Клептопластика - Kleptoplasty

C = хлоропласт,
N = ядро клетки.
Электронная микрофотография: масштабная линейка 3 мкм.
Клептопластика или клептопластидия это симбиотический явление, посредством которого пластиды, особенно хлоропласты от водоросли, секвестрируются организмами-хозяевами. Слово происходит от Клептес (κλέπτης), который является Греческий для вор. Водоросль поедается нормально и частично переваривается, оставляя неповрежденной пластиду. Пластиды сохраняются внутри хозяина, временно продолжая фотосинтез и принося пользу хищнику.[1] Этот термин был придуман в 1990 году для описания симбиоза хлоропластов.[2][3]
Инфузории
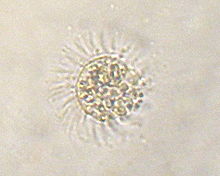
Mesodinium rubrum это инфузория крадет хлоропласты из криптомонада Геминигера криофила.[4] М. rubrum участвует в дополнительном эндосимбиозе, передавая свои пластиды своим хищникам, планктонам динофлагеллят, принадлежащим к роду Динофиз.[5]
Кариоклепты - это связанный процесс, в котором ядро клетки-жертвы также сохраняется за хищником. Впервые это было описано в М. rubrum.[6]
Динофлагелляты
Стабильность временных пластид значительно различается у разных видов, сохраняющих пластиды. в динофлагелляты Гимнодиниум виды и Pfisteria piscicida, клептопластиды фотосинтетически активны всего несколько дней, в то время как клептопластиды в Динофиз виды может быть стабильным в течение 2 месяцев.[1] Предполагается, что у других динофлагеллят клептопластика представляет собой либо механизм, обеспечивающий функциональную гибкость, либо, возможно, ранний эволюционный этап в постоянном приобретении хлоропластов.[7]
Сакоглоссанские слизни

Одними из немногих известных животных, которые практикуют клептопластику, являются морские слизни в кладе Сакоглосса.[8] Некоторые виды морских слизней Sacoglossan захватывают неповрежденные функциональные хлоропласты из пищевых источников водорослей, удерживая их в специализированных клетках, выстилающих моллюск пищеварительный дивертикулы. Самая продолжительная известная ассоциация клептопластики, которая может длиться до десяти месяцев, обнаружена у Elysia chlorotica,[2] который приобретает хлоропласты, поедая водоросли Vaucheria litorea, храня хлоропласты в клетках, выстилающих кишечник.[9] Молодые морские слизни вызывают клептопластический эндосимбиоз, питаясь клетками водорослей, высасывая их содержимое и выбрасывая все, кроме хлоропластов. Хлоропласты фагоцитированный пищеварительными клетками, заполняя широко разветвленные пищеварительные канальцы, обеспечивая своего хозяина продуктами фотосинтеза.[10] Однако не решено, секретируют ли украденные пластиды активно фотосинтат или слизни косвенно извлекают выгоду из медленно разрушающихся клептопластов.[11]

