК реплицируется) и часто сопровождается телофаза и цитокинез; что делит цитоплазма, органеллы и клеточная мембрана из одной ячейки в две новые клетки содержащие примерно равные доли этих клеточных компонентов. Различные стадии митоза вместе определяют митотический (M) фаза цикла животных клеток - разделение материнской клетки на две дочерние клетки, генетически идентичные друг другу[нужна цитата ]. Мейоз приводит к появлению четырех гаплоидных дочерних клеток в результате одного раунда репликации ДНК, за которым следуют два деления. Гомологичные хромосомы разделены в первом делении, а сестринские хроматиды разделены во втором делении. Оба этих цикла деления клеток используются в процессе полового размножения в какой-то момент своего жизненного цикла. Считается, что оба они присутствовали в последнем общем предке эукариот.
Прокариоты (бактерии и археи ) обычно подвергаются делению вегетативных клеток, известному как двойное деление, где их генетический материал разделен поровну на две дочерние клетки. Хотя бинарное деление может быть средством деления большинства прокариот, существуют альтернативные способы деления, такие как подающий надежды, которые наблюдались. Всем клеточным делениям, независимо от организма, предшествует один раунд Репликация ДНК.
Для простого одноклеточный микроорганизмы такой как амеба, одно деление клетки эквивалентно воспроизведение - создается совершенно новый организм. В более крупном масштабе деление митотических клеток может создавать потомство от многоклеточных организмов, например растений, вырастающих из черенков. Деление митотических клеток позволяет половым путем организмов развиваться из одноклеточных зигота, который сам был произведен мейотическим делением клеток из гаметы.[3][4] После роста клеточное деление путем митоза позволяет непрерывно строить и восстанавливать организм.[5] Человеческое тело испытывает около 10 квадриллион деления клеток в течение жизни.[6]
Первоочередная задача клеточного деления - поддержание исходной клеточной геном. Прежде чем может произойти деление, геномная информация, которая хранится в хромосомах, должна быть реплицирована, и дублированный геном должен быть четко разделен между клетками.[7] Большая часть клеточной инфраструктуры участвует в поддержании согласованности геномной информации между поколениями.
Деление клеток у бактерий
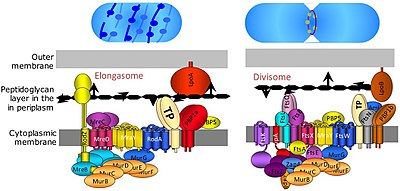
Деление бактериальной клетки происходит посредством бинарного деления или почкования. В Divisome представляет собой белковый комплекс бактерий, который отвечает за деление клеток, сужение внутренней и внешней мембран во время деления и синтез пептидогликана (PG) в месте деления. Тубулиноподобный белок, FtsZ играет решающую роль в формировании сократительного кольца для деления клеток. [9]
Деление клеток у эукариот
Деление клеток у эукариотов намного сложнее, чем у прокариот. В зависимости от количества хромосом уменьшено или нет; Деления эукариотических клеток можно классифицировать как Митоз (эквациональное деление) и Мейоз (редукционное деление). Встречается также премитивная форма деления клеток, которая называется амитоз. Амитотическое или митотическое деление клеток более нетипично и разнообразно у различных групп организмов, таких как простейшие (а именно диатомеи, динофлагелляты и т. Д.) И грибы.
- Формы митоза (ступени кариокинеза) у эукариот

закрыто
внутриядерный
плевромитоз
закрыто
внеядерный
плевромитоз
закрыто
ортомитоз
полуоткрытый
плевромитоз
полуоткрытый
ортомитоз
открыто
ортомитоз
В митотической метафазе (см. Ниже) обычно хромосомы (каждая с двумя сестринскими хроматидами, которые они развили в результате репликации в S-фазе интерфазы) располагаются, а сестринские хроматиды расщепляются и распределяются по дочерним клеткам.
