Юрский - Jurassic - Wikipedia
| Юрский | |
|---|---|
| 201.3 ± 0.2 – ~145.0 Ма | |
| Хронология | |
Ключевые события юрского периода -205 — – -200 — – -195 — – -190 — – -185 — – -180 — – -175 — – -170 — – -165 — – -160 — – -155 — – -150 — – -145 — – как приблизиться к временной шкале ключевых событий юрского периода. Вертикальная ось: миллионы лет назад. | |
| Этимология | |
| Формальность имени | Формальный |
| Информация об использовании | |
| Небесное тело | земной шар |
| Региональное использование | Глобальный (ICS ) |
| Используемая шкала времени | Шкала времени ICS |
| Определение | |
| Хронологическая единица | Период |
| Стратиграфическая единица | Система |
| Формальность промежутка времени | Формальный |
| Определение нижней границы | Первое появление Аммонит Psiloceras spelae tirolicum. |
| Нижняя граница ГССП | Секция Кухйох, Карвендельские горы, Северные известняковые Альпы, Австрия 47 ° 29′02 ″ с.ш. 11 ° 31′50 ″ в.д. / 47.4839 ° с.ш. 11.5306 ° в. |
| ГССП ратифицирован | 2010 |
| Определение верхней границы | Формально не определен |
| Кандидаты в определение верхней границы | Никто |
| Верхняя граница раздела (-ов) кандидата GSSP |
|
| Атмосферные и климатические данные | |
| Среднее атмосферное О 2 содержание | c. 26 об.% (130% современных) |
| Среднее атмосферное CO 2 содержание | c. 1950 промилле (7 раз доиндустриальный) |
| Средняя температура поверхности | c. 16,5 ° С (На 3 ° C выше современного) |
В Юрский (/dʒʊˈрæs.sɪk/ юу-РАСС-ik;[1]) это геологический период и система который охватывает 56 миллионов лет с конца Триасовый Период 201,3 миллиона лет назад (Mya ) к началу Меловой Период 145 млн лет назад.[примечание 1] Юрский период составляет средний период Мезозойская эра. Юрский период назван в честь Горы Джура в Европейские Альпы, куда известняк пласты этого периода были впервые идентифицированы.
Начало периода ознаменовалось крупными Триас-юрское вымирание. Два других вымирание события произошли в период: Плинсбахско-тоарское вымирание в ранней юре и Титонский событие в конце;[4] ни одно событие не входит в число Массовые вымирания "большой пятерки", тем не мение.
Юрский период делится на три эпохи: Рано, Середина, и Поздно. Аналогичным образом в стратиграфия, юра делится на нижнюю, среднюю и верхнюю юрские. серии горных пород.
К началу юрского периода суперконтинент Пангея начался рифтинг на две суши: Лавразия на север, и Гондвана На юг. Это создало больше береговых линий и сместило континентальный климат с сухого на влажный, а многие из засушливый пустыни триаса сменились пышными тропическими лесами.
На суше фауна перешла от фауны триаса, в которой преобладали оба вида. динозавроморф и крокодиломорф архозавры, к тому, где доминирует динозавры один. Первый птицы также появился в юрском периоде, развившись из ветви теропод динозавры. Другие важные события включают появление самых ранних ящериц и эволюцию терианец млекопитающие. Крокодилы перешли от наземного образа жизни к водному. Океаны населяли морские рептилии Такие как ихтиозавры и плезиозавры, пока птерозавры были доминирующими летающими позвоночные.
Этимология
В хроностратиграфический термин «юрский период» напрямую связан с Горы Джура, а горный хребет в основном следуя курсу Граница между Францией и Швейцарией. Во время экскурсии по области в 1795 г.[заметка 2] Александр фон Гумбольдт признал в основном известняк Доминировал горный массив Джурских гор как отдельное образование, не включенное в сложившуюся стратиграфическую систему, определенную Авраам Готтлоб Вернер, и в 1799 году он назвал его «Юра-Калькштейн» («Юрский известняк»).[заметка 3][7][8][9]
Тридцать лет спустя, в 1829 году, французский натуралист Александр Бронгниар опубликовал обзор различных ландшафтов, составляющих земную кору. В этой книге Бронгниарт называл территории гор Юра как Terrains jurassiques, таким образом, вводится в обращение и публикуется термин впервые.[10]
Название «Джура» происходит от кельтского корня. * Джор через галльский * iuris "лесистая гора", заимствованная из латинского названия места, превратилась в Юрия и наконец Юра.[7][8][11]
Геология
Юрский период делится на три эпохи: Ранний, Средний и Поздний. Аналогичным образом в стратиграфия юрский период делится на Нижняя юра, Средняя юра, и Верхняя юра серии скальных образований, также известных в Европа в качестве Lias, Доггер и Мальм.[12] Разделение юрского периода на три части началось с Леопольд фон Бух.[9] Три эпохи подразделяются на более короткие отрезки времени, называемые возраст. Возраст юрского периода от самого молодого до самого старого:
| Верхняя / Поздняя юра | Титонский | (152.1 ± 4 – 145 ± 4 Mya ) |
| Кимериджский | (157,3 ± 4 - 152,1 ± 4 млн лет назад) | |
| Оксфордский | (163,5 ± 4 - 157,3 ± 4 млн лет назад) | |
| Средняя юра | Келловейский | (166,1 ± 4 - 163,5 ± 4 млн лет назад) |
| Батонский | (168,3 ± 3,5 - 166,1 ± 4 млн лет назад) | |
| Байосский | (170,3 ± 3 - 168,3 ± 3,5 млн лет назад) | |
| Ааленский | (174,1 ± 2 - 170,3 ± 3 млн лет назад) | |
| Нижняя / ранняя юра | Тоарский | (182,7 ± 1,5 - 174,1 ± 2 млн лет назад) |
| Плинсбахский | (190,8 ± 1,5 - 182,7 ± 1,5 млн лет назад) | |
| Синемурийский | (199,3 ± 1 - 190,8 ± 1,5 млн лет назад) | |
| Геттангский | (201,3 ± 0,6 - 199,3 ± 1 млн лет назад) |
Стратиграфия
Стратиграфия юрского периода в основном основана на использовании аммониты в качестве индекс окаменелости, с Датум первого появления конкретных таксонов аммонитов, используемых для обозначения начала стадий, а также меньших промежутков времени внутри стадий, называемых «Аммонитовые зоны», они, в свою очередь, также иногда подразделяются на подзоны. Глобальная стратиграфия основана на стандартных европейских аммонитовых зонах, а другие регионы откалиброваны по европейским сукцессиям.[13]

