Диатомовый - Diatom
| Диатомеи | |
|---|---|
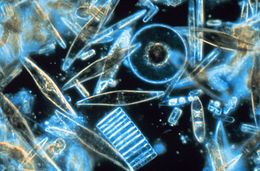 | |
| Световая микроскопия образцов морских диатомовых водорослей, обитающих между кристаллами годового морского льда в Антарктиде, демонстрирует разнообразие размеров, форм и цветов. | |
| Научная классификация | |
| Clade: | SAR |
| Тип: | Охрофита |
| Подтип: | Хакиста |
| Класс: | Bacillariophyceae Dangeard, 1933[1] |
| Синонимы | |
| |
Диатомеи (диа-том-ос "разрезать пополам", от диа, «через» или «отдельно»; и корень tém-n-ō, 'Я режу'.)[6] являются основной группой водоросли,[7] конкретно микроводоросли, найденные в океанах, водных путях и почвах мира. Живые диатомовые водоросли составляют значительную часть земного биомасса: они генерируют от 20 до 50 процентов кислород производятся на планете каждый год,[8][9] принять более 6,7 млрд метрических тонн кремний каждый год из вод, в которых они живут,[10] и составляют почти половину органического материала, обнаруженного в океанах. Раковины мертвых диатомовых водорослей могут достигать глубины 800 м на дне океана, и весь Бассейн Амазонки удобряется ежегодно 27 миллионами тонн диатомовой пыли, переносимой трансатлантическими ветрами с африканских Сахара, большая часть из Депрессия Боделе, который когда-то представлял собой систему пресноводных озер.[11][12]
Диатомовые водоросли одноклеточный: они встречаются либо в одиночных камерах, либо в колонии, которые могут иметь форму лент, вееров, зигзагов или звезд. Размер отдельных ячеек составляет от 2 до 200 микрометров.[13] При наличии достаточного количества питательных веществ и солнечного света количество живых диатомовых водорослей удваивается примерно каждые 24 часа на бесполый множественное деление; максимальная продолжительность жизни отдельных клеток составляет около шести дней.[14] Диатомовые водоросли имеют две различные формы: несколько (центрические диатомеи) находятся радиально симметричный, а большинство (пеннатные диатомеи) в целом двусторонне симметричный. Уникальная особенность диатомовой водоросли анатомия в том, что они окружены клеточная стенка сделано из кремнезем (гидратированный диоксид кремния), называемый панцирь.[15] Эти панцири имеют структурная окраска из-за их фотонный наноструктура, что побудило их называть «драгоценностями моря» и «живыми опалами». Движение диатомовых водорослей в основном происходит пассивно в результате обоих факторов. водные течения и ветровая вода турбулентность; однако мужчина гаметы центрических диатомей имеют жгутики, позволяющие активно двигаться в поисках женских гамет. Похожий на растения, диатомеи конвертируют энергия света к химическая энергия от фотосинтез, хотя это общее автотрофия развился независимо в обоих родословная. Необычно для автотрофных организмов, диатомеи обладают цикл мочевины, функция, с которой они делятся животные, хотя у диатомовых водорослей этот цикл используется для разных метаболических целей. Семья Rhopalodiaceae также обладают цианобактериальный эндосимбионт называется сфероидным телом. Этот эндосимбионт утратил фотосинтетические свойства, но сохранил способность выполнять азотфиксация, позволяя диатомею фиксировать атмосферный азот.[16]
Изучение диатомовых водорослей - это раздел психология. Диатомовые водоросли классифицируются как эукариоты, организмы с мембраносвязанный ядро клетки, что отделяет их от прокариоты археи и бактерии. Диатомовые водоросли - это разновидность планктон называется фитопланктон, самый распространенный из видов планктона. Диатомовые водоросли также прикрепляются к бентосный субстратах, плавающем мусоре и на макрофиты. Они составляют неотъемлемую часть перифитон сообщество.[17] Другая классификация делит планктон на восемь типов в зависимости от размера: в этой схеме диатомовые водоросли классифицируются как микроводоросли. Несколько систем для классификации отдельных диатомовых водорослей виды существует. Ископаемое данные свидетельствуют о том, что диатомовые водоросли возникли во время или до Юрский период, что было от 150 до 200 миллионов лет назад. Древнейшее ископаемое свидетельство диатомовых водорослей - это образец существующего рода Hemiaulus в позднеюрском янтаре из Таиланда.[18]
Диатомовые водоросли используются для мониторинга прошлых и настоящих условий окружающей среды и обычно используются для изучения качества воды. Кизельгур (диатомит) представляет собой набор раковин диатомовых водорослей, обнаруженных в земной коре. Это мягкие осадочные породы, содержащие кремнезем, которые легко крошатся в мелкий порошок и обычно имеют размер частиц от 10 до 200 мкм. Кизельгур используется для различных целей, в том числе для фильтрации воды, в качестве мягкого абразива, в наполнителе для кошачьих туалетов и в качестве стабилизатора динамита.

Отображает наложения с четырех флуоресцентных каналов
(b) Голубой: [флуоресценция PLL-A546] - общий контраст для визуализации поверхностей эукариотических клеток.
(c) Синий: [флуоресценция Hoechst] - окрашивает ДНК, идентифицирует ядра.
(d) Красный: [автофлуоресценция хлорофилла] - рассасывает хлоропласты.[19]

Структура
Диатомовые водоросли обычно имеют размер от 2 до 200 микрометров,[13] с несколькими более крупными видами. Их желтовато-коричневый хлоропласты, место фотосинтеза, типичны для гетероконты, имея четыре мембраны и содержащий пигменты такой как каротиноид фукоксантин. Людям обычно не хватает жгутики, но они присутствуют в мужском гаметы центрических диатомовых водорослей и имеют обычное гетероконтовое строение, включая волоски (мастигонемы ) характеристика в других группах.
Диатомовые водоросли часто называют «драгоценностями моря» или «живыми опалами» из-за их оптических свойств.[22] Биологическая функция этого структурная окраска не ясно, но предполагается, что это может быть связано с коммуникацией, маскировкой, теплообменом и / или защитой от ультрафиолета.[23]
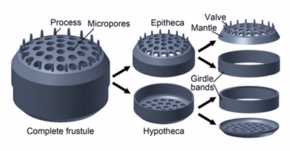
Диатомовые водоросли образуют сложные твердые, но пористые стенки клеток, называемые панцири состоит в основном из кремнезем.[24]:25–30 Эта кремнистая стена[25] может иметь четкий узор с множеством пор, ребер, мелких шипов, краевых выступов и возвышений; все это можно использовать для определения родов и видов.
Сама ячейка состоит из двух половин, каждая из которых содержит по существу плоскую пластину, или клапан, и краевое соединение, или поясную ленту. Одна половина ипотека, немного меньше другой половины, эпитека. Морфология диатомовых водорослей различна. Хотя форма ячейки обычно круглая, некоторые ячейки могут быть треугольными, квадратными или эллиптическими. Их отличительная особенность - твердая минеральная оболочка или панцирь, состоящий из опала (гидратированной полимеризованной кремниевой кислоты).

2) Ядрышко; Расположение хромосомы
3) аппарат Гольджи; изменяет белки и отправляет их из камеры
4) Клеточная стенка; Наружная мембрана клетки
5) Пиреноид; центр фиксация углерода
6) Хроматофор; структура мембраны, несущей пигмент
7) Вакуоли; везикула клетки, содержащая жидкость, связанную мембраной
8) Цитоплазматические тяжи; удерживать ядро
9) Митохондрии; Создайте АТФ (энергия) для клетки
10) Клапаны / Стрии; впускать питательные вещества в клетку и выводить ее из клетки
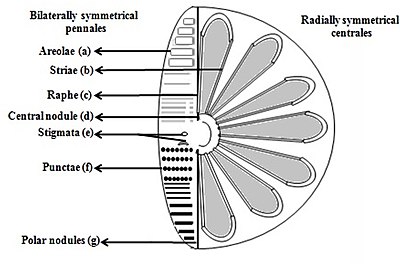
б) Стрии (поры, точки, пятна или точки в линию на поверхности
в) Raphe (прорезь в клапанах
г) Центральный узелок (утолщение стенки посередине шва
д) Стигматы (отверстия в поверхности клапана, которые выглядят округлыми снаружи, но с прорезью, похожей на внутреннюю
е) Точки (пятна или небольшие перфорации на поверхности
ж) Полярные узелки (утолщение стенки на дистальных концах шва) [26][27]

Диатомовые водоросли делятся на две группы, которые различаются формой панциря: центрические диатомеи и пеннатные диатомеи.
Пеннатные диатомеи двусторонне симметричны. Каждый из их клапанов имеет отверстия, которые являются прорезями вдоль швыряет и их раковины обычно вытянуты параллельно этим швам. Они вызывают движение клеток через цитоплазму, которая течет по швам, всегда двигаясь по твердым поверхностям.
Центрические диатомеи радиально симметричны. Они состоят из верхнего и нижнего клапанов - эпитека и ипотека - каждый из них состоит из клапана и пояса, которые могут легко скользить друг под другом и расширяться, увеличивая содержание клеток по мере развития диатомовых водорослей. Цитоплазма центрической диатомовой водоросли расположена вдоль внутренней поверхности раковины и образует полую выстилку вокруг большой вакуоли, расположенной в центре клетки. Эта большая центральная вакуоль заполнена жидкостью, известной как «клеточный сок», которая похожа на морскую воду, но изменяется в зависимости от содержания определенных ионов. Цитоплазматический слой является домом для нескольких органелл, таких как хлоропласты и митохондрии. Прежде чем центрическая диатомовая водоросль начинает расширяться, ее ядро оказывается в центре одного из створок и начинает двигаться к центру цитоплазматического слоя до завершения деления. Центрические диатомеи имеют различные формы и размеры, в зависимости от того, от какой оси простирается раковина и присутствуют ли шипы.

Поведение диатомовых водорослей
Наиболее центрические и пеннатные диатомеи арафида неподвижный, а их относительно плотные клеточные стенки заставляют их легко тонуть. Планктонный формы в открытой воде обычно полагаются на бурный перемешивание верхних слоев океанических вод ветром, чтобы они оставались взвешенными в освещенных солнцем поверхностных водах. Многие планктонные диатомовые водоросли также развили особенности, замедляющие скорость их опускания, такие как шипы или способность расти колониальными цепями.[29] Эти приспособления увеличивают их отношение площади поверхности к объему и тянуть, позволяя им дольше оставаться во взвешенном состоянии в толще воды. Отдельные клетки могут регулировать плавучесть через ионный насос.[30]
Некоторые пеннатные диатомовые водоросли способны к так называемому «скольжению», что позволяет им перемещаться по поверхности с помощью клея. слизь секретный через раф (удлиненная прорезь в торце клапана).[31][32] Чтобы клетка диатомовой водоросли могла скользить, она должна иметь твердый субстрат, на который будет прилипать слизь.
Клетки одиночны или объединены в колонии различного типа, которые могут быть связаны кремнистыми структурами; слизь подушечки, стебли или трубочки; аморфные массы слизи; или потоками хитин (полисахарид), которые секретируются в клеточных процессах.


