Дилофозавр - Dilophosaurus
| Дилофозавр | |
|---|---|
 | |
| Реконструированный слепок образец голотипа (UCMP 37302) в месте захоронения, Королевский музей Онтарио | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Семья: | †Дилофозавриды |
| Род: | †Дилофозавр Уэллс, 1970 |
| Разновидность: | †D. Wherilli |
| Биномиальное имя | |
| †Дилофозавр ветхерилли (Уэллс, 1954) | |
| Синонимы | |
| |
Дилофозавр (/dаɪˌлoʊжəˈsɔːrəs,-жoʊ-/[1] dy-LOHF-о-SOR-s ) это род из теропод динозавры что жил в том, что сейчас Северная Америка вовремя Раннеюрский период около 193 миллионов лет назад. Три скелета были обнаружены в северная Аризона в 1940 г., а два наиболее сохранившихся экземпляра были собраны в 1942 г. Самым полным экземпляром стал голотип нового вида в роду Мегалозавр, названный М. ветхерилли к Сэмюэл П. Уэллс в 1954 году. Уэллс нашел более крупный скелет, принадлежащий тому же виду, в 1964 году. Понимая, что на черепе есть гребни, он отнес этот вид к новому роду. Дилофозавр в 1970 году, когда Дилофозавр ветхерилли. Название рода означает «двуногая ящерица», а название вида дано в честь Джона Уэтерилла, Навахо советник. С тех пор были обнаружены новые образцы, в том числе младенец. Следы животного также были приписаны животному, включая следы отдыха. Другой вид, Дилофозавр китайский из Китая, был назван в 1993 году, но позже был обнаружен как принадлежащий к роду Синозавр. Он был обозначен как государственный динозавр из Коннектикут на основе найденных там следов.
При длине около 7 м (23 фута) при весе около 400 кг (880 фунтов), Дилофозавр был одним из первых крупных хищных динозавров и самым крупным известным наземным животным в Северной Америке того времени. Он был стройным и легко сложенным, а череп пропорционально большим, но изящным. Морда была узкой, а верхняя челюсть имела щель или перегиб ниже ноздри. На черепе у него была пара продольных изогнутых гребней; их полная форма неизвестна, но, вероятно, они были увеличены кератин. В нижняя челюсть была тонкой и нежной спереди, но глубокой сзади. Зубы были длинными, изогнутыми, тонкими и сжатыми вбок. Те, что в нижней челюсти, были намного меньше, чем в верхней. Большинство зубов имели зубцы по их переднему и заднему краям. Шея была длинной, а позвонки полые и очень легкие. Руки были мощными, с длинной и тонкой костью плеча. На руках было по четыре пальца; первый был коротким, но сильным и нес большой коготь, два следующих пальца были длиннее и тоньше с меньшими когтями; четвертый был рудиментарный. Бедренная кость была массивной, лапы крепкими, а на пальцах ног были большие когти.
Дилофозавр является членом семьи Дилофозавриды вместе с Драковенатор, группа, помещенная между Coelophysidae а позже - теропод. Дилофозавр были бы активными и двуногими, возможно, охотились на крупных животных; он также мог питаться более мелкими животными и рыбой. Из-за ограниченного диапазона движений и короткости передних конечностей рот мог вместо этого впервые контактировать с добычей. Функция гребней неизвестна; они были слишком слабы для битвы, но, возможно, использовались в визуальный дисплей, Такие как признание видов и половой отбор. Возможно, он быстро рос, достигнув в начале жизни темпов роста от 30 до 35 кг (от 66 до 77 фунтов) в год. Образец голотипа имел несколько палеопатологии, в том числе зажившие травмы и признаки аномалии развития. Дилофозавр известно из Кайентская свита, и жили рядом с динозаврами, такими как Мегапнозавр и Саразавр. Дилофозавр был показан в романе парк Юрского периода и это экранизация, в котором ему были даны вымышленные способности плевать ядом и расширять оборка на шее, а также меньше, чем настоящее животное.
История открытия

Летом 1942 г. палеонтолог Чарльз Л. Кэмп возглавил полевую вечеринку из Музей палеонтологии Калифорнийского университета (UCMP) в поисках окаменелости позвоночные в Округ Навахо в северная Аризона. Слово об этом распространилось среди Коренные американцы там и Навахо Джесси Уильямс привел трех членов экспедиции к некоторым ископаемым костям, которые он обнаружил в 1940 году. Этот район был частью формации Кайента, примерно в 32 км к северу от Кэмерон возле Туба Сити в Резервация индейцев навахо. Три скелета динозавров были обнаружены в багровых тонах. сланец, расположенные в виде треугольника, длиной около 9,1 м (30 футов) с одной стороны. Первая была почти полной, без передней части черепа, частей таза и некоторых позвонков. Второй был сильно разрушен, включая переднюю часть черепа, нижнюю челюсть, некоторые позвонки, кости конечностей и сочлененную руку. Третий был настолько разрушен, что состоял только из фрагментов позвонков. Первый хороший скелет был заключен в гипсовый блок после 10 дней работы и загружен на грузовик, второй скелет был легко собран, так как он почти полностью выветрился из-под земли, а третий скелет почти исчез.[2][3][4]
Практически полный первый образец был очищен и установлен на UCMP под наблюдением палеонтолога. Ванн Лэнгстон, процесс, который занял у трех человек два года. Каркас крепился к стене в барельеф, с загнутым вверх хвостом, шея выпрямлена, а левая нога приподнята для обзора, но остальная часть скелета оставалась в погребальном положении. Поскольку череп был раздавлен, он был реконструирован на основе задней части черепа первого экземпляра и передней части второго. Таз реконструирован после Аллозавр, и стопы также были реконструированы. В то время это был один из наиболее хорошо сохранившихся скелетов динозавра-теропод, хотя и неполный. В 1954 году палеонтолог Сэмюэл П. Уэллс, входивший в группу раскопок скелетов, предварительно описал и назвал этого динозавра новым видом в существующих род Мегалозавр, М. ветхерилли. Почти полный экземпляр (каталогизированный как UCMP 37302) был сделан голотипом вида, а второй экземпляр (UCMP 37303) был сделан паратип. В конкретное имя почитал Джона Уэзерилла, советника навахо, которого Уэллс назвал «исследователем, другом ученых и надежным торговцем». Племянник Уэзерилла, Милтон, первым сообщил экспедиции об окаменелостях. Уэллс поместил новый вид в Мегалозавр из-за схожих пропорций конечностей и M. bucklandii, и потому что он не обнаружил между ними больших различий. В то время, Мегалозавр использовался как "таксон для мусорной корзины ", куда были помещены многие виды теропод, независимо от их возраста и местонахождения.[2][5][3][6]