В мейозе, обычно в мейозе-I, гомологичные хромосомы спариваются, а затем разделяются и распределяются по дочерним клеткам. Мейоз-II подобен митозу, в котором хроматиды разделены. У человека и других высших животных и многих других организмов мейоз называется гаметным мейозом, то есть мейоз дает начало гаметам. В то время как у многих групп организмов, особенно у растений (наблюдаемых у низших растений, но рудиментарная стадия у высших растений), мейоз дает начало видам спор, которые прорастают в гаплоидную вегетативную фазу (гаметофит). Этот вид мейоза называется споровым мейозом.
Фазы деления эукариотических клеток
Эта статья слишком полагается на Рекомендации к основные источники. (Июль 2017 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |
Межфазный
Межфазный это процесс, через который клетка должна пройти перед митозом, мейозом и цитокинез.[10] Интерфаза состоит из трех основных фаз: грамм1, S, и грамм2. грамм1 это время роста клетки, когда выполняются специализированные клеточные функции, чтобы подготовить клетку к репликации ДНК.[11] Во время интерфазы есть контрольные точки, которые позволяют клетке либо продвигаться, либо останавливать дальнейшее развитие. В S-фазе хромосомы реплицируются для сохранения генетического содержимого.[12] Во время G2, клетка проходит заключительную стадию роста перед переходом в фазу М, где шпиндели синтезированы. Фаза M может быть митозом или мейозом в зависимости от типа клетки. Стволовые клетки, или гаметы, претерпевают мейоз, а соматические клетки подвергнется митозу. После того, как клетка успешно пройдет через М-фазу, она может подвергнуться клеточному делению посредством цитокинеза. Контроль каждого КПП контролируется циклин и циклин-зависимые киназы. Развитие интерфазы является результатом увеличения количества циклина. По мере увеличения количества циклина все больше и больше циклинзависимых киназ присоединяются к циклину, передавая сигнал клетке дальше в интерфазу. На пике циклина, связанного с циклинзависимыми киназами, эта система выталкивает клетку из интерфазы в фазу М, где происходят митоз, мейоз и цитокинез.[13] Есть три контрольных точки перехода, которые клетка должна пройти перед переходом в фазу M. Самый важный из них - G1-S переходная контрольная точка. Если клетка не проходит эту контрольную точку, это приводит к выходу клетки из клеточного цикла.[14]
Профаза
Профаза это первый этап деления. На этом этапе ядерная оболочка разрушается, длинные нити хроматина конденсируются, образуя более короткие и более заметные нити, называемые хромосомами, ядрышко исчезает, а микротрубочки прикрепляются к хромосомам в дискообразной области. кинетохоры присутствует в центромере.[15] Микротрубочки, связанные с выравниванием и разделением хромосом, называются веретенообразными и веретеновыми волокнами. Хромосомы также будут видны под микроскопом и будут соединены в центромере. Во время этого периода конденсации и выравнивания в мейозе гомологичные хромосомы претерпевают разрыв в своей двухцепочечной ДНК в одних и тех же местах, за которым следует рекомбинация уже фрагментированных родительских цепей ДНК в не-родительские комбинации, известные как кроссинговер.[16] Доказано, что этот процесс в значительной степени вызван высококонсервативными Spo11 белок по механизму, подобному тому, что наблюдается с топосомераза в репликации и транскрипции ДНК.[17]
Метафаза
