Эукариот - Eukaryote
| Эукариот | |
|---|---|
 | |
| Эукариоты и некоторые примеры их разнообразия - по часовой стрелке сверху слева: Красная пчела каменщика, Белый гриб, шимпанзе, Изотриха кишечника, Ranunculus asiaticus, и Volvox carteri | |
| Научная классификация | |
| Домен: | Эукариоты (Chatton, 1925) Whittaker & Маргулис, 1978 |
| Супергруппы[2] и королевства | |
Эукариотические организмы, которые не могут быть отнесены к царствам Plantae, Animalia или Fungi, иногда группируются в королевстве. Протиста. | |
Эукариоты (/juˈkærяoʊтs,-əтs/) находятся организмы чей клетки есть ядро заключен в ядерная оболочка.[3][4][5] Эукариоты принадлежат к домен Эукариоты или же Эукария; их имя происходит от Греческий εὖ (Европа, "хорошо" или "хорошо") и κάρυον (карион, «орех» или «ядро»).[6] Домен Eukaryota составляет одну из сфер жизни в ныне устаревшем трехдоменная система: Два других домена Бактерии и Археи (вместе известные как прокариоты ), а эукариоты обычно рассматриваются как возникшие в архее или как сестра ныне культивируемых Асгард Археи.[7][8][9][10][11] Эукариоты представляют крошечное меньшинство среди живых организмов;[12] однако из-за их, как правило, гораздо большего размера, их коллектив во всем мире биомасса оценивается примерно так же, как у прокариот.[12] Эукариоты возникли примерно 2,1-1,6 миллиарда лет назад, во время Протерозойский эон, вероятно, как бичеванный фаготрофы.[13]
Эукариотические клетки обычно содержат мембраносвязанные органеллы Такие как митохондрии и аппарат Гольджи, и хлоропласты можно найти в растения и водоросли; эти органеллы уникальны для эукариот, хотя примитивные органеллы можно найти у прокариот.[14] А также быть одноклеточный, эукариоты также могут быть многоклеточный и включать много типы клеток формирование различных видов ткань; для сравнения, прокариоты обычно одноклеточные. Животные, растения, и грибы самые знакомые эукариоты; другие эукариоты иногда называют протисты.[15]
Эукариоты могут воспроизводить оба бесполым путем через митоз и сексуально через мейоз и гамета слияние. В митозе одна клетка делится, чтобы произвести две генетически идентичные клетки. В мейозе Репликация ДНК за ним следуют два раунда деление клеток произвести четыре гаплоидный дочерние клетки. Они действуют как половые клетки (гаметы). Каждая гамета имеет только один набор хромосом, каждая из которых представляет собой уникальное сочетание соответствующей пары родительских хромосомы в результате генетическая рекомбинация во время мейоза.[16]
История концепции

Концепция эукариота была приписана французскому биологу. Эдуард Чаттон (1883–1947). Термины прокариот и эукариот были более окончательно введены канадским микробиологом. Роджер Станье и голландско-американский микробиолог К. Б. ван Ниль в 1962 г. В его работе 1937 г. Titres et Travaux Scientifiques,[17] Чаттон предложил два термина, назвав бактерии прокариотами и организмы с ядрами в своих клетках эукариотами. Однако он упомянул об этом только в одном абзаце, и идея фактически игнорировалась до тех пор, пока заявление Чаттона не было переоткрыто Станье и ван Нилом.[18]
В 1905 и 1910 годах русский биолог Константин Мерещковский (1855–1921) утверждал, что пластиды были сокращены цианобактерии в симбиоз с не-фотосинтетический (гетеротрофный ) хозяин, который сам был сформирован симбиозом между амебоподобным хозяином и бактериоподобной клеткой, которая сформировала ядро. Таким образом, растения унаследовали фотосинтез от цианобактерий.[19]
В 1967 г. Линн Маргулис предоставил микробиологические доказательства для эндосимбиоз как происхождение хлоропластов и митохондрий в эукариотических клетках в ее статье, О происхождении митозирующих клеток.[20] В 1970-е годы Карл Вёзе исследованный микробный филогенетика, изучая вариации в 16S рибосомная РНК. Это помогло раскрыть происхождение эукариот и симбиогенез двух важных эукариот органеллы, митохондрии и хлоропласты. В 1977 году Вёзе и Джордж Фокс представили «третью форму жизни», которую они назвали архебактериями; в 1990 году Вёзе, Отто Кандлер и Марк Л. Уилис переименовал это в Архею.[21][18]
В 1979 г. Дж. У. Гулд и Дж. Дж. Дринг предположили, что эукариотическая клетка ядро пришло из способности Грамположительные бактерии формировать эндоспоры. В статьях 1987 г. и позже, Томас Кавалье-Смит предложил вместо этого, чтобы мембраны ядра и эндоплазматический ретикулум сначала образуется в результате складывания плазматической мембраны прокариот. В 1990-х годах несколько других биологов предложили эндосимбиотическое происхождение ядра, эффективно возродив теорию Мерешковского.[19]
Особенности ячейки

Эукариотические клетки обычно намного больше, чем у прокариоты, имеющий объем примерно в 10 000 раз больше, чем прокариотическая клетка.[22] Они имеют множество внутренних мембраносвязанных структур, называемых органеллы, а цитоскелет состоит из микротрубочки, микрофиламенты, и промежуточные нити, которые играют важную роль в определении организации и формы клетки. Эукариотический ДНК делится на несколько линейных пучков, называемых хромосомы, которые разделены микротрубчатый шпиндель во время ядерного деления.
Внутренняя мембрана

Клетки эукариотов включают множество мембраносвязанных структур, вместе называемых эндомембранная система.[23] Простые отсеки, называемые пузырьки и вакуоли, может образоваться за счет отпочкования других мембран. Многие клетки поглощают пищу и другие материалы в процессе эндоцитоз, где внешняя мембрана инвагинирует а затем отщипывает, образуя пузырек.[24] Вероятно[нужна цитата ] что большинство других мембраносвязанных органелл в конечном итоге происходит из таких везикул. Альтернативно, некоторые продукты, продуцируемые клеткой, могут покидать пузырьки через экзоцитоз.
Ядро окружено двойной мембраной (обычно называемой ядерная мембрана или ядерная оболочка) с порами, которые позволяют материалу входить и выходить.[25] Различные трубчатые и пластинчатые продолжения ядерной мембраны образуют эндоплазматический ретикулум, который участвует в транспорте и созревании белка. Он включает грубую эндоплазматическую сеть, где рибосомы прикрепляются для синтеза белков, которые попадают во внутреннее пространство или просвет. Впоследствии они обычно попадают в пузырьки, которые отпочковываются от гладкой эндоплазматической сети.[26] У большинства эукариот эти несущие белок везикулы высвобождаются и далее модифицируются в стопки уплощенных везикул (цистерны ), аппарат Гольджи.[27]
Везикулы могут быть специализированы для различных целей. Например, лизосомы содержат пищеварительный ферменты что ломается больше всего биомолекулы в цитоплазме.[28] Пероксисомы используются для разрушения перекись, который в остальном токсичен. Много простейшие имеют сократительные вакуоли, которые собирают и выводят лишнюю воду, и экструзомы, которые изгоняют материал, используемый для отражения хищников или поимки добычи. У высших растений большую часть объема клетки занимает центральная вакуоль, которая в основном содержит воду и в первую очередь поддерживает ее осмотическое давление.
Митохондрии и пластиды

Митохондрии органеллы найдены во всех, кроме одного[примечание 1] эукариот. Митохондрии обеспечивают энергией клетки эукариотов, превращая сахара в АТФ.[30] У них есть два окружающих мембраны, каждый фосфолипидный бислой; то внутренний из которых складывается в инвагинации, называемые кристы куда аэробного дыхания происходит.
Наружная митохондриальная мембрана свободно проницаема и позволяет практически всему проникать в межмембранное пространство в то время как внутренняя митохондриальная мембрана полупроницаема, поэтому пропускает только некоторые необходимые вещи в матрицу митохондрий.
Митохондрии содержат их собственная ДНК, который имеет близкое структурное сходство с бактериальной ДНК и кодирует рРНК и тРНК гены, производящие РНК, которая по структуре ближе к бактериальной РНК, чем к РНК эукариот.[31] В настоящее время считается, что они произошли от эндосимбиотический прокариоты, вероятно протеобактерии.
Некоторые эукариоты, такие как метамонады Такие как Лямблии и Трихомонада, а амебозой Пеломикс, по-видимому, лишены митохондрий, но все они содержат органеллы, происходящие из митохондрий, такие как гидрогеносомы и митосомы, и, таким образом, вторично потеряли свои митохондрии.[29] Они получают энергию за счет ферментативного воздействия на питательные вещества, поступающие из окружающей среды. Метамонада Monocercomonoides также приобрел боковой перенос гена, цитозольный сера система мобилизации, которая обеспечивает кластеры железа и серы, необходимые для синтеза белка. Нормальный путь митохондриального кластера железа и серы был вторично утрачен.[29][32]
Растения и различные группы водоросли Также есть пластиды. Пластиды также имеют их собственная ДНК и разработаны из эндосимбионты, в этом случае цианобактерии. Обычно они имеют форму хлоропласты которые, как и цианобактерии, содержат хлорофилл и производят органические соединения (такие как глюкоза ) через фотосинтез. Остальные занимаются хранением еды. Хотя пластиды, вероятно, имели одно происхождение, не все пластидсодержащие группы тесно связаны. Вместо этого некоторые эукариоты получили их от других через вторичный эндосимбиоз или проглатывание.[33] Захват и секвестрация фотосинтетических клеток и хлоропластов происходит во многих типах современных эукариотических организмов и известна как клептопластика.
Эндосимбиотическое происхождение также было предложено для ядра и эукариотического происхождения. жгутики.[34]
Цитоскелетные структуры

Многие эукариоты имеют длинные тонкие подвижные цитоплазматические выступы, называемые жгутики, или аналогичные структуры, называемые реснички. Жгутики и реснички иногда называют ундулиподия,[35] и по-разному вовлечены в движение, питание и ощущения. Они состоят в основном из тубулин. Они полностью отличаются от жгутиков прокариот. Они поддерживаются связкой микротрубочек, возникающих из центриоль, характерно расположенные в виде девяти дублетов, окружающих два синглета. Жгутики также могут иметь волоски или мастигонемы, и чешуйки, соединяющие мембраны и внутренние стержни. Их интерьер является продолжением камеры цитоплазма.
Микрофиламентные структуры, состоящие из актин и актин-связывающие белки, например, α-актинин, фимбрин, филамин также присутствуют в субмембранозных корковых слоях и пучках. Моторные белки микротрубочек, например, динеин или же кинезин и актин, например, миозины обеспечить динамичный характер сети.
Центриоли часто присутствуют даже в клетках и группах, не имеющих жгутиков, но хвойные породы и цветущие растения нет ни того, ни другого. Обычно они встречаются группами, которые дают начало различным корням микротрубочек. Они образуют первичный компонент структуры цитоскелета и часто собираются в ходе нескольких клеточных делений, при этом один жгутик сохраняется от родителя, а другой происходит от него. Центриоли образуют веретено во время деления ядра.[36]
Важность цитоскелетных структур подчеркивается в определении формы клеток, а также в том, что они являются важными компонентами миграционных реакций, таких как хемотаксис и хемокинез. Немного протисты имеют различные другие органеллы, поддерживаемые микротрубочками. К ним относятся радиолярии и гелиозоа, которые производят аксоподия используется для плавания или для ловли добычи, а гаптофиты, которые имеют своеобразную органеллу, похожую на жгутик, называемую гаптонема.