Уэллс вернулся в Туба-Сити в 1964 г., чтобы определить возраст формации Кайента (предполагалось, Поздний триас в возрасте, тогда как Уэллс думал, что это Рано к Средняя юра ), и обнаружил еще один скелет около 400 м (1⁄4 mi) к югу от того места, где были обнаружены экземпляры 1942 года. Почти полный экземпляр (каталогизированный как UCMP 77270) был собран с помощью Уильяма Брида из Музей Северной Аризоны и другие. При подготовке этого экземпляра выяснилось, что это более крупная особь М. ветхерилли, и что у него было два гребня на верхней части черепа. Первоначально считалось, что один гребень, представляющий собой тонкую костную пластину, является частью отсутствующей левой стороны черепа, которая была вытащена из своего положения с помощью мусорщик. Когда стало очевидно, что это гребень, также стало понятно, что соответствующий гребень должен был находиться с левой стороны, поскольку правый гребень находился справа от средней линии и был вогнутым по своей средней длине. Это открытие привело к повторному исследованию экземпляра голотипа, у которого было обнаружено, что основания из двух тонких, вытянутых вверх костей были раздавлены вместе. Они также представляли собой гребни, но раньше считалось, что они являются частью неправильно расположенной скулы. Два экземпляра 1942 года также оказались молодые, в то время как особь 1964 г. была взрослой особью, примерно на треть крупнее остальных.[2][7][8] Позже Уэллс вспоминал, что считал гребни столь же неожиданными, как нахождение «крыльев у червя».[9]

Впоследствии Уэллс и его помощник исправили настенное крепление голотипа на основе нового скелета, восстановив гребни, переделав таз, удлинив ребра шеи и поместив их ближе друг к другу. Изучив скелеты североамериканских и европейских теропод, Уэллс понял, что динозавр не принадлежал Мегалозавр, и потребовалось новое название рода. В то время не было известно никаких других теропод с большими продольными гребнями на голове, поэтому динозавр вызвал интерес палеонтологов. Был изготовлен слепок с голотипа, слепки которого были розданы по разным экспонатам; чтобы упростить маркировку этих слепков, Уэллс решил дать название новому роду в краткой заметке, а не ждать публикации подробного описания. В 1970 году Уэллс придумал новое название рода Дилофозавр, от греческих слов ди (δι) означает "два", лофос (λόφος), что означает «гребень», и sauros (σαυρος), что означает «ящерица»: «двуглавая ящерица». Уэллс опубликовал подробный остеологический описание Дилофозавр в 1984 году, но не включил экземпляр 1964 года, так как он думал, что он принадлежит к другому роду.[2][7][10][8][11] Дилофозавр был первым хорошо известным тероподом ранней юры и остается одним из наиболее хорошо сохранившихся образцов того времени.[5]
В 2001 году палеонтолог Роберт Дж. Гей идентифицировали останки не менее трех новых Дилофозавр экземпляров (это количество основано на наличии трех фрагментов лобковой кости и двух бедренных костей разного размера) в коллекциях Музея Северной Аризоны. Образцы были найдены в 1978 году в четырехугольнике Рок-Хед, в 190 км (120 миль) от того места, где были обнаружены оригинальные образцы, и были отмечены как «большой теропод». Хотя большая часть материала повреждена, он включает элементы, не сохранившиеся в более ранних образцах, в том числе часть таза и несколько ребер. Некоторые элементы в коллекции принадлежали экземпляру младенца (MNA P1.3181), самому молодому известному экземпляру этого рода и одному из самых ранних известных тероподов из Северной Америки, которому предшествовали лишь некоторые из них. Целофиз образцы. Ювенильный экземпляр включает частичную плечевую кость, частичную малоберцовую кость и фрагмент зуба.[12] В 2005 году палеонтолог Рональд С. Тыкоски назначил образец (TMM 43646-140) из Голд-Спринг, штат Аризона, Дилофозавр, но в 2012 году палеонтолог Мэтью Т. Каррано и его коллеги обнаружили, что он отличается в некоторых деталях.[13][14]
В 2020 году палеонтологи Адам Д. Марш и Тимоти Б. Роу всесторонне переописали Дилофозавр на основе известных к тому времени образцов, включая образец UCMP 77270, который оставался неописанным с 1964 года. Они также удалили некоторые ранее назначенные образцы, посчитав их слишком фрагментарными для идентификации, и переместили типовой карьер.[6] В интервью Марш позвонил Дилофозавр "самый худший из известных динозавров", поскольку животное было плохо изучено, несмотря на то, что оно было обнаружено 80 лет назад. Основная проблема заключалась в том, что предыдущие исследования образцов не прояснили, какие части были оригинальными окаменелостями, а какие были реконструированы в гипсе, однако последующим исследователям оставалась только монография Уэллса 1984 года, на которую можно было положиться для последующих исследований, запутывая понимание анатомии динозавра. Марш провел семь лет, изучая образцы, чтобы прояснить проблемы, связанные с динозавром, в том числе два образца, обнаруженные двумя десятилетиями ранее Роу, его доктором философии. советник.[15]
Ранее присвоенные виды