Из-за этой необычной способности сакоглоссанов иногда называют «морскими слизнями, работающими на солнечной энергии», хотя фактическая польза от фотосинтеза для выживания некоторых из проанализированных видов кажется в лучшем случае незначительной.[12] Фактически, некоторые виды могут даже погибнуть в присутствии клептопластов, фиксирующих диоксид углерода, в результате повышенных уровней активных форм кислорода.[13]
Было показано, что изменения температуры отрицательно влияют на клептопластические способности сакоглоссана. Показано, что показатели эффективности фотосинтеза, а также численность клептопластов снижаются в корреляции с понижением температуры. Однако характер и скорость этих изменений различаются у разных видов морских слизней.[14]
Голожаберники
Некоторые виды голожаберники такие как Pteraeolidia ianthina секвестр всего живого симбиотика зооксантеллы в их пищеварительных дивертикулах и, следовательно, также "питаются солнечной энергией".
Фораминиферы
Некоторые виды фораминифер роды Булимина, Эльфидиум, Хайнесина, Неион, Nonionella, Нонионеллина, Реофакс, и Stainforthia были показаны секвестру диатомовый хлоропласты.[15]
Плоские черви Rhabdocoel
Два вида из группы морских плоских червей, известных как рабдокоэльс, Baicalellia solaris и Pogaina paranygulgus, было обнаружено, что они представляют собой другой тип животных, у которых наблюдается клептопластика. Группа ранее была классифицирована как имеющая эндосимбионтов водорослей, хотя уже было обнаружено, что эндосимбионты не содержат ядер.[16]
Потребляя диатомовые водоросли, Б. солярис и P. paranygulusв процессе, который еще не открыт, извлекают пластиды из их добычи, включая их надэпидермально, разделяя и переваривая панцирь и остальную часть диатомовых водорослей. В Б. солярис извлеченные пластиды, или клептопласты, продолжают проявлять функциональный фотосинтез в течение короткого периода примерно 7 дней. Поскольку эти две группы не являются сестринскими таксонами, и этот признак не является общим для более близких групп, есть свидетельства того, что клептопластика развивалась независимо внутри этих двух таксонов.[17]
Смотрите также
использованная литература
- ^ а б Миннхаген С., Карвалью В. Ф., Саломон П. С., Янсон С. (сентябрь 2008 г.). «Содержание ДНК хлоропластов в Dinophysis (Dinophyceae) на разных стадиях клеточного цикла согласуется с клептопластикой». Environ. Микробиол. 10 (9): 2411–7. Дои:10.1111 / j.1462-2920.2008.01666.x. PMID 18518896.
- ^ а б С. К. Пирс; С. Э. Мэсси; J. J. Hanten; Н. Э. Кертис (1 июня 2003 г.). «Горизонтальный перенос функциональных ядерных генов между многоклеточными организмами». Биол. Бык. 204 (3): 237–240. Дои:10.2307/1543594. JSTOR 1543594. PMID 12807700. S2CID 9671982. Получено 2008-11-24.
- ^ Кларк, К. Б.; Jensen, K. R .; Стриц, Х. М. (1990). «Исследование функциональной клептопластики среди аскоглоссы Западной Атлантики (= Sacoglossa) (Mollusca: Opisthobranchia)». Велигер. 33: 339–345. ISSN 0042-3211. S2CID 87182226.
- ^ Джонсон, Мэтью Д .; Олдах, Дэвид; Чарльз, Ф. Делвиче; Стокер, Дайан К. (январь 2007 г.). «Удержание транскрипционно активных ядер криптофита реснитчатым Myrionecta rubra». Природа. 445 (7126): 426–8. Bibcode:2007Натура.445..426J. Дои:10.1038 / природа05496. PMID 17251979. S2CID 4410812.
- ^ Nishitani, G .; Nagai, S .; Баба, К .; Киёкава, S .; Kosaka, Y .; Миямура, К .; Nishikawa, T .; Сакурада, К .; Shinada, A .; Камияма, Т. (2010). "Соответствие высокого уровня Myrionecta rubra добыча и Динофиз видовые пластидные идентичности, выявленные генетическим анализом изолятов из прибрежных вод Японии ". Прикладная и экологическая микробиология. 76 (9): 2791–2798. Дои:10.1128 / AEM.02566-09. ЧВК 2863437. PMID 20305031.
- ^ Джонсон, Мэтью Д .; Олдах, Дэвид; и другие. (25 января 2007 г.). «Удержание транскрипционно активных ядер криптофита инфузорией Myrionecta rubra". Природа. 445 (7126): 426–428. Bibcode:2007Натура.445..426J. Дои:10.1038 / природа05496. PMID 17251979. S2CID 4410812. Получено 4 февраля 2015.
- ^ Гаст Р.Дж., Моран Д.М., Деннетт М.Р., Карон Д.А. (январь 2007 г.). «Клептопластика у динофлагеллат Антарктики: в процессе эволюционного перехода?». Environ. Микробиол. 9 (1): 39–45. CiteSeerX 10.1.1.490.54. Дои:10.1111 / j.1462-2920.2006.01109.x. PMID 17227410.
- ^ Händeler, K .; Grzymbowski, Y.P .; Krug, P.J .; Вегеле, Х. (2009). «Функциональные хлоропласты в клетках многоклеточных животных - уникальная эволюционная стратегия в жизни животных». Границы зоологии. 6: 28. Дои:10.1186/1742-9994-6-28. ЧВК 2790442. PMID 19951407.
- ^ Екатерина Брахич (24 ноября 2008 г.). «Морской слизняк на солнечных батареях использует украденные гены растений». Новый ученый. Получено 24 ноября 2008.
- ^ «СимБио: Введение-Клептопластика». Университет штата Мэн. Архивировано из оригинал на 2008-12-02. Получено 2008-11-24.
- ^ де Фрис, Ян; Криста, Грегор; Гулд, Свен Б. (2014). «Выживание пластидов в цитозоле клеток животных». Тенденции в растениеводстве. 19 (6): 347–350. Дои:10.1016 / j.tplants.2014.03.010. ISSN 1360-1385. PMID 24767983.
- ^ Де Врис, Ян; Rauch, Cessa; Криста, Грегор; Гулд, Свен Б. (2014). "Гид по пластидному симбиозу". Acta Societatis Botanicorum Poloniae. 83 (4): 415–421. Дои:10.5586 / asbp.2014.042. ISSN 2083-9480.
- ^ de Vries, J .; Woehle, C .; Christa, G .; Wagele, H .; Tielens, A.G.M .; Jahns, P .; Гулд, С. Б. (2015). «Сравнение родственных видов выявляет факторы, лежащие в основе совместимости пластид у зеленых морских слизней». Труды Королевского общества B: биологические науки. 282 (1802): 20142519. Дои:10.1098 / rspb.2014.2519. ISSN 0962-8452. ЧВК 4344150. PMID 25652835.
- ^ Лаец, Элиза; Вагеле, Хайке (2018). «Как температура влияет на функциональную клептопластику? Сравнение популяций видов-сестер, работающих на солнечной энергии, Elysia timida Risso, 1818 и Elysa cornigera Nuttall, 1989 (Gastopoda: Sacoglossa)». Границы зоологии. 15: 17. Дои:10.1186 / s12983-018-0264-у. ЧВК 5937827. PMID 29760759.
- ^ Бернхард, Джоан М .; Баузер, Сэмюэл С. (1999). «Бентические фораминиферы дизоксических отложений: секвестрация хлоропластов и функциональная морфология». Обзоры наук о Земле. 46 (1–4): 149–165. Bibcode:1999ESRv ... 46..149B. Дои:10.1016 / с0012-8252 (99) 00017-3.
- ^ Э. Маркус, Turbellaria Brasileiros (9). Бол. Фак. Fil. Ci. Letras Univ. Сан-Паулу 16, 5–215 (1951).
- ^ Van Steenkiste, Niels W. L .; Стефенсон, Индия; Эрранц, Мария; Хусник, Филип; Килинг, Патрик Дж .; Леандер, Брайан С. (2019). «Новый случай клептопластики у животных: морские плоские черви крадут функциональные пластиды у диатомовых водорослей». Достижения науки. 5 (7): eaaw4337. Bibcode:2019SciA .... 5.4337V. Дои:10.1126 / sciadv.aaw4337.
внешние ссылки
- "Морские слизни на солнечных батареях". ABC Science Online. Июнь 2007 г.. Получено 2008-11-24.