Клеточная стенка
Клетки растений и водорослей, грибов и большинства хромальвеолаты имеют клеточную стенку, слой за пределами клеточная мембрана, обеспечивая ячейку структурной опорой, защитой и фильтрующим механизмом. Стенка клетки также предотвращает чрезмерное расширение при попадании воды в клетку.[37]
Главная полисахариды составляя первичную клеточную стенку наземные растения находятся целлюлоза, гемицеллюлоза, и пектин. Целлюлоза микрофибриллы связаны гемицеллюлозными связями с образованием целлюлозно-гемицеллюлозной сети, которая встроена в пектиновую матрицу. Наиболее распространенная гемицеллюлоза в первичной клеточной стенке - это ксилоглюкан.[38]
Различия между эукариотическими клетками
Существует множество различных типов эукариотических клеток, хотя животные и растения являются наиболее знакомыми эукариотами и, таким образом, являются отличной отправной точкой для понимания структуры эукариот. Однако у грибов и многих протистов есть существенные различия.
Животная клетка
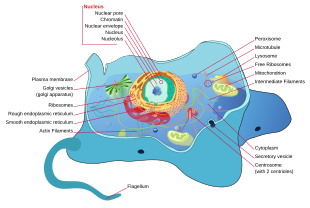
Все животные эукариоты. Клетки животных отличаются от клеток других эукариот, в первую очередь растения, поскольку им не хватает клеточные стенки и хлоропласты и иметь меньше вакуоли. Из-за отсутствия клеточная стенка клетки животных могут принимать самые разные формы. А фагоцитарный клетка может даже поглотить другие структуры.
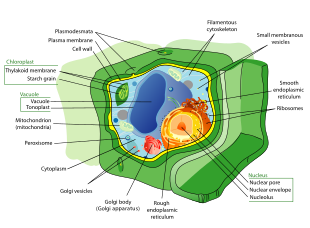
Растительная клетка
Клетки растений сильно отличаются от клеток других эукариотических организмов. Их отличительными особенностями являются:
- Большой центральный вакуоль (заключены в мембрану, тонопласт ), который поддерживает клеточную тургор и контролирует движение молекулы между цитозоль и сок[39]
- Первичный клеточная стенка содержащий целлюлоза, гемицеллюлоза и пектин, сданные на хранение протопласт на внешней стороне клеточная мембрана; это контрастирует с клеточными стенками грибы, которые содержат хитин, а конверты ячеек прокариот, у которых пептидогликаны основные структурные молекулы
- В плазмодесматы, поры в клеточной стенке, которые связывают соседние клетки и позволяют клеткам растений связываться с соседними клетками.[40] У животных другая, но функционально аналогичная система щелевые соединения между соседними ячейками.
- Пластиды, особенно хлоропласты, органеллы которые содержат хлорофилл, пигмент, дающий растения их зеленый цвет и позволяет им выполнять фотосинтез
- Мохообразные и бессемянные сосудистые растения в сперматозоидах есть только жгутики и центриоли.[41] Сперма саговники и Гинкго представляют собой большие сложные клетки, в которых плавают от сотен до тысяч жгутиков.[42]
- Хвойные (Pinophyta) и цветущие растения (Покрытосеменных) не хватает жгутики и центриоли которые присутствуют в клетках животных.
Грибковая клетка

Ячейки грибы наиболее похожи на клетки животных, за исключением следующих случаев:[43]
- Стенка клетки, содержащая хитин
- Меньшее разделение между ячейками; то гифы высших грибов имеют пористые перегородки, называемые септа, которые позволяют прохождение цитоплазмы, органелл и, иногда, ядер; так что каждый организм по сути гигантский многоядерный supercell - эти грибы описываются как ценоцитарный. У примитивных грибов мало или нет перегородок.
- Только самые примитивные грибы, хитриды, есть жгутики.
Другие эукариотические клетки
У некоторых групп эукариот есть уникальные органеллы, такие как цианеллы (необычные хлоропласты) глаукофиты,[44] гаптонема гаптофиты, или эжектосомы из криптомонады. Другие конструкции, такие как псевдоподия, встречаются в различных группах эукариот в разных формах, например, лобозной амебозойные или сеточка фораминиферы.[45]
Размножение

Деление клеток обычно имеет место бесполым путем к митоз, процесс, который позволяет каждому дочернему ядру получать по одной копии каждого хромосома. У большинства эукариот также есть жизненный цикл, который включает: половое размножение, чередование между гаплоидный фаза, когда только одна копия каждой хромосомы присутствует в каждой клетке и диплоид фаза, при которой две копии каждой хромосомы присутствуют в каждой клетке. Диплоидная фаза образуется путем слияния двух гаплоидных гамет с образованием зиготы, которая может делиться митозом или подвергаться редукции хромосомы за счет мейоз. Этот паттерн сильно различается. У животных нет многоклеточной гаплоидной фазы, но каждое поколение растений может состоять из гаплоидной и диплоидной многоклеточных фаз.
Эукариоты имеют меньшее отношение площади поверхности к объему, чем прокариоты, и, следовательно, имеют более низкие скорости метаболизма и более длительное время генерации.[46]
В эволюция полового размножения может быть изначальной и фундаментальной характеристикой эукариот. На основании филогенетического анализа, Dacks and Роджер предположили, что факультативный пол присутствовал у общего предка всех эукариот.[47] Основной набор генов, которые функционируют в мейозе, присутствует в обоих Влагалищная трихомонада и Лямблии кишечные, два организма, которые ранее считались бесполыми.[48][49] Поскольку эти два вида являются потомками линий, которые рано разошлись с эволюционным деревом эукариот, был сделан вывод, что основные мейотические гены и, следовательно, пол, вероятно, присутствовали у общего предка всех эукариот.[48][49] Эукариотические виды, которые когда-то считались бесполыми, например, паразитические простейшие из рода Лейшмания, было показано, что у них есть половой цикл.[50] Кроме того, теперь данные показывают, что амебы, ранее считавшиеся бесполыми, в древности являются половыми и что большинство современных бесполых групп, вероятно, возникли недавно и независимо.[51]
Классификация



В древность, две линии животные и растения были признаны. Им дали таксономический ранг из Королевство к Линней. Хотя он включил грибы Что касается растений с некоторыми оговорками, то позже выяснилось, что они совершенно разные и требуют отдельного царства, состав которого не был полностью ясен до 1980-х годов.[52] Когда стало известно, различные одноклеточные эукариоты изначально были помещены вместе с растениями или животными. В 1818 году немецкий биолог Георг А. Гольдфусс придумал слово простейшие для обозначения организмов, таких как инфузории,[53] и эта группа была расширена до тех пор, пока она не охватила всех одноклеточных эукариот, и получила свое собственное царство, Протиста, к Эрнст Геккель в 1866 г.[54][55] Таким образом, эукариоты состояли из четырех царств:
Протистов считали «примитивными формами», и, следовательно, эволюционная ступень, объединенные своей примитивной одноклеточной природой.[55] Распутывание глубоких трещин в Дерево жизни только действительно началось с Секвенирование ДНК, что приводит к системе домены а не королевства в качестве ранга высшего уровня, выдвигаемого Карл Вёзе, объединяющий все царства эукариотов под доменом эукариотов.[21] В то же время активизировались работы над деревом протистов, которые активно продолжаются и сегодня. Было предложено несколько альтернативных классификаций, хотя единого мнения в этой области нет.
Эукариоты клада обычно считается сестрой Heimdallarchaeota в Асгард группировка в Археи.[56][57][58] В одной предложенной системе базовые группы представляют собой Opimoda, Дифода, то Дискоба, а Лукозоа. Корень эукариот обычно считается находящимся рядом или даже в Дискобе.
А классификация, разработанная в 2005 году для Международного общества протистологов,[59] который отражал консенсус того времени, разделил эукариот на шесть якобы монофилетических «супергрупп». Однако в том же году (2005 г.) были высказаны сомнения относительно того, являются ли некоторые из этих супергрупп монофилетический, особенно Хромальвеолаты,[60] а обзор, проведенный в 2006 году, отметил отсутствие доказательств для некоторых из предполагаемых шести супергрупп.[61] Пересмотренная классификация 2012 г.[2] распознает пять супергрупп.
| Archaeplastida (или Primoplantae) | Наземные растения, зеленые водоросли, красные водоросли, и глаукофиты |
| SAR супергруппа | Страменопилы (бурые водоросли, диатомеи, так далее.), Альвеолаты, и Ризария (Фораминиферы, Радиолярии, и различные другие амебовидный простейшие) |
| Экскавата | Разные жгутик простейшие |
| Амебозоа | Самый лобозный амебоиды и слизевые формы |
| Опистоконта | Животные, грибы, хоанофлагелляты, так далее. |
Существуют также более мелкие группы эукариот, положение которых неясно или, кажется, выходит за рамки основных групп.[62] - особенно, Гаптофита, Криптофита, Centrohelida, Телонемия, Пикозоа,[63] Апусомонадида, Анциромонадиды, Breviatea, а род Коллодиктион.[64] В целом кажется, что, несмотря на достигнутый прогресс, все еще существуют очень значительные неопределенности в истории эволюции и классификации эукариот. В качестве Роджер & Симпсон сказал в 2009 году, что «с нынешними темпами изменений в нашем понимании дерева жизни эукариот, мы должны действовать осторожно».[65]
В статье, опубликованной в Природная микробиология в апреле 2016 года авторы «еще раз подтвердили, что жизнь, которую мы видим вокруг себя - растения, животные, люди и другие так называемые эукариоты, - представляют крошечный процент мирового биоразнообразия».[66] Они классифицировали эукариот «на основе наследования их информационных систем в отличие от липидов или других клеточных структур». Джиллиан Ф. Бэнфилд из Калифорнийский университет в Беркли и коллеги-ученые использовали суперкомпьютер для создания схемы нового древа жизни на основе ДНК 3000 видов, включая 2072 известных вида и 1011 недавно зарегистрированных микробных организмов, ДНК которых они собрали из различных сред.[8][67] Поскольку способность секвенировать ДНК стала проще, Бэнфилд и его команда смогли выполнить метагеномное секвенирование - «секвенировать целые сообщества организмов одновременно и выбирать отдельные группы только на основе их генов».[66]
Филогения
В рРНК деревья, построенные в 1980-х и 1990-х годах, оставили большинство эукариот в нерешенной группе "кроны" (технически не Корона ), который обычно делили по форме митохондриальных крист; видеть коронные эукариоты. Несколько групп, которым не хватает митохондрии разветвлялись отдельно, поэтому отсутствие считалось примитивным; но теперь это считается артефактом притяжение длинных ветвей, и они, как известно, потеряли их вторично.[68][69]
По состоянию на 2011 г.[Обновить], широко распространено мнение, что ризарии принадлежат к Stramenopiles и Alveolata в клады назвал SAR супергруппа, так что Rhizaria не является одной из основных групп эукариот; также что Amoebozoa и Opisthokonta являются монофилетическими и образуют кладу, часто называемую unikonts.[70][71][72][73][74] Помимо этого, похоже, нет консенсуса.