Самая старая часть юрского периода исторически упоминалась как Lias или лиасовый, примерно эквивалентный по протяженности раннеюрскому периоду, но также включающий часть предшествующих Ретиан. Геттангский этап был назван швейцарским палеонтологом. Эжен Реневье в 1864 г. после Hettange-Grande в Северо-Восточной Франции. В Разрез и точка стратотипа глобальной границы (GSSP) для основания геттанга находится на перевале Кухйох, Карвендельские горы, Северные известняковые Альпы, Австрия, ратифицированная в 2010 году. Начало геттанга, а значит и юрского периода в целом, отмечено Первое появление из аммонит Psiloceras spelae tirolicum.[14] Основание юрского периода ранее определялось как первое появление Psiloceras planorbis Альбертом Оппелем в 1856-58 гг., но это было изменено, поскольку внешний вид рассматривался как слишком локальное событие для международной границы.[13]
Синемурийский этап был определен и введен в научную литературу французским палеонтологом. Альсид д'Орбиньи в 1842 году. Название происходит от французского города Semur-en-Auxois, возле Дижон. Первоначальное определение синемурийского языка включало то, что сейчас является геттангским. GSSP Синемурийского периода расположен на скале к северу от деревни East Quantoxhead, 6 км к востоку от г. Watchet, Сомерсет, Англия в рамках Blue Lias. Начало синемурийского периода определяется первым появлением аммонита. Vermiceras quantoxense.[13][15]
Плейнсбахи был назван немецким палеонтологом. Альберт Оппель в 1858 г. после села Pliensbach в сообществе Целль-унтер-Айхельберг в Швабский Альб, возле Штутгарт, Германия. GSSP для основания Pleinsbachian находится в местности Wine Haven в Залив Робин Гуда, Йоркшир, Англия, в Формирование аргиллита Редкар. Начало плейнсбаха определено первым появлением аммонита. Bifericeras donovani.[16]
Тоарский назван в честь деревни Thouars (Латинский: Тоарций), к югу от Сомюр в Долина Луары из Франция, он был определен Альсидом д'Орбиньи в 1842 году первоначально из карьера Вринес примерно в 2 км к северо-западу от деревни. GSSP для базы Toarcian находится в Пенише, Португалия. Граница определяется по первому появлению аммонитов, относящихся к подроду Дактилиоцерасы (Eodactylioceras).[17]
Ааленский город назван в честь города Аален в Германии. Ааленский ярус определил швейцарский геолог. Карл Майер-Эймар в 1864 году. Изначально нижняя граница проходила между темными глинами Черный юрский и вышележащий глинистый песчаник и железистый оолит из Коричневый юрский период разрезы юго-западной Германии.[13] ГССП для базы Аален находится на Fuentelsaz в Иберийский диапазон возле Гвадалахара, Испания. Основание ааленского яруса определяется первым появлением аммонита. Leioceras opalinum.[18]
Байосиан назван в честь города Байе (Латинский: Bajoce) в Нормандии, Франция, и был определен Альсидом д'Орбиньи в 1842 году. GSSP для базы Байос расположен в Муртинхейре в Португалии и был определен в 1997 году. аммонит Hyperlioceras mundum.[19]
Bathonian назван в честь города Ванна, Англия, представленный бельгийским геологом d'Omalius d'Halloy в 1843 г. после неполного вскрытия оолитовых известняков в нескольких карьерах региона. GSSP для основания Бата - Равен-дю-Бес, область Нижняя Ауран, Альпы Верхнего Прованса, Франция, который был определен в 2009 году. Фундамент Батона определяется первым появлением аммонита. Гонолкиты сходятся, в основе Зигзагообразные зигзаги аммонитовая зона.[20]
Келловей происходит от латинизированного названия деревни Kellaways в Уилтшире, Англия, и был определен Альсидом д'Орбиньи в 1852 году, первоначально с основанием на стыке между Формирование лесного мрамора и Формация Корнбраш. Однако позже было установлено, что эта граница находится в верхней части Бата. База келловейцев еще не имеет сертифицированного GSSP, по состоянию на 2019 год.[13]
Oxfordian назван в честь города Оксфорд в Англии, и был назван Альсидом д'Орбиньи в 1844 году в связи с Оксфордская глина. В основе оксфорда отсутствует определенный GSSP. У. Дж. Аркелл в исследованиях 1939 и 1946 гг. нижняя граница оксфорда была обозначена как первое появление аммонита. Quenstedtoceras mariae (затем помещается в род Vertumniceras ). Последующие предложения предполагали первое появление Cardioceras redcliffense как нижняя граница.[13]
Кимериджан назван в честь села Киммеридж на побережье Дорсет, Англия. Он был назван Альсидом д'Орбиньи в 1842 году в связи с Киммеридж Клэй. Хотя это не подтверждено, секция Flodigarry на Стаффин-Бэй на Остров Скай, Шотландия был представлен как GSSP для основания кимериджа.[21]
Титониан был введен в научную литературу Альбертом Оппелем в 1865 году. Имя Титониан необычно для геологических сценических псевдонимов, потому что оно происходит от Греческая мифология а не название места. Тифон был сыном Лаомедон из Трой и влюбился в Эос, греческая богиня Рассвет. Его имя было выбрано Альбертом Оппелем для этого стратиграфический стадии, потому что Титониан находится рука об руку с рассветом мелового периода. База Титониан в настоящее время не имеет GSSP.[13] Верхняя граница юрского периода также в настоящее время не определена. Кальпионеллиды, загадочная группа пелагических протистов с кальцитовыми тесты кратковременно обильные в период от поздней юры до раннего мелового периода, были предложены как наиболее многообещающие кандидаты на установление границы J / K. [22]
Месторождения полезных ископаемых и углеводородов
Киммериджская глина и ее эквиваленты являются основными материнская порода для Нефть Северного моря.[23] Аравийский межшельфовый бассейн, отложившийся от конца средней до верхней юры, является местом расположения крупнейших в мире запасов нефти, включая Гавар Филд, крупнейшее в мире месторождение нефти.[24] Юрский возраст Саргелу[25] и наокелеканские свиты[26] являются основными нефтематеринскими породами Ирака. Более 1500 гигатонн запасов юрского угля находится в Северо-Западном Китае, в основном в Турпан-Хамиский бассейн и Бассейн Ордос.[27]
Кратеры от удара
Основные ударные кратеры включают Кратер Мороквенг кратер диаметром 70 км, погребенный под пустыней Калахари на севере Южной Африки. Воздействие датируется границей юры и мела, около 145 млн лет. Было высказано предположение, что кратер Мороквенг сыграл роль в круговороте юрско-мелового перехода.[28] Другой крупный ударный кратер - Кратер Пучеж-Катунки Диаметром 40-80 км, погребенный под Нижегородская область, Россия. Удар датирован синемурским периодом, примерно 192–196 млн лет назад.[29]
Палеогеография и тектоника


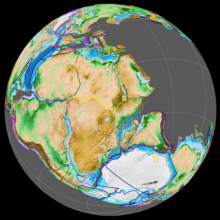
В раннеюрский период суперконтинент Пангея распался на северный суперконтинент Лавразия и южный суперконтинент Гондвана; то Мексиканский залив открылся в новом расколе между Северная Америка и что сейчас в Мексике Полуостров Юкатан. Юрский север Атлантический океан была относительно узкой, тогда как Южная Атлантика не открывалась до следующего мелового периода, когда Африка и Южная Америка разделились.[30] Континенты были окружены Панталасса, с Тетис Океан между Гондваной и Азией. Западная и Центральная Европа образовали архипелаг островов, окруженных мелководными морями. Мадагаскар и Антарктида начали отделяться от Африки во время ранней юры, начав фрагментацию Гондваны.[31][32] Климат был теплый, без признаков ледник появившись. Как и в триасовом периоде, по-видимому, не было суши ни над одним полюсом, и не было обширных ледяных шапок.
Основываясь на оценках кривых уровня моря, уровень моря был близок к нынешним уровням во время геттангского и синемюрского периодов, поднявшись на несколько десятков метров в течение позднего синемура-плейнсбаха, прежде чем к концу плейнсбаха снизился до почти современных уровней. Похоже, что в тоарский период происходил постепенный подъем до пика ~ 75 м над нынешним уровнем моря. Во время последней части тоаса уровень моря снова понижается на несколько десятков метров. Уровень моря постепенно повышался с ааленского яруса и далее, за исключением падений на несколько десятков метров в байосе и вокруг границы келловея и оксфорда, достигая высшей точки в уровне моря, возможно, на высоте 140 метров над нынешним уровнем моря в кимериджско-титонском периоде. граница. Уровень моря падает в позднем титоне, возможно, примерно до 100 метров, прежде чем подняться примерно до 110 метров на границе титона и берриаса.[33]
Геологическая летопись юрского периода хороша на западе. Европа, где обширные морские последовательности указывают на время, когда большая часть этой будущей суши была затоплена мелкими тропическими морями; известные места включают Юрское побережье Объект всемирного наследия в южной Англии и знаменитом конце юрского периода lagerstätten из Holzmaden и Solnhofen в Германии.[34] Напротив, летопись юры в Северной Америке является самой бедной из мезозойских отложений, с небольшими выходами на поверхность.[35] Хотя эпиконтинентальный Сандэнс Море оставили морские отложения в частях северных равнин Соединенных Штатов и Канады в конце юры, наиболее обнаженные отложения этого периода являются континентальными, такими как аллювиальный депозиты Формация Моррисон.
Юрский период был временем кальцитовое море геохимия, в которой низкомагнезийный кальцит был первичным неорганическим морским осадком карбоната кальция. Карбонатные грунты были поэтому очень распространены наряду с кальцитовыми ооиды, кальцитовые цементы и фауны беспозвоночных с преобладанием кальцитового скелета.[36]
Первый из нескольких массивных батолиты были размещены на севере Американские кордильеры начиная с середины юры, отмечая Неваданская орогения.[37]
В Африке слои ранней юры распределены аналогично слоям позднего триаса, с более частыми выходами на поверхность на юге и менее распространенными слоями окаменелостей, в которых преобладают следы на севере.[38] По мере развития юрского периода в Африке размножались более крупные и знаковые группы динозавров, такие как зауроподы и орнитоподы.[38] Отложения средней юры в Африке мало представлены и мало изучены.[38] Позднеюрские отложения также слабо представлены, если не считать впечатляющих Фауна Тендагуру в Танзании.[38] Позднеюрская жизнь Тендагуру очень похожа на который найдено в западной части Северной Америки Формация Моррисон.[38]

Юрские известняки и мергели ( Формация Матмор ) на юге Израиль

Верхняя юра Формация Моррисон в Колорадо является одним из самых плодородных источников окаменелостей динозавров в Северной Америке.