Эта конфокальное изображение показывает диатомовые водоросли ' клеточная стенка (голубой), хлоропласты (красный), ДНК (синий), мембраны и органеллы (зеленый).
Жизненный цикл
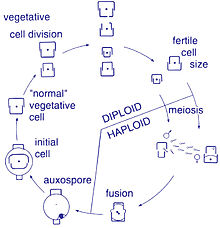
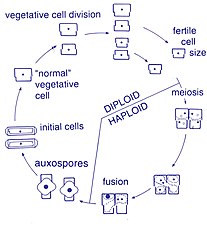
Размножение и размер клеток
Размножение среди этих организмов происходит бесполым путем. двойное деление, во время которого диатомовые водоросли делятся на две части, давая два «новых» диатомовых водоросли с идентичными генами. Каждый новый организм получает один из двух панцири - один больше, другой меньше - принадлежит родителю, который теперь называется эпитека; и используется для создания второй, меньшей панциря, ипотека. Диатомовая водоросль, получившая более крупную панцирь, становится того же размера, что и ее родитель, но диатомовая водоросль, получившая более мелкую панцирь, остается меньше своего родителя. Это приводит к уменьшению среднего размера клеток этой популяции диатомовых водорослей.[13] Однако было замечено, что некоторые таксоны обладают способностью делиться, не вызывая уменьшения размера клеток.[33] Тем не менее, чтобы восстановить размер клеток популяции диатомовых водорослей для тех, кто действительно переносит уменьшение размера, половое размножение и вспомогательная спора формирование должно произойти.[13]
Деление клеток
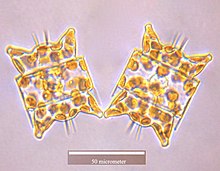
Вегетативные клетки диатомовых водорослей диплоид (2N) и так мейоз может происходить, производя мужские и женские гаметы, которые затем сливаются, образуя зигота. Зигота сбрасывает свою кремнеземную теку и превращается в большую сферу, покрытую органической мембраной, ауксоспорой. Новая клетка диатомовых водорослей максимального размера, исходная клетка, формируется внутри ауксоспоры, таким образом, начинается новое поколение. Покоящиеся споры также могут образовываться в ответ на неблагоприятные условия окружающей среды, при этом прорастание происходит при улучшении условий.[24]
Подвижность сперматозоидов
Диатомовые водоросли в основном неподвижный; однако сперма, обнаруженная у некоторых видов, может быть бичеванный, хотя подвижность обычно ограничивается скольжением.[24] У центрических диатомовых небольшой самец гаметы Имеется жгутик в то время как женские гаметы большие и неподвижные (оогамный ). Напротив, у пеннатных диатомовых водорослей обе гаметы лишены жгутиков (изогамный ).[13] Некоторые виды арафид, то есть перистые диатомеи без раф (шов), были задокументированы как анизогамные и, следовательно, считаются переходной стадией между центрическими и рафидными пеннатными диатомовыми водорослями, диатомовыми водорослями со швом.[33]
Разложение микробами
Определенные виды бактерий в океанах и озерах могут ускорить растворение кремнезема в мертвых и живых диатомовых водорослях, используя гидролитический ферменты, разрушающие органический материал водорослей.[34][35]
Экология


Потоки в Т моль Si y−1 (28 миллионов метрических тонн кремния в год)


по сравнению с концентрацией силиката [38]
Распределение
Диатомовые водоросли - широко распространенная группа, их можно найти в океаны, в пресная вода, в почвы, и на влажных поверхностях. Они являются одним из основных компонентов фитопланктон в богатых питательными веществами прибрежных водах и во время весеннего цветения в океане, поскольку они могут делиться быстрее, чем другие группы фитопланктона.[39] Самый живой пелагически в открытой воде, хотя некоторые из них живут как поверхностные пленки на границе раздела вода-осадок (бентосный ), или даже во влажных атмосферных условиях. Они особенно важны в океанах, где, по оценкам, на них приходится 45% всей океанической основное производство из органического материала.[40] Пространственное распространение видов морского фитопланктона ограничено как по горизонтали, так и по вертикали.[41][24]
Рост
Планктонный диатомовые водоросли в пресноводных и морских средах обычно проявляют "бум и спад " (или "цветение и бюст ») образ жизни. Когда условия в верхнем смешанном слое (питательные вещества и свет) благоприятны (как на весна ), их конкурентное преимущество и быстрый рост[39] позволяет им доминировать в сообществах фитопланктона («бум» или «цветение»). Как таковые, их часто называют оппортунистическими. р-стратеги (т.е. те организмы, экология которых определяется высокой скоростью роста, р).
Вклад в современный океанический цикл кремния
Диатомовые водоросли вносят значительный вклад в современный океанический цикл кремния: они являются источником подавляющего большинства биологических продуктов.
Влияние
Пресноводная диатомовая водоросль Didymosphenia geminata, широко известный как Дидимо, вызывает серьезную деградацию окружающей среды в водотоках, где цветет, образуя большие количества коричневого желеобразного материала, называемого «коричневыми соплями» или «каменными соплями». Эта диатомовая водоросль произрастает в Европе и является инвазивные виды как в антиподы и в части Северная Америка.[42][43] Проблема чаще всего регистрируется из Австралия и Новая Зеландия.[44]
Когда условия становятся неблагоприятными, как правило, при истощении питательных веществ, клетки диатомовых водорослей обычно увеличивают скорость погружения и выходят из верхнего смешанного слоя («разрушение»). Это опускание вызвано либо потерей контроля над плавучестью, синтезом слизи, склеивающей клетки диатомовых водорослей, либо производством тяжелых отдыхающие споры. Погружение из верхнего перемешанного слоя удаляет диатомеи из условий, неблагоприятных для роста, включая популяции травоядных животных и более высокие температуры (которые в противном случае увеличили бы рост клеток. метаболизм ). Затем клетки, достигающие более глубокой воды или мелкого морского дна, могут отдыхать, пока условия снова не станут более благоприятными. В открытом океане многие тонущие клетки теряются на глубине, но популяции убежищ могут сохраняться вблизи термоклин.
В конечном итоге клетки диатомовых водорослей в этих покоящихся популяциях повторно попадают в верхний перемешанный слой, когда их захватывает вертикальное перемешивание. В большинстве случаев это смешивание также пополняет питательные вещества в верхнем перемешанном слое, создавая основу для следующего раунда цветения диатомовых водорослей. В открытом океане (вдали от участков непрерывного апвеллинг[45]), этот цикл цветения, спада и затем возврата к условиям до цветения обычно происходит в течение годового цикла, причем диатомеи преобладают только весной и в начале лета. Однако в некоторых местах может произойти осеннее цветение, вызванное нарушением летней стратификации и увлечением питательных веществ, когда уровень освещенности все еще достаточен для роста. Поскольку вертикальное перемешивание увеличивается, а уровень освещенности падает с приближением зимы, эти цветы меньше и короче, чем их весенние эквиваленты.
В открытом океане цветение диатомовых водорослей (весеннее) обычно заканчивается нехваткой кремния. В отличие от других минералов, потребность в кремнии уникальна для диатомовых водорослей, и он не регенерируется в экосистеме планктона так эффективно, как, например, азот или фосфор питательные вещества. Это можно увидеть на картах поверхностных концентраций питательных веществ - по мере того, как питательные вещества снижаются по градиенту, кремний обычно исчерпывается первым (за ним обычно идет азот, затем фосфор).
Считается, что из-за этого цикла цветения и спада диатомовые водоросли играют непропорционально важную роль в экспорте углерода из поверхностных вод океана.[45][46] (см. также биологический насос ). Примечательно, что они также играют ключевую роль в регулировании биогеохимический цикл из кремний в современном океане.[40][36]
Причина успеха
Диатомовые водоросли экологически успешны и встречаются практически в любой среде, содержащей воду - не только в океанах, морях, озерах и ручьях, но также в почве и водно-болотных угодьях.[нужна цитата ] Многие исследователи считают, что использование кремния диатомовыми водорослями является ключом к этому экологическому успеху. Ворон (1983)[47] отметил, что по сравнению с органическими клеточные стенки, для синтеза панцирей кремнезема требуется меньше энергии (примерно 8% от сопоставимой органической стенки), что потенциально дает значительную экономию общего энергетического бюджета клетки. В уже ставшем классическим исследовании Эгге и Акснес (1992)[38] нашел эту диатомовую водоросль господство из мезокосм сообщества были напрямую связаны с доступностью кремниевой кислоты - когда концентрации превышали 2 мкмоль м−3они обнаружили, что диатомовые водоросли обычно составляют более 70% сообщества фитопланктона. Другие исследователи[48] предположили, что биогенный кремнезем в клеточных стенках диатомовых водорослей действует как эффективный pH буферный агент, облегчая преобразование бикарбонат к растворенному CO2 (который легче усваивается). В более общем плане, несмотря на эти возможные преимущества, связанные с использованием кремния, диатомеи обычно имеют более высокую скорость роста, чем другие водоросли того же соответствующего размера.[39]
Источники для сбора
Диатомовые водоросли можно получить из нескольких источников.[49] Морские диатомеи можно собирать прямым отбором проб воды, а бентические формы - соскобом. ракушки, устрица и другие снаряды. Диатомовые водоросли часто присутствуют в виде коричневого скользкого налета на погруженных в воду камнях и палках, и можно увидеть, как они «текут» с течением реки. На поверхности ила пруда, канавы или лагуны почти всегда можно найти диатомовые водоросли. Живые диатомеи часто в большом количестве цепляются за нитчатые водоросли или образуют студенистые массы на различных погруженных в воду растениях. Кладофора часто покрывается Кокконеис, диатомовая водоросль эллиптической формы; Vaucheria часто покрывается мелкими формами. Поскольку диатомовые водоросли составляют важную часть пищи моллюски, оболочки, и Рыбы, то пищеварительные тракты от этих животных часто получаются формы, которые нелегко закрепить другими способами. Диатомовые водоросли можно заставить появиться, если наполнить кувшин водой и грязью, обернуть его черной бумагой и позволить прямым солнечным лучам падать на поверхность воды. В течение дня диатомеи поднимутся наверх в виде накипи, и их можно будет изолировать.[49]
Биохимия