В 1984 году Уэллс предположил, что образец 1964 года (UCMP 77270) не принадлежит Дилофозавр, но к новому роду, основанному на различиях в черепе, позвонках и бедрах. Он утверждал, что у обоих родов были гребни, но точная форма их была неизвестна в Дилофозавр.[2] Уэллс умер в 1997 году, прежде чем он смог назвать этого предполагаемого нового динозавра, но идея о том, что это два разных рода, с тех пор обычно игнорировалась или забывалась.[5] В 1999 году палеонтолог-любитель Стефан Пикеринг в частном порядке опубликовал новое название. Дилофозавр "Breedorum" основан на образце 1964 года, названном в честь Брида, который помогал в его сборе. Это имя считается nomen nudum, неверно опубликованное имя, и Гей указал в 2005 году, что не существует существенных различий между D. "бородорум" и другие D. Wherilli образцы.[16][17] В 2012 году Каррано и его коллеги обнаружили различия между экземпляром 1964 года и голотипом, но приписали их различиям между людьми, а не видами.[13] В 2014 году палеонтологи Кристоф Хендрикс и Октавио Матеус предположили, что известные образцы могут представлять два вида Дилофозавр основаны на различных характеристиках черепа и стратиграфическом разделении до подробного описания отобранных образцов.[18] В 2020 году Марш и Роу пришли к выводу, что существует только один таксон среди известных Дилофозавр образцы, и что различия между ними были связаны с их разной степенью зрелости и сохранности. Они также не обнаружили значительного стратиграфического разделения между экземплярами.[6]
Практически полный скелет теропод (KMV 8701) был обнаружен в Формация Луфэн, в Провинция Юньнань, Китай, 1987 г. Он похож на Дилофозавр, с парой гребней и промежутком, отделяющим предчелюстную кость от верхней челюсти, но отличается в некоторых деталях. Палеонтолог Шаоцзинь Ху назвал его новым видом Дилофозавр в 1993 г. D. sinensis (с греческого Синай, имея в виду Китай).[19] В 1998 году палеонтолог Мэтью К. Ламанна и его коллеги обнаружили D. sinensis быть идентичным Sinosaurus triassicus, теропод из той же формации, названный в 1940 году.[20] Этот вывод был подтвержден палеонтологом Лидой Син и его коллегами в 2013 году, и хотя палеонтолог Го-Фу Ван и его коллеги согласились, что этот вид принадлежит к Синозавр в 2017 году они предположили, что это может быть отдельный вид, S. sinensis.[21][22]
Описание
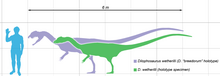
Дилофозавр был одним из первых крупных хищников динозавры, среднего размера теропод хотя и небольшой по сравнению с некоторыми из более поздних теропод.[2][5] Это также было самое крупное из известных наземных животных Северной Америки в раннеюрский период.[6] Стройный и легкий, его размер был сопоставим с размером бурый медведь.[5][23][24] Самый крупный известный экземпляр весил около 400 килограммов (880 фунтов), имел длину около 7 метров (23 фута), а его череп составлял 590 миллиметров (23 1⁄4 в) длинный. Меньший голотип образец весил около 283 кг (624 фунта), был 6,03 метра (19 футов 9 1⁄2 дюймов) в длину, с высотой бедра около 1,36 метра (4 фута 5 1⁄2 дюймов), а его череп был 523 миллиметра (1 фут 8 1⁄2 в) длинный.[23][25] След покоя теропод, похожий на Дилофозавр и Liliensternus интерпретируется некоторыми исследователями как демонстрация впечатлений от перья вокруг живота и ступней, похожие на вниз.[26][27] Другие исследователи вместо этого интерпретируют эти впечатления как седиментологический артефакты, созданные при перемещении динозавра, хотя эта интерпретация не исключает, что следопыт мог иметь перья.[28][29]
Череп