Было подсчитано, что может существовать 75 различных линий эукариот.[75] Большинство этих линий - протисты.
Известные размеры генома эукариот варьируются от 8,2 мегабайт (МБ) в Babesia bovis до 112 000–220 050 МБ в динофлагелляте Prorocentrum micans, показывая, что геном предкового эукариота претерпел значительные изменения в ходе своей эволюции.[75] Считается, что последним общим предком всех эукариот был фаготрофный протист с ядром, по крайней мере, одним центриоль и ресничка, факультативно аэробные митохондрии, пол (мейоз и сингамия ), спящий киста с клеточной стенкой из хитин и / или целлюлоза и пероксисомы.[75] Позднее эндосимбиоз привел к распространению пластид в некоторых линиях.
Пять супергрупп
Глобальное дерево эукариот на основе консенсуса филогенетических данных (в частности, филогеномики), редких геномных сигнатур и морфологических характеристик представлено в Adl. и другие. 2012[2] и Бурки 2014/2016, когда пикозоа появились в Archaeplastida, и Криптиста как его сестра.[62][76][77][78][79][80][29][81] Возможно, ЦАР - сестра Гаптисты.[82][83]
| Эукариоты |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




В некоторых исследованиях Хакробия группа (Гаптофита + Криптофита ) находится рядом с Archaeplastida,[70] но в других он гнездится внутри Archaeplastida.[84] Однако несколько недавних исследований пришли к выводу, что Haptophyta и Cryptophyta не образуют монофилетическую группу.[85] Первая могла быть сестринской группой Группа SAR, последний кластер с Archaeplastida (растения в широком смысле).[86]
Разделение эукариот на две основные клады, биконты (Archaeplastida + SAR + Экскавата ) и unikonts (Амебозоа + Опистоконта ), происходящие от предкового двухжеллярного организма и предкового однофлагеллярного организма, соответственно, были предложены ранее.[84][87][88] В исследовании 2012 года было получено несколько аналогичное разделение, хотя и отмечалось, что термины «униконты» и «биконты» не использовались в первоначальном смысле.[63]
Набор сильно сходящихся и совпадающих деревьев представлен у Derelle et al. (2015), Ren et al. (2016), Ян и др. (2017) и Cavalier-Smith (2015), включая дополнительную информацию, что привело к более консервативному и консолидированному дереву. Он сочетается с некоторыми результатами Кавальер-Смита для базальной Opimoda.[89][90][91][92][93][79][94] Основными остающимися противоречиями являются корень и точное расположение Rhodophyta и биконты Rhizaria, Haptista, Cryptista, Picozoa и Telonemia, многие из которых могут быть эндосимбиотическими гибридами эукариот-эукариот.[95] Приобретена археопластида хлоропласты вероятно, эндосимбиозом прокариотического предка, связанного с ныне существующими цианобактерии, Gloeomargarita lithophora.[96][97][95]
| Эукариоты |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Дерево кавалера-смита
Томас Кавалье-Смит 2010,[98] 2013,[99] 2014,[100] 2017[90] и 2018[101] помещает корень эукариотического дерева между Экскавата (с брюшной канавкой для кормления, поддерживаемой корнем микротрубочки) и без бороздки Эвгленозоа, и монофилетический Chromista, коррелированный с одним эндосимбиотическим событием поимки красных водорослей. He et al.[102] специально поддерживает укоренение эукариотического дерева между монофилетическими Дискоба (Discicristata + Якобида ) и Аморфея -Потогонные клады.
| Эукариоты |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Происхождение эукариот



Происхождение эукариотической клетки является важной вехой в эволюции жизни, поскольку эукариоты включают все сложные клетки и почти все многоклеточные организмы. Для поиска первых эукариотов и их ближайших родственников был использован ряд подходов. В последний общий предок эукариот (LECA) - гипотетический последний общий предок всех эукариот, которые когда-либо жили, и, скорее всего, были биологическим численность населения.[105]
Эукариоты обладают набором характерных черт, которые отличают их от других сфер жизни, включая эндомембранная система и уникальные биохимические пути, такие как стеран синтез.[106] Набор белков, называемых эукариотическими сигнатурными белками (ESP), был предложен для идентификации эукариотических родственников в 2002 году: к тому времени у них не было гомологии с белками, известными в других сферах жизни, но они оказались универсальными среди эукариот. Они включают белки, которые составляют цитоскелет, сложный механизм транскрипции, системы мембранной сортировки, ядерную пору, а также некоторые ферменты биохимических путей.[107]
Окаменелости
Трудно определить время этой серии событий; Knoll (2006) предполагает, что они возникли примерно 1,6–2,1 миллиарда лет назад. Немного акритархи известны не менее 1,65 миллиарда лет назад, а возможные водоросли Грипания был обнаружен еще 2,1 миллиарда лет назад.[108] В Геосифон -подобное ископаемое грибок Дискагма был обнаружен в палеопочвах возрастом 2,2 миллиарда лет.[109]
Организованные жилые конструкции обнаружены в черные сланцы палеопротерозойской франсвильской формации B в Габоне, возраст которой составляет 2,1 миллиарда лет. В то время могла возникнуть эукариотическая жизнь.[110] Окаменелости, которые явно связаны с современными группами, начали появляться примерно 1,2 миллиарда лет назад в виде красные водоросли, хотя недавняя работа предполагает существование окаменелых нитчатые водоросли в Виндхья бассейн, датируемый примерно 1,6–1,7 миллиарда лет назад.[111]
Биомаркеры предположить, что по крайней мере корень эукариоты возникли еще раньше. Наличие стераны в Австралийский сланцы указывает на то, что в этих породах присутствовали эукариоты возрастом 2,7 миллиарда лет,[106][112] хотя предполагалось, что они могли возникнуть из-за загрязнения образцов.[113]
Когда бы они ни возникли, эукариоты могли стать экологически доминирующими гораздо позже; массивный рост цинкового состава морских отложений 800 миллион лет назад было приписано росту значительных популяций эукариот, которые преимущественно потребляют и включают цинк по сравнению с прокариотами.[114]
В апреле 2019 года биологи сообщили, что очень большой медузавирус, или родственник, возможно, несут ответственность, по крайней мере частично, за эволюционное возникновение сложных эукариотических клеток из более простых прокариотические клетки.[115]
Отношение к архее
Ядерная ДНК и генетический аппарат эукариот больше похож на Археи чем Бактерии, что привело к противоречивому предположению, что эукариоты должны быть сгруппированы с археями в кладу Неомура. В остальном, например, по составу мембран, эукариоты похожи на Бактерии. Было предложено три основных объяснения этого:
- Эукариоты возникли в результате полного слияния двух или более клеток, при этом цитоплазма образовалась из эубактерии, и ядро из архея,[116] от вируса,[117][118] или из предварительная ячейка.[119][120]
- Эукариоты произошли от архей и приобрели свои эубактериальные характеристики благодаря эндосимбиоз прото-митохондрии эубактериального происхождения.[121]
- Эукариоты и археи развивались отдельно от модифицированных эубактерий.

Альтернативные предложения включают:
- Гипотеза хроноцитов постулирует, что примитивная эукариотическая клетка была сформирована в результате эндосимбиоза как архей, так и бактерий третьим типом клеток, называемых клетками. хроноцит. В основном это связано с тем, что к 2002 году сигнатурные белки эукариот не были обнаружены где-либо еще.[107]
- Универсальный общий предок (UCA) нынешнего древа жизни был сложным организмом, пережившим событие массового вымирания, а не раннюю стадию эволюции жизни. Эукариоты и, в частности, акариоты (бактерии и археи) эволюционировали за счет редукционной потери, так что сходство является результатом дифференциального сохранения исходных характеристик.[123]
Если предположить, что никакая другая группа не участвует, существует три возможных филогении для бактерий, архей и эукариот, каждая из которых является монофилетической. В таблице ниже они обозначены от 1 до 3. Гипотеза эоцитов является модификацией гипотезы 2, согласно которой археи являются парафилетическими. (Таблица и названия гипотез основаны на Harish and Kurland, 2017.[124])
| 1 - Две империи | 2 - Три домена | 3 - Гупта | 4 - Эоцит | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
В последние годы большинство исследователей отдали предпочтение либо трех доменам (3D), либо гипотезе эоцитов. An рРНК анализы подтверждают сценарий эоцитов, очевидно, с корнем эукариот в Экскавата.[67][98][99][100][90] Кладограмма, подтверждающая гипотезу эоцитов, позиционирующая эукариот внутри архей, основана на филогеномном анализе Асгард архей, это:[56][57][58][10]
| Протеоархеи |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
В этом сценарии группа Асгарда рассматривается как родственный таксон TACK группа, в которую входят Crenarchaeota (ранее назывался эоциты ), Таумархеота, и другие. Сообщается, что эта группа содержит много сигнатурных белков эукариот и продуцирует пузырьки.[125]
В 2017 году этот сценарий получил серьезный отпор, утверждая, что эукариоты не появились в архее. Cunha и другие. провели анализы, подтверждающие три области (3D) или гипотезу Вёза (2 в таблице выше) и отвергающие гипотезу эоцитов (4 выше).[126] Хариш и Курланд нашли сильную поддержку двух предыдущих империй (2D) или гипотезы Майра (1 в таблице выше), основываясь на анализе кодирующих последовательностей белковых доменов. Они отвергли гипотезу эоцитов как наименее вероятную.[127][124] Возможная интерпретация их анализа состоит в том, что универсальный общий предок (UCA) нынешнего древа жизни был сложным организмом, который пережил эволюционное узкое место, а не более простой организм, возникший в начале истории жизни.[123] С другой стороны, исследователи, придумавшие Асгард, подтвердили свою гипотезу дополнительными образцами Асгарда.[128]
Детали взаимоотношений представителей архей Асгарда и эукариот все еще рассматриваются.[129] хотя в январе 2020 года ученые сообщили, что Candidatus Prometheoarchaeum syntrophicum, тип культурных архей Асгарда, может быть возможным связующим звеном между простыми прокариотический и сложный эукариотический микроорганизмов около двух миллиардов лет назад.[130][125]
Эндомембранная система и митохондрии
Происхождение эндомембранной системы и митохондрий также неясно.[131] В фаготрофная гипотеза предполагает, что мембраны эукариотического типа, лишенные клеточной стенки, возникли первыми, с развитием эндоцитоза, тогда как митохондрии были приобретены при приеме внутрь в виде эндосимбионтов.[132] В синтрофическая гипотеза предполагает, что прото-эукариот полагался на прото-митохондрии в качестве пищи, и поэтому в конечном итоге вырос, чтобы окружить его. Здесь мембраны образовались после поглощения митохондрии, отчасти благодаря митохондриальным генам ( водородная гипотеза это одна конкретная версия).[133]
В исследовании с использованием геномов для построения супердеревья, Пизани и другие. (2007) предполагают, что наряду с доказательствами того, что никогда не существовало эукариот без митохондрий, эукариоты произошли от синтрофия между археями, тесно связанными с Thermoplasmatales и α-протеобактерии, вероятно, симбиоз управляемый серой или водородом. Митохондрия и ее геном - это остатки α-протеобактериального эндосимбионта.[134] Большинство генов симбионта перенесено в ядро. Они составляют большую часть метаболических и энергетических путей эукариотической клетки, в то время как информационная система сохраняется от архей.[135]
Гипотезы
Разные гипотезы были высказаны предположения о том, как возникли эукариотические клетки. Эти гипотезы можно разделить на два разных класса - автогенные модели и химерные модели.