| Часть серии по |
| Планктон |
|---|
 |
Энергетический ресурс
Диатомовые водоросли в основном фотосинтезируют; однако некоторые обязательны гетеротрофы и могут жить в отсутствие света при наличии подходящего источника органического углерода.[50][51]
Метаболизм кремнезема
Клетки диатомовых водорослей содержатся в уникальном кремнеземе. клеточная стенка известный как усадьба состоит из двух клапанов, называемых теки, которые обычно перекрывают друг друга.[52] В биогенный кремнезем составляющая клеточная стенка синтезированный внутриклеточно посредством полимеризация из кремниевая кислота мономеры. Затем этот материал выдавливается на внешнюю поверхность ячейки и добавляется к стене. У большинства видов, когда диатомовые водоросли делятся с образованием двух дочерних клеток, каждая клетка сохраняет одну из двух половинок и вырастает в ней меньшую половину. В результате после каждого цикла деления средний размер диатомовых клеток в популяции становится меньше. Как только такие клетки достигают определенного минимального размера, вместо того, чтобы просто делиться, они обращают вспять это уменьшение, формируя вспомогательная спора. Это увеличивается в размере, давая начало гораздо большей клетке, которая затем возвращается к уменьшающимся в размерах делениям.[нужна цитата ] Производство ауксоспор почти всегда связано с мейоз и половое размножение.
Точный механизм передачи кремнезем поглощается диатомовыми водорослями клеточная стенка неизвестно. Большая часть секвенирования генов диатомовых водорослей происходит в результате поиска механизма поглощения и отложения кремнезема в наноразмерных структурах в усадьба. Наибольшего успеха в этой области достигли два вида: Thalassiosira pseudonana, который стал модельным видом, поскольку был секвенирован весь геном и были установлены методы генетического контроля, и Cylindrotheca fusiformis, в котором впервые были обнаружены важные белки осаждения кремнезема - силафины.[53] Силафины, наборы поликатионов пептиды, были найдены в С. fusiformis стенки ячеек и могут образовывать сложные кремнеземные структуры. В этих структурах наблюдались поры размеров, характерных для рисунков диатомей. Когда T. pseudonana прошел анализ генома, было обнаружено, что он кодирует цикл мочевины, в том числе большее количество полиамины чем большинство геномов, а также три различных гена транспорта кремнезема.[54] В филогенетический При исследовании генов транспорта кремнезема у 8 различных групп диатомовых водорослей было обнаружено, что транспорт кремнезема обычно группируется с видами.[53] Это исследование также обнаружило структурные различия между переносчиками кремнезема пеннатных (двусторонняя симметрия) и центрических (радиальная симметрия) диатомовых водорослей. Последовательности, сравниваемые в этом исследовании, были использованы для создания разнообразного фона, чтобы идентифицировать остатки, которые дифференцируют функцию в процессе осаждения кремнезема. Кроме того, в том же исследовании было обнаружено, что некоторые области сохраняются внутри видов, вероятно, это основная структура транспорта кремнезема.
Эти транспортные белки кремнезема уникальны для диатомовых водорослей и не имеют гомологи встречается у других видов, таких как губки или рис.Дивергенция этих генов транспорта кремнезема также указывает на структуру белка, происходящего из двух повторяющихся единиц, состоящих из пяти мембраносвязанных сегментов, что указывает либо на дупликацию гена, либо на димеризация.[53] Предполагается, что отложение кремнезема из мембраносвязанных везикул у диатомовых водорослей является результатом активности силлафинов и длинноцепочечных полиаминов. Эта везикула отложения кремнезема (SDV) была охарактеризована как кислый компартмент, слитый с везикулами, полученными из Гольджи.[55] Было показано, что эти две белковые структуры создают листы узорчатого кремнезема. in vivo с порами неправильной формы по шкале диатомовых водорослей панцири. Одна из гипотез о том, как эти белки работают для создания сложной структуры, заключается в том, что остатки консервативны в SDV, что, к сожалению, трудно идентифицировать или наблюдать из-за ограниченного числа доступных разнообразных последовательностей. Хотя точный механизм очень однородного осаждения кремнезема пока неизвестен, Thalassiosira pseudonana гены, связанные с силуффинами, рассматриваются как мишени для генетического контроля наноразмерных отложений кремнезема.
Цикл мочевины
Особенностью диатомовых водорослей является цикл мочевины, что эволюционно связывает их с животными. Это было обнаружено в ходе исследования, проведенного Эндрю Алленом, Крисом Боулером и его коллегами. Их результаты, опубликованные в 2011 году, о том, что диатомовые водоросли имеют функционирующий цикл мочевины, были очень важны, поскольку до этого считалось, что цикл мочевины возник из многоклеточные животные который появился через несколько сотен миллионов лет после диатомовых водорослей. Их исследование показало, что, хотя диатомовые водоросли и животные используют цикл мочевины для разных целей, считается, что они эволюционно связаны таким образом, как животные и растения - нет.[56]
Пигменты
Главный пигменты диатомовых водорослей хлорофиллы а и в, бета-каротин, фукоксантин, диатоксантин и диадиноксантин.[13]
Товары для хранения
Продукты для хранения хризоламинарин и липиды.[24]
Таксономия



Стефанодискус hantzschii

Нервная истмия
Odontella aurita
Диатомовые водоросли принадлежат к большой группе, называемой гетероконты, которые включают как автотрофы такие как золотые водоросли и ламинария; и гетеротрофы такие как водяные формы. Классификация гетероконтов до сих пор не определена: их можно обозначить как деление, филюм, Королевство, или что-то среднее между ними. Следовательно, диатомеи ранжируются где угодно из класса, обычно называемого Diatomophyceae или Bacillariophyceae, в подразделение (= тип), обычно называемое Bacillariophyta, с соответствующими изменениями в рангах своих подгрупп.
Роды и виды
Приблизительно 20 000 сохранившийся диатомовый виды считаются существующими, из которых около 12000 названы на сегодняшний день, согласно Guiry, 2012[57] (другие источники дают более широкий диапазон оценок[13][58][59][60]). Описано около 1000-1300 родов диатомовых, как существующих, так и ископаемых,[61][62] из которых около 250-300 существуют только в виде окаменелостей.[63]
Классы и заказы
В течение многих лет диатомовые водоросли, рассматриваемые либо как класс (Bacillariophyceae), либо как тип (Bacillariophyta), были разделены всего на 2 отряда, соответствующих центрическим и пеннатным диатомовым (Centrales и Pennales ). Эта классификация была тщательно пересмотрена Раундом, Кроуфордом и Манном в 1990 году, которые рассматривали диатомовые водоросли в более высоком ранге (подразделение, соответствующее филюму в зоологической классификации) и преобразовали основные классификационные единицы в классы, сохранив центрические диатомеи как единый класс. Coscinodiscophyceae, но разделение бывших пеннатных диатомовых водорослей на 2 отдельных класса, Fragilariophyceae и Bacillariophyceae (последнее старое название сохранено, но с измененным определением), между ними насчитывается 45 отрядов, большинство из которых являются новыми.
Сегодня (написано в середине 2020 года) признано, что система Round et al. нуждается в пересмотре с появлением новых молекулярных исследований, однако лучшая система для его замены неясна, и современные системы широко используются, такие как AlgaeBase, то Всемирный регистр морских видов и его база данных DiatomBase, и система «все живое», представленная в Ruggiero et al., 2015, все сохраняют Round et al. лечение в качестве их основы, хотя диатомовые водоросли в целом рассматриваются как класс, а не подраздел / тип, а классы Round et al. сведены к подклассам для лучшего согласования с трактовкой филогенетически смежных групп и содержащихся в них таксонов. (Ссылки см. В отдельных разделах ниже).
Одно из предложений, сделанное Линдой Медлин и соавторами в 2004 г., состоит в том, чтобы некоторые из центрических отрядов диатомовых, которые, как считается, более тесно связаны с перистыми, должны быть выделены в новый класс, Mediophyceae, который сам более тесно связан с перистыми диатомовыми водорослями, чем с перистыми диатомовыми. остальные центрики. Эта гипотеза, позже обозначенная как Coscinodiscophyceae-Mediophyceae-Bacillariophyceae или Coscinodiscophyceae + (Mediophyceae + Bacillariophyceae) (CMB), была принята Д.Г. Mann, среди прочих, который использует его в качестве основы для классификации диатомовых водорослей, представленной в Adl. и др. в серии синтезов (2005, 2012, 2019), а также в главе о Bacillariophyta 2017 г. Справочник протистов под редакцией Archibald et al., с некоторыми модификациями, отражающими очевидную немонофилию Medlin et al. оригинал "Coscinodiscophyceae". Между тем, группа, возглавляемая EC Theriot, поддерживает другую гипотезу филогении, которая была названа гипотезой структурной градации (SGH), и не признает Mediophyceae как монофилетическую группу, в то время как другой анализ, проведенный Parks et al., 2018, находит, что радиально-центрические диатомеи (Coscinodiscophyceae Medlin et al.) не являются монофилетическими, но поддерживают монофилию Mediophyceae минус Аттея, который является аномальным родом. Обсуждение относительных достоинств этих конфликтующих схем продолжается различными вовлеченными сторонами.[64][65][66][67]
Adl et al., 2019 лечение
В 2019 году Адл и другие.[68] представил следующую классификацию диатомовых водорослей, отметив при этом: «Этот пересмотр отражает многочисленные достижения в филогении диатомовых водорослей за последнее десятилетие. Из-за плохого отбора образцов таксонов за пределами Mediophyceae и перистых диатомовых, а также известного и ожидаемого разнообразия всех диатомовых водорослей. многие клады находятся на высоком уровне классификации (а классификация на более высоком уровне довольно плоская) ». Эта классификация рассматривает диатомовые водоросли как тип (Diatomeae / Bacillariophyta), принимает класс Mediophyceae Медлина и его сотрудников, вводит новые подтипы и классы для ряда изолированных родов и перераспределяет ряд ранее установленных таксонов в качестве подклассов. но не перечисляет заказы или семьи. Предполагаемые ранги были добавлены для ясности (Адл. И др. Не используют ранги, но предполагаемые ранги в этой части классификации очевидны из выбора используемых окончаний в рамках используемой системы ботанической номенклатуры).
- Clade Диатомиста Derelle et al. 2016, поправить. Cavalier-Smith 2017 (диатомовые водоросли плюс часть других групп охрофитов)
- Тип Diatomeae Dumortier 1821 [= Bacillariophyta Haeckel 1878] (диатомовые водоросли)
- Подтип Leptocylindrophytina D.G. Mann в Adl et al. 2019 г.
- Класс Leptocylindrophyceae D.G. Mann в Adl et al. 2019 (Лептоцилиндры, Tenuicylindrus )
- Класс Corethrophyceae D.G. Mann в Adl et al. 2019 (Коретрон )
- Подтип Ellerbeckiophytina D.G. Mann в Adl et al. 2019 (Эллербекия )
- Подтип Probosciophytina D.G. Mann в Adl et al. 2019 (Хобоща )
- Подтип Melosirophytina D.G. Mann в Adl et al. 2019 (Aulacoseira, Мелосира, Гиалодискус, Стефанопиксис, Паралия, Endictya )
- Подтип Coscinodiscophytina Medlin & Kaczmarska 2004, исправил. (Актиноптих, Coscinodiscus, Актиноциклус, Астеромфал, Авлакодискус, Stellarima )
- Подтип Rhizosoleniophytina D.G. Mann в Adl et al. 2019 (Guinardia, Ризосоления, Псевдосоления )
- Подтип Arachnoidiscophytina D.G. Mann в Adl et al. 2019 (Арахноидискус )
- Подтип Bacillariophytina Medlin & Kaczmarska 2004, исправил.
- Класс Mediophyceae Джоус и Прошкина-Лавренко в Medlin & Kaczmarska 2004
- Подкласс Chaetocerotophycidae Round & R.M. Crawford в Round et al. 1990 г., поправить.
- Подкласс Lithodesmiophycidae Round & R.M. Crawford в Round et al. 1990 г., поправить.
- Подкласс Thalassiosirophycidae Round & R.M. Кроуфорд в Round et al. 1990 г.
- Подкласс Cymatosirophycidae Round & R.M. Crawford в Round et al. 1990 г.
- Подкласс Odontellophycidae D.G. Mann в Adl et al. 2019 г.
- Подкласс Chrysanthemodiscophycidae D.G. Mann в Adl et al. 2019 г.
- Класс Biddulphiophyceae D.G. Mann в Adl et al. 2019 г.
- Подкласс Biddulphiophycidae Round и R.M. Кроуфорд в Round et al. 1990 г., поправить.
- Biddulphiophyceae incertae sedis (Аттея )
- Класс Bacillariophyceae Геккель 1878 г., исправил.
- Bacillariophyceae incertae sedis (Striatellaceae)
- Подкласс Urneidophycidae Medlin 2016
- Подкласс Fragilariophycidae Round in Round, Crawford & Mann 1990, исправлено.
- Подкласс Bacillariophycidae D.G. Манн в Раунде, Кроуфорд и Манн, 1990, исправьте.
Увидеть таксономия диатомовых водорослей Больше подробностей.
Галерея
- Изображения с растрового электронного микроскопа