Гигандипус, а динозавр след в нижней юре Формация Моэнав в месте обнаружения динозавров Святого Георгия на ферме Джонсон, юго-запад Юта

В Пермский период стратиграфией юрского периода Плато Колорадо область юго-востока штата Юта




Флора


Засушливые континентальные условия, характерные для Триасовый неуклонно снижалась в юрский период, особенно в более высоких широтах; теплый влажный климат позволял пышным джунглям покрывать большую часть ландшафта.[39] Голосеменные были относительно разнообразны в юрский период.[12] Хвойные в особенности преобладала флора, как во время триаса; они были самой разнообразной группой и составляли большинство крупных деревьев.
Сохранившиеся семейства хвойных, которые процветали в юрский период, включали Араукариевые, Cephalotaxaceae, Pinaceae, Подокарповые, Taxaceae и Taxodiaceae.[40] Вымершее мезозойское семейство хвойных Хейролепидиевые преобладала низкоширотная растительность, а также кустарниковая Bennettitales.[41] Cycads, похожие на пальмы, также были обычным явлением, как и гинкго и древовидные папоротники В лесу.[12] Меньше папоротники были, вероятно, преобладающим подростом. Семенной папоротник малиновый были еще одной группой важных растений в то время и, как полагают, были от кустарников до небольших деревьев.[42] Гинкго были особенно распространены в средних и высоких северных широтах.[12] В Южном полушарии, подокарпы были особенно успешными, в то время как Гинкго и Чекановскиалес были редкостью.[39][41]
В океанах современные коралловые водоросли появился впервые.[12]
Фауна
Водные и морские
В юрский период основные позвоночные животные, обитавшие в море, были рыбы и морской рептилии. К последним относятся ихтиозавры, которые были на пике своего разнообразия, плезиозавры, включая плиозавры, и морской талатто крокодилиобразные семей Телеозавры, Machimosauridae и Metriorhynchidae.[43] Во время поздней юры фильтрующая подача толстокожий рыба начнет разнообразиться, включая самую большую костную рыбу, которая существовала, Leedsichthys, с расчетной максимальной длиной более 15 метров.[44] В среднеюрский период самые древние из известных макрель акула появился, представленный родом Палеокархарии.[45]
в беспозвоночный world появилось несколько новых групп, в том числе рудисты (а риф -образующее разнообразие двустворчатые моллюски ) и белемниты. Известковый сабеллиды (Гломерула ) появился в ранней юре.[46][47] Юрский период также отличался разнообразием корковых и скучный (склеробионтов), и наблюдался значительный рост биоэрозия карбонатных оболочек и твердых грунтов. Особенно распространен ихноген (след окаменелости ) Гастрохенолиты.[48] Аммониты были многочисленны в юрском периоде, все юрские аммониты происходят из Филлоцератина, единственная группа аммонитов, пережившая вымирание TJ.[49]
В юрский период около четырех или пяти из двенадцати клады планктонных организмов, которые существуют в летописи окаменелостей, либо испытали массивную эволюционную радиацию, либо появились впервые.[12]

Плезиозавры подобно Мураенозавр бродил по юрским океанам.

Брюхоногие моллюски и прикрепил mytilid двустворчатые моллюски на плоскости залегания юрских известняков на юге Израиль.


Наземный

На суше различные архозавр рептилии оставались доминирующими. Юрский период был золотым веком для крупных травоядных динозавров, известных как зауроподы —Камаразавр, Апатозавр, Диплодок, Брахиозавр и многие другие, бродившие по стране в конце того периода; их кормовыми угодьями были либо прерии из папоротники ладони саговники и Bennettitales, или более высокие хвойные породы, в зависимости от их приспособлений. Меньший Орнитисхий травоядные динозавры, как стегозавры и маленький орнитоподы были менее преобладающими, но играли важные роли. На них охотились в основном тероподы, Такие как Цератозавр, Мегалозавр, Торвозавр и Аллозавр, все они относятся к «ящерице бедренной» или заурисхий филиал динозавры.[50]
Во время поздней юры первые авиалайнеры, подобно Археоптерикс, развился с малого целурозавр динозавры. В воздухе, птерозавры были обычными; они правили небесами, выполняя многие экологические роли, птицы,[51] и, возможно, уже произвел на свет некоторых из крупнейших летающих животных всех времен.[52][53] В подлеске были разные виды ранних млекопитающие, а также тритилодонты, похожий на ящерицу сфенодонты, и рано лиссамфибии. Остаток от Лиссамфибия эволюционировали в этот период, представив первые саламандры и цецилии.[54]

Аллозавр был одним из крупнейших наземных хищников юрского периода.

Стегозавр - один из самых узнаваемых родов динозавров, живший в середине и конце юрского периода.

Aurornis xui, который жил в конце юрского периода, может быть самым примитивный Avialan динозавр, известный на сегодняшний день, и является одним из самых первых птиц, обнаруженных на сегодняшний день.




Рептилии
Черепахи
Стебель-группа черепахи (Testudinata ) будет разнообразить во время юрского периода. Юрские стволовые черепахи принадлежат к двум более развитым кладам: Мезохелидия и Перихелидия.[55] Считается, что родовое состояние мезохелидцев - водное, в отличие от земного у Testudinata.[56] Две современные группы черепах (Testudines ), Pleurodires и Крипто-каталоги, расходится к средней юре.[57][58] Ранние криптографические линии, такие как Xinjiangchelyidae известны со средней юры,[59] в то время как ранняя стволовая линия плевродира, Platychelyidae известен с поздней юры.[60]
Лепидозавры

Лепидозавры, который включает в себя чешуйчатые (ящерицы и змеи) и ринхоцефалы (который сегодня включает только Туатара ) будет разнообразить во время юрского периода. В юрском периоде ринхоцефалы занимали широкий спектр морфологий и образов жизни, например, специализированные водные виды. Pleurosauridae а так же травоядные Опистодонтия.[61] Скваматические породы также будут диверсифицироваться во время юрского периода, предположительно, происхождение живых ящериц - в раннеюрский период (~ 190 млн лет назад) и расхождение большинства основных групп чешуекрылых во время раннего-среднего юрского периода.[62] Многие чешуйки юрского периода имеют неясные отношения к живым группам.[63] Ранние представители змеиной линии Офидия появляются в средней юре.[64]
Амфибии