Автогенные модели
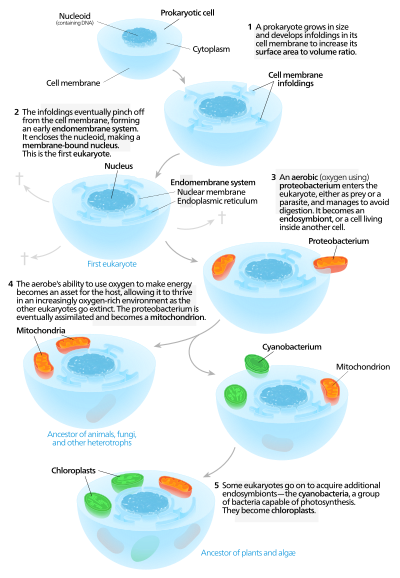
Аутогенные модели предполагают, что протоэукариотическая клетка, содержащая ядро существовал сначала, а потом приобрел митохондрии.[136] Согласно этой модели, большой прокариот развитый инвагинации в его плазматическая мембрана чтобы получить достаточно площадь поверхности для обслуживания своего цитоплазматический объем. По мере того, как инвагинации дифференцировались по функциям, некоторые из них стали отдельными отделениями, что привело к возникновению эндомембранная система, в том числе эндоплазматический ретикулум, аппарат Гольджи, ядерная мембрана, и одинарные мембранные конструкции, такие как лизосомы.[137]
Митохондрии предлагается поступить из эндосимбиоз аэробного протеобактерии, и предполагается, что все эукариотические линии, которые не приобрели митохондрии, вымерли,[138] заявление, подвергшееся критике за отсутствие фальсифицируемость. Хлоропласты возникли в результате другого эндосимбиотического события, связанного с цианобактерии. Поскольку все известные эукариоты имеют митохондрии, но не все имеют хлоропласты, серийный эндосимбиоз теория предполагает, что митохондрии были первыми.
Химерные модели
Химерные модели утверждают, что изначально существовали две прокариотические клетки - одна Археон и бактерия. Ближайшие из ныне живущих родственников этих Асгардархеота и (отдаленно связанные) альфапротеобактерии.[139][140] Эти клетки подверглись процессу слияния, либо путем физического слияния, либо путем эндосимбиоз, тем самым приводя к образованию эукариотической клетки. В рамках этих химерных моделей некоторые исследования также утверждают, что митохондрии произошли от бактериального предка, в то время как другие подчеркивают роль эндосимбиотических процессов в происхождении митохондрий.
Гипотеза наизнанку
Гипотеза наизнанку, разработанная двоюродными братьями Дэвидом и Баззом Баумом, предполагает, что слияние свободноживущих митохондриоподобных бактерий и архея в эукариотическую клетку происходило постепенно, в течение длительного периода времени, а не фагоцитоз залпом. В этом сценарии археон ловит аэробные бактерии с помощью выступы клеток, а затем поддерживайте их в живых, чтобы черпать из них энергию, а не переваривать. На ранних стадиях бактерии все еще частично находились бы в прямом контакте с окружающей средой, и археону не нужно было бы обеспечивать их всеми необходимыми питательными веществами. Но в конечном итоге археон полностью поглотит бактерии, создав внутренние мембранные структуры и мембрану ядра.[141]
Предполагается, что архейская группа под названием галофилы прошли через аналогичную процедуру, в которой они приобрели целую тысячу генов от бактерии, намного больше, чем с помощью обычных горизонтальный перенос генов that often occurs in the microbial world, but that the two microbes separated again before they had fused into a single eukaryote-like cell.[142]
Based on the process of mutualistic symbiosis, the hypotheses can be categorized as – сериал эндосимбиотический гипотеза or theory (SET),[143][144][145] то hydrogen hypothesis (mostly a process of symbiosis where hydrogen transfer takes place among different species),[133] и syntrophy hypothesis.[146][147] These hypotheses are discussed separately in the following sections.
An expanded version of the inside-out hypothesis proposes that the eukaryotic cell was created by physical interactions between two prokaryotic organisms and that the last common ancestor of eukaryotes got its genome from a whole population or community of microbes participating in cooperative relationships to thrive and survive in their environment. The genome from the various types of microbes would complement each other, and occasional horizontal gene transfer between them would be largely to their own benefit. This accumulation of beneficial genes gave rise to the genome of the eukaryotic cell, which contained all the genes required for independence.[148]
The serial endosymbiotic hypothesis
According to serial endosymbiotic theory (championed by Линн Маргулис ), a union between a motile анаэробный bacterium (like Спирохета) and a thermoacidophilic crenarchaeon (подобно Термоплазма which is sulfidogenic in nature) gave rise to the present day eukaryotes. This union established a motile organism capable of living in the already existing acidic and sulfurous waters. Oxygen is known to cause toxicity to organisms that lack the required метаболический машины. Thus, the archaeon provided the bacterium with a highly beneficial reduced environment (sulfur and sulfate were reduced to sulfide). In microaerophilic conditions, oxygen was reduced to water thereby creating a mutual benefit platform. The bacterium on the other hand, contributed the necessary ферментация продукты и электрон acceptors along with its motility feature to the archaeon thereby gaining a swimming подвижность для организма.
From a consortium of bacterial and archaeal DNA originated the nuclear геном эукариотических клеток. Спирохеты gave rise to the motile features of eukaryotic cells. Endosymbiotic unifications of the ancestors of alpha-proteobacteria and cyanobacteria, led to the origin of mitochondria and пластиды соответственно. Например, Thiodendron has been known to have originated via an ectosymbiotic process based on a similar синтрофия of sulfur existing between the two types of bacteria – Desulphobacter и Спирохета.
However, such an association based on motile symbiosis has never been observed practically. Also there is no evidence of archaeans and spirochetes adapting to intense acid-based environments.[136]
The hydrogen hypothesis
In the hydrogen hypothesis, the symbiotic linkage of an anaerobic and автотрофный methanogenic archaeon (host) with an alpha-proteobacterium (the symbiont) gave rise to the eukaryotes. The host utilized водород (ЧАС2) и углекислый газ (CO
2) производить метан while the symbiont, capable of aerobic respiration, expelled H2 и CO
2 as byproducts of anaerobic fermentation process. The host's methanogenic environment worked as a sink for H2, which resulted in heightened bacterial fermentation.
Endosymbiotic gene transfer (EGT) acted as a catalyst for the host to acquire the symbionts' углевод metabolism and turn гетеротрофный in nature. Subsequently, the host's methane forming capability was lost. Thus, the origins of the heterotrophic organelle (symbiont) are identical to the origins of the eukaryotic родословная. In this hypothesis, the presence of H2 represents the selective force that forged eukaryotes out of prokaryotes.[нужна цитата ]
The syntrophy hypothesis
The syntrophy hypothesis was developed in contrast to the hydrogen hypothesis and proposes the existence of two symbiotic events. According to this theory, the origin of eukaryotic cells was based on metabolic symbiosis (syntrophy) between a methanogenic archaeon and a delta-proteobacterium. This syntrophic symbiosis was initially facilitated by H2 transfer between different species under anaerobic environments. In earlier stages, an alpha-proteobacterium became a member of this integration, and later developed into the mitochondrion. Ген transfer from a delta-proteobacterium to an archaeon led to the methanogenic archaeon developing into a nucleus. The archaeon constituted the genetic apparatus, while the delta-proteobacterium contributed towards the цитоплазматический Особенности.
This theory incorporates two selective forces at the time of nucleus эволюция
- presence of metabolic partitioning to avoid the harmful effects of the co-existence of анаболический и катаболический cellular pathways, and
- prevention of abnormal биосинтез белка due to a vast spread of интроны in the archaeal genes after acquiring the mitochondrion and losing метаногенез.[нужна цитата ]
6+ serial endosymbiosis scenario
Pitts and Galbanón propose a complex scenario of 6+ serial endosymbiotic events of Archaea and bacteria in which mitochondria and an asgard related archaeota were acquired at a late stage of eukaryogenesis, possibly in combination, as a secondary endosymbiont.[149][150] The findings have been rebuked as an artefact.[151]
Смотрите также
- Eukaryote hybrid genome
- Эволюция полового размножения
- Список секвенированных эукариотических геномов
- Parakaryon myojinensis
- Прокариот
- Таумархеота
- Хранилище (органелла)
Примечания
- ^ To date, only one eukaryote, Monocercomonoides, is known to have completely lost its mitochondria.[29]
Рекомендации
- ^ Sakaguchi M, Takishita K, Matsumoto T, Hashimoto T, Inagaki Y (July 2009). "Tracing back EFL gene evolution in the cryptomonads-haptophytes assemblage: separate origins of EFL genes in haptophytes, photosynthetic cryptomonads, and goniomonads". Ген. 441 (1–2): 126–31. Дои:10.1016/j.gene.2008.05.010. PMID 18585873.
- ^ а б c Adl SM, Simpson AG, Lane CE, Lukeš J, Bass D, Bowser SS, et al. (Сентябрь 2012 г.). «Пересмотренная классификация эукариот» (PDF). Журнал эукариотической микробиологии. 59 (5): 429–93. Дои:10.1111 / j.1550-7408.2012.00644.x. ЧВК 3483872. PMID 23020233. Архивировано из оригинал (PDF) on 16 June 2016.
- ^ Янгсон Р.М. (2006). Словарь Коллинза по биологии человека. Глазго: HarperCollins. ISBN 978-0-00-722134-9.
- ^ Нельсон Д.Л., Кокс М.М. (2005). Принципы биохимии Ленингера (4-е изд.). Нью-Йорк: W.H. Фримен. ISBN 978-0-7167-4339-2.
- ^ Martin EA, ed. (1983). Macmillan Dictionary of Life Sciences (2-е изд.). Лондон: Macmillan Press. ISBN 978-0-333-34867-3.
- ^ Харпер, Дуглас. "eukaryotic". Интернет-словарь этимологии.
- ^ Woese CR, Kandler O, Wheelis ML (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев». Труды Национальной академии наук Соединенных Штатов Америки. 87 (12): 4576–9. Bibcode:1990PNAS ... 87,4576 Вт. Дои:10.1073/pnas.87.12.4576. ЧВК 54159. PMID 2112744.
- ^ а б Zimmer C (11 April 2016). "Scientists Unveil New 'Tree of Life'". Нью-Йорк Таймс. Получено 11 апреля 2016.
- ^ Gribaldo, Simonetta; Brochier-Armanet, Céline (1 January 2020). "Evolutionary relationships between Archaea and eukaryotes". Природа Экология и эволюция. 4 (1): 20–21. Дои:10.1038/s41559-019-1073-1. ISSN 2397-334X.
- ^ а б Уильямс, Том А .; Кокс, Саймон Дж .; Фостер, Питер Дж .; Szöllsi, Gergely J .; Embley, T. Martin (9 December 2019). «Филогеномика обеспечивает надежную поддержку двухдоменного древа жизни». Природа Экология и эволюция. 4 (1): 138–147. Дои:10.1038 / с41559-019-1040-х. ISSN 2397-334X. ЧВК 6942926. PMID 31819234.