Диатомовый Surirella spiralis

Диатомеи Chaetoceros виды на мембранном фильтре, размер пор 0,4 мкм.

Диатомовый Паралия бороздчатая.

Диатомовый Ахантес тринодис




Эволюция и летопись окаменелостей
Происхождение
Хлоропласты гетероконта происходят от хлоропластов красные водоросли, а не напрямую из прокариоты как произошло в растения. Это говорит о том, что они имели более недавнее происхождение, чем многие другие водоросли. Однако окаменелости скудны, и только с развитием самих диатомовых водорослей гетероконты оказывают серьезное влияние на летопись окаменелостей.
Самые ранние окаменелости
Самые ранние известные ископаемые диатомовые водоросли датируются ранним Юрский (~185 Ма назад ),[69] Хотя молекулярные часы[69] и осадочный[70] данные свидетельствуют о более раннем происхождении. Было высказано предположение, что их происхождение может быть связано с массовое вымирание в конце перми (~ 250 млн лет назад), после чего многие морские ниши были открыты.[71] Разрыв между этим событием и временем первого появления ископаемых диатомовых водорослей может указывать на период, когда диатомеи не были кремнеземными, а их эволюция была загадочный.[72] С момента появления кремнезема диатомовые водоросли оказали значительное влияние на летопись окаменелостей, при этом крупные залежи окаменелостей были обнаружены еще в ранние годы. Меловой, и с некоторыми камнями, такими как диатомитовая земля, почти полностью состоящий из них.
Отношение к кремниевому циклу
Хотя диатомеи могли существовать с Триасовый, время их господства и «захвата» кремниевого цикла произошло совсем недавно. До Фанерозой (до 544 млн лет) считается, что микробный или неорганический процессы слабо регулируют кремниевый цикл океана.[73][74][75] Впоследствии в цикле преобладают (и более строго регулируются) радиолярии и кремнистые губки, первый как зоопланктон, последний как сидячий фильтраторы в первую очередь на континентальные шельфы.[76] Считается, что в течение последних 100 млн. Лет цикл кремния попал под еще более жесткий контроль, и что это происходит из-за экологического превосходства диатомовых водорослей.
Однако точные сроки «захвата» остаются неясными, и разные авторы имеют противоречивые интерпретации летописи окаменелостей. Некоторые свидетельства, такие как смещение кремнистых губок с полок,[77] предполагает, что это поглощение началось в меловом периоде (146–66 млн лет), в то время как данные радиолярий предполагают, что «захват» не начался до Кайнозойский (66 млн лет по настоящее время).[78]
Отношение к лугам
Расширение пастбище биомы и эволюционное излучение из травы в течение Миоцен считается, что это увеличило приток растворимого кремния в океаны, и утверждалось, что это способствовало развитию диатомовых водорослей в кайнозойскую эру.[79][80] Недавняя работа предполагает, что успех диатомовых водорослей не связан с эволюцией трав, хотя разнообразие диатомовых водорослей и пастбищ сильно увеличилось со среднего миоцена.[81]
Отношение к климату
Разнообразие диатомовых водорослей в кайнозое очень чувствительно к глобальной температуре, особенно к градиенту температуры на экваториальном полюсе. В более теплых океанах, особенно в более теплых полярных регионах, в прошлом было показано, что разнообразие диатомовых водорослей значительно ниже. Будущие теплые океаны с усилением полярного потепления, как это прогнозируется в сценариях глобального потепления,[82] таким образом, теоретически может привести к значительной утрате разнообразия диатомовых водорослей, хотя, исходя из имеющихся данных, невозможно сказать, произойдет ли это быстро или только в течение многих десятков тысяч лет.[81]
Метод исследования
Летопись окаменелостей диатомовых водорослей в основном была установлена благодаря извлечению их кремнистых водорослей. панцири в морских и неморских отложениях. Хотя диатомеи имеют стратиграфическую летопись как морских, так и неморских, диатомовые биостратиграфия, который основан на ограниченном по времени эволюционном происхождении и исчезновении уникальных таксонов, только хорошо разработан и широко применим в морских системах. Продолжительность ареалов диатомовых водорослей была документально подтверждена путем изучения кернов океана и горных пород, обнаженных на суше.[83] Где диатомовые биозоны хорошо зарекомендовали себя и откалиброваны шкала времени геомагнитной полярности (например., Южный океан, Северная часть Тихого океана, восточная экваториальная Тихий океан ), оценки возраста на основе диатомовых водорослей могут быть разрешены с точностью до <100000 лет, хотя типичное разрешение возраста для Кайнозойский диатомовые сообщества насчитывают несколько сотен тысяч лет.
Сохранившиеся в озерных отложениях диатомеи широко используются для палеоэкологические реконструкции из Четвертичный климат, особенно для озер с закрытыми бассейнами, которые подвержены колебаниям глубины и солености воды.
Диверсификация
В Меловой Сведения о диатомовых водорослях ограничены, но недавние исследования показывают прогрессирующую диверсификацию видов диатомовых водорослей. В Меловое – палеогеновое вымирание, который в океанах сильно повлиял на организмы с известковыми скелетами, по-видимому, оказал относительно небольшое влияние на эволюцию диатомовых водорослей.[84]
Оборот
Хотя массового вымирания морских диатомовых водорослей в период Кайнозойский, времена относительно быстрого эволюционного круговорота в сообществах морских диатомей приходились на Палеоцен –эоцен граница[85] и на эоцен –Олигоцен граница.[86] Дальнейший оборот сборок происходил в разное время между серединой Миоцен и поздно Плиоцен,[87] в ответ на прогрессирующее похолодание полярных регионов и развитие более эндемичных сообществ диатомовых водорослей.
Глобальная тенденция к более нежным панцирям диатомовых водорослей была отмечена с Олигоцен к Четвертичный.[83] Это совпадает со все более активной циркуляцией поверхности океана и глубинных вод, вызванной увеличением широтных температурных градиентов в начале крупных ледяной покров расширение на Антарктида и прогрессивное охлаждение через Неоген и Четвертичный к биполярному ледниковому миру. Это привело к тому, что диатомовые водоросли потребляли меньше кремния для образования своих панцирей. Повышенное перемешивание океанов обновляет кремнезем и другие питательные вещества, необходимые для роста диатомей в поверхностных водах, особенно в прибрежных и океанических регионах. апвеллинг.
Генетика
Маркировка выраженной последовательности
В 2002 году были впервые обнаружены свойства Phaeodactylum tricornutum репертуар генов был описан с использованием 1000 выраженные теги последовательности (EST).[88] Впоследствии количество EST было увеличено до 12 000, и была создана база данных EST диатомовых водорослей для функционального анализа.[89] Эти последовательности были использованы для проведения сравнительного анализа между П. tricornutum и предполагаемые полные протеомы из зеленая водоросль Chlamydomonas reinhardtii, то красная водоросль Cyanidioschyzon merolae, и диатомовые Thalassiosira pseudonana.[90] База данных диатомовых EST в настоящее время состоит из более чем 200 000 EST из П. tricornutum (16 библиотек) и T. pseudonana (7 библиотек) клетки, выращенные в различных условиях, многие из которых соответствуют различным абиотическим стрессам.[91]
Секвенирование генома
В 2004 г. весь геном центрической диатомеи, Thalassiosira pseudonana (32,4 МБ) был секвенирован,[92] затем в 2008 году секвенировали пеннатную диатомовую водоросль, Phaeodactylum tricornutum (27,4 Мб).[93] Сравнение двух показывает, что П. tricornutum геном включает меньше генов (10 402 против 11 776), чем T. pseudonana; между двумя геномами не было обнаружено никакой основной синтении (порядка генов). T. pseudonana гены показывают в среднем ~ 1,52 интрона на ген по сравнению с 0,79 интрона на ген. П. tricornutum, что указывает на недавнее широко распространенное увеличение интронов у центрических диатомовых водорослей.[93][94] Несмотря на относительно недавнюю эволюционную дивергенцию (90 миллионов лет), степень молекулярной дивергенции между центриками и перистыми крыльями указывает на быстрые темпы эволюции в пределах Bacillariophyceae по сравнению с другими группами эукариот.[93] Сравнительная геномика также установила, что определенный класс сменные элементы, ретротранспозоны, подобные Diatom Copia (или CoDis), значительно усилились в П. tricornutum геном в отношении T. pseudonana, что составляет 5,8 и 1% геномов соответственно.[95]
Перенос эндосимбиотического гена
Геномика диатомовых водорослей принесла много информации о масштабах и динамике эндосимбиотических заболеваний. передача гена (EGT) процесс. Сравнение T. pseudonana белки с гомологами у других организмов предполагают, что сотни имеют своих ближайших гомологов в линии Plantae. EGT по отношению к геномам диатомовых может быть проиллюстрировано тем фактом, что T. pseudonana геном кодирует шесть белков, которые наиболее тесно связаны с генами, кодируемыми Гиллардия тета (криптомонада ) нуклеоморф геном. Четыре из этих генов также обнаружены в геномах пластид красных водорослей, демонстрируя, таким образом, последовательный EGT от пластиды красных водорослей к ядру красных водорослей (нуклеоморфу) и к ядру гетероконта-хозяина.[92] Более свежий филогеномный анализ диатомовых протеомов предоставили доказательства празинофит -подобный эндосимбионт у общего предка хромальвеолаты что подтверждается тем фактом, что 70% генов диатомовых водорослей, происходящих из растений, имеют происхождение от зеленой линии, и что такие гены также обнаруживаются в геномах других страменопилы. Поэтому было высказано предположение, что хромальвеолаты являются продуктом серийного вторичного эндосимбиоз сначала с зеленые водоросли, за которым следует второй с красные водоросли который сохранил геномные следы предыдущего, но вытеснил зеленую пластиду.[96] Однако филогеномный анализ протеомов диатомовых водорослей и истории эволюции хромальвеолитов, вероятно, будет использовать преимущества дополнительных геномных данных из недостаточно секвенированных линий, таких как красные водоросли.
Горизонтальный перенос генов
Помимо EGT, горизонтальный перенос генов (HGT) может происходить независимо от эндосимбиотического события. Публикация П. tricornutum геном сообщил, что не менее 587 П. tricornutum гены, по-видимому, наиболее тесно связаны с бактериальными генами, составляя более 5% всех П. tricornutum протеом. Около половины из них также находятся в T. pseudonana геном, что свидетельствует об их древнем включении в род диатомовых водорослей.[93]
Генная инженерия
Чтобы понять биологические механизмы, лежащие в основе большого значения диатомовых водорослей в геохимических циклах, ученые использовали Phaeodactylum tricornutum и Thalassiosira виды виды как модельные организмы с 90-х гг.[97]В настоящее время доступно несколько инструментов молекулярной биологии для создания мутантов или трансгенных линий: плазмиды содержащие трансгены вставляются в клетки с помощью биолистический метод[98] или транскингдом бактериальная конъюгация[99] (с доходностью 10⁻⁶ и 10⁻⁴ соответственно[98][99]) и другие классические методы трансфекции, такие как электропорация или использование ПЭГ сообщалось, что они дают результаты с более низкой эффективностью.[99]
Трансфицированные плазмиды могут быть либо случайным образом интегрированы в хромосомы диатомовых водорослей, либо поддерживаться как стабильные кольцевые эписомы (благодаря центромерной последовательности дрожжей CEN6-ARSH4-HIS3[99]). Флеомицин /ген устойчивости к зеоцину Sh Ble обычно используется как маркер выбора,[97][100] и различные трансгены были успешно введены и экспрессированы в диатомовых водорослях со стабильной передачей из поколения в поколение,[99][100] или с возможностью его удаления.[100]
Кроме того, эти системы теперь позволяют использовать Инструмент редактирования генома CRISPR-Cas, что приводит к быстрому производству функциональных нокаут-мутанты[100][101] и более точное понимание клеточных процессов диатомовых водорослей.
Человеческое использование
Палеонтология
Разложение и разложение диатомовых водорослей приводит к органический и неорганический (в виде силикаты ) отложения, неорганический компонент которых может привести к методу анализа прошлой морской среды по кернам дна океана или заливные грязи, поскольку неорганическое вещество внедряется в отложения глины и илы и формирует постоянную геологическую запись таких морских пластов (см. кремнистый ил ).
Промышленное