В метафаза, то центромеры хромосом собираются на метафазная пластинка (или же экваториальная плита), воображаемая линия, которая находится на равном расстоянии от двух центросома полюса и удерживаются вместе сложными комплексами, известными как когезины. Хромосомы выстраиваются в середине клетки за счет центры организации микротрубочек (MTOC) толкают и притягивают центромеры обеих хроматид, заставляя хромосому перемещаться к центру. На данный момент хромосомы все еще конденсируются и в настоящее время находятся в одном шаге от того, чтобы быть наиболее свернутыми и конденсированными, как они будут, а волокна веретена уже соединены с кинетохорами.[18] Во время этой фазы все микротрубочки, за исключением кинетохор, находятся в состоянии нестабильности, способствуя их продвижению к анафазе.[19] В этот момент хромосомы готовы разделиться на противоположные полюса клетки по отношению к веретену, с которым они связаны.[20]
Анафаза
Анафаза это очень короткая стадия клеточного цикла, которая происходит после выравнивания хромосом в митотической пластинке. Кинетохоры испускают сигналы ингибирования анафазы до момента их прикрепления к митотическому веретену. Как только последняя хромосома правильно выровнена и прикреплена, последний сигнал рассеивается и вызывает резкий переход к анафазе.[19] Этот резкий сдвиг вызван активацией комплекс, способствующий анафазе и его функция маркировки деградации белков, важных для перехода от метафазы к анафазе. Один из этих белков, который расщепляется, - Securin который при его распаде высвобождает фермент отделить который расщепляет когезиновые кольца, удерживающие вместе сестринские хроматиды, тем самым приводя к разделению хромосом.[21] После того, как хромосомы выстроятся в одну линию в середине клетки, волокна веретена разорвут их. Хромосомы разделяются, а сестринские хроматиды перемещаются в противоположные стороны клетки.[22] По мере того как сестринские хроматиды раздвигаются, клетка и плазма удлиняются за счет некинетохорных микротрубочек.[23]
Телофаза
Телофаза это последняя стадия клеточного цикла, в которой борозда расщепления расщепляет цитоплазму клеток (цитокинез) и хроматин. Это происходит за счет синтеза новых ядерных оболочек, которые формируются вокруг хроматина, который собирается на каждом полюсе, и реформирования ядрышка, когда хромосомы возвращают свой хроматин обратно в рыхлое состояние, которым он обладал во время интерфазы.[24][25] Разделение клеточного содержимого не всегда одинаково и может варьироваться в зависимости от типа клеток, как видно на примере образования ооцитов, когда одна из четырех дочерних клеток обладает большей частью цитоплазмы.[26]
Цитокинез
Последний этап процесса деления клеток - это цитокинез. На этой стадии происходит деление цитоплазмы в конце митоза или мейоза. На этом этапе происходит необратимое разделение, ведущее к двум дочерним клеткам. Деление клетки играет важную роль в определении судьбы клетки. Это связано с возможностью асимметричного разделения. В результате это приводит к цитокинезу, продуцирующему неравные дочерние клетки, содержащие совершенно разные количества или концентрации определяющих судьбу молекул.[27]
У животных цитокинез заканчивается образованием сократительного кольца, а затем и расщеплением. Но у растений бывает иначе. Сначала образуется клеточная пластинка, а затем между двумя дочерними клетками развивается клеточная стенка.
В делящихся дрожжах (С. Помбе ) цитокинез происходит в фазе G1 [28]
Варианты
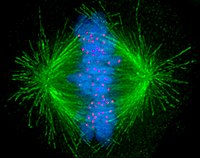
Клетки в общих чертах подразделяются на две основные категории: простые без зародышей прокариотический клетки и комплекс ядерные эукариотический клетки. Из-за их структурных различий эукариотические и прокариотические клетки не делятся одинаково. Кроме того, паттерн деления клеток, трансформирующий эукариотические клетки стволовые клетки в гаметы (сперма клетки у мужчин или яйцо клеток у самок), называемый мейозом, отличается от деления соматический клетки в теле. Изображение митотическое веретено в клетке человека микротрубочки показаны зеленым, хромосомы (ДНК) - синим, а кинетохоры - красным.