Подавляющее большинство темноспондилы вымерли в конце триаса, только брахиопоиды дожить до юрского периода и за его пределами. Члены семьи Brachyopidae известны из юрских отложений Азии,[65] в то время как чигутизаврид Siderops известен с ранней юры Австралии.[66] Современное лиссамфибии также начал бы диверсифицироваться во время юрского периода. Ранняя юра Просалирус Считается, что он представляет собой первого родственника лягушки с морфологией, способной прыгать, как живые лягушки.[67] Морфогически узнаваемые стволовые лягушки, такие как Нотобатрахус известны из средней юры.[68] В то время как самые ранние земноводные саламандры известны еще в триасе.[69] Корона группа саламандры впервые появляются в средней-поздней юре Евразии вместе с родственниками стволовой группы. Многие юрские саламандры стволовой группы, например Марморерпетон и Кокартус считается, что были неотенический.[70] К ранним представителям саламандр коронной группы относятся Chunerpeton, Pangerpeton и Linglongtriton из средней-поздней юры Янляо Биота Китая, которые относятся к Криптобранхоидея, который содержит живые азиатский и гигантские саламандры.[71] Пока Beiyanerpeton, также из той же биоты считается одним из первых членов Саламандройда, группа, в которую входят все остальные живые саламандры.[72] Саламандры рассеялись бы в Северной Америке к концу юрского периода, о чем свидетельствуют Иридотритон найден в позднеюрской формации Моррисон.[73] Самый старый бесспорный ствол -цецилий это раннеюрский период Eocaecilia из Аризоны.[74] Четвертая группа лиссамфибий, вымершие альбанерпетонтиды, впервые появится в средней юре, представленный Anoualerpeton priscus из бата в Британии, а также неопределенные останки равно возрастных Ануальная формация Марокко.[75]
Млекопитающие
Млекопитающие широко разнообразились в течение юрского периода, важные группы юрских млекопитающих включают Морганукодонта, Докодонта, Eutriconodonta, Dryolestida, Харамийида и Мультитуберкулята. В то время как большинство юрских млекопитающих известны исключительно по изолированным зубам и фрагментам челюстей, исключительно сохранившиеся останки свидетельствуют о разнообразии образа жизни. Докодонтан Castorocauda был адаптирован для водных организмов, как и утконос и выдры.[76] Некоторые члены Харамийиды[77] и племя eutriconodontan Volaticotherini[78] обладал патагиум сродни тем из белки-летяги, позволяя им скользить по воздуху. Фруитафоссор был, вероятно, специалистом по колониальным насекомым, похожим на живых муравьеды.[79] Не млекопитающие цинодонты семьи Tritylodontidae пережили вымирание TJ и продолжали существовать в раннем меловом периоде. Theriiform млекопитающие, представленные сегодня живыми плаценты и сумчатые, также появится в начале поздней юры, представленный Джурамая, а евтерский млекопитающее ближе к предкам плацентарных, чем сумчатые.[80] Джурамая намного более развитый, чем ожидалось для своего возраста, поскольку другие териформные млекопитающие не появляются до раннего мелового периода.[81]
Насекомые и паукообразные
Многочисленные важные местонахождения окаменелостей насекомых известны из юрского периода Евразии, наиболее важными из которых являются Карабастауская свита Казахстана, а также различные месторождения биоты Яньляо во Внутренней Монголии, Китай, такие как Кровать Daohugou, датируемые Келловейский -Оксфордский. Разнообразие насекомых оставалось неизменным на протяжении всей ранней и средней юры, но во второй трети юрского периода темпы их возникновения существенно возросли, в то время как темпы исчезновения оставались неизменными.[82] Средняя и поздняя юра была временем больших диверсификаций для жуки.[83] Долгоносики впервые появляются в летописи окаменелостей в период средней-верхней юры, но предполагается, что они возникли в период позднего триаса-ранней юры.[84] Самый старый из известных чешуекрылые (группа бабочек и мотыльков) известны с границы триаса и юры, с чешуей крыльев, принадлежащими подотряду Глоссата и Micropterigidae -состава бабочек из отложений этого возраста в Германии.[85] Хотя современные представители не известны до кайнозоя, эктопаразитарный насекомые, которые, как считается, представляют собой стволовая группа к блохи впервые появляются во время юрского периода, например Pseudopulex jurassicus. Эти насекомые существенно отличаются от современных блох, поскольку у них отсутствует специализированная морфология современных блох, и они больше по размеру.[86][87] Самая ранняя группа Фазматода (палочники), крылатые Susumanioidea, внешняя группа современных фазматод, впервые появилась в средней юре.[88] Самый старый член Mantophasmatidae (гладиаторы) также появились в это время.[89]
Известно лишь несколько записей о клещах из юрского периода, в том числе Юреремус, ан Орибатид клещ, принадлежащий семье Cymbaeremaeidae известен из верхней юры Великобритании и России.[90] В юрском периоде пауки расширились.[91] Ранняя юра Сеппо Копонени Считается, что возможно представляет основную группу для Пальпиманоидея.[92] Eoplectreurys из средней юры Китая считается стволовой линией Synspermiata. Самый старший член семьи Archaeidae, Патархей, известен из средней юры.[91] Монголарачне из средней юры Китая - один из крупнейших известных ископаемых пауков с ногами более 5 сантиметров.[93] Единственный скорпион, известный из юрского периода, - это Лиассоскорпиониды из нижней юры Германии, местонахождение не определено.[94] Eupnoi Opiliones известны из средней юры, включая членов семейства Склеросоматиды.[95][96]
В популярной культуре
С начала 1990-х годов термин Юрский был популяризирован парк Юрского периода франшизы, которая началась в 1990 году с Майкл Крайтон с роман с таким же названием и это экранизация, впервые выпущенный в 1993 году.
Смотрите также
Рекомендации
Примечания
- ^ Возраст юры-мела 140 млн лет вместо обычно принимаемых 145 млн лет был предложен в 2014 г. на основе стратиграфического исследования Формация Вака Муэрта в Бассейн Неукен, Аргентина.[2] Виктор Рамос, один из авторов исследования, предлагающего пограничный возраст 140 млн лет, рассматривает исследование как «первый шаг» к формальному изменению возраста в Международный союз геологических наук.[3]
- ^ "Ich hatte mich auf einer geognostischen Reise, die ich 1795 durch das südliche Franken, die westliche Schweiz und Ober-Italien machte, davon überzeugt, daß der Jura-Kalkstein, welchen Werner zu seinem Muschelkalk rechnete, eine eineiligntee Formation. В meiner Schrift über die unterirdischen Gasarten, welche mein Bruder Wilhelm von Humboldt 1799 während meines Aufenthalts в Südamerika herausgab, wird der Formation, die ich vorläufig mit dem Namen Jura-Kalkstein bezeichnete, zuerichnete.«(« Во время геологического тура, который я совершил в 1795 году через южную Францию, западную Швейцарию и верхнюю Италию, я убедился, что юрский известняк, который Вернер включил в свой ракушечный известняк, представляет собой отдельное образование. В моей статье о подземных типах Газы, которые мой брат Вильгельм фон Гумбольдт опубликовал в 1799 году во время моего пребывания в Южной Америке, впервые возникла формация, которую я условно обозначил названием «юрский известняк».[5]