Диатомовые водоросли и их раковины (панцири) как диатомит или диатомовая земля, являются важными производственными ресурсами, используемыми для тонкой полировки и фильтрации жидкостей. Сложная структура их микроскопических оболочек была предложена в качестве материала для нанотехнологий.[102]
Диатомит считается естественным нано материалом и имеет множество применений и приложений, таких как: производство различных керамических изделий, строительной керамики, термостойкой керамики, специальных оксидной керамики, для производства контроля влажности материалов, используемых в качестве фильтрующего материала, материала в цементе производственная промышленность, исходный материал для производства носителей лекарственных средств с пролонгированным высвобождением, абсорбционный материал в промышленных масштабах, производство пористой керамики, стекольная промышленность, используемая в качестве носителя катализатора, в качестве наполнителя в пластмассах и красках, очистка промышленных вод, держатель пестицидов, а также для улучшения физических и химических характеристик определенных почв и других целей. [103] [104] [105]
Диатомовые водоросли также используются для определения происхождения содержащих их материалов, в том числе морской воды.
Судебно-медицинская экспертиза
Основная цель анализа диатомовых водорослей в криминалистика заключается в том, чтобы отличить смерть от погружения в воду от посмертного погружения тела в воду. Лабораторные тесты могут выявить присутствие диатомовых водорослей в организме. Поскольку скелеты диатомовых водорослей на основе кремнезема не подвержены быстрому разложению, их иногда можно обнаружить даже в сильно разложившихся телах. Поскольку они не встречаются в организме естественным образом, если лабораторные тесты показывают в трупе диатомовые водоросли того же вида, что и в воде, где было извлечено тело, тогда это может быть хорошо. доказательства утопления как причина смерти. Смесь видов диатомовых водорослей, обнаруженных в трупе, может быть такой же или отличаться от окружающей воды, что указывает на то, утонула ли жертва в том же месте, где было найдено тело.[106]
Нанотехнологии
Осаждение кремнезема диатомовыми водорослями также может оказаться полезным для нанотехнологии.[107] Клетки диатомовых водорослей многократно и надежно производят клапаны различных форм и размеров, что потенциально позволяет диатомовым водорослям создавать микро- или наноразмерные структуры, которые могут быть использованы в ряде устройств, включая: оптические системы; полупроводник нанолитография; и даже автомобили для Доставка наркотиков. При соответствующем искусственный отбор процедуры, диатомовые водоросли, которые производят створки определенных форм и размеров, могут быть выведены для выращивания в хемостат культур для массовое производство наноразмерные компоненты.[108] Также было предложено использовать диатомовые водоросли в качестве компонента солнечных элементов, заменив их светочувствительный оксид титана для диоксида кремния, который диатомовые водоросли обычно используют для создания своих клеточных стенок.[109] Также были предложены солнечные панели для производства диатомового биотоплива.[110]
История открытия
Первая диатомовая водоросль, официально описанная в научной литературе, колониальная Bacillaria paradoxa, был открыт в 1783 году датским натуралистом Отто Фридрих Мюллер.
использованная литература
- ^ Дангерд, П. (1933). Traite d'Algologie. Поль Лешвалье и Филс, Париж, [1].
- ^ Дюмортье, Б.-К. (1822). Commentationes botanicae. Наблюдения за ботаникой, dédiées à la Société d'Horticulture de Tournay (PDF). Турне: Imprimerie de Ch. Casterman-Dieu, Rue de pont No. 10. pp. [I], [1] -116, [1, tabl., Err.] Архивировано из оригинал (PDF) 6 октября 2015 г. - через Algaebase.
- ^ Рабенхорст, Л. Флора europaea algarum aquae dulcis et submarinae (1864–1868). Sectio I. Algas diatomaceas completectens, cum figuris generum omnium xylographice impressionis (1864). С. 1–359. Lipsiae [Leipzig]: Apud Eduardum Kummerum.
- ^ Геккель, Э. (1878). Das Protistenreich.
- ^ Энглер А. и Гилг Э. (1919). Syllabus der Pflanzenfamilien: eine Übersicht über das gesamte Pflanzensystem mit besonderer Berücksichtigung der Medizinal- und Nutzpflanzen, nebst einer Übersicht über die Florenreiche und Florengebiete Pharmübermechizin und Florengebiete derbrachizin und Florengebiete derbrachizinderde zum, 8-е изд., Gebrüder Borntraeger Verlag, Берлин, 395 с.
- ^ диа-том-ос "разрезать пополам" (= дихо-том-ос ) – диа "через" или "отдельно" и корень tém-n-ō "Я режу". Чередование между е и о в корне глагола аблаут.
- ^ «Определение ДИАТОМ». www.merriam-webster.com. Архивировано из оригинал 29 июля 2018 г.. Получено 30 июля 2018.
- ^ Воздух, которым вы дышите? Из диатомовой водоросли
- ^ "Что такое диатомовые водоросли?". Диатомовые водоросли Северной Америки. Архивировано из оригинал 25 января 2020 г.. Получено 28 января 2020.
- ^ Treguer, P .; Нельсон, Д. М .; Van Bennekom, A.J .; Демастер, Д. Дж .; Leynaert, A .; Кегинер, Б. (1995). «Баланс кремнезема в Мировом океане: переоценка». Наука. 268 (5209): 375–9. Bibcode:1995Научный ... 268..375T. Дои:10.1126 / science.268.5209.375. PMID 17746543. S2CID 5672525.
- ^ "Королевский колледж Лондона - озеро Мегачад". www.kcl.ac.uk. Получено 5 мая 2018.
- ^ Bristow, C.S .; Hudson-Edwards, K.A .; Чаппелл, А. (2010). «Удобрение Амазонки и экваториальной Атлантики западноафриканской пылью». Geophys. Res. Латыш. 37 (14): L14807. Bibcode:2010GeoRL..3714807B. Дои:10.1029 / 2010GL043486.
- ^ а б c d е ж г Грет Р. Хасл; Эрик Э. Сивертсен; Карен А. Стейдингер; Карл Танген (25 января 1996 г.). «Морские диатомеи». В Кармело Р. Томас (ред.). Определение морских диатомовых водорослей и динофлагеллят. Академическая пресса. С. 5–385. ISBN 978-0-08-053441-1. Получено 13 ноября 2013.
- ^ "Пожиратели газа".
- ^ «Подробнее о диатомовых водорослях». Музей палеонтологии Калифорнийского университета. Архивировано из оригинал 4 октября 2012 г.. Получено 20 мая 2015.
- ^ Накаяма, Т .; Kamikawa, R .; Tanifuji, G .; Kashiyama, Y .; Ohkouchi, N .; Archibald, J.M .; Инагаки, Ю. (2014). «Полный геном нефотосинтетической цианобактерии диатомовых водорослей свидетельствует о недавней адаптации к внутриклеточному образу жизни». Труды Национальной академии наук Соединенных Штатов Америки. 111 (31): 11407–11412. Bibcode:2014ПНАС..11111407Н. Дои:10.1073 / pnas.1405222111. ЧВК 4128115. PMID 25049384.
- ^ Wehr, J.D .; Sheath, R. G .; Коциолек, Дж. П., ред. (2015). Пресноводные водоросли Северной Америки: экология и классификация (2-е изд.). Сан-Диего: Academic Press. ISBN 978-0-12-385876-4.
- ^ Жирар, Винсент; Сен-Мартен, Симона; Баффето, Эрик; Сен-Мартен, Жан-Поль; Нераудо, Дидье; Пейро, Даниэль; Роги, Гвидо; Рагацци, Эухенио; Сутихорн, Варавуд (2020). "Тайский янтарь: понимание ранней истории диатомовых водорослей?". BSGF - Бюллетень наук о Земле. 191: 23. Дои:10.1051 / bsgf / 2020028. ISSN 1777-5817.
- ^ Колин, С., Коэльо, Л.П., Сунагава, С., Боулер, К., Карсенти, Э., Борк, П., Пепперкок, Р. и Де Варгас, К. (2017) «Количественная 3D-визуализация для клеточной биологии. и экология экологических микробных эукариот ». eLife, 6: e26066. Дои:10.7554 / eLife.26066.002.
 Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0. - ^ Внутреннее пространство субарктического Тихого океана Земные экспедиции НАСА, 4 сентября 2018 г.
 Эта статья включает текст из этого источника, который находится в всеобщее достояние.
Эта статья включает текст из этого источника, который находится в всеобщее достояние. - ^ Rousseaux, Cecile S .; Грегг, Уотсон В. (2015). «Последние десятилетние тенденции в глобальном составе фитопланктона». Глобальные биогеохимические циклы. 29 (10): 1674–1688. Bibcode:2015GBioC..29.1674R. Дои:10.1002 / 2015GB005139.
- ^ Паркер, Эндрю Р .; Таунли, Хелен Э. (2007). «Биомиметика фотонных наноструктур». Природа Нанотехнологии. 2 (6): 347–53. Bibcode:2007НатНа ... 2..347П. Дои:10.1038 / nnano.2007.152. PMID 18654305.
- ^ Гордон, Ричард; Лошич, Душан; Тиффани, Мэри Энн; Надь, Стивен С .; Стерренбург, Фритьоф А.С. (2009). «Стеклянный зверинец: диатомеи для новых применений в нанотехнологиях». Тенденции в биотехнологии. 27 (2): 116–27. Дои:10.1016 / j.tibtech.2008.11.003. PMID 19167770.
- ^ а б c d е Рита А. Хорнер (2002). Таксономический справочник по распространенному морскому фитопланктону.. Биопресс. С. 25–30. ISBN 978-0-948737-65-7. Получено 13 ноября 2013.
- ^ «Стекло в природе». Музей стекла Корнинг. Архивировано из оригинал 7 марта 2013 г.. Получено 19 февраля 2013.
- ^ Тейлор, Дж. К., Хардинг, У. Р. и Арчибальд, К. (2007). Иллюстрированное руководство по некоторым распространенным видам диатомовых водорослей из Южной Африки. Гезина: Комиссия по водным исследованиям. ISBN 9781770054844.
- ^ Мишра, М., Аруха, А.П., Башир, Т., Ядав, Д., и Прасад, Г.Б.К.С. (2017) «Все новые грани диатомовых водорослей: потенциальный источник наноматериалов и не только». Границы микробиологии, 8: 1239. Дои:10.3389 / fmicb.2017.01239. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ а б Zhang, D .; Wang, Y .; Cai, J .; Pan, J .; Цзян, X .; Цзян, Ю. (2012). «Технология биопроизводства на основе микро- и наноструктуры диатомовых водорослей». Китайский научный бюллетень. 57 (30): 3836–3849. Bibcode:2012ЧСБУ..57.3836З. Дои:10.1007 / с11434-012-5410-х.
- ^ Падисак, Юдит; Сороки-Пинтер, Эва; Резнер, Жужанна (2003), Мартенс, Коэн (ред.), «Тонущие свойства некоторых форм фитопланктона и связь устойчивости формы с морфологическим разнообразием планктона - экспериментальное исследование» (PDF), Водное биоразнообразие: праздничный том в честь Анри Ж. Дюмона, Разработки в гидробиологии, Springer, Нидерланды, стр. 243–257, Дои:10.1007/978-94-007-1084-9_18, ISBN 9789400710849, заархивировано из оригинал (PDF) 23 июля 2018 г., получено 4 октября 2019
- ^ Андерсон, Ларс В. Дж .; Суини, Беатрис М. (1 мая 1977 г.). "Diel изменения в седиментационных характеристиках Ditylum brightwelli: изменения клеточного липида и эффекты респираторных ингибиторов и модификаторов ионного транспорта1". Лимнология и океанография. 22 (3): 539–552. Bibcode:1977LimOc..22..539A. Дои:10.4319 / lo.1977.22.3.0539. ISSN 1939-5590.