Череп Дилофозавр был большим по сравнению с общим скелетом, но хрупким. Морда при виде спереди была узкой, сужаясь к закругленной вершине. В предчелюстная кость (передняя кость верхней челюсти) была длинной и низкой при осмотре сбоку, выпуклой спереди, а ее внешняя поверхность стала менее выпуклой от морды до ноздри (костной ноздри). Ноздри располагались дальше назад, чем у большинства других теропод. Предчелюстные кости были в тесном сочленении друг с другом, а предчелюстные кости соединялись только с верхняя челюсть (следующая кость верхней челюсти) в середине неба, без каких-либо боковых соединений, они образовали прочный сустав через прочное, взаимосвязанное сочленение между направленными назад и вперед отростками этих костей. Сзади и снизу верхняя челюсть образовывала стенку для промежутка между собой и верхней челюстью, называемого субнариальным промежутком (также называемым «перегибом»). Такой разрыв присутствует и в целофизоиды, а также другие динозавры. Субнарный разрыв привел к диастема, разрыв в зубном ряду (который также называют «выемкой»). Внутри субнариальной щели была глубокая выемка позади зубного ряда предчелюстной кости, называемая субнариальной ямкой, которая была окружена нижним килем предчелюстной кости.[2][23][30][7][6]
Наружная поверхность предчелюстной кости была покрыта отверстие (отверстия) разного размера. Верхний из двух идущих назад отростков предчелюстной кости был длинным и низким и составлял большую часть верхней границы удлиненного носа. Он имел наклон в сторону купели, что делало область у основания вогнутой в профиль. Нижняя часть предчелюстной кости, содержащая альвеолы (лунки зубов) овальные. Верхняя челюсть неглубокая и вдавлена вокруг анторбитальное окно (большое отверстие перед глазом), образуя углубление, которое закруглено спереди и более гладкое, чем остальная часть верхней челюсти. В это углубление на переднем изгибе выходило отверстие, называемое преанторбитальным отверстием. Большие отверстия проходили сбоку от верхней челюсти над альвеолами. Глубокая питательная канавка шла назад от субнариальной ямки вдоль основания межзубные пластины (или морщинистые) верхней челюсти.[2]
Дилофозавр носил пару высоких, тонких и изогнутых (или пластинчатых) гребней на крыше черепа. Гребни (называемые носослезными гребнями) начинались как низкие гребни на предчелюстных костях и в основном образовывались расширенными вверх. носовой и слезные кости. Эти кости были освеженный вместе (слияние при образовании костной ткани), поэтому швы между ними невозможно определить. Слезная кость превратилась в толстую, морщинистую. предглазничный выступ, образующий дугу на верхней передней границе орбита (глазница) и поддерживала нижнюю часть задней части гребня. Уникально для этого рода ободок над орбитой продолжался назад и заканчивался небольшим, почти треугольным отростком позади орбиты, который слегка изгибался наружу. Поскольку только короткая часть верхней поверхности этого отростка не повреждена, остальная часть гребня могла подниматься над черепом на расстояние ~ 12 миллиметров (0,47 дюйма). Сохранившаяся часть гребня в UCMP 77270 является самой высокой в районе середины длины анторбитального отверстия. UCMP 77270 сохраняет вогнутую полку между основаниями гребней, и, если смотреть спереди, они выступают вверх и в стороны под углом ~ 80 °. Уэллс обнаружил, что гребешки напоминают двугорбый казуар, а Марш и Роу заявили, что они, вероятно, были покрыты кератин или ороговевшая кожа. Они отметили, что по сравнению с цесарка в шлеме, кератин на гребнях Дилофозавр мог бы увеличить их намного больше, чем указано на кости. Поскольку только один экземпляр сохранил большую часть гребней, неизвестно, различались ли они у разных особей.[6][2][7][10][5][13]
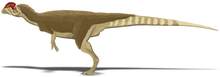
Глазница была овальной, к низу суженной. В скуловая кость имел два обращенных вверх отростка, первый из которых составлял часть нижнего края анторбитального отверстия и часть нижнего края орбиты. Проекция из квадратная кость в латеральное височное отверстие (открывая глаза) дал этому почковидный (почковидный) очертание. В большое затылочное отверстие (большое отверстие в задней части черепная коробка ) составлял примерно половину ширины затылочного мыщелка, который сам сердцевидный (в форме сердца), имел короткую шейку и бороздку сбоку.[2] В нижняя челюсть был тонким и тонким спереди, но суставная область (там, где он соединялась с черепом) была массивной, а нижняя челюсть была глубокой вокруг нижнечелюстное отверстие (отверстие сбоку). Нижнечелюстное отверстие было маленьким в Дилофозаврпо сравнению с целофизоидами и уменьшена спереди назад, что является уникальным для этого рода. В зубная кость (передняя часть нижней челюсти, где было прикреплено большинство зубов) имела скорее загнутый вверх, чем заостренный подбородок. Подбородок имел большое отверстие на кончике, и ряд маленьких отверстий шел примерно параллельно верхнему краю зубной кости. На внутренней стороне нижнечелюстной симфиз (где две половины нижней челюсти соединены) была плоской и гладкой и не показывала никаких признаков слияния с противоположной половиной. А Меккелевское отверстие проходил по внешней стороне зубной кости. Боковая поверхность надугловая кость имел уникальный пирамидальный отросток перед сочленением с квадратом, и этот горизонтальный гребень образовывал полку. В ретроартикулярный отросток нижней челюсти (обратная проекция) длинный.[2][30][6]
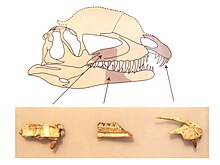
Дилофозавр имел четыре зуба в каждой предчелюстной кости, 12 в каждой верхней челюсти и 17 в каждой зубной кости. Зубы обычно были длинными, тонкими и загнутыми назад, с относительно небольшими основаниями. Они сжатые вбок, овальные в поперечном сечении у основания, линзовидные (линзовидные) сверху и слегка вогнутые с внешней и внутренней стороны. Самый крупный зуб верхней челюсти находился либо в четвертой альвеоле, либо рядом с ней, а высота коронок зубов уменьшалась кзади. Первый зуб верхней челюсти немного направлен вперед от альвеолы, потому что нижняя граница прексамиллярного отростка (которая выступала назад к верхней челюсти) была перевернута. Зубы зубной кости были намного меньше, чем зубы верхней челюсти. Третий или четвертый зуб в зубной части Дилофозавр и некоторые целофизоиды были там самыми крупными и, кажется, вошли в субнариальную щель верхней челюсти. Большинство зубов имело зазубрины на переднем и заднем краях, которые были смещены вертикальными канавками и были меньше на передней части. На передних краях было от 31 до 41 зубцов, а сзади от 29 до 33. По крайней мере, второй и третий зубы предчелюстной кости имели зубцы, а четвертый - нет. Зубы были покрыты тонким слоем эмаль Толщиной от 0,1 до 0,15 мм (от 0,0039 до 0,0059 дюйма), которая простиралась далеко к их основаниям. Альвеолы были эллиптическими или почти круглыми, и все они были больше, чем основания содержащихся в них зубов, которые, следовательно, могли свободно удерживаться в челюстях. Хотя количество альвеол в зубной части, казалось бы, указывает на то, что зубы были очень скученными, они находились довольно далеко друг от друга из-за большего размера их альвеол. Челюсти содержали замена зубов на разных стадиях прорезывания. Межзубные пластины между зубами очень низкие.[2][30][12]
Посткраниальный скелет