- ^ "[…] Die ausgebreitete Formation, все, что нужно, - это все, что есть, унд neueren Sandstein liegt, und welchen их vorläufig mit dem Nahmen Jura-Kalkstein bezeichne.«… Широко распространенное образование, которое лежит между старым гипсом и более поздним песчаником и которое я условно обозначил названием« юрский известняк »».[6]
Цитаты
- ^ "Юрский". Dictionary.com Несокращенный. Случайный дом.
- ^ Vennari et al. 2014 г. С. 374-385.
- ^ Харамилло 2014.
- ^ Халлам 1986, стр. 765-768.
- ^ фон Гумбольдт 1858, п. 632.
- ^ фон Гумбольдт 1799, п. 39.
- ^ а б Гёльдер 1964.
- ^ а б Аркелл 1956.
- ^ а б Pieńkowski et al. 2008 г. С. 823–922.
- ^ Brongniart 1829.
- ^ Rollier 1903.
- ^ а б c d е ж Казлев 2002.
- ^ а б c d е ж грамм Ogg, J.G .; Hinnov, L.A .; Хуанг, К. (2012), "Юрский", Шкала геологического времени, Elsevier, стр. 731–791, Дои:10.1016 / b978-0-444-59425-9.00026-3, ISBN 978-0-444-59425-9, получено 2020-12-05
- ^ Hillebrandt, A.v .; Krystyn, L .; Kürschner, W.M .; Bonis, N.R .; Ruhl, M .; Richoz, S .; Schobben, M.A.N .; Урлихс, М .; Bown, P.R .; Кмент, К .; МакРобертс, К.А. (01.09.2013). «Глобальные стратотипические разрезы и точки (GSSP) для основания юрской системы в Кухйохе (горы Карвендель, Северные известняковые Альпы, Тироль, Австрия)». Эпизоды. 36 (3): 162–198. Дои:10.18814 / epiiugs / 2013 / v36i3 / 001. ISSN 0705-3797.
- ^ Блус, Герт; Пейдж, Кевин Н. (2002-03-01). «Глобальный стратотипический разрез и точка основания синемурийского яруса (нижняя юра)». Эпизоды. 25 (1): 22–28. Дои:10.18814 / epiiugs / 2002 / v25i1 / 003. ISSN 0705-3797.
- ^ Мейстер, Кристиан; Аберхан, Мартин; Блау, Иоахим; Доммерг, Жан-Луи; Файст-Буркхард, Сюзанна; Hailwood, Ernie A .; Харт, Малком; Hesselbo, Stephen P .; Хаунслоу, Марк У .; Хилтон, Марк; Мортон, Никол (01.06.2006). «Глобальный стратотипический разрез и точка (GSSP) для основания плинсбахского яруса (нижняя юра), Wine Haven, Йоркшир, Великобритания». Эпизоды. 29 (2): 93–106. Дои:10.18814 / epiiugs / 2006 / v29i2 / 003. ISSN 0705-3797.
- ^ Фантазия, Алисия; Адатте, Тьерри; Спангенберг, Хорхе Э .; Шрифт, Эрик; Дуарте, Луис В .; Фёльми, Карл Б. (ноябрь 2019 г.). «Глобальные процессы в сравнении с локальными во время перехода от плинсбаха к тоарскому веку в Пенише-ГССП, Португалия: запись с несколькими прокси». Обзоры наук о Земле. 198: 102932. Bibcode:2019ESRv..19802932F. Дои:10.1016 / j.earscirev.2019.102932.
- ^ Баррон, Эдуардо; Урета, Соледад; Гой, Антонио; Лассалетта, Луис (август 2010). «Палинология разреза и точки стратотипа глобальной границы тоара-аалена (GSSP) в Фуэнтельсасе (нижняя – средняя юра, Иберийский хребет, Испания)». Обзор палеоботаники и палинологии. 162 (1): 11–28. Дои:10.1016 / j.revpalbo.2010.04.003.
- ^ Павия, G .; Эней, Р. (1997-03-01). «Определение границы аален-байосского яруса». Эпизоды. 20 (1): 16–22. Дои:10.18814 / epiiugs / 1997 / v20i1 / 004. ISSN 0705-3797.
- ^ Лопес, Фернандес; Рафаэль, Сиксто; Павия, Джулио; Эрба, Элизабетта; Guiomar, Myette; Пайва Энрикес, Мария Хелена; Ланца, Роберто; Мангольд, Чарльз; Мортон, Никол; Оливеро, Давиде; Тирабоски, Даниэле (2009). «Глобальный стратотипический разрез и точка (GSSP) для подошвы батского яруса (средняя юра), разрез Равен-дю-Бес, Юго-Восточная Франция» (PDF). Эпизоды. 32 (4): 222–248. Дои:10.18814 / epiiugs / 2009 / v32i4 / 001. Архивировано из оригинал (PDF) 4 марта 2016 г.. Получено 5 июн 2015.
- ^ БАРСКИЙ, Марцин (2018-09-06). «Наборы цист динофлагеллат на границе оксфорда и кимериджа (верхняя юра) в Флодигарри, Стаффин-Бей, остров Скай, Шотландия - предлагаемый GSSP для основания кимериджа». Volumina Jurassica. XV (1): 51–62. Дои:10.5604/01.3001.0012.4594. ISSN 1731-3708.
- ^ УИМБЛДОН, Уильям А.П. (27 декабря 2017 г.). «События, связанные с установлением границы титона и берриаса (J / K)». Volumina Jurassica (1): 0. Дои:10.5604/01.3001.0010.7467. ISSN 1731-3708.
- ^ Готье Д.Л. (2005). «Общая нефтегазовая система кимериджских сланцев провинции Грабен Северного моря» (PDF). Геологическая служба США. Получено 2 ноября 2018.
- ^ Уилсон, А. О. (2020). «Глава 1: Введение в юрский арабский внутришельфовый бассейн». Геологическое общество, Лондон, Мемуары. 53 (1): 1–19. Дои:10.1144 / M53.1. ISSN 0435-4052. S2CID 226967035.
- ^ Абдула, Рзгер А. (август 2015 г.). «Углеводородный потенциал формации Саргелу и корреляция источников нефти, Иракский Курдистан». Арабский журнал наук о Земле. 8 (8): 5845–5868. Дои:10.1007 / s12517-014-1651-0. ISSN 1866-7511. S2CID 129120960.
- ^ Соранский университет; Абдула, Рзгер А. (16.10.2016). «Оценка материнской породы формации Наокелекан в Иракском Курдистане». Журнал Занкой Сулаймани - Часть A. 19 (1): 103–124. Дои:10.17656 / jzs.10589.
- ^ Ао, Вэйхуа; Хуанг, Вэньхуэй; Вен, Чэнминь; Сяо, Сюлин; Лю, Даменг; Тан, Сюи; Чен, Пинг; Чжао, Чжиген; Ван, Хуан; Финкельман, Роберт Б. (январь 2012 г.). «Петрология угля и генезис юрских углей в бассейне Ордос, Китай». Границы геонаук. 3 (1): 85–95. Дои:10.1016 / j.gsf.2011.09.004.
- ^ Теннант, Джонатан П .; Маннион, Филип Д .; Апчерч, Пол (2016-09-02). «Уровень моря регулирует динамику разнообразия четвероногих в юрском / меловом интервале». Nature Communications. 7 (1): 12737. Bibcode:2016НатКо ... 712737T. Дои:10.1038 / ncomms12737. ISSN 2041-1723. ЧВК 5025807. PMID 27587285.
- ^ Holm ‐ Alwmark, S .; Alwmark, C .; Ferrière, L .; Lindström, S .; Мейер, М. М. М .; Scherstén, A .; Herrmann, M .; Masaitis, V. L .; Мащак, М. С .; Наумов, М. В .; Журдан, Ф. (август 2019 г.). «Раннеюрский возраст импактной структуры Пучеж-Катунки (Россия) по данным 40 Ar / 39 Ar и палинологии». Метеоритика и планетология. 54 (8): 1764–1780. Bibcode:2019M & PS ... 54.1764H. Дои:10.1111 / maps.13309. ISSN 1086-9379.
- ^ Скотез 2003.
- ^ Гейгер, Маркус; Кларк, Дэвид Норман; Метте, Вольфганг (март 2004 г.). «Переоценка времени распада Гондваны на основе седиментологических и сейсмических данных из бассейна Морондава, Мадагаскар». Журнал африканских наук о Земле. 38 (4): 363–381. Bibcode:2004JAfES..38..363G. Дои:10.1016 / j.jafrearsci.2004.02.003.
- ^ Nguyen, Luan C .; Холл, Стюарт А .; Bird, Dale E .; Болл, Филип Дж. (Июнь 2016 г.). «Реконструкция континентальных окраин Восточной Африки и Антарктиды: РЕКОНСТРУКЦИЯ АФРИКИ-АНТАРКТИКИ». Журнал геофизических исследований: твердая Земля. 121 (6): 4156–4179. Дои:10.1002 / 2015JB012776.