- ^ Поульсен, Николь С .; Спектор, Илан; Сперк, Тимоти П .; Шульц, Томас Ф .; Уэтерби, Ричард (1 сентября 1999 г.). «Скольжение диатомей является результатом актин-миозиновой системы подвижности». Подвижность клеток и цитоскелет. 44 (1): 23–33. Дои:10.1002 / (SICI) 1097-0169 (199909) 44: 1 <23 :: AID-CM2> 3.0.CO; 2-D. ISSN 1097-0169. PMID 10470016.
- ^ Манн, Дэвид Г. (февраль 2010 г.). "рафидные диатомеи". Веб-проект "Древо жизни". Получено 27 сентября 2019.
- ^ а б Г. Дребес (1 января 1977 г.). «Глава 9: Сексуальность». В Дитрих Вернер (ред.). Биология диатомовых водорослей. Ботанические монографии. 13. Калифорнийский университет Press. С. 250–283. ISBN 978-0-520-03400-6. Получено 14 ноября 2013.
- ^ Азам, Фарук; Бидл, Кей Д. (1999). «Ускоренное растворение диатомового кремнезема сообществами морских бактерий». Природа. 397 (6719): 508–12. Bibcode:1999Натура.397..508Б. Дои:10.1038/17351. S2CID 4397909. ИНИСТ:1755031.
- ^ Захарова Юлия Р .; Галачянц, Юрий П .; Курилкина Мария И .; Лихошвай, Александр В .; Петрова, Дарья П .; Шишлянников, Сергей М .; Равин, Николай В .; Марданов, Андрей В .; Белецкий, Алексей В .; Лихошвей, Елена В. (2013). «Структура микробного сообщества и деградация диатомовых водорослей в глубоком придонном слое озера Байкал». PLOS ONE. 8 (4): e59977. Bibcode:2013PLoSO ... 859977Z. Дои:10.1371 / journal.pone.0059977. ЧВК 3613400. PMID 23560063.
- ^ а б Treguer, P .; Нельсон, Д. М .; Van Bennekom, A.J .; Демастер, Д. Дж .; Leynaert, A .; Кегинер, Б. (1995). «Баланс кремнезема в Мировом океане: переоценка». Наука. 268 (5209): 375–9. Bibcode:1995Научный ... 268..375T. Дои:10.1126 / science.268.5209.375. PMID 17746543. S2CID 5672525.
- ^ Килиас, Эстель С .; Юнгес, Леандро; Шупраха, Лука; Леонард, Гай; Метфиес, Катя; Ричардс, Томас А. (2020). «Распространение хитридовых грибов и их совместное появление с диатомовыми водорослями коррелируют с таянием морского льда в Северном Ледовитом океане». Биология коммуникации. 3 (1): 183. Дои:10.1038 / с42003-020-0891-7. ЧВК 7174370. PMID 32317738. S2CID 216033140.
- ^ а б Egge, J. K .; Акснес Д. Л. (1992). «Силикат как регулирующее питательное вещество в конкуренции фитопланктона». Mar. Ecol. Прог. Сер. 83: 281–289. Bibcode:1992MEPS ... 83..281E. Дои:10.3354 / meps083281.
- ^ а б c Фурнас, Майлз Дж. (1990). «Темпы роста морского фитопланктона in situ: подходы к измерению, скорости роста сообществ и видов». Журнал исследований планктона. 12 (6): 1117–51. Дои:10.1093 / планкт / 12.6.1117. ИНИСТ:5474600.
- ^ а б Йул, Эндрю; Тиррелл, Тоби (2003). «Роль диатомовых водорослей в регулировании цикла кремния океана». Глобальные биогеохимические циклы. 17 (4): н / д. Bibcode:2003GBioC..17.1103Y. CiteSeerX 10.1.1.394.3912. Дои:10.1029 / 2002GB002018.
- ^ Липпс, Джери Х. (1970). «Эволюция планктона». Эволюция. 24 (1): 1–22. Дои:10.2307/2406711. JSTOR 2406711. PMID 28563010.
- ^ Дидимо, «Иностранцы среди нас». В архиве 7 октября 2015 г. Wayback Machine Виртуальная выставка Виртуальный музей Канады
- ^ "DEP сообщает, что Didymo обнаружен в реке Фармингтон в западном ответвлении. Проверено 15 января 2014 г.". Архивировано из оригинал 16 февраля 2015 г.. Получено 27 апреля 2015.
- ^ «Отчет для заинтересованных сторон Didymo - 31 октября 2008 г.». MAF Biosecurity New Zealand www.biosecurity.govt.nz. Архивировано из оригинал 12 февраля 2013 г.. Получено 1 декабря 2013.
- ^ а б Dugdale, R.C .; Вилкерсон, Ф. П. (1998). «Силикатное регулирование новой добычи в экваториальном тихоокеанском апвеллинге». Природа. 391 (6664): 270–273. Bibcode:1998Натура.391..270D. Дои:10.1038/34630. S2CID 4394149.
- ^ Сметачек, В. С. (1985). «Роль погружения в жизненных циклах диатомовых водорослей: экологическое, эволюционное и геологическое значение». Mar. Biol. 84 (3): 239–251. Дои:10.1007 / BF00392493. S2CID 85054779.
- ^ Рэйвен, Дж. А. (1983). «Транспорт и функция кремния в растениях». Биол. Rev. 58 (2): 179–207. Дои:10.1111 / j.1469-185X.1983.tb00385.x. S2CID 86067386.
- ^ Миллиган, А. Дж .; Морель, Ф. М. М. (2002). «Буферная роль протонов для кремнезема в диатомовых водорослях». Наука. 297 (5588): 1848–1850. Bibcode:2002Наука ... 297.1848M. Дои:10.1126 / science.1074958. PMID 12228711. S2CID 206507070.
- ^ а б Чемберлен, К. Дж. (1901) Методы гистологии растений, University of Chicago Press, США
- ^ Армстронг, Э; Роджерсон, А; Лефтли, Jw (2000). «Использование углерода морских водорослей тремя связанными с поверхностью гетеротрофными протистами: Stereomyxa ramosa, Nitzschia alba и Labyrinthula sp.». Экология водных микробов. 21: 49–57. Дои:10.3354 / ame021049. ISSN 0948-3055.
- ^ ЛЮИН, ДЖОЙС; ЛЮИН, Р. А. (1967). «Культура и питание некоторых апохлорных диатомовых водорослей рода Nitzschia». Микробиология. 46 (3): 361–367. Дои:10.1099/00221287-46-3-361. ISSN 1350-0872.
- ^ «Диатомовые водоросли». Получено 13 февраля 2016.
- ^ а б c Thamatrakoln, K .; Alverson, A.J .; Хильдебранд, М. (2006). "Сравнительный анализ последовательности диатомовых переносчиков кремния: к механистической модели транспорта кремния". Журнал психологии. 42 (4): 822–834. Дои:10.1111 / j.1529-8817.2006.00233.x. S2CID 86674657.
- ^ Крегер, Нильс; Дойцманн, Райнер; Манфред, Сампер (ноябрь 1999 г.). «Поликатионные пептиды из диатомовой двуокиси кремния, которые направляют формирование наносферы кремнезема». Наука. 286 (5442): 1129–1132. Дои:10.1126 / science.286.5442.1129. PMID 10550045. S2CID 10925689.[постоянная мертвая ссылка ]
- ^ Крогер, Нильс (2007). Справочник по биоминерализации: биологические аспекты и формирование структуры. Вайнхайм, Германия: Wiley-VCH Verlag GmbH. стр. глава 3.
- ^ Аллен, Эндрю Э .; Дюпон, Кристофер Л .; Оборник, Мирослав; Хорак, Алеш; Нунес-Неси, Адриано; МакКроу, Джон П .; Чжэн, Хун; Johnson, Daniel A .; Ху, Ханьхуа; Fernie, Alisdair R .; Боулер, Крис (2011). «Эволюция и метаболическое значение цикла мочевины в фотосинтетических диатомовых водорослях». Природа. 473 (7346): 203–7. Bibcode:2011Натура.473..203А. Дои:10.1038 / природа10074. PMID 21562560. S2CID 4350884. Сложить резюме – Science Daily (12 мая 2011 г.).
- ^ Гайри, доктор медицины (2012). «Сколько существует видов водорослей?». Журнал психологии. 48 (5): 1057–1063. Дои:10.1111 / j.1529-8817.2012.01222.x. PMID 27011267. S2CID 30911529.
- ^ Фрэнк Эрик Раунд; Р. М. Кроуфорд; Д. Г. Манн (1990). Диатомовые водоросли: биология и морфология родов. Издательство Кембриджского университета. ISBN 978-0-521-36318-1. Получено 13 ноября 2013.[страница нужна ]
- ^ Кантер-Лунд, Х. и Лунд, Дж. У. Г. (1995). Пресноводные водоросли: объяснение их микроскопического мира, Биопресс Лимитед. ISBN 0-948737-25-5.[страница нужна ]
- ^ Манн, Дэвид Г. (2005). «Видовая концепция диатомовых водорослей: данные о морфологически различных симпатрических гамодемах у четырех эпипелических видов». Систематика и эволюция растений. 164 (1/4): 215–37. Дои:10.1007 / BF00940439. JSTOR 23675282. S2CID 37684109.
- ^ Фуртанье, Элизабет; Коциолек, Дж. Патрик (1999). "Каталог диатомовых родов". Диатомовые исследования. 14 (1): 1–190. Дои:10.1080 / 0269249X.1999.9705462.
- ^ В Всемирный регистр морских видов перечисляет 1356 названий родов диатомовых из всех местообитаний по состоянию на июль 2020 года, из которых 1248 являются «принятыми».
- ^ Запросы во Всемирный регистр морских видов, июль 2020 года, возвращают 299 названий родов «только ископаемые», из которых 285 «признаны».
- ^ Териот, Эдвард С .; Кэнноне, Джейми Дж .; Гутелл, Робин Р .; Алверсон, Эндрю Дж. (2009). «Пределы ядерно-кодируемой рДНК SSU для разрешения филогении диатомовых водорослей». Европейский журнал психологии. 4 (3): 277–290. Дои:10.5091 / плесево.2010.418. ЧВК 2835975. PMID 20224747.
- ^ Териот, Эдвард С .; Эшворт, Мэтт; Рак, Элизабет; Наков, Теофил; Янсен, Роберт К. (2010). «Предварительная мультигенная филогения диатомовых водорослей (Bacillariophyta): проблемы для будущих исследований». Экология и эволюция растений. 143 (3). Дои:10.1080/09670260902749159. PMID 20224747.
- ^ Парки, Мэтью Б.; Wickett, Norman J .; Алверсон, Эндрю Дж. (2018). «Сигнал, неопределенность и конфликт в филогеномных данных для различных линий микробных эукариот (Diatoms, Bacillariophyta)». Молекулярная биология и эволюция. 35 (1): 80–93. Дои:10.1093 / molbev / msx268. ЧВК 5850769. PMID 29040712.
- ^ Medlin, L.K .; Desdevises, Y. (2020). «Обзор филогенетической реконструкции диатомовых водорослей с использованием молекулярных инструментов с анализом набора данных из семи генов с использованием нескольких внешних групп и морфологических данных для подхода к полному доказательству» (PDF). Phycologia. под давлением.
- ^ Adl, Sina M .; Басс, Дэвид; Lane, Christopher E .; Лукеш, Юлий; Schoch, Conrad L .; Смирнов, Алексей; Агата, Сабина; Берни, Седрик; Браун, Мэтью В. (26 сентября 2018 г.). «Поправки к классификации, номенклатуре и разнообразию эукариот». Журнал эукариотической микробиологии. 66 (1): 4–119. Дои:10.1111 / jeu.12691. ЧВК 6492006. PMID 30257078.
- ^ а б Kooistra, Wiebe H.C.F .; Медлин, Линда К. (1996). «Эволюция диатомовых водорослей (Bacillariophyta)». Молекулярная филогенетика и эволюция. 6 (3): 391–407. Дои:10.1006 / mpev.1996.0088. PMID 8975694.
- ^ Шибер, Юрген; Кринсли, Дэйв; Рикпути, Ли (2000). «Диагенетическое происхождение кварцевого ила в аргиллитах и влияние на круговорот кремнезема». Природа. 406 (6799): 981–5. Bibcode:2000Натура.406..981С. Дои:10.1038/35023143. PMID 10984049. S2CID 4417951.
- ^ Medlin, L.K .; Kooistra, W.H.C.F .; Gersonde, R .; Sims, P. A .; Уэллброк, У. (1997). «Связано ли происхождение диатомовых водорослей с массовым вымиранием в конце перми?». Nova Hedwigia. 65 (1–4): 1–11. Дои:10.1127 / nova.hedwigia / 65/1997/1. HDL:10013 / epic.12689.
- ^ Raven, J. A .; Уэйт, А. М. (2004). «Эволюция окварцевания диатомовых водорослей: неизбежное погружение и погружение как побег?». Новый Фитолог. 162 (1): 45–61. Дои:10.1111 / j.1469-8137.2004.01022.x. JSTOR 1514475.