Дилофозавр имел 10 шейных (шея), 14 спинных (спинных) и 45 хвостовых (хвостовых) позвонков. У него была длинная шея, которая, вероятно, была изогнута почти на 90 ° черепом и плечом, удерживая череп в горизонтальном положении. Шейные позвонки необычайно легкие; их центры ("тела" позвонков) были выдолблены плевроцели (впадины по бокам) и центроцели (полости внутри). Дуги шейных позвонков также имели хонозы, конические углубления, настолько большие, что разделяющие их кости иногда были тонкими как бумага. Центры были плоско-вогнутыми, плоскими или слабо выпуклыми спереди и глубоко чашевидными (или вогнутыми) сзади, как и Цератозавр. Это указывает на то, что шея была гибкой, хотя у нее были длинные перекрывающиеся шейные ребра, которые срослись с центральными. Шейные ребра были тонкими и могли легко сгибаться.[2][30]
В атласная кость (первый шейный позвонок, который прикрепляется к черепу) имел небольшой кубический центр и вогнутость спереди, где он образовывал чашу для затылочный мыщелок (выпуклость, которая соединяется с атласным позвонком) в задней части черепа. В ось кость (второй шейный позвонок) имел тяжелый позвоночник, а его постзигапофизы (отростки позвонков, которые сочленяются с презигапофизами следующих позвонков) были встречены длинными презигапофизами, которые изгибались вверх от третьего шейного позвонка. Центральные и нервные отростки шейных позвонков были длинными и низкими, а при виде сбоку они были ступенчатыми, образуя «плечи» спереди и сзади, а также более высокие центральные «шапки», которые создавали вид Мальтийский крест (крестообразный) при взгляде сверху отличительные черты этого динозавра. Задняя центродиапофизарная пластинка шейных позвонков показала серию вариаций, раздвоение и воссоединение вниз по шее, что является уникальной особенностью. Нервные отростки спинных позвонков также были низкими и расширенными спереди и сзади, которые образовывали прочные прикрепления для связки. Уникально для этого рода дополнительные пластинки исходят из передней центродиапофизарной пластинки среднего туловища и задней центродиапофизарной пластинки. В крестцовые позвонки который занимал длину подвздошная кость лезвие, похоже, не переплавилось. Ребро первого крестцового позвонка сочленено с преацетабулярным отростком подвздошной кости - отличительный признак. Центры хвостовых позвонков были очень одинаковой длины, но их диаметр стал меньше к спине, и они перешли от эллиптических к круглым в поперечном сечении.[2][30][6]

В лопатки (лопатки) были средней длины и вогнуты с внутренней стороны, что соответствовало кривизне тела. Лопатки были широкими, особенно их верхняя часть была прямоугольной (или квадратной), что было уникальной особенностью. В коракоиды были эллиптическими и не срослись с лопатками. Нижние задние части коракоидов имели «горизонтальную опору» рядом с бугорком двуглавой мышцы, уникальную для этого рода. Руки были мощными, с глубокими ямками и крепкими отростками для прикрепления мышц и связок. В плечевая кость (кость плеча) была большой и тонкой, с толстыми надпочечниками и локтевая кость (нижняя кость) была толстой и прямой, с толстым олекранон. На руках было четыре пальца: первый был короче, но сильнее следующих двух пальцев, с большим когтем, а два следующих пальца были длиннее и тоньше, с меньшими когтями.Когти были изогнутыми и острыми. Третий палец был уменьшен, а четвертый - рудиментарный (сохранено, но без функции).[2][30][6]
В гребень подвздошной кости была выше подвздошной цветонос (нижний отросток подвздошной кости), а его внешняя сторона вогнутая. Подножие лобковая кость был лишь немного расширен, тогда как нижний конец был значительно расширен на седалищная кость, который также имел очень тонкий вал. Задние лапы были большими, чуть длиннее. бедренная кость (бедренная кость) чем большеберцовая кость (кость голени), противоположность, например, Целофиз. Бедренная кость была массивной; его вал был сигмовидный -образный (изогнутый как 'S'), и его большой вертел был отцентрован на валу. Большеберцовая кость имела развитую бугристость и был расширен на нижнем конце. В кость астрагала (лодыжка) была отделена от большеберцовой кости и пяточная кость, и сформировал половину лунки для малоберцовой кости. У него были длинные крепкие лапы с тремя хорошо развитыми пальцами с большими когтями, которые были гораздо менее изогнутыми, чем у кисти. Третий палец был самым крепким, а первый палец меньшего размера ( Hallux ) был оторван от земли.[2][30][31][6]
Классификация

Уэллс подумал Дилофозавр а мегалозавр в 1954 г., но пересмотрел свое мнение в 1970 г. после обнаружения гребней.[7][3] К 1974 году Уэллс и палеонтолог Роберт А. Лонг обнаружили Дилофозавр быть цератозавроид.[32] В 1984 году Уэллс обнаружил, что Дилофозавр выставлены черты обоих Целурозаврия и Карнозаврия, две основные группы, на которые до сих пор делились теропод, основанные на размере тела, и он предположил, что это разделение было неточным. Он нашел Дилофозавр быть ближе всего к тем тероподам, которые обычно помещались в семейство Halticosauridae, особенно Liliensternus.[2]
В 1988 г. палеонтолог Грегори С. Пол классифицировал гальтикозавров как подсемейство семейства Coelophysidae, и предположил, что Дилофозавр мог быть прямым потомком Целофиз. Павел также рассмотрел возможность того, что спинозавры были поздно выжившими дилофозаврами, основываясь на сходстве изогнутой морды, положения ноздрей и тонких зубов Барионикс.[23] В 1994 г. палеонтолог Томас Р. Хольц размещен Дилофозавр в группе Coelophysoidea, вместе с Coelophysidae, но отдельно от них. Он поместил Coelophysoidea в группу Ceratosauria.[33] В 2000 году палеонтолог Джеймс Х. Мэдсен и Уэллс разделили Ceratosauria на семьи Ceratosauridae и Дилофозавриды, с Дилофозавр как единственный член последней семьи.[34]