- ^ Doolittle, W. Ford (24 February 2020). "Evolution: Two Domains of Life or Three?". Текущая биология. 30 (4): R177–R179. Дои:10.1016/j.cub.2020.01.010.
- ^ а б Whitman WB, Coleman DC, Wiebe WJ (June 1998). "Prokaryotes: the unseen majority" (PDF). Труды Национальной академии наук Соединенных Штатов Америки. 95 (12): 6578–6583. Bibcode:1998PNAS ... 95,6578 Вт. Дои:10.1073 / пнас.95.12.6578. ЧВК 33863. PMID 9618454.
- ^ Leander, Brian S. (18 May 2020). "Predatory protists". Текущая биология. 30 (10): R510–R516. Дои:10.1016/j.cub.2020.03.052.
- ^ Мурат, Дороти; Бирн, Меган; Komeili, Arash (1 October 2010). «Клеточная биология прокариотических органелл». Перспективы Колд-Спринг-Харбор в биологии. 2 (10): a000422. Дои:10.1101 / cshperspect.a000422. ЧВК 2944366. PMID 20739411.
- ^ Whittaker, R.H. (January 1969). «Новые концепции царств или организмов. Эволюционные отношения лучше представлены новыми классификациями, чем традиционными двумя царствами». Наука. 163 (3863): 150–60. Bibcode:1969Sci ... 163..150W. CiteSeerX 10.1.1.403.5430. Дои:10.1126 / science.163.3863.150. PMID 5762760.
- ^ Campbell NA, Cain ML, Minorsky PV, Reece JB, Urry LA (2018). "Chapter 13: Sexual Life Cycles and Meiosis". Biology: A Global Approach (11-е изд.). Нью-Йорк: образование Пирсона. ISBN 978-1-292-17043-5.
- ^ Chatton, Édouard (1937). Titres Et Travaux Scientifiques (1906-1937) De Edouard Chatton. Sète: Impr. E. Sottano.
- ^ а б Sapp J (June 2005). "The prokaryote-eukaryote dichotomy: meanings and mythology". Обзоры микробиологии и молекулярной биологии. 69 (2): 292–305. Дои:10.1128 / MMBR.69.2.292-305.2005. ЧВК 1197417. PMID 15944457.
- ^ а б Мартин В.Ф., Гарг С., Зиморский В. (сентябрь 2015 г.). "Эндосимбиотические теории происхождения эукариот". Философские труды Лондонского королевского общества. Серия B, Биологические науки. 370 (1678): 20140330. Дои:10.1098 / rstb.2014.0330. ЧВК 4571569. PMID 26323761.
- ^ Sagan L (March 1967). «О происхождении митозирующих клеток». Журнал теоретической биологии. 14 (3): 255–274. Дои:10.1016/0022-5193(67)90079-3. PMID 11541392.
- ^ а б Woese CR, Kandler O, Wheelis ML (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев». Труды Национальной академии наук Соединенных Штатов Америки. 87 (12): 4576–4579. Bibcode:1990PNAS ... 87,4576 Вт. Дои:10.1073/pnas.87.12.4576. ЧВК 54159. PMID 2112744.
- ^ Yamaguchi M, Worman CO (2014). "Deep-sea microorganisms and the origin of the eukaryotic cell" (PDF). Jpn. J. Protozool. 47 (1, 2): 29–48. Архивировано из оригинал (PDF) 9 августа 2017 г.. Получено 24 октября 2017.
- ^ Linka M, Weber AP (2011). "Evolutionary Integration of Chloroplast Metabolism with the Metabolic Networks of the Cells". In Burnap RL, Vermaas WF (eds.). Functional Genomics and Evolution of Photosynthetic Systems. Springer. п. 215. ISBN 978-9400715332.
- ^ Marsh M (2001). Эндоцитоз. Издательство Оксфордского университета. п. vii. ISBN 978-0-19-963851-2.
- ^ Hetzer MW (March 2010). "The nuclear envelope". Перспективы Колд-Спринг-Харбор в биологии. 2 (3): a000539. Дои:10.1101/cshperspect.a000539. ЧВК 2829960. PMID 20300205.
- ^ «Эндоплазматическая сетка (грубая и гладкая)». Британское общество клеточной биологии. Получено 12 ноября 2017.
- ^ "Golgi Apparatus". British Society for Cell Biology. Архивировано из оригинал 13 ноября 2017 г.. Получено 12 ноября 2017.
- ^ "Lysosome". British Society for Cell Biology. Архивировано из оригинал 13 ноября 2017 г.. Получено 12 ноября 2017.
- ^ а б c d Karnkowska A, Vacek V, Zubáčová Z, Treitli SC, Petrželková R, Eme L, Novák L, árský V, Barlow LD, Herman EK, Soukal P, Hroudová M, Doležal P, Stairs CW, Roger AJ, Eliáš M, Dacks , Влчек Ч, Хампл В. (май 2016 г.). «Эукариот без митохондриальной органеллы». Текущая биология. 26 (10): 1274–1284. Дои:10.1016 / j.cub.2016.03.053. PMID 27185558.
- ^ Mack, Steve (1 May 2006). "Re: Are there eukaryotic cells without mitochondria?". madsci.org.
- ^ Watson J, Hopkins N, Roberts J, Steitz JA, Weiner A (1988). "28: The Origins of Life". Молекулярная биология гена (Четвертое изд.). Менло-Парк, Калифорния: Издательство Benjamin / Cummings Publishing Company, Inc. стр.1154. ISBN 978-0-8053-9614-0.
- ^ Davis JL (13 May 2016). "Scientists Shocked To Discover Eukaryote With NO Mitochondria". IFL Science. Архивировано из оригинал 17 февраля 2019 г.. Получено 13 мая 2016.
- ^ Sato N (2006). "Origin and Evolution of Plastids: Genomic View on the Unification and Diversity of Plastids". В Wise RR, Hoober JK (ред.). Строение и функции пластидов. Достижения в фотосинтезе и дыхании. 23. Springer Нидерланды. pp. 75–102. Дои:10.1007/978-1-4020-4061-0_4. ISBN 978-1-4020-4060-3.
- ^ Margulis L (1998). Symbiotic planet: a new look at evolution. Нью-Йорк: Основные книги. ISBN 978-0-465-07271-2. OCLC 39700477.[страница нужна ]
- ^ Линн Маргулис, Heather I. McKhann & Lorraine Olendzenski (ed.), Illustrated Glossary of Protoctista, Jones and Bartlett Publishers, Boston, 1993, p. xviii. ISBN 0-86720-081-2
- ^ Vorobjev IA, Nadezhdina ES (1987). The centrosome and its role in the organization of microtubules. Международный обзор цитологии. 106. С. 227–293. Дои:10.1016 / S0074-7696 (08) 61714-3. ISBN 978-0-12-364506-7. PMID 3294718.
- ^ Хауленд Дж. Л. (2000). Удивительные археи: открытие еще одной области жизни. Оксфорд: Издательство Оксфордского университета. С. 69–71. ISBN 978-0-19-511183-5.
- ^ Fry, Stephen C. (1989). "The Structure and Functions of Xyloglucan". Журнал экспериментальной ботаники. 40 (1): 1–11. Дои:10.1093/jxb/40.1.1.
- ^ Raven J (July 1987). «Роль вакуолей». Новый Фитолог. 106 (3): 357–422. Дои:10.1111 / j.1469-8137.1987.tb00149.x.
- ^ Oparka K (2005). Плазмодесматы. Оксфорд, Великобритания: Blackwell Publishing.
- ^ Raven PH, Evert RF, Eichorm SE (1999). Биология растений. Нью-Йорк: W.H. Фримен.
- ^ Silflow CD, Lefebvre PA (December 2001). «Сборка и подвижность ресничек и жгутиков эукариот. Уроки Chlamydomonas reinhardtii». Физиология растений. 127 (4): 1500–1507. Дои:10.1104 / стр.010807. ЧВК 1540183. PMID 11743094.
- ^ Дьякон Дж. (2005). Грибковая биология. Кембридж, Массачусетс: Blackwell Publishers. pp. 4 and passim. ISBN 978-1-4051-3066-0.
- ^ Килинг П.Дж. (октябрь 2004 г.). «Разнообразие и эволюционная история пластид и их хозяев». Американский журнал ботаники. 91 (10): 1481–1493. Дои:10.3732 / ajb.91.10.1481. PMID 21652304.
- ^ Patterson DJ. "Amoebae: Protists Which Move and Feed Using Pseudopodia". Веб-проект "Древо жизни". Получено 12 ноября 2017.
- ^ Lane N (June 2011). "Energetics and genetics across the prokaryote-eukaryote divide". Биология Директ. 6 (1): 35. Дои:10.1186/1745-6150-6-35. ЧВК 3152533. PMID 21714941.
- ^ Dacks J, Roger AJ (June 1999). "The first sexual lineage and the relevance of facultative sex". Журнал молекулярной эволюции. 48 (6): 779–783. Bibcode:1999JMolE..48..779D. Дои:10.1007/PL00013156. PMID 10229582. S2CID 9441768.
- ^ а б Ramesh MA, Malik SB, Logsdon JM (January 2005). "A phylogenomic inventory of meiotic genes; evidence for sex in Giardia and an early eukaryotic origin of meiosis". Текущая биология. 15 (2): 185–191. Дои:10.1016/j.cub.2005.01.003. PMID 15668177. S2CID 17013247.
- ^ а б Malik SB, Pightling AW, Stefaniak LM, Schurko AM, Logsdon JM (August 2007). Hahn MW (ed.). "An expanded inventory of conserved meiotic genes provides evidence for sex in Trichomonas vaginalis". PLOS ONE. 3 (8): e2879. Bibcode:2008PLoSO...3.2879M. Дои:10.1371/journal.pone.0002879. ЧВК 2488364. PMID 18663385.
- ^ Akopyants NS, Kimblin N, Secundino N, Patrick R, Peters N, Lawyer P, Dobson DE, Beverley SM, Sacks DL (April 2009). "Demonstration of genetic exchange during cyclical development of Leishmania in the sand fly vector". Наука. 324 (5924): 265–268. Bibcode:2009Sci...324..265A. Дои:10.1126/science.1169464. ЧВК 2729066. PMID 19359589.
- ^ Лар DJ, Парфри Л.В., Митчелл Е.А., Кац Л.А., Лара Е. "The chastity of amoebae: re-evaluating evidence for sex in amoeboid organisms". Труды: Биологические науки.. 278 (1715): 2081–2090. Дои:10.1098/rspb.2011.0289. ЧВК 3107637. PMID 21429931.
- ^ Moore RT (1980). "Taxonomic proposals for the classification of marine yeasts and other yeast-like fungi including the smuts". Ботаника Марина. 23: 361–373.
- ^ Goldfuß (1818). "Ueber die Classification der Zoophyten" [On the classification of zoophytes]. Isis, Oder, Encyclopädische Zeitung von Oken (на немецком). 2 (6): 1008–1019. С п. 1008: "Erste Klasse. Urthiere. Protozoa." (First class. Primordial animals. Protozoa.) [Note: each column of each page of this journal is numbered; there are two columns per page.]
- ^ Scamardella JM (1999). «Не растения или животные: краткая история происхождения Kingdoms Protozoa, Protista и Protoctista» (PDF). Международная микробиология. 2 (4): 207–221. PMID 10943416. Архивировано из оригинал (PDF) 14 июня 2011 г.