- ^ Р. Сивер; Стивен Генри Шнайдер; Пенелопа Дж. Бостон (январь 1993 г.). «Кремнезем в Мировом океане: биогеологическое взаимодействие». Ученые на Гайе. MIT Press. стр.287–295. ISBN 978-0-262-69160-4. Получено 14 ноября 2013.
- ^ Киддер, Дэвид Л .; Эрвин, Дуглас Х. (2001). «Вековое распространение биогенного кремнезема в фанерозое: сравнение замещенных кремнеземом окаменелостей и слоистых кремнеземов на уровне серии». Журнал геологии. 109 (4): 509–22. Bibcode:2001JG .... 109..509K. Дои:10.1086/320794. S2CID 128401816.
- ^ Гренн, Тор; Слэк, Джон Ф. (2003). «Палеозойская и мезозойская морская вода, богатая кремнеземом: данные по месторождениям гематитового кремня (яшмы)». Геология. 31 (4): 319–22. Bibcode:2003Гео .... 31..319Г. Дои:10.1130 / 0091-7613 (2003) 031 <0319: PAMSRS> 2.0.CO; 2. ИНИСТ:14692468.
- ^ Racki, G; Корди, Фабрис (2000). «Радиоляриевая палеоэкология и радиоляриты: настоящее - ключ к прошлому?». Обзоры наук о Земле. 52 (1): 83–120. Bibcode:2000ESRv ... 52 ... 83R. Дои:10.1016 / S0012-8252 (00) 00024-6.
- ^ Мальдонадо, Мануэль; Кармона, М. Кармен; Уриз, Мария Дж .; Крузадо, Антонио (1999). «Уменьшение количества мезозойских губок, строящих рифы, объясняется ограничением содержания кремния». Природа. 401 (6755): 785–8. Bibcode:1999Натура.401..785М. Дои:10.1038/44560. S2CID 205034177. ИНИСТ:1990263.
- ^ Харпер, Ховард Э .; Нолл, Эндрю Х. (1975). «Кремнезем, диатомовые водоросли и эволюция кайнозойских радиолярий». Геология. 3 (4): 175–7. Bibcode:1975Гео ..... 3..175H. Дои:10.1130 / 0091-7613 (1975) 3 <175: SDACRE> 2.0.CO; 2.
- ^ Falkowski, P.G .; Кац, Мириам Э .; Knoll, Andrew H .; Куигг, Антониетта; Рэйвен, Джон А .; Шофилд, Оскар; Тейлор, Ф. Дж. Р. (2004). «Эволюция современного эукариотического фитопланктона». Наука. 305 (5682): 354–60. Bibcode:2004Наука ... 305..354F. CiteSeerX 10.1.1.598.7930. Дои:10.1126 / science.1095964. PMID 15256663. S2CID 451773.
- ^ Kidder, D. L .; Герловски-Кордеш, Э. Х. (2005). "Воздействие радиации пастбищ на неморский цикл кремнезема и миоценовый диатомит". ПАЛАИ. 20 (2): 198–206. Bibcode:2005Палай..20..198K. Дои:10.2110 / palo.2003.p03-108. JSTOR 27670327. S2CID 140584104.
- ^ а б Лазарь, Давид; Бэррон, Джон; Реноди, Йохан; Дайвер, Патрик; Тюрке, Андреас (2014). "Кайнозойское планктонное разнообразие морских диатомовых водорослей и корреляция с изменением климата". PLOS ONE. 9 (1): e84857. Bibcode:2014PLoSO ... 984857L. Дои:10.1371 / journal.pone.0084857. ЧВК 3898954. PMID 24465441.
- ^ Основная группа авторов МГЭИК, 2007 г. Изменение климата 2007 г .: Сводный отчет. 104.
- ^ а б Scherer, R.P .; Гладенков, А.Ю .; Бэррон, Дж. А. (2007). «Методы и приложения кайнозойской биостратиграфии морских диатомей». Документы Палеонтологического общества. 13: 61–83. Дои:10.1017 / S1089332600001467.
- ^ Harwood, D.M .; Николаев, В. А .; Уинтер, Д. М. (2007). «Меловые записи эволюции, радиации и расширения диатомовых водорослей». Документы Палеонтологического общества. 13: 33–59. Дои:10.1017 / S1089332600001455.
- ^ Стрельникова, Н. И. (1990). «Эволюция диатомовых водорослей в меловой и палеогеновый периоды». В Симоле, Х. (ред.). Материалы Десятого Международного симпозиума по диатомовым водам. Кенигштейн: Научные книги Кельца. С. 195–204. ISBN 3-87429-307-6.
- ^ Балдауф, Дж. Г. (1993). «Средний эоцен - раннемиоценовый круговорот диатомовых водорослей». In Prothero, D .; Берггрен, У. Х. (ред.). Эоцен-олигоцен климатическая и биотическая эволюция. Принстон: Издательство Принстонского университета. С. 310–326. ISBN 0-691-02542-8.
- ^ Бэррон, Дж. А. (2003). «Появление и исчезновение планктонных диатомовых водорослей за последние 18 млн лет в Тихом и Южном океанах». Диатомовые исследования. 18: 203–224. Дои:10.1080 / 0269249x.2003.9705588. S2CID 84781882.
- ^ Scala, S .; Carels, N; Falciatore, A; Chiusano, M. L .; Боулер, C (2002). «Свойства генома диатомовой водоросли Phaeodactylum tricornutum». Физиология растений. 129 (3): 993–1002. Дои:10.1104 / стр.010713. ЧВК 166495. PMID 12114555.
- ^ Maheswari, U .; Montsant, A; Голл, Дж; Кришнасами, S; Раджяшри, К. Р .; Patell, V. M .; Боулер, C (2004). "База данных Diatom EST". Исследования нуклеиновых кислот. 33 (Выпуск базы данных): D344–7. Дои:10.1093 / нар / gki121. ЧВК 540075. PMID 15608213.
- ^ Montsant, A .; Джаббари, К. Maheswari, U; Боулер, C (2005). «Сравнительная геномика пеннатной диатомовой водоросли Phaeodactylum tricornutum». Физиология растений. 137 (2): 500–13. Дои:10.1104 / стр. 104.052829. ЧВК 1065351. PMID 15665249.
- ^ Maheswari, U .; Mock, T .; Armbrust, E. V .; Боулер, К. (2009). «Обновление базы данных Diatom EST: новый инструмент для цифровой транскриптомики». Исследования нуклеиновых кислот. 37 (Выпуск базы данных): D1001–5. Дои:10.1093 / nar / gkn905. ЧВК 2686495. PMID 19029140.
- ^ а б Armbrust, E. V .; Berges, John A .; Боулер, Крис; Грин, Беверли Р.; Мартинес, Диего; Putnam, Николас H .; Чжоу, Шигуо; Аллен, Эндрю Э .; Apt, Kirk E .; Бехнер, Майкл; Бжезинский, Марк А .; Chaal, Balbir K .; Чиовитти, Энтони; Дэвис, Обри К .; Демарест, Марк С .; Деттер, Дж. Крис; Главина, Тихана; Гудштейн, Дэвид; Хади, Масуд З .; Хеллстен, Уффе; Хильдебранд, Марк; Jenkins, Bethany D .; Юрка, Ежи; Капитонов Владимир В .; Крегер, Нильс; Lau, Winnie W. Y .; Лейн, Тодд У .; Лаример, Франк В .; Липпмайер, Дж. Кейси; и другие. (2004). «Геном диатомовой водоросли Thalassiosira Pseudonana: экология, эволюция и метаболизм». Наука. 306 (5693): 79–86. Bibcode:2004 Наука ... 306 ... 79А. CiteSeerX 10.1.1.690.4884. Дои:10.1126 / science.1101156. PMID 15459382. S2CID 8593895.
- ^ а б c d Боулер, Крис; Аллен, Эндрю Э .; Барсук, Джонатан Х .; Гримвуд, Джейн; Джаббари, Камель; Куо, Алан; Махешвари, Ума; Мартенс, Синди; Маумус, Флориан; Otillar, Robert P .; Райко, Эдда; Саламов, Асаф; Вандепоэле, Клаас; Beszteri, банк; Грубер, Ансгар; Хейде, Марк; Катинка, Михаил; Mock, Томас; Валентин, Клаус; Верре, Фредерик; Berges, John A .; Браунли, Колин; Кадоре, Жан-Поль; Чиовитти, Энтони; Чой, Чанг Джэ; Козель, Саша; Де Мартино, Алессандра; Деттер, Дж. Крис; Дуркин, Коллин; и другие. (2008). «Геном Phaeodactylum раскрывает эволюционную историю геномов диатомовых водорослей» (PDF). Природа. 456 (7219): 239–44. Bibcode:2008Натура.456..239Б. Дои:10.1038 / природа07410. PMID 18923393. S2CID 4415177. Архивировано из оригинал (PDF) 12 августа 2017 г.. Получено 20 сентября 2019.
- ^ Рой, С. У .; Пенни, Д. (2007). «Очень высокая доля уникальных позиций интронов в богатой интроном диатоме Thalassiosira pseudonana указывает на широко распространенное усиление интронов». Молекулярная биология и эволюция. 24 (7): 1447–57. Дои:10.1093 / молбев / msm048. PMID 17350938.
- ^ Маумус, Флориан; Аллен, Эндрю Э; Мхири, Коринн; Ху, Ханьхуа; Джаббари, Камель; Варди, Ассаф; Гранбастьен, Мари-Анжель; Боулер, Крис (2009). «Возможное влияние ретротранспозонов, активированных стрессом, на эволюцию генома морской диатомеи». BMC Genomics. 10: 624. Дои:10.1186/1471-2164-10-624. ЧВК 2806351. PMID 20028555.
- ^ Мустафа, А .; Beszteri, B .; Maier, U.G .; Bowler, C .; Валентин, К .; Бхаттачарья, Д. (2009). «Геномные следы криптического пластидного эндосимбиоза у диатомовых водорослей» (PDF). Наука. 324 (5935): 1724–6. Bibcode:2009Научный ... 324.1724M. Дои:10.1126 / science.1172983. PMID 19556510. S2CID 11408339. Архивировано из оригинал (PDF) 21 апреля 2014 г.. Получено 13 января 2019.
- ^ а б Kroth, Peter G .; Bones, Atle M .; и другие. (Октябрь 2018 г.). «Редактирование генома в данных: достижения и цели». Отчеты о растительных клетках. 37: 1401-1408. Дои:10.1007 / s00299-018-2334-1.
- ^ а б Фальсиатор, Анджела; Casotti, Raffaella; и другие. (Май 2015 г.). "Трансформация генов неселектаблпортеров в морской диатоме Re". Морская биотехнология. 1 (3): 239–251. Дои:10.1007 / PL00011773.
- ^ а б c d е Карас, Богумил Дж .; Diner, Рэйчел Э .; и другие. (21 апреля 2015 г.). «Дизайнерские эписомы диатомовых водорослей, полученные путем бактериальной конъюгации». Nature Communications. 6 (1): 6925. Дои:10.1038 / ncomms7925. ISSN 2041-1723.
- ^ а б c d Слэттери, Сэмюэл С .; Даймонд, Андрей; и другие. (16 февраля 2018 г.). «Расширенный набор генетических инструментов на основе плазмид позволяет редактировать геном Cas9 и стабильно поддерживать синтетические пути у Phaeodactylum tricornutum». Синтетическая биология ACS. 7 (2): 328–338. Дои:10.1021 / acssynbio.7b00191.
- ^ Нимарк, Марианна; Шарма, Амит Кумар; и другие. (Июль 2016 г.). «Система CRISPR / Cas9, адаптированная для редактирования генов морских водорослей». Научные отчеты. 6 (1): 24951. Дои:10.1038 / srep24951.
- ^ Мишра, М; Аруха, А.П .; Башир, Т; Ядав, Д; Гбкс, Прасад (2017). «Все новые грани диатомовых водорослей: потенциальный источник наноматериалов и не только». Передний микробиол. 8: 1239. Дои:10.3389 / fmicb.2017.01239. ЧВК 5496942. PMID 28725218.
- ^ Reka, Arianit A .; Павловский, Благой; Макрески, Петре (октябрь 2017 г.). «Новый оптимизированный метод низкотемпературного гидротермального производства пористой керамики с использованием диатомовой земли». Керамика Интернэшнл. 43 (15): 12572–12578. Дои:10.1016 / j.ceramint.2017.06.132.
- ^ Река, Арианит; Ановски, Тодор; Богоевский, Слободан; Павловский, Благой; Бошковски, Бошко (29 декабря 2014 г.). «Физико-химические и минералого-петрографические исследования диатомита из месторождения близ села Рожден, Республика Македония». Geologica Macedonica. 28 (2): 121–126. ISSN 1857-8586. Архивировано из оригинал 18 апреля 2020 г.. Получено 8 апреля 2020.
- ^ Reka, Arianit A .; Павловский, Благой; Адеми, Эгзон; Джашари, Ахмед; Боев, Блазо; Боев, Иван; Макрески, Петре (31 декабря 2019 г.). «Эффект термической обработки трепеля в диапазоне температур 800-1200˚С». Открытая химия. 17 (1): 1235–1243. Дои:10.1515 / chem-2019-0132.
- ^ Ауэр, Антти (1991). «Качественный анализ диатомовых водорослей как инструмент диагностики утопления». Американский журнал судебной медицины и патологии. 12 (3): 213–8. Дои:10.1097/00000433-199109000-00009. PMID 1750392. S2CID 38370984.
- ^ Брэдбери, Дж. (2004). "Нанотехнологи природы: раскрывая секреты диатомовых водорослей". PLOS Биология. 2 (10): 1512–1515. Дои:10.1371 / journal.pbio.0020306. ЧВК 521728. PMID 15486572.