- ^ Хак, Билал У. (01.01.2018). «Юрские колебания уровня моря: переоценка». GSA сегодня: 4–10. Дои:10.1130 / GSATG359A.1.
- ^ «Земля и море в юрский период». urweltmuseum.de. Архивировано из оригинал на 2007-07-14.
- ^ "Североамериканский гобелен времени и ландшафта: скалы юрского периода - 208–146 миллионов лет назад". nationalatlas.gov. Архивировано из оригинал на 2007-07-15.
- ^ Стэнли и Харди 1998.
- ^ Монро и Викандер 1997, п. 607.
- ^ а б c d е Джейкобс 1997, стр. 2-4.
- ^ а б Haines 2000, п. 65.
- ^ Behrensmeyer et al. 1992 г., п. 349.
- ^ а б Behrensmeyer et al. 1992 г., п. 352.
- ^ Behrensmeyer et al. 1992 г., п. 353.
- ^ Мотани 2000.
- ^ Листон Дж., Ньюбри М., Чалландс Т. и Адамс К., 2013 г., «Рост, возраст и размер юрских пахикормид. Leedsichthys problematicus (Osteichthyes: Actinopterygii) в: Arratia, G., Schultze, H. and Wilson, M. (eds.) Мезозойские рыбы 5 - глобальное разнообразие и эволюция. Verlag Dr. Friedrich Pfeil, Мюнхен, Германия, стр. 145–175.
- ^ Джамбура, Патрик Л .; Киндлиманн, Рене; Лопес-Ромеро, Фавьель; Маррама, Джузеппе; Пфафф, Катрин; Штумпф, Себастьян; Тюрчер, Юлия; Андервуд, Чарли Дж .; Уорд, Дэвид Дж .; Кривет, Юрген (декабрь 2019 г.). «Визуализация микрокомпьютерной томографии выявляет развитие уникального паттерна минерализации зубов у скумбрийских акул (Chondrichthyes; Lamniformes) в глубоком времени». Научные отчеты. 9 (1): 9652. Bibcode:2019НатСР ... 9.9652J. Дои:10.1038 / с41598-019-46081-3. ISSN 2045-2322. ЧВК 6609643. PMID 31273249.
- ^ Винн и Мутвей 2009 С. 286-296.
- ^ Винн, тен Хоув и Мутвей 2008 С. 295–301.
- ^ Тейлор и Уилсон, 2003 г., стр. 1-103.
- ^ Пейдж, Кевин Н. (январь 2008 г.). «Эволюция и география юрских аммоноидей». Труды ассоциации геологов. 119 (1): 35–57. Дои:10.1016 / S0016-7878 (08) 80257-X.
- ^ Haines 2000.
- ^ Feduccia 1996.
- ^ Виттон, Мартил и Ловеридж 2010 С. 79-81.
- ^ Виттон 2016.
- ^ Кэрролл 1988.
- ^ Джойс, Уолтер Г. (апрель 2017 г.). "Обзор летописи окаменелостей базальных мезозойских черепах". Бюллетень музея естественной истории Пибоди. 58 (1): 65–113. Дои:10.3374/014.058.0105. ISSN 0079-032X. S2CID 54982901.
- ^ Стерли, Джулиана; de la Fuente, Marcelo S .; Ружье, Гильермо В. (4 июля 2018 г.). «Новые останки Condorchelys antiqua (Testudinata) из ранней-средней юры Патагонии: анатомия, филогения и педоморфоз в ранней эволюции черепах». Журнал палеонтологии позвоночных. 38 (4): (1)–(17). Дои:10.1080/02724634.2018.1480112. ISSN 0272-4634. S2CID 109556104.
- ^ Wings et al. 2012 г., стр. 925-935.
- ^ Гэннон 2012.
- ^ Раби, Мартон; Джойс, Уолтер Дж .; Крылья, Оливер (сентябрь 2010 г.). «Обзор мезозойских черепах в бассейне Джунгар (Синьцзян, Северо-Западный Китай) и палеобиогеография от юрских до раннемеловых азиатских тестудинатов». Палеобиоразнообразие и палеоокружение. 90 (3): 259–273. Дои:10.1007 / s12549-010-0031-3. ISSN 1867-1594. S2CID 128805609.
- ^ Салливан, Патрик М .; Джойс, Уолтер Г. (август 2017 г.). «Анатомия панциря и таза позднеюрской черепахи Platychelys oberndorferi на основе материала из Золотурна, Швейцария». Швейцарский журнал палеонтологии. 136 (2): 323–343. Дои:10.1007 / s13358-017-0136-7. ISSN 1664-2376. S2CID 90587841.
- ^ Эррера-Флорес, Хорхе А .; Стаббс, Томас Л .; Бентон, Майкл Дж. (2017). «Макроэволюционные закономерности в Rhynchocephalia: является ли Tuatara (Sphenodon punctatus) живым ископаемым?». Палеонтология. 60 (3): 319–328. Дои:10.1111 / pala.12284. ISSN 1475-4983.
- ^ Бербринк, Фрэнк Т; Грацциотин, Фелипе Джи; Пайрон, Р. Александр; Кандалл, Дэвид; Доннеллан, Стив; Ирландская, Фрэнсис; Кио, Дж. Скотт; Краус, Фред; Мерфи, Роберт В; Нунан, Брайс; Раксуорти, Кристофер Дж (01.05.2020). Томсон, Роберт (ред.). «Изучение данных в геномном масштабе для чешуекрылых (ящериц, змей и амфисбенов) не обнаруживает поддержки ключевых традиционных морфологических связей». Систематическая биология. 69 (3): 502–520. Дои:10.1093 / sysbio / syz062. ISSN 1063-5157. PMID 31550008.
- ^ Донг, Липин; Ван, Юань; Mou, Lijie; Чжан, Гоцзе; Эванс, Сьюзан Э. (13 сентября 2019 г.). «Новая юрская ящерица из Китая». Геодиверситас. 41 (16): 623. Дои:10.5252 / geodiversitas2019v41a16. ISSN 1280-9659. S2CID 204256127.
- ^ Caldwell, M. W .; Nydam, R. L .; Palci, A .; Апестегия, С. Н. (2015). «Самые старые известные змеи из средней юры-нижнего мела дают представление об эволюции змей». Nature Communications. 6: 5996. Bibcode:2015 НатКо ... 6.5996C. Дои:10.1038 / ncomms6996. PMID 25625704.
- ^ Аверьянов Александр Олегович; Мартин, Томас; Скутчас, Павел П .; Резвой, Антон С .; Бакиров, Айзек А. (март 2008 г.). "Amphibians from the Middle Jurassic Balabansai Svita in the Fergana Depression, Kyrgyzstan (Central Asia)". Палеонтология. 51 (2): 471–485. Дои:10.1111/j.1475-4983.2007.00748.x.
- ^ Warren, A. A.; Hutchinson, M. N.; Hill, Dorothy (1983-09-13). "The last Labyrinthodont? A new brachyopoid (Amphibia, Temnospondyli) from the early Jurassic Evergreen formation of Queensland, Australia". Философские труды Лондонского королевского общества. B, биологические науки. 303 (1113): 1–62. Bibcode:1983RSPTB.303....1W. Дои:10.1098/rstb.1983.0080.
- ^ Reilly, Stephen M.; Jorgensen, Michael E. (February 2011). "The evolution of jumping in frogs: Morphological evidence for the basal anuran locomotor condition and the radiation of locomotor systems in crown group anurans". Журнал морфологии. 272 (2): 149–168. Дои:10.1002/jmor.10902. PMID 21210487. S2CID 14217777.
- ^ Báez, Ana Maria; Nicoli, Laura (March 2008). "A new species of Notobatrachus (Amphibia, Salientia) from the Middle Jurassic of northwestern Patagonia". Журнал палеонтологии. 82 (2): 372–376. Дои:10.1666/06-117.1. ISSN 0022-3360. S2CID 130032431.
- ^ Schoch, Rainer R.; Werneburg, Ralf; Voigt, Sebastian (2020-05-26). "A Triassic stem-salamander from Kyrgyzstan and the origin of salamanders". Труды Национальной академии наук. 117 (21): 11584–11588. Дои:10.1073/pnas.2001424117. ISSN 0027-8424. ЧВК 7261083. PMID 32393623.
- ^ Skutschas, Pavel; Stein, Koen (April 2015). "Long bone histology of the stem salamander Kokartus honorarius (Amphibia: Caudata) from the Middle Jurassic of Kyrgyzstan". Journal of Anatomy. 226 (4): 334–347. Дои:10.1111/joa.12281. ЧВК 4386933. PMID 25682890.
- ^ Jia, Jia; Gao, Ke-Qin (2019-03-04). "A new stem hynobiid salamander (Urodela, Cryptobranchoidea) from the Upper Jurassic (Oxfordian) of Liaoning Province, China". Журнал палеонтологии позвоночных. 39 (2): e1588285. Дои:10.1080/02724634.2019.1588285. ISSN 0272-4634. S2CID 164310171.
- ^ Gao, K.-Q .; Shubin, N. H. (2012-04-10). "Late Jurassic salamandroid from western Liaoning, China". Труды Национальной академии наук. 109 (15): 5767–5772. Дои:10.1073/pnas.1009828109. ISSN 0027-8424. ЧВК 3326464. PMID 22411790.