- ^ а б Rothschild LJ (1989). «Простейшие, протисты, протоктисты: что в названии?». Журнал истории биологии. 22 (2): 277–305. Дои:10.1007 / BF00139515. PMID 11542176. S2CID 32462158.
- ^ а б Спанг А., Пила Дж. Х., Йоргенсен С. Л., Заремба-Недзведзка К., Мартейн Дж., Линд А. Е., ван Эйк Р., Шлепер К., Гай Л., Эттема Т. Дж. (Май 2015 г.). «Сложные археи, которые преодолевают разрыв между прокариотами и эукариотами». Природа. 521 (7551): 173–179. Bibcode:2015Натура.521..173S. Дои:10.1038 / природа14447. ЧВК 4444528. PMID 25945739.
- ^ а б Zaremba-Niedzwiedzka K, Caceres EF, Saw JH, Bäckström D, Juzokaite L, Vancaester E, Seitz KW, Anantharaman K, Starnawski P, Kjeldsen KU, Stott MB, Nunoura T, Banfield JF, Schramm A, Baker BJ, Spang A, Ettema TJ (January 2017). «Археи Асгарда проливают свет на происхождение эукариотической клеточной сложности». Природа. 541 (7637): 353–358. Bibcode:2017Натура.541..353Z. Дои:10.1038 / природа21031. OSTI 1580084. PMID 28077874. S2CID 4458094.
- ^ а б Liu Y, Zhou Z, Pan J, Baker BJ, Gu JD, Li M (April 2018). "Comparative genomic inference suggests mixotrophic lifestyle for Thorarchaeota". Журнал ISME. 12 (4): 1021–1031. Дои:10.1038/s41396-018-0060-x. ЧВК 5864231. PMID 29445130.
- ^ Adl SM, Simpson AG, Farmer MA, Andersen RA, Anderson OR, Barta JR, et al. (2005). «Новая классификация эукариот более высокого уровня с упором на таксономию простейших». Журнал эукариотической микробиологии. 52 (5): 399–451. Дои:10.1111 / j.1550-7408.2005.00053.x. PMID 16248873. S2CID 8060916.
- ^ Harper JT, Waanders E, Keeling PJ (January 2005). "On the monophyly of chromalveolates using a six-protein phylogeny of eukaryotes" (PDF). Международный журнал систематической и эволюционной микробиологии. 55 (Pt 1): 487–496. Дои:10.1099/ijs.0.63216-0. PMID 15653923. Архивировано из оригинал (PDF) 17 декабря 2008 г.
- ^ Parfrey LW, Barbero E, Lasser E, Dunthorn M, Bhattacharya D, Patterson DJ, Katz LA (December 2006). "Evaluating support for the current classification of eukaryotic diversity". PLOS Genetics. 2 (12): e220. Дои:10.1371 / journal.pgen.0020220. ЧВК 1713255. PMID 17194223.
- ^ а б Burki F (May 2014). "The eukaryotic tree of life from a global phylogenomic perspective". Перспективы Колд-Спринг-Харбор в биологии. 6 (5): a016147. Дои:10.1101/cshperspect.a016147. ЧВК 3996474. PMID 24789819.
- ^ а б Zhao S, Burki F, Bråte J, Keeling PJ, Klaveness D, Shalchian-Tabrizi K (June 2012). "Collodictyon – an ancient lineage in the tree of eukaryotes". Молекулярная биология и эволюция. 29 (6): 1557–1568. Дои:10.1093/molbev/mss001. ЧВК 3351787. PMID 22319147.
- ^ Romari K, Vaulot D (2004). "Composition and temporal variability of picoeukaryote communities at a coastal site of the English Channel from 18S rDNA sequences". Лимнол Океаногр. 49 (3): 784–798. Bibcode:2004LimOc..49..784R. Дои:10.4319/lo.2004.49.3.0784. S2CID 86718111.
- ^ Roger AJ, Simpson AG (February 2009). "Evolution: revisiting the root of the eukaryote tree". Текущая биология. 19 (4): R165–67. Дои:10.1016 / j.cub.2008.12.032. PMID 19243692. S2CID 13172971.
- ^ а б Sanders R (11 April 2016). "Wealth of unsuspected new microbes expands tree of life". Новости Беркли. Архивировано из оригинал 20 апреля 2016 г.. Получено 11 апреля 2016.
- ^ а б c Hug LA, Baker BJ, Anantharaman K, Brown CT, Probst AJ, Castelle CJ, Butterfield CN, Hernsdorf AW, Amano Y, Ise K, Suzuki Y, Dudek N, Relman DA, Finstad KM, Amundson R, Thomas BC, Banfield JF (April 2016). "A new view of the tree of life". Природная микробиология. 1 (5): 16048. Дои:10.1038/nmicrobiol.2016.48. PMID 27572647.
- ^ Tovar J, Fischer A, Clark CG (June 1999). "The mitosome, a novel organelle related to mitochondria in the amitochondrial parasite Entamoeba histolytica". Молекулярная микробиология. 32 (5): 1013–1021. Дои:10.1046/j.1365-2958.1999.01414.x. PMID 10361303. S2CID 22805284.
- ^ Boxma B, de Graaf RM, van der Staay GW, van Alen TA, Ricard G, Gabaldón T, van Hoek AH, Moon-van der Staay SY, Koopman WJ, van Hellemond JJ, Tielens AG, Friedrich T, Veenhuis M, Huynen MA, Hackstein JH (March 2005). «Анаэробная митохондрия, производящая водород» (PDF). Природа. 434 (7029): 74–79. Bibcode:2005Натура.434 ... 74Б. Дои:10.1038 / природа03343. PMID 15744302. S2CID 4401178.
- ^ а б Burki F, Shalchian-Tabrizi K, Minge M, Skjaeveland A, Nikolaev SI, Jakobsen KS, Pawlowski J (August 2007). Butler G (ed.). "Phylogenomics reshuffles the eukaryotic supergroups". PLOS ONE. 2 (8): e790. Bibcode:2007PLoSO ... 2..790B. Дои:10.1371 / journal.pone.0000790. ЧВК 1949142. PMID 17726520.
- ^ Burki F, Shalchian-Tabrizi K, Pawlowski J (August 2008). «Филогеномика открывает новую« мегагруппу », включающую большинство фотосинтезирующих эукариот». Письма о биологии. 4 (4): 366–369. Дои:10.1098 / рсбл.2008.0224. ЧВК 2610160. PMID 18522922.
- ^ Burki F, Inagaki Y, Bråte J, Archibald JM, Keeling PJ, Cavalier-Smith T, Sakaguchi M, Hashimoto T, Horak A, Kumar S, Klaveness D, Jakobsen KS, Pawlowski J, Shalchian-Tabrizi K (July 2009). "Large-scale phylogenomic analyses reveal that two enigmatic protist lineages, telonemia and centroheliozoa, are related to photosynthetic chromalveolates". Геномная биология и эволюция. 1: 231–238. Дои:10.1093 / gbe / evp022. ЧВК 2817417. PMID 20333193.
- ^ Hackett JD, Yoon HS, Li S, Reyes-Prieto A, Rümmele SE, Bhattacharya D (August 2007). "Phylogenomic analysis supports the monophyly of cryptophytes and haptophytes and the association of rhizaria with chromalveolates". Молекулярная биология и эволюция. 24 (8): 1702–1713. Дои:10.1093/molbev/msm089. PMID 17488740.
- ^ Кавалер-Смит Т (Июнь 2010 г.). «Царства Protozoa и Chromista и эозойный корень эукариотического дерева». Письма о биологии. 6 (3): 342–345. Дои:10.1098 / rsbl.2009.0948. ЧВК 2880060. PMID 20031978.
- ^ а б c Jagus R, Bachvaroff TR, Joshi B, Place AR (2012). "Diversity of Eukaryotic Translational Initiation Factor eIF4E in Protists". Сравнительная и функциональная геномика. 2012: 1–21. Дои:10.1155/2012/134839. ЧВК 3388326. PMID 22778692.
- ^ Burki F, Kaplan M, Tikhonenkov DV, Zlatogursky V, Minh BQ, Radaykina LV, Smirnov A, Mylnikov AP, Keeling PJ (January 2016). "Untangling the early diversification of eukaryotes: a phylogenomic study of the evolutionary origins of Centrohelida, Haptophyta and Cryptista". Труды: Биологические науки.. 283 (1823): 20152802. Дои:10.1098 / rspb.2015.2802. ЧВК 4795036. PMID 26817772.
- ^ Janouškovec J, Tikhonenkov DV, Burki F, Howe AT, Rohwer FL, Mylnikov AP, Keeling PJ (December 2017). "A New Lineage of Eukaryotes Illuminates Early Mitochondrial Genome Reduction" (PDF). Текущая биология. 27 (23): 3717–24.e5. Дои:10.1016 / j.cub.2017.10.051. PMID 29174886. S2CID 37933928.
- ^ Bodył A (февраль 2018 г.). «Некоторые пластиды, полученные из красных водорослей, эволюционировали посредством клептопластии? Гипотеза». Биологические обзоры Кембриджского философского общества. 93 (1): 201–222. Дои:10.1111 / brv.12340. PMID 28544184. S2CID 24613863.
- ^ а б Brown MW, Heiss AA, Kamikawa R, Inagaki Y, Yabuki A, Tice AK, Shiratori T, Ishida KI, Hashimoto T, Simpson AG, Roger AJ (February 2018). "Phylogenomics Places Orphan Protistan Lineages in a Novel Eukaryotic Super-Group". Геномная биология и эволюция. 10 (2): 427–433. Дои:10.1093/gbe/evy014. ЧВК 5793813. PMID 29360967.
- ^ Lax G, Eglit Y, Eme L, Bertrand EM, Roger AJ, Simpson AG (November 2018). "Hemimastigophora is a novel supra-kingdom-level lineage of eukaryotes". Природа. 564 (7736): 410–414. Bibcode:2018Natur.564..410L. Дои:10.1038/s41586-018-0708-8. PMID 30429611. S2CID 205570993.
- ^ Ирисарри, Икер; Strassert, Jürgen F. H.; Burki, Fabien (4 August 2020). "Phylogenomic Insights into the Origin of Primary Plastids". bioRxiv: 2020.08.03.231043. Дои:10.1101/2020.08.03.231043.
- ^ Strassert, Jürgen F. H.; Ирисарри, Икер; Уильямс, Том А .; Burki, Fabien (21 August 2020). "A molecular timescale for the origin of red algal-derived plastids". bioRxiv: 2020.08.20.259127. Дои:10.1101/2020.08.20.259127.
- ^ Бурки, Фабьен; Roger, Andrew J.; Браун, Мэтью У .; Simpson, Alastair G.B. (1 января 2020 г.). "The New Tree of Eukaryotes". Тенденции в экологии и эволюции. 35 (1): 43–55. Дои:10.1016/j.tree.2019.08.008. ISSN 0169-5347.
- ^ а б Kim E, Graham LE (July 2008). Redfield RJ (ed.). "Анализ EEF2 бросает вызов монофилии Archaeplastida и Chromalveolata". PLOS ONE. 3 (7): e2621. Bibcode:2008PLoSO...3.2621K. Дои:10.1371 / journal.pone.0002621. ЧВК 2440802. PMID 18612431.