Ламанна и его коллеги отметили в 1998 г., что с тех пор, как Дилофозавр было обнаружено, что на черепе были гребни, были обнаружены другие тероподы с таким же гребнем (в том числе Синозавр), и, следовательно, эта особенность не является уникальной для данного рода и имеет ограниченное использование для определения взаимоотношений внутри их группы.[20] Палеонтолог Адам М. Йейтс описал род Драковенатор из Южной Африки в 2005 году и обнаружил, что он тесно связан с Дилофозавр и Зупайзавр. Его кладистический анализ предположили, что они принадлежат не к Coelophysoidea, а скорее к Neotheropoda, более полученный (или «продвинутая») группа. Он предложил, если Дилофозавр был более производным, чем Coelophysoidea, черты, которые он разделял с этой группой, могли быть унаследованы от базальный (или «примитивные») тероподы, что указывает на то, что тероподы, возможно, прошли «стадию целофизоидов» в своей ранней эволюции.[35]
В 2007 году палеонтолог Натан Д. Смит и его коллеги обнаружили хохлатого теропода. Криолофозавр быть родственные виды из Дилофозавр, и сгруппировал их с Драковенатор и Синозавр. Эта клада была более производной, чем Coelophysoidea, но более базальной, чем Ceratosauria, тем самым размещая базальные теропод в виде лестницы.[36] В 2012 году Каррано и его коллеги обнаружили, что группа гребенчатых теропод, предложенная Смитом и его коллегами, основана на особенностях, которые связаны с наличием таких гребней, но что особенности остальной части скелета были менее согласованными. Вместо этого они обнаружили, что Дилофозавр был целофизоидом, с Криолофозавр и Синозавр будучи более производными, так как основные члены группы Столбняк.[13]
Палеонтолог Кристоф Хендрикс и его коллеги определили, что Dilophosauridae включают Дилофозавр и Драковенатор в 2015 г. и отметил, что, хотя существует общая неопределенность в отношении размещения этой группы, она, по-видимому, несколько более производная, чем Coelophysoidea, и сестринская группа по отношению к Averostra. У Dilophosauridae есть общие черты с Coelophysoidea, такие как субнариальная щель и передние зубы верхней челюсти, направленные вперед, в то время как черты, общие с Averostra, включают в себя отверстие в передней части верхней челюсти и уменьшенное количество зубов в верхней челюсти. Они предположили, что черепные гребни Криолофозавр и Синозавр имел либо развивались конвергентно, или были функцией, унаследованной от общего предка. Следующее кладограмма основан на опубликованном Hendrickx и его коллегами, основанном на более ранних исследованиях:[37]
| Neotheropoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
В 2019 году палеонтологи Марион Захнер и Винанд Бринкманн обнаружили, что представители Dilophosauridae являются последовательными базальными сестринскими таксонами аверостры, а не монофилетический clade (естественная группа), но отметили, что некоторые из их анализов действительно нашли группу действительной, содержащую Дилофозавр, Драковенатор, Криолофозавр, и возможно Notatesseraeraptor как самый базальный член. Поэтому они поставили диагноз Dilophosauridae на основании особенностей нижней челюсти.[38] В филогенетическом анализе, сопровождавшем их редескрипцию 2020 г., Марш и Роу обнаружили все образцы Дилофозавр сформировать монофилетическую группу, сестру Аверостры и более производную, чем Криолофозавр. Их анализ не нашел подтверждения для Dilophosauridae, и они предположили, что черепные гребни были плезиоморфный (наследственный) признак Ceratosauria и Tetanurae.[6]
Технология

Разные ихнотакса (таксоны на основе следы окаменелостей ) были отнесены к Дилофозавр или подобные теропод. В 1971 году Уэллс сообщил о следах динозавров из формации Кайента в северной Аризоне, на двух уровнях, на 14 м (45 футов) и 112 м (367 футов) ниже того места, где находился оригинал. Дилофозавр экземпляров не обнаружено. Нижние следы были тридактиль (трехпалый), и мог быть сделан Дилофозавр; Уэллс создал новый ихноген и вид Dilophosauripus williamsi на их основе, в честь Уильямса, первооткрывателя первого Дилофозавр скелеты. Типовой образец представляет собой слепок большого отпечатка, занесенного в каталог как UCMP 79690-4, с слепками трех других отпечатков, включенных в гиподигму.[39] В 1984 году Уэллс признал, что не было найдено способа доказать или опровергнуть, что следы принадлежали Дилофозавр.[2] В 1996 году палеонтологи Майкл Моралес и Скотт Балки сообщили о путь ихногена Eubrontes из кайентской свиты сделан очень крупным тероподом. Они отметили, что это могло быть сделано очень большим Дилофозавр человек, но обнаружил, что маловероятно, по их оценке, высота гусеницы составляла 2,83–2,99 м (9 футов 3 1⁄2 дюйм – 9 футов 9 3⁄4 дюйма) высотой до бедер по сравнению с 1,50–1,75 м (4 фута 11 дюймов - 5 футов 9 дюймов) Дилофозавр.[40]