Деградация
Многоклеточные организмы заменяют изношенные клетки путем деления клеток. Однако у некоторых животных деление клеток со временем останавливается. В люди это происходит в среднем после 52 делений, известных как Лимит Хейфлика. Тогда ячейка называется дряхлый. С каждым делением клетки теломеры, защитные последовательности ДНК на конце хромосома которые предотвращают деградацию хромосомной ДНК, сокращать. Это сокращение коррелирует с негативными эффектами, такими как возрастные заболевания и сокращение продолжительности жизни у людей.[30][31] Рак клетки, с другой стороны, не думают, что деградируют таким образом, если вообще. An фермент комплекс называется теломераза, присутствующий в больших количествах в раковых клетках, восстанавливает теломеры за счет синтеза теломерных повторов ДНК, позволяя делению продолжаться бесконечно.[32]
История

Деление клетки под микроскоп был впервые обнаружен немецким ботаником Хьюго фон Моль в 1835 году, когда он работал над зеленая водоросль Кладофора клубочки.[33]
В 1943 году впервые было заснято деление клеток.[34] к Курт Мишель используя фазово-контрастный микроскоп.[35]
Смотрите также
- Двойное деление
- Клеточная биология
- Слияние клеток
- Рост клеток
- Циклинзависимая киназа
- Лабильные клетки, клетки, которые постоянно делятся
Рекомендации
- ^ Мартин Э.А., Хайн Р. (2020). Словарь биологии (6-е изд.). Оксфорд: Издательство Оксфордского университета. ISBN 9780199204625. OCLC 176818780.
- ^ Гриффитс AJ (2012). Введение в генетический анализ (10-е изд.). Нью-Йорк: W.H. Фриман и Ко. ISBN 9781429229432. OCLC 698085201.
- ^ Гилберт С.Ф. (2000). «Сперматогенез». Биология развития (6-е изд.).
- ^ Гилберт С.Ф. (2000). «Оогенез». Биология развития (6-е изд.).
- ^ Матон, Антея. (1997). Клетки: строительные блоки жизни (3-е изд.). Река Аппер Сэдл, Нью-Джерси: Прентис-Холл. стр.70–74. ISBN 978-0134234762. OCLC 37049921.
- ^ Quammen D (апрель 2008 г.). «Заразный рак». Журнал Harper's. ISSN 0017-789X. Получено 2019-04-14.
- ^ Голицын, Юрий Н .; Крылов, Михаил С. (2010). Деление клетки: теория, варианты и деградация. Нью-Йорк: Nova Science Publishers. п. 137. ISBN 9781611225938. OCLC 669515286.
- ^ Hugonnet, Жан-Эммануэль; Mengin-Lecreulx, Dominique; Монтон, Алехандро; Blaauwen, Tanneke den; Карбоннель, Этьен; Векерле, Кэрол; Ив; Brun, V .; Nieuwenhze, Майкл ван (2016-10-21). «Факторы, существенные для опосредованного L, D-транспептидазой сшивания пептидогликанов и устойчивости к β-лактамам у Escherichia coli». eLife. 5. Дои:10.7554 / elife.19469. ISSN 2050-084X. ЧВК 5089857. PMID 27767957.
- ^ Подразделение клеток: Цикл кольца, Лоуренс Ротфилд и Шерил Джастис, КЛЕТКА, DOI
- ^ Мариеб EN (2000). Основы анатомии и физиологии человека (6-е изд.). Сан-Франциско: Бенджамин Каммингс. ISBN 978-0805349405. OCLC 41266267.
- ^ Парди AB (ноябрь 1989 г.). «События G1 и регуляция пролиферации клеток». Наука. 246 (4930): 603–8. Bibcode:1989Научный ... 246..603P. Дои:10.1126 / science.2683075. PMID 2683075.
- ^ Морган Д.О. (2007). Клеточный цикл: принципы контроля. Лондон: New Science Press. ISBN 9780199206100. OCLC 70173205.
- ^ Линдквист А., Ван Зон В., Карлссон Розенталь С., Вольтуис Р.М. (май 2007 г.). «Активация циклина B1-Cdk1 продолжается после разделения центросом для контроля митотической прогрессии». PLOS Биология. 5 (5): e123. Дои:10.1371 / journal.pbio.0050123. ЧВК 1858714. PMID 17472438.