- ^ Evans, S.E .; Lally, C .; Chure, D. C.; Elder, A.; Maisano, J. A. (2005). "A Late Jurassic salamander (Amphibia: Caudata) from the Morrison Formation of North America". Зоологический журнал Линнеевского общества. 143 (4): 599–616. Дои:10.1111/j.1096-3642.2005.00159.x.
- ^ Santos, Rodolfo Otávio; Лорин, Мишель; Zaher, Hussam (2020-11-03). "A review of the fossil record of caecilians (Lissamphibia: Gymnophionomorpha) with comments on its use to calibrate molecular timetrees". Биологический журнал Линнеевского общества. 131 (4): 737–755. Дои:10.1093/biolinnean/blaa148. ISSN 0024-4066.
- ^ Haddoumi, Hamid; Аллен, Ронан; Meslouh, Said; Metais, Grégoire; Monbaron, Michel; Понс, Дениз; Rage, Jean-Claude; Вулло, Ромен; Zouhri, Samir (January 2016). "Guelb el Ahmar (Bathonian, Anoual Syncline, eastern Morocco): First continental flora and fauna including mammals from the Middle Jurassic of Africa" (PDF). Исследования Гондваны. 29 (1): 290–319. Bibcode:2016GondR..29..290H. Дои:10.1016/j.gr.2014.12.004. ISSN 1342-937X.
- ^ Ji, Q .; Luo, Z.-X.; Yuan, C.-X.; Tabrum, A. R. (2006). "A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals". Наука. 311 (5, 764): 1, 123–1, 127. Bibcode:2006Sci ... 311.1123J. Дои:10.1126/science.1123026. PMID 16497926. S2CID 46067702.
- ^ Meng, Qing-Jin; Grossnickle, David M.; Лю, Ди; Чжан, Ю-Гуан; Neander, April I.; Цзи, Цян; Luo, Zhe-Xi (August 2017). "New gliding mammaliaforms from the Jurassic". Природа. 548 (7667): 291–296. Bibcode:2017Natur.548..291M. Дои:10.1038/nature23476. ISSN 1476-4687. PMID 28792929. S2CID 205259206.
- ^ Meng, J .; Hu, Y.-M.; Wang, Y.-Q.; Wang, X.-L.; Li, C.-K. (2007). "Corrigendum: A Mesozoic gliding mammal from northeastern China". Nature 446 (7131): 102. Bibcode:2007 Натур.446Q.102M. Дои:10.1038 / природа05639.
- ^ Luo, Z.-X.; Wible, J.R. (2005). "A Late Jurassic Digging Mammal and Early Mammalian Diversification". Наука. 308 (5718): 103–107. Bibcode:2005Sci...308..103L. Дои:10.1126/science.1108875. ISSN 0036-8075. PMID 15802602. S2CID 7031381.
- ^ Zhe-Xi Luo; Chong-Xi Yuan; Qing-Jin Meng; Qiang Ji (25 August 2011). "A Jurassic eutherian mammal and divergence of marsupials and placentals" (PDF). Природа. 476 (7361): 442–445. Bibcode:2011Natur.476..442L. Дои:10.1038/nature10291. PMID 21866158. S2CID 205225806. Архивировано из оригинал (PDF) 10 ноября 2013 г. Electronic supplementary material
- ^ King, Benedict; Beck, Robin M. D. (2020-06-10). «Датирование подсказок поддерживает новые решения противоречивых отношений между ранними млекопитающими». Труды Королевского общества B: биологические науки. 287 (1928): 20200943. Дои:10.1098 / rspb.2020.0943. ЧВК 7341916. PMID 32517606.
- ^ Labandeira, Conrad C. (2018-05-23), "The Fossil History of Insect Diversity", Insect Biodiversity, Chichester, UK: John Wiley & Sons, Ltd, pp. 723–788, Дои:10.1002/9781118945582.ch24, ISBN 978-1-118-94558-2, получено 2020-12-06
- ^ McKenna, Duane D.; Шин, Сынгван; Ahrens, Dirk; Балке, Майкл; Beza-Beza, Cristian; Clarke, Dave J.; Донат, Александр; Escalona, Hermes E .; Friedrich, Frank; Летч, Харальд; Liu, Shanlin (2019-12-03). "The evolution and genomic basis of beetle diversity". Труды Национальной академии наук. 116 (49): 24729–24737. Дои:10.1073/pnas.1909655116. ISSN 0027-8424. ЧВК 6900523. PMID 31740605.
- ^ Шин, Сынгван; Clarke, Dave J; Lemmon, Alan R; Moriarty Lemmon, Emily; Aitken, Alexander L; Haddad, Stephanie; Farrell, Brian D; Marvaldi, Adriana E; Oberprieler, Rolf G; McKenna, Duane D (2018-04-01). "Phylogenomic Data Yield New and Robust Insights into the Phylogeny and Evolution of Weevils". Молекулярная биология и эволюция. 35 (4): 823–836. Дои:10.1093/molbev/msx324. ISSN 0737-4038. PMID 29294021. S2CID 4366092.
- ^ van Eldijk, Timo J. B.; Вапплер, Торстен; Strother, Paul K.; van der Weijst, Carolien M. H.; Раджаи, Хоссейн; Visscher, Henk; van de Schootbrugge, Bas (January 2018). "A Triassic-Jurassic window into the evolution of Lepidoptera". Достижения науки. 4 (1): e1701568. Bibcode:2018SciA....4.1568V. Дои:10.1126/sciadv.1701568. ISSN 2375-2548. ЧВК 5770165. PMID 29349295.
- ^ Huang, DiYing; Engel, Michael S.; Cai, ChenYang; Nel, André (May 2013). "Mesozoic giant fleas from northeastern China (Siphonaptera): Taxonomy and implications for palaeodiversity". Китайский научный бюллетень. 58 (14): 1682–1690. Bibcode:2013ChSBu..58.1682H. Дои:10.1007/s11434-013-5769-3. ISSN 1001-6538. S2CID 53578959.
- ^ Gao, Taiping; Shih, Chungkun; Rasnitsyn, Alexandr P.; Сюй, Син; Wang, Shuo; Ren, Dong (July 2013). "New Transitional Fleas from China Highlighting Diversity of Early Cretaceous Ectoparasitic Insects". Текущая биология. 23 (13): 1261–1266. Дои:10.1016/j.cub.2013.05.040. PMID 23810530. S2CID 9646168.
- ^ Yang, Hongru; Shi, Chaofan; Engel, Michael S; Zhao, Zhipeng; Рен, Донг; Gao, Taiping (2020-04-02). "Early specializations for mimicry and defense in a Jurassic stick insect". Национальный научный обзор: nwaa056. Дои:10.1093/nsr/nwaa056. ISSN 2095-5138.
- ^ Huang, Di-ying; Nel, André; Zompro, Oliver; Waller, Alain (2008-06-11). "Mantophasmatodea now in the Jurassic". Naturwissenschaften. 95 (10): 947–952. Bibcode:2008NW.....95..947H. Дои:10.1007/s00114-008-0412-x. ISSN 0028-1042. PMID 18545982. S2CID 35408984.
- ^ Селден, Пол А .; Baker, Anne S.; Phipps, Kenneth J. (2008). "An Oribatid Mite (arachnida: Acari) from the Oxford Clay (jurassic: Upper Callovian) of South Cave Station Quarry, Yorkshire, Uk". Палеонтология. 51 (3): 623–633. Дои:10.1111/j.1475-4983.2008.00769.x. HDL:1808/8353. ISSN 1475-4983.
- ^ а б Magalhaes, Ivan L. F.; Azevedo, Guilherme H. F.; Michalik, Peter; Ramírez, Martín J. (February 2020). "The fossil record of spiders revisited: implications for calibrating trees and evidence for a major faunal turnover since the Mesozoic". Биологические обзоры. 95 (1): 184–217. Дои:10.1111/brv.12559. ISSN 1464-7931. PMID 31713947. S2CID 207937170.
- ^ Селден, Пол А .; Dunlop, Jason A. (2014). "The first fossil spider (Araneae: Palpimanoidea) from the Lower Jurassic (Grimmen, Germany)". Zootaxa. 3894 (1): 161–168. Дои:10.11646/zootaxa.3894.1.13. PMID 25544628.
- ^ Selden, P. A.; Shih, C.K.; Ren, D. (2013). "A giant spider from the Jurassic of China reveals greater diversity of the orbicularian stem group". Naturwissenschaften. 100 (12): 1171–1181. Bibcode:2013NW....100.1171S. Дои:10.1007/s00114-013-1121-7. ЧВК 3889289. PMID 24317464.
- ^ Данлоп, Джейсон А .; Kamenz, Carsten; Scholtz, Gerhard (June 2007). "Reinterpreting the morphology of the Jurassic scorpion Liassoscorpionides". Строение и развитие членистоногих. 36 (2): 245–252. Дои:10.1016/j.asd.2006.09.003. PMID 18089103.