Палеонтолог Герард Герлинский исследовал следы трехактилей на Горы Святого Креста в Польша и пришел к выводу в 1991 году, что они принадлежали тероподу, подобному Дилофозавр. Он назвал новые их виды Grallator (Eubrontes) soltykovensis на их основе, с голотипом отпечатка ноги MGIW 1560.11.12.[41] В 1994 году Герлинский также установил следы Формация Хёганас в Швеции обнаружили в 1974 г. G. (E.) soltykovensis.[42] В 1996 году Герлински приписал трассу AC 1/7 от Формация Тернерс Фолс Массачусетса, след покоя, который, как он считал, показывает отпечатки перьев теропод, похожий на Дилофозавр и Liliensternus, и отнесли его к ихнотаксону Grallator minisculus.[26] Палеонтолог Мартин Кундрат согласился, что на следе были отпечатки перьев в 2004 году, но палеонтолог оспорил эту интерпретацию. Мартин Локли и коллеги в 2003 г. и палеонтолог Энтони Дж. Мартин и коллегами в 2004 г., которые считали их седиментологическими артефактами. Мартин и его коллеги также переназначили трек ихнотаксону. Fulicopus lyellii.[27][28][29]
Палеонтолог Роберт Э. Вимс в 2003 г. предположил, что Eubrontes следы были произведены не тероподом, а завроподоморф похожий на Платеозавр, без учета Дилофозавр как возможный трекмейкер. Вместо этого Вимс предложил Kayentapus hopii, еще один ихнотаксон, названный Уэллсом в 1971 г. лучшим соответствием Дилофозавр.[43] Приписывание Дилофозавр в первую очередь было основано на большом угле между отпечатками третьего и четвертого пальцев, показанных этими следами, и наблюдении, что на ноге голотипа виден четвертый палец с аналогичным расширением. Также в 2003 году палеонтолог Эмма Рейнфорт утверждала, что растяжение стопы голотипа было всего лишь результатом искажения, и что Eubrontes действительно будет хорошей парой для Дилофозавр.[44][45]Палеонтолог Спенсер Дж. Лукас и коллеги заявили в 2006 году, что существует практически универсальное соглашение о том, что Eubrontes следы были сделаны тероподом как Дилофозавр, и что они и другие исследователи отклонили претензии Уимса.[46]
В 2006 году Уимс защитил свою оценку 2003 года. Eubrontes, и предложил животное вроде Дилофозавр как возможный создатель множества Кайентапус следы карьера Калпепер в Вирджинии. Вимс предложил округлые отпечатки, связанные с некоторыми из этих путей, чтобы представить отпечатки рук без отпечатков пальцев, которые он интерпретировал как след движения на четвероногих.[45] Милнер и его коллеги использовали новая комбинация Kayentapus soltykovensis в 2009 г. и предложил Дилофозаврип не может отличаться от Eubrontes и Кайентапус. Они предположили, что длинные следы когтей, которые использовались для различения Дилофозаврип может быть артефактом перетаскивания. Они обнаружили, что Гигандипус и Анхизаврип треки могут также просто представлять вариации Eubrontes. Они указали, что различия между ихнотаксонами могут отражать скорее то, как трекмейкер взаимодействует с субстратом, чем таксономию. Они также нашли Дилофозавр быть подходящей партией для Eubrontes путь и след покоя (SGDS 18.T1) от Место открытия динозавров Святого Георгия в Формация Моэнав Юты, хотя сам динозавр неизвестен по формации, которая немного старше формации Кайента.[47] Вимс заявил в 2019 году, что Eubrontes следы не отражают изящных ног Дилофозавр, и утверждал, что они были созданы двуногим зауроподопормфом. Анхизавр.[48]
Палеобиология

Кормление и диета
Уэллс обнаружил, что Дилофозавр не имел сильного прикуса из-за слабости, вызванной субнарным разрывом. Он думал, что он использовал свои передние предчелюстные зубы для выщипывания и разрыва, а не кусания, а верхние зубы, расположенные дальше назад, для прокалывания и надрезания. Он думал, что это, вероятно, падальщик, а не хищник, и что если бы он действительно убивал крупных животных, то делал бы это руками и ногами, а не челюстями. Уэллс не нашел доказательств черепной кинезис в черепе Дилофозавр, функция, которая позволяет отдельным костям черепа перемещаться относительно друг друга.[2] В 1986 году палеонтолог Роберт Т. Баккер вместо этого нашел Дилофозаврс массивной шеей и черепом и большими верхними зубами, он был приспособлен для убийства крупной добычи и достаточно силен, чтобы атаковать любых травоядных животных ранней юры.[49] В 1988 году Пол отверг идею о том, что Дилофозавр был мусорщиком и утверждал, что наземные животные - это просто миф. Он заявил, что морда Дилофозавр был закреплен лучше, чем считалось ранее, и что очень большие и тонкие зубы верхней челюсти были более смертоносными, чем когти. Пол предположил, что он охотился на крупных животных, таких как прозауроподы, и что он был более способен хватать мелких животных, чем другие тероподы такого же размера.[23]
2005 г. теория пучка исследование палеонтолога Франсуа Терриена и его коллег показало, что сила укуса в нижней челюсти Дилофозавр быстро уменьшалась кзади при броске зубов. Это указывает на то, что передняя часть нижней челюсти с вздернутым подбородком "розетка "зубов и усиленная симфизальная область (подобная спинозавридам), использовалась для захвата и манипулирования добычей, вероятно, относительно меньшего размера. Свойства его нижнечелюстного симфиза были аналогичны свойствам кошачьи и крокодилы, которые используют переднюю часть своих челюстей, чтобы нанести мощный укус при покорении добычи. Нагрузки, оказываемые на челюсти, соответствовали борьбе с мелкой добычей, на которую, возможно, охотились, нанося рубящие укусы, чтобы поранить ее, а затем захватывать передней частью челюстей после того, как они были слишком ослаблены, чтобы сопротивляться. Добыча могла быть перемещена дальше назад в челюсти, где были расположены самые большие зубы, и убита путем нарезания укусов (как у некоторых крокодилов) сжатыми вбок зубами. Авторы предположили, что если Дилофозавр действительно питались мелкой добычей, возможные охотничьи стаи были бы ограниченного размера.[50]
Милнер и палеонтолог Джеймс И. Киркланд предложил в 2007 г. Дилофозавр имел признаки, указывающие на то, что он, возможно, ел рыбу. Они отметили, что концы челюстей были расширены в стороны, образуя «розетку» сцепленных зубов, подобных тем, что у спинозаврид, которые, как известно, поедали рыбу, и гавиалы, который является современным крокодил который ест больше всего рыбы. Носовые отверстия также были втянуты назад на челюстях, подобно спинозавридам, у которых носовые отверстия еще больше втянуты, и это могло ограничивать попадание воды в ноздри во время рыбалки. У обеих групп также были длинные руки с хорошо развитыми когтями, которые могли помочь при ловле рыбы. Озеро Дикси, большое озеро, простирающееся от Юты до Аризоны и Невады, должно было стать источником обилия рыбы в «посткатаклизмическом», биологически более бедном мире, который последовал за Триас-юрское вымирание.[51]