- ^ Драм, Райан В .; Гордон, Ричард (2003). «Репликаторы из« Звездного пути »и нанотехнологии диатомовых водорослей». Тенденции в биотехнологии. 21 (8): 325–8. Дои:10.1016 / S0167-7799 (03) 00169-0. PMID 12902165.
- ^ Джонсон, Р. (9 апреля 2009 г.). «Диатомовые водоросли могут утроить эффективность солнечных батарей». EE Times. Архивировано из оригинал 31 июля 2012 г.. Получено 13 апреля 2009.
- ^ Рамачандра, Т. В .; Махапатра, Дурга Мадхаб; б, Картик; Гордон, Ричард (2009). «Доение диатомовых водорослей для устойчивой энергетики: биохимическая инженерия по сравнению с солнечными панелями из диатомовых водорослей, секретирующих бензин». Промышленные и инженерные химические исследования. 48 (19): 8769–88. Дои:10.1021 / ie900044j.
внешние ссылки
- База данных Diatom EST, École Normale Supérieure
- Планктон * Сеть, таксономический база данных, включающая изображения видов диатомовых водорослей
- История жизни и экология диатомовых водорослей, Музей палеонтологии Калифорнийского университета
- Диатомовые водоросли: "Природные шарики", Сайт Эврика, Бергенский университет
- История жизни и экология диатомовых водорослей, Восстановление и распространение изображений микрофоссилий для обучения и образования (ЧУДО), Университетский колледж Лондона
- Страница диатомовых водорослей, Королевский ботанический сад Эдинбурга
- Геометрия и закономерности в природе 3: дыры в тестах радиолярий и диатомей
- Краткие факты о диатоме, Научно-исследовательский институт аквариума Монтерей-Бэй
- База данных изображений водорослей Академия естественных наук Филадельфии (ПАНО)
- Таксоны диатомовых водорослей Академия естественных наук Филадельфии (ПАНО)