- ^ Паулович А.Г., Точиски Д.П., Хартвелл Л.Х. (февраль 1997 г.). «Когда пропускные пункты выходят из строя». Клетка. 88 (3): 315–21. Дои:10.1016 / S0092-8674 (00) 81870-X. PMID 9039258. S2CID 5530166.
- ^ Schermelleh L, Carlton PM, Haase S, Shao L, Winoto L, Kner P, Burke B, Cardoso MC, Agard DA, Gustafsson MG, Leonhardt H, Sedat JW (июнь 2008 г.). «Субдифракционная многоцветная визуализация периферии ядра с помощью микроскопии со структурированным освещением в 3D». Наука. 320 (5881): 1332–6. Bibcode:2008Научный ... 320.1332S. Дои:10.1126 / science.1156947. ЧВК 2916659. PMID 18535242.
- ^ Левонтин Р.К., Миллер Дж. Х., Гелбарт В. М., Гриффитс А. Дж. (1999). «Механизм перехода». Современный генетический анализ.
- ^ Кини С (2001). Механизм и контроль инициации мейотической рекомбинации. Актуальные темы биологии развития. 52. Эльзевир. стр.1–53. Дои:10.1016 / с0070-2153 (01) 52008-6. ISBN 9780121531522. PMID 11529427.
- ^ «Исследователи пролили свет на уменьшение размеров хромосом». ScienceDaily. Получено 2019-04-14.
- ^ а б Уолтер П., Робертс К., Рафф М., Льюис Дж., Джонсон А., Альбертс Б. (2002). "Митоз". Молекулярная биология клетки (4-е изд.).
- ^ Элрод С (2010). Очертания Шаума: генетика (5-е изд.). Нью-Йорк: Макгроу-Хилл. п. 8. ISBN 9780071625036. OCLC 473440643.
- ^ Брукер А.С., Берковиц К.М. (2014). «Роль когезинов в митозе, мейозе, здоровье и болезни человека». Контроль клеточного цикла. Методы молекулярной биологии. 1170. Нью-Йорк: Спрингер. С. 229–66. Дои:10.1007/978-1-4939-0888-2_11. ISBN 9781493908875. ЧВК 4495907. PMID 24906316.
- ^ «Клеточный цикл». www.biology-pages.info. Получено 2019-04-14.
- ^ Урри, Лиза А .; Каин, Майкл Л .; Джексон, Роберт Б .; Вассерман, Стивен А .; Минорский, Петр V .; Рис, Джейн Б. (2014). «Биология Кэмпбелла в фокусе. Лиза А. Урри, Майкл Л. Кейн, Стивен А. Вассерман, Питер В. Минорски, Роберт Б. Джексон и Джейн Б. Рис. Бостон (Массачусетс): Пирсон. $ 146,67. Xxxix + 905 п .; ил. + A-1 - A-34; B-1; C-1; D-1; E-1 - E-2; F-1 - F-3; CR-1 - CR-6; Г-1 - Г-34; И-1 - И-48 (указатель) ». Ежеквартальный обзор биологии. 88 (3): 242. Дои:10.1086/671541. ISBN 978-0-321-81380-0.
- ^ Деккер Дж. (25 ноября 2014 г.). «Два способа свернуть геном во время клеточного цикла: идеи, полученные с помощью захвата конформации хромосомы». Эпигенетика и хроматин. 7 (1): 25. Дои:10.1186/1756-8935-7-25. ЧВК 4247682. PMID 25435919.
- ^ Hetzer MW (март 2010 г.). "Ядерная оболочка". Перспективы Колд-Спринг-Харбор в биологии. 2 (3): a000539. Дои:10.1101 / cshperspect.a000539. ЧВК 2829960. PMID 20300205.