- ^ Baurain D, Brinkmann H, Petersen J, Rodríguez-Ezpeleta N, Stechmann A, Demoulin V, Roger AJ, Burger G, Lang BF, Philippe H (July 2010). "Phylogenomic evidence for separate acquisition of plastids in cryptophytes, haptophytes, and stramenopiles". Молекулярная биология и эволюция. 27 (7): 1698–1709. Дои:10.1093/molbev/msq059. PMID 20194427.
- ^ Burki F, Okamoto N, Pombert JF, Keeling PJ (June 2012). "The evolutionary history of haptophytes and cryptophytes: phylogenomic evidence for separate origins". Труды: Биологические науки.. 279 (1736): 2246–2254. Дои:10.1098/rspb.2011.2301. ЧВК 3321700. PMID 22298847.
- ^ Кавалер-Смит Т (2006). "Protist phylogeny and the high-level classification of Protozoa". Европейский журнал протистологии. 39 (4): 338–348. Дои:10.1078/0932-4739-00002. S2CID 84403388.
- ^ Burki F, Pawlowski J (October 2006). "Monophyly of Rhizaria and multigene phylogeny of unicellular bikonts". Молекулярная биология и эволюция. 23 (10): 1922–1930. Дои:10.1093/molbev/msl055. PMID 16829542.
- ^ Ren R, Sun Y, Zhao Y, Geiser D, Ma H, Zhou X (September 2016). "Phylogenetic Resolution of Deep Eukaryotic and Fungal Relationships Using Highly Conserved Low-Copy Nuclear Genes". Геномная биология и эволюция. 8 (9): 2683–2701. Дои:10.1093/gbe/evw196. ЧВК 5631032. PMID 27604879.
- ^ а б c Cavalier-Smith T (January 2018). «Kingdom Chromista и его восемь типов: новый синтез, подчеркивающий нацеливание на перипластидные белки, эволюцию цитоскелета и перипластид, а также древние расхождения». Протоплазма. 255 (1): 297–357. Дои:10.1007 / s00709-017-1147-3. ЧВК 5756292. PMID 28875267.
- ^ Derelle R, Torruella G, Klimeš V, Brinkmann H, Kim E, Vlček Č, Lang BF, Eliáš M (February 2015). "Bacterial proteins pinpoint a single eukaryotic root". Труды Национальной академии наук Соединенных Штатов Америки. 112 (7): E693–699. Bibcode:2015PNAS..112E.693D. Дои:10.1073/pnas.1420657112. ЧВК 4343179. PMID 25646484.
- ^ Yang J, Harding T, Kamikawa R, Simpson AG, Roger AJ (May 2017). "Mitochondrial Genome Evolution and a Novel RNA Editing System in Deep-Branching Heteroloboseids". Геномная биология и эволюция. 9 (5): 1161–1174. Дои:10.1093/gbe/evx086. ЧВК 5421314. PMID 28453770.
- ^ Cavalier-Smith T, Fiore-Donno AM, Chao E, Kudryavtsev A, Berney C, Snell EA, Lewis R (February 2015). "Multigene phylogeny resolves deep branching of Amoebozoa". Молекулярная филогенетика и эволюция. 83: 293–304. Дои:10.1016/j.ympev.2014.08.011. PMID 25150787.
- ^ Torruella G, de Mendoza A, Grau-Bové X, Antó M, Chaplin MA, del Campo J, Eme L, Pérez-Cordón G, Whipps CM, Nichols KM, Paley R, Roger AJ, Sitjà-Bobadilla A, Donachie S, Ruiz-Trillo I (September 2015). «Филогеномика раскрывает конвергентную эволюцию образа жизни у близких родственников животных и грибов». Текущая биология. 25 (18): 2404–2410. Дои:10.1016 / j.cub.2015.07.053. PMID 26365255.
- ^ а б Лопес-Гарсия П., Эме Л., Морейра Д. (декабрь 2017 г.). «Симбиоз в эволюции эукариот». Журнал теоретической биологии. 434: 20–33. Дои:10.1016 / j.jtbi.2017.02.031. ЧВК 5638015. PMID 28254477.
- ^ Понсе-Толедо Р.И., Дешам П., Лопес-Гарсия П., Живанович Ю., Бензерара К., Морейра Д. (февраль 2017 г.). «Пресноводная цианобактерия с ранним ветвлением и у истоков пластид». Текущая биология. 27 (3): 386–391. Дои:10.1016 / j.cub.2016.11.056. ЧВК 5650054. PMID 28132810.
- ^ де Фриз Дж, Арчибальд Дж. М. (февраль 2017 г.). «Эндосимбиоз: развились ли пластиды из пресноводных цианобактерий?». Текущая биология. 27 (3): R103–105. Дои:10.1016 / j.cub.2016.12.006. PMID 28171752.
- ^ а б Кавальер-Смит Т. (июнь 2010 г.). «Царства Protozoa и Chromista и эозойный корень эукариотического дерева». Письма о биологии. 6 (3): 342–345. Дои:10.1098 / rsbl.2009.0948. ЧВК 2880060. PMID 20031978.
- ^ а б Cavalier-Smith T (May 2013). «Ранняя эволюция режимов питания эукариот, структурное разнообразие клеток и классификация простейших типов Loukozoa, Sulcozoa и Choanozoa». Европейский журнал протистологии. 49 (2): 115–178. Дои:10.1016 / j.ejop.2012.06.001. PMID 23085100.
- ^ а б Cavalier-Smith T, Chao EE, Snell EA, Berney C, Fiore-Donno AM, Lewis R (December 2014). "Multigene eukaryote phylogeny reveals the likely protozoan ancestors of opisthokonts (animals, fungi, choanozoans) and Amoebozoa". Молекулярная филогенетика и эволюция. 81: 71–85. Дои:10.1016/j.ympev.2014.08.012. PMID 25152275.
- ^ Cavalier-Smith T, Chao EE, Lewis R (April 2018). "Multigene phylogeny and cell evolution of chromist infrakingdom Rhizaria: contrasting cell organisation of sister phyla Cercozoa and Retaria". Протоплазма. 255 (5): 1517–1574. Дои:10.1007 / s00709-018-1241-1. ЧВК 6133090. PMID 29666938.
- ^ He D, Fiz-Palacios O, Fu CJ, Fehling J, Tsai CC, Baldauf SL (February 2014). "An alternative root for the eukaryote tree of life". Текущая биология. 24 (4): 465–470. Дои:10.1016/j.cub.2014.01.036. PMID 24508168.
- ^ Cox CJ, Foster PG, Hirt RP, Harris SR, Embley TM (December 2008). «Архебактериальное происхождение эукариот». Труды Национальной академии наук Соединенных Штатов Америки. 105 (51): 20356–20361. Bibcode:2008PNAS..10520356C. Дои:10.1073 / pnas.0810647105. ЧВК 2629343. PMID 19073919.
- ^ Ciccarelli FD, Doerks T., von Mering C, Creevey CJ, Snel B, Bork P (март 2006 г.). «К автоматической реконструкции дерева жизни с высоким разрешением». Наука. 311 (5765): 1283–1287. Bibcode:2006Научный ... 311.1283C. CiteSeerX 10.1.1.381.9514. Дои:10.1126 / science.1123061. PMID 16513982. S2CID 1615592.
- ^ O’Malley, Maureen A.; Leger, Michelle M.; Wideman, Джереми Дж .; Ruiz-Trillo, Iñaki (18 February 2019). "Concepts of the last eukaryotic common ancestor". Природа Экология и эволюция. ООО "Спрингер Сайенс энд Бизнес Медиа". 3 (3): 338–344. Дои:10.1038/s41559-019-0796-3. HDL:10261/201794. ISSN 2397-334X. PMID 30778187. S2CID 67790751.
- ^ а б Brocks JJ, Logan GA, Buick R, Summons RE (август 1999 г.). «Архейские молекулярные окаменелости и ранний рост эукариот». Наука. 285 (5430): 1033–1036. CiteSeerX 10.1.1.516.9123. Дои:10.1126 / science.285.5430.1033. PMID 10446042.
- ^ а б Hartman H, Fedorov A (February 2002). "The origin of the eukaryotic cell: a genomic investigation". Труды Национальной академии наук Соединенных Штатов Америки. 99 (3): 1420–5. Bibcode:2002PNAS...99.1420H. Дои:10.1073/pnas.032658599. ЧВК 122206. PMID 11805300.
- ^ Knoll AH, Javaux EJ, Hewitt D, Cohen P (June 2006). «Эукариотические организмы в протерозойских океанах». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 361 (1470): 1023–1038. Дои:10.1098 / rstb.2006.1843. ЧВК 1578724. PMID 16754612.
- ^ Retallack GJ, Krull ES, Thackray GD, Parkinson DH (2013). «Проблемные окаменелости в форме урн из палеопротерозойских (2,2 млрд лет) палеопочв в Южной Африке». Докембрийские исследования. 235: 71–87. Bibcode:2013PreR..235 ... 71R. Дои:10.1016 / j.precamres.2013.05.015.
- ^ El Albani A, Bengtson S, Canfield DE, Bekker A, Macchiarelli R, Mazurier A, Hammarlund EU, Boulvais P, Dupuy JJ, Fontaine C, Fürsich FT, Gauthier-Lafaye F, Janvier P, Javaux E, Ossa FO, Pierson-Wickmann AC, Riboulleau A, Sardini P, Vachard D, Whitehouse M, Meunier A (July 2010). «Крупные колониальные организмы с координированным ростом в насыщенной кислородом среде 2,1 млрд лет назад». Природа. 466 (7302): 100–104. Bibcode:2010Натура.466..100А. Дои:10.1038 / природа09166. PMID 20596019. S2CID 4331375.
- ^ Bengtson S, Belivanova V, Rasmussen B, Whitehouse M (May 2009). «Спорные« кембрийские »окаменелости Виндхьяна реальны, но более чем на миллиард лет старше». Труды Национальной академии наук Соединенных Штатов Америки. 106 (19): 7729–7734. Bibcode:2009PNAS..106.7729B. Дои:10.1073 / pnas.0812460106. ЧВК 2683128. PMID 19416859.
- ^ Ward P (9 февраля 2008 г.). "Mass extinctions: the microbes strike back". Новый ученый: 40–43.
- ^ French KL, Hallmann C, Hope JM, Schoon PL, Zumberge JA, Hoshino Y, Peters CA, George SC, Love GD, Brocks JJ, Buick R, Summons RE (May 2015). "Reappraisal of hydrocarbon biomarkers in Archean rocks". Труды Национальной академии наук Соединенных Штатов Америки. 112 (19): 5915–5920. Bibcode:2015PNAS..112.5915F. Дои:10.1073/pnas.1419563112. ЧВК 4434754. PMID 25918387.
- ^ Isson TT, Love GD, Dupont CL, Reinhard CT, Zumberge AJ, Asael D, et al. (Июнь 2018). "Tracking the rise of eukaryotes to ecological dominance with zinc isotopes". Геобиология. 16 (4): 341–352. Дои:10.1111/gbi.12289. PMID 29869832.
- ^ Yoshikawa G, Blanc-Mathieu R, Song C, Kayama Y, Mochizuki T, Murata K, Ogata H, Takemura M (April 2019). "Medusavirus, a Novel Large DNA Virus Discovered from Hot Spring Water". Журнал вирусологии. 93 (8). Дои:10.1128 / JVI.02130-18. ЧВК 6450098. PMID 30728258. Сложить резюме – EurekAlert! (30 апреля 2019 г.).