- ^ Хуанг, Дийин; Селден, Пол А .; Dunlop, Jason A. (August 2009). "Harvestmen (Arachnida: Opiliones) from the Middle Jurassic of China". Naturwissenschaften. 96 (8): 955–962. Bibcode:2009NW.....96..955H. Дои:10.1007/s00114-009-0556-3. ISSN 0028-1042. PMID 19495718. S2CID 9570512.
- ^ Гирибет, Гонсало; Tourinho, Ana Lúcia; Ши, ЧунгКун; Ren, Dong (March 2012). "An exquisitely preserved harvestman (Arthropoda, Arachnida, Opiliones) from the Middle Jurassic of China". Разнообразие и эволюция организмов. 12 (1): 51–56. Дои:10.1007/s13127-011-0067-x. ISSN 1439-6092. S2CID 15658216.
Источники
- Arkell, W.J. (1956). Jurassic Geology of the World. Эдинбург: Оливер и Бойд.
- Behrensmeyer, A.K.; Damuth, J.D.; DiMichele, W.A.; Potts, R.; Sues, H.D.; Wing, S.L., eds. (1992). Terrestrial Ecosystems through Time: the Evolutionary Paleoecology of Terrestrial Plants and Animals. Чикаго: Издательство Чикагского университета. ISBN 0-226-04154-9.
- Brongniart, Alexandre (1829). Tableau des terrains qui composent l'écorce du globe ou essai sur la structure de la partie connue de la terre [Description of the Terrains that Constitute the Crust of the Earth or Essay on the Structure of the Known Lands of the Earth] (На французском). Strasbourg – via Gallica.
- Carroll, R. L. (1988). Vertebrate Paleontology and Evolution. Нью-Йорк: WH Freeman. ISBN 978-0-7167-1822-2.
- Федучча, А. (1996). The Origin and Evolution of Birds. Нью-Хейвен: издательство Йельского университета. ISBN 978-0-300-06460-5.
- Gannon, Megan (31 October 2012). "Jurassic turtle graveyard found in China". cbsnews.com. Получено 10 октября 2019.
- Haines, Tim (2000). Walking with Dinosaurs: A Natural History. Нью-Йорк: Дорлинг Киндерсли. ISBN 978-0-7894-5187-3.
- Hallam, A. (1986). "The Pliensbachian and Tithonian extinction events". Природа. 319 (6056): 765–768. Bibcode:1986Natur.319..765H. Дои:10.1038/319765a0. S2CID 4310433.
- Hölder, H. (1964). Jura – Handbuch der stratigraphischen Geologie (на немецком). IV. Stuttgart: Enke-Verlag.
- Jacobs, Louis, L. (1997). "African Dinosaurs". In Currie, Phillip J.; Падиан, Кевин (ред.). Энциклопедия динозавров. Академическая пресса.
- Jaramillo, Jessica (March–April 2014). "Entrevista al Dr. Víctor Alberto Ramos, Premio México Ciencia y Tecnología 2013" (на испанском). Vol. 17 нет. 66.
Si logramos publicar esos nuevos resultados, sería el primer paso para cambiar officialmente la edad del Jurásico-Cretácico. A partir de ahí, la Unión Internacional de la Ciencias Geológicas y la Comisión Internacional de Estratigrafía Certificaría o no, depende de los resultados, ese cambio.
Журнал Cite требует| журнал =(помощь) - Kazlev, M. Alan (2002). "Palaeos Mesozoic: Jurassic: The Jurassic Period". Палеос. Архивировано из оригинал 5 января 2006 г.. Получено 8 января, 2006.
- Pieńkowski, G.; Schudack, M.E.; Bosák, P.; Enay, R.; Feldman-Olszewska, A.; Golonka, J.; Gutowski, J.; и другие. (2008). "Jurassic". В Макканн, Т. (ред.). The Geology of Central Europe. Mesozoic and Cenozoic. Лондон: Геологическое общество.
- Rollier, L. (1903). "Das Schweizerische Juragebirge". Sonderabdruck aus dem Geographischen Lexikon der Schweiz (на немецком). Attinger, Neuenburg: von Gebr.
- Monroe, James S.; Wicander, Reed (1997). The Changing Earth: Exploring Geology and Evolution (2-е изд.). Belmont: West Publishing Company. ISBN 0-314-09577-2.
- Motani, R. (2000). "Rulers of the Jurassic Seas". Scientific American. Vol. 283 no. 6. Дои:10.1038/scientificamerican1200-52.
- Scotese, Christopher R. (2003). "Pangea Begins to Rift Apart". scotese.com. Получено 2019-10-10.
- Stanley, S.M.; Hardie, L.A. (1998). "Secular oscillations in the carbonate mineralogy of reef-building and sediment-producing organisms driven by tectonically forced shifts in seawater chemistry". Палеогеография, палеоклиматология, палеоэкология. 144 (1–2): 3–19. Bibcode:1998PPP...144....3S. Дои:10.1016/s0031-0182(98)00109-6.
- Taylor, P.D .; Wilson, M.A. (2003). "Palaeoecology and evolution of marine hard substrate communities" (PDF). Обзоры наук о Земле. 62 (1): 1–103. Bibcode:2003ESRv ... 62 .... 1Т. Дои:10.1016/s0012-8252(02)00131-9. Архивировано из оригинал (PDF) на 25 марта 2009 г.
- Веннари, Вероника В .; Лескано, Марина; Найпауэр, Максимилиано; Агирре-Уррета, Беатрис; Кончайро, Андреа; Шальтеггер, Урс; Армстронг, Ричард; Пиментель, Марсио; Рамос, Виктор А. (2014). «Новые ограничения на границе юры и мела в Высоких Андах с использованием высокоточных U – Pb данных». Исследования Гондваны. 26 (1): 374–385. Bibcode:2014ГондР..26..374В. Дои:10.1016 / j.gr.2013.07.005.
- Винн, О .; ten Hove, H.A.; Mutvei, H. (2008). "On the tube ultrastructure and origin of calcification in sabellids (Annelida, Polychaeta)". Палеонтология. 51 (2): 295–301. Дои:10.1111/j.1475-4983.2008.00763.x.
- Винн, О .; Mutvei, H. (2009). "Calcareous tubeworms of the Phanerozoic" (PDF). Estonian Journal of Earth Sciences. 58 (4): 286–296. Дои:10.3176/earth.2009.4.07. Гейл A216178929.
- von Humboldt, Alexander (1799). Ueber die unterirdischen Gasarten und die Mittel, ihren Nachteil zu vermindern, ein Beitrag zur Physik der praktischen Bergbaukunde [On the types of subterranean gases and means of minimizing their harm, a contribution to the physics of practical mining] (на немецком). Брауншвейг: Vieweg.
- von Humboldt, Alexander (1858). Космос (на немецком). 4. Stuttgart: Cotta. п. 632 – via HathiTrust.
- Wings, Oliver; Rabi, Márton; Schneider, Jörg W.; Schwermann, Leonie; Солнце, Ге; Zhou, Chang-Fu; Joyce, Walter G. (2012). "An enormous Jurassic turtle bone bed from the Turpan Basin of Xinjiang, China". Naturwissenschaften. 114 (11): 925–935. Bibcode:2012NW.....99..925W. Дои:10.1007/s00114-012-0974-5. PMID 23086389. S2CID 17423081.
- Witton, Mark P.; Мартилла, Дэвид М .; Loveridge, Robert F. (2010). "Clipping the Wings of Giant Pterosaurs: Comments on Wingspan Estimations and Diversity". Acta Geoscientica Sinica. 31 (Supp 1): 79–81.
- Witton, Mark P. (23 June 2016). "Why the giant azhdarchid Arambourgiania philadelphiae needs a fanclub". markwitton-com.blogspot.com.
дальнейшее чтение
- Mader, Sylvia (2004). Биология (eighth ed.).
- Огг, Джим (июнь 2004 г.). Overview of Global Boundary Stratotype Sections and Points (GSSP's). International Commission on Stratigraphy. п. 17.
- Stanley, S.M.; Hardie, L.A. (1999). "Hypercalcification; paleontology links plate tectonics and geochemistry to sedimentology". GSA сегодня. 9: 1–7.
внешняя ссылка
- Examples of Jurassic Fossils
- Jurassic (chronostratigraphy scale)
- Jurassic fossils in Harbury, Warwickshire
- Jurassic Microfossils: 65+ images of Foraminifera
- . Британская энциклопедия. 15 (11-е изд.). 1911 г. With map and table.