- ^ Гилберт С.Ф. (2000). «Оогенез». Биология развития (6-е изд.).
- ^ Guertin, David A .; Траутманн, Сюзанна; Макколлум, Даннел (01.06.2002). «Цитокинез у эукариот». Обзоры микробиологии и молекулярной биологии. 66 (2): 155–178. Дои:10.1128 / MMBR.66.2.155-178.2002. ISSN 1092-2172. ЧВК 120788. PMID 12040122.
- ^ Клетка, Г. Купер; изд 2 Книжная полка NCBI, Эукариотический клеточный цикл, Рисунок 14.7
- ^ Фазовая голографическая визуализация. Отделение клеток
- ^ Jiang H, Schiffer E, Song Z, Wang J, Zürbig P, Thedieck K, Moes S, Bantel H, Saal N, Jantos J, Brecht M, Jenö P, Hall MN, Hager K, Manns MP, Hecker H, Ganser A , Дёнер К., Бартке А., Мейснер К., Мишак Х., Джу З., Рудольф К.Л. (август 2008 г.). «Белки, вызванные дисфункцией теломер и повреждением ДНК, представляют собой биомаркеры старения и болезней человека». Труды Национальной академии наук Соединенных Штатов Америки. 105 (32): 11299–304. Bibcode:2008PNAS..10511299J. Дои:10.1073 / pnas.0801457105. ЧВК 2516278. PMID 18695223.
- ^ Коутон Р.М., Смит К.Р., О'Брайен Э., Сиваченко А., Кербер Р.А. (февраль 2003 г.). «Связь между длиной теломер в крови и смертностью людей в возрасте 60 лет и старше». Ланцет. 361 (9355): 393–5. Дои:10.1016 / S0140-6736 (03) 12384-7. PMID 12573379. S2CID 38437955.
- ^ Джафри М.А., Ансари С.А., Алкахтани М.Х., Шай Дж.В. (июнь 2016 г.). «Роль теломер и теломеразы в развитии рака и достижения в области терапии, направленной на теломеразу». Геномная медицина. 8 (1): 69. Дои:10.1186 / s13073-016-0324-х. ЧВК 4915101. PMID 27323951.
- ^ Биография, Deutsche. "Моль, Хьюго фон - Deutsche Biographie". www.deutsche-biographie.de (на немецком). Получено 2019-04-15.
- ^ Мастерс BR (15.12.2008). «История оптического микроскопа в клеточной биологии и медицине». Энциклопедия наук о жизни. John Wiley & Sons, Ltd. Дои:10.1002 / 9780470015902.a0003082. ISBN 978-0470016176.
- ^ ZEISS Microscopy (01.06.2013), Исторический замедленный фильм доктора Курта Мишеля, Carl Zeiss Jena (около 1943 г.), получено 2019-04-15
дальнейшее чтение
- Морган Привет. (2007). «Клеточный цикл: принципы контроля» Лондон: New Science Press.
- Дж. М. Тернер Плод в мужчину (1978, 1989). Издательство Гарвардского университета. ISBN 0-674-30692-9
- Деление клетки: бинарное деление и митоз
- В Секция митоза и контроля клеточного цикла от Основные статьи по клеточной биологии (Gall JG, McIntosh JR, ред.) Содержит комментарии и ссылки на основополагающие исследовательские работы по митозу и делению клеток. Опубликовано в Интернете в Библиотека изображений и видео из Американское общество клеточной биологии
- В Библиотека изображений и видео из Американское общество клеточной биологии содержит множество видеороликов, показывающих деление клеток.
- В Отделение клеток из Библиотека изображений ячеек
- Изображений : Calanthe обесцвечивание Линдл. - Секретный цветочный сад Флавона
- Модель деления клеток Тайсона и Описание на База данных биомоделей
- WormWeb.org: интерактивная визуализация C. elegans Клеточная линия - Визуализируйте весь набор делений клеток нематоды C. elegans