- ^ Мартин В. (декабрь 2005 г.). «Архебактерии (Археи) и происхождение ядра эукариот». Текущее мнение в микробиологии. 8 (6): 630–637. Дои:10.1016 / j.mib.2005.10.004. PMID 16242992.
- ^ Такемура М (май 2001 г.). «Поксвирусы и происхождение ядра эукариот». Журнал молекулярной эволюции. 52 (5): 419–425. Bibcode:2001JMolE..52..419T. Дои:10.1007 / s002390010171. PMID 11443345. S2CID 21200827.
- ^ Белл PJ (сентябрь 2001 г.). «Вирусный эукариогенез: был ли предком ядра сложный ДНК-вирус?». Журнал молекулярной эволюции. 53 (3): 251–256. Bibcode:2001JMolE..53..251L. Дои:10.1007 / s002390010215. PMID 11523012. S2CID 20542871.
- ^ Wächtershäuser G (январь 2003 г.). «От предклеток до эукарии - сказка о двух липидах». Молекулярная микробиология. 47 (1): 13–22. Дои:10.1046 / j.1365-2958.2003.03267.x. PMID 12492850. S2CID 37944519.
- ^ Wächtershäuser G (октябрь 2006 г.). «От вулканического происхождения хемоавтотрофной жизни до бактерий, архей и эукарий». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 361 (1474): 1787–1806, обсуждение 1806–1808. Дои:10.1098 / rstb.2006.1904. ЧВК 1664677. PMID 17008219.
- ^ Лейн, Ник (2016). Главный вопрос: почему жизнь такая, какая она есть? (под ред. в мягкой обложке). Профильные книги. С. 157–91. ISBN 978-1-781-25037-2.
- ^ Эгель Р. (январь 2012 г.). «Первичный эукариогенез: о коллективной природе доклеточных состояний, предков современной жизни». Жизнь. 2 (1): 170–212. Дои:10.3390 / жизнь2010170. ЧВК 4187143. PMID 25382122.
- ^ а б Хариш А., Тунлид А., Курланд К.Г. (август 2013 г.). «Укоренившаяся филогения трех сверхцарств». Биохимия. 95 (8): 1593–1604. Дои:10.1016 / j.biochi.2013.04.016. PMID 23669449.
- ^ а б Хариш А., Курляндская CG (июль 2017 г.). «Акариоты и эукариоты - независимые потомки универсального общего предка». Биохимия. 138: 168–183. Дои:10.1016 / j.biochi.2017.04.013. PMID 28461155.
- ^ а б Имачи Х., Нобу М.К., Накахара Н., Мороно Й., Огавара М., Такаки Й. и др. (Январь 2020 г.). «Изоляция архея на границе прокариот-эукариот». Природа. 577 (7791): 519–525. Bibcode:2020Натура.577..519I. Дои:10.1038 / s41586-019-1916-6. ЧВК 7015854. PMID 31942073.
- ^ Да Кунья В., Гайя М., Гадель Д., Насир А., Фортер П. (июнь 2017 г.). «Локиархеи - близкие родственники Euryarchaeota, не ликвидирующие разрыв между прокариотами и эукариотами». PLOS Genetics. 13 (6): e1006810. Дои:10.1371 / journal.pgen.1006810. ЧВК 5484517. PMID 28604769.
- ^ Хариш А., Курляндская CG (июль 2017 г.). «Эмпирические модели эволюции генома укореняют древо жизни». Биохимия. 138: 137–155. Дои:10.1016 / j.biochi.2017.04.014. PMID 28478110.
- ^ Spang A, Eme L, Saw JH, Caceres EF, Zaremba-Niedzwiedzka K, Lombard J, et al. (Март 2018 г.). «Асгарские археи - ближайшие прокариотические родственники эукариот». PLOS Genetics. 14 (3): e1007080. Дои:10.1371 / journal.pgen.1007080. ЧВК 5875740. PMID 29596421.
- ^ Маклеод Ф., Киндлер Г.С., Вонг Х.Л., Чен Р., Бернс Б.П. (2019). «Археи Асгарда: разнообразие, функции и эволюционные последствия для ряда микробиомов». AIMS Microbiology. 5 (1): 48–61. Дои:10.3934 / microbiol.2019.1.48. ЧВК 6646929. PMID 31384702.
- ^ Циммер, Карл (15 января 2020 г.). «Этот странный микроб может означать один из больших скачков жизни - организм, живущий в океанской иле, дает ключ к разгадке происхождения сложных клеток всех животных и растений». Нью-Йорк Таймс. Получено 18 января 2020.
- ^ Джекели Джи (2007). «Происхождение эукариотических эндомембран: критическая оценка различных модельных сценариев». Эукариотические мембраны и цитоскелет. Успехи экспериментальной медицины и биологии. 607. Нью-Йорк, Нью-Йорк: Springer Science + Business Media; Остин, Техас: Landes Bioscience. стр.38–51. Дои:10.1007/978-0-387-74021-8_3. ISBN 978-0-387-74020-1. PMID 17977457.
- ^ Кавальер-Смит Т. (март 2002 г.). «Фаготрофное происхождение эукариот и филогенетическая классификация простейших». Международный журнал систематической и эволюционной микробиологии. 52 (Pt 2): 297–354. Дои:10.1099/00207713-52-2-297. PMID 11931142.
- ^ а б Мартин В., Мюллер М. (март 1998 г.). «Гипотеза водорода для первого эукариота». Природа. 392 (6671): 37–41. Bibcode:1998Натура.392 ... 37М. Дои:10.1038/32096. PMID 9510246. S2CID 338885.
- ^ Пизани Д., Коттон Дж. А., Макинерни Дж. О. (август 2007 г.). «Супердеревья раскрывают химерическое происхождение геномов эукариот». Молекулярная биология и эволюция. 24 (8): 1752–1760. Дои:10.1093 / молбев / msm095. PMID 17504772.
- ^ Брюкнер Дж., Мартин В.Ф. (апрель 2020 г.). «Бактериальные гены превосходят по численности гены архей в эукариотических геномах». Геномная биология и эволюция. 12 (4): 282–292. Дои:10.1093 / gbe / evaa047. ЧВК 7151554. PMID 32142116.
- ^ а б Латорре А, Дурбан А, Моя А, Перето Дж (2011). «Роль симбиоза в эволюции эукариот». В Gargaud M, López-Garcìa P, Martin H (ред.). Происхождение и эволюция жизни: астробиологическая перспектива. Кембридж: Издательство Кембриджского университета. С. 326–339. ISBN 978-0-521-76131-4.
- ^ Аяла Дж. (Апрель 1994 г.). «Транспорт и внутренняя организация мембран: везикулы, мембранные сети и GTP-связывающие белки». Журнал клеточной науки. 107 (Pt 4) (107): 753–763. PMID 8056835. Архивировано из оригинал 29 апреля 2012 г.. Получено 27 марта 2013.
- ^ Мартин В.Ф. «Происхождение митохондрий». Scitable. Природное образование. Получено 27 марта 2013.
- ^ Dacks JB, Field MC (август 2018). «Эволюционное происхождение и специализация мембранного транспорта». Текущее мнение в области клеточной биологии. 53: 70–76. Дои:10.1016 / j.ceb.2018.06.001. ЧВК 6141808. PMID 29929066.
- ^ Мартейн Дж., Фоссеберг Дж., Гай Л., Оффре П., Эттема Т. Дж. (Май 2018 г.). «Глубокое митохондриальное происхождение за пределами отобранных альфа-протеобактерий». Природа. 557 (7703): 101–105. Bibcode:2018Натура.557..101M. Дои:10.1038 / s41586-018-0059-5. PMID 29695865. S2CID 13740626. Сложить резюме – Ученый.
- ^ Баум Д.А., Баум Б. (октябрь 2014 г.). «Вывернутое наизнанку происхождение эукариотической клетки». BMC Биология. 12: 76. Дои:10.1186 / s12915-014-0076-2. ЧВК 4210606. PMID 25350791. Сложить резюме – Университет Висконсин-Мэдисон.
- ^ Брауэрс Л. (12 апреля 2013 г.). «Как генетическое ограбление превратило микроб в розового соленого падальщика». Scientific American. 109 (50): 20537–20542. Архивировано из оригинал 10 октября 2018 г.. Получено 21 апреля 2019.
- ^ Маргулис Л (1970). Происхождение эукариотических клеток. Нью-Хейвен, Лондон: Издательство Йельского университета.
- ^ Маргулис Л (1993). Симбиоз в эволюции клетки. Нью-Йорк: W.H. Фримен.
- ^ Маргулис Л., Долан М. Ф., Герреро Р. (июнь 2000 г.). «Химерный эукариот: происхождение ядра от кариомастигонта у амитохондриальных протистов». Труды Национальной академии наук Соединенных Штатов Америки. 97 (13): 6954–6959. Bibcode:2000PNAS ... 97.6954M. Дои:10.1073 / пнас.97.13.6954. ЧВК 34369. PMID 10860956.
- ^ Морейра Д., Лопес-Гарсия П. (ноябрь 1998 г.). «Симбиоз между метаногенными археями и дельта-протеобактериями как происхождение эукариот: синтрофическая гипотеза». Журнал молекулярной эволюции. 47 (5): 517–530. Bibcode:1998JMolE..47..517M. Дои:10.1007 / PL00006408. PMID 9797402. S2CID 3911443.
- ^ Лопес-Гарсия П., Морейра Д. (май 2006 г.). «Селективные силы для происхождения эукариотического ядра». BioEssays. 28 (5): 525–533. Дои:10.1002 / bies.20413. PMID 16615090.
- ^ «Переосмысление происхождения эукариот». Журнал Quanta. Архивировано из оригинал 9 мая 2019 г.. Получено 8 мая 2019.
- ^ Питтис А.А., Габальдон Т. (март 2016 г.). «Позднее приобретение митохондрий хозяином с химерными прокариотическими предками». Природа. 531 (7592): 101–104. Bibcode:2016Натура.531..101П. Дои:10.1038 / природа16941. ЧВК 4780264. PMID 26840490.
- ^ Burton ZF (1 августа 2017 г.). Эволюция с момента написания кода: люльки, нимбы, стволы и крылья.. Академическая пресса. ISBN 9780128130346.
- ^ Мартин В.Ф., Рёттгер М., Ку К., Гарг С.Г., Нельсон-Сати С., Ландан Г. (февраль 2017 г.). «Позднее митохондриальное происхождение - артефакт». Геномная биология и эволюция. 9 (2): 373–379. Дои:10.1093 / gbe / evx027. ЧВК 5516564. PMID 28199635.
![]() Эта статья включаетматериалы общественного достояния от NCBI документ: «Учебник по науке».
Эта статья включаетматериалы общественного достояния от NCBI документ: «Учебник по науке».
внешняя ссылка
- «Эукариоты» (Веб-проект "Древо жизни" )
- «Эукариот» на Энциклопедия жизни

- Влечение и секс среди наших последних общих предков-эукариот-микробов, Атлантика, 11 ноября 2020 г.
#invoke: Navbox#invoke: Navbox#invoke: Navbox#invoke: Taxonbar#invoke: Авторитетный контроль