В 2018 году Марш и Роу сообщили, что экземпляр голотипа зауроподоморфа Саразавр имел возможные следы зубов, разбросанные по скелету, которые могли быть оставлены Дилофозавр (Syntarus был слишком мал, чтобы произвести их), собирая образец после того, как он умер (положение костей также могло быть нарушено в результате уборки). Пример таких следов можно увидеть на левой лопатке, которая имеет овальное углубление на поверхности ее верхней стороны и большое отверстие на нижнем переднем конце правой большеберцовой кости. Карьер, где найдены голотип и паратипные экземпляры Саразавр были раскопаны также содержали частично незрелые Дилофозавр образец.[52] В 2020 году Марш и Роу предположили, что многие отличительные особенности Дилофозавр от более ранних теропод были связаны с увеличением размеров тела и макропредами (охотой на крупных животных). Хотя Марш и Роу согласились, что Дилофозавр могли питаться рыбой и мелкой добычей в речной системе в окружающей среде, они отметили, что сочленение между предчелюстной и верхней челюстью было неподвижным и гораздо более прочным, чем считалось ранее, и что крупная добыча могла быть хватали и манипулировали передними конечностями во время хищничества и уборки мусора. Они считали большие следы укусов на Саразавр образцы рядом с поливными зубами и наличие Дилофозавр образец в том же карьере как подтверждение этой идеи.[6]
Движение

Уэллс предвидел Дилофозавр как активное, явно двуногое животное, похожее на увеличенное страус. Он обнаружил, что передние конечности были мощным оружием, сильным и гибким, и не использовались для передвижения. Он отметил, что руки были способны хватать и рубить, встречаться друг с другом и достигать двух третей шеи. Он предложил, чтобы в сидячем положении животное опиралось на большую «ступню» седалищной кости, а также на хвост и лапы.[2] В 1990 году палеонтологи Стивен и Сильвия Черкас предположили, что слабый таз Дилофозавр мог быть адаптацией к водному образу жизни, когда вода поддерживала его вес, и что он мог быть хорошим пловцом. Однако они сочли сомнительным, что он мог быть ограничен водной средой из-за силы и пропорций его задних конечностей, которые сделали бы его быстроногим и подвижным во время двуногого передвижения.[53] Павел изобразил Дилофозавр подпрыгивая на хвосте, набрасываясь на врага, как кенгуру.[54]
В 2005 году палеонтологи Фил Сентер и Джеймс Х. Робинс исследовали диапазон движений передних конечностей Дилофозавр и другие теропод. Они обнаружили, что Дилофозавр мог бы отвести плечевую кость назад до тех пор, пока она не стала почти параллельной лопатке, но не мог переместить ее вперед до более чем вертикальной ориентации. Локоть может приближаться к полному разгибанию и сгибанию под прямым углом, но не полностью. Пальцы, по-видимому, не были произвольно чрезмерно растянуты (могли вытягиваться назад за пределы их нормального диапазона), но они могли быть пассивно гиперрастяжимыми, чтобы противостоять вывихам во время резких движений захваченной добычи.[55] В статье Сентер и Робинс, опубликованной в 2015 году, были даны рекомендации по восстановлению осанки передних конечностей у двуногих динозавров, основанные на исследовании различных таксонов, включая Дилофозавр. Лопатки держались очень горизонтально, положение локтя в покое было близко к прямому углу, а положение руки не сильно отклонялось от ориентации предплечья.[56]

В 2018 году Сентер и Корвин Салливан исследовали диапазон движений в суставах передних конечностей. Дилофозавр манипулируя костями, чтобы проверить предполагаемые функции передних конечностей. Они также учли, что эксперименты с аллигатор туши показывают, что диапазон движений больше в локтях, покрытых мягкими тканями (такими как хрящ, связки и мышцы), чем то, на что указывает манипуляция с голыми костями. Они обнаружили, что плечевая кость Дилофозавр мог быть втянут в положение, которое было почти параллельно лопатке, вытягивалось почти до вертикального уровня и поднималось на 65 °. Локоть нельзя было согнуть под прямым углом к плечевой кости. Пронация и супинация запястий (пересечение радиус и локтевая кость нижней части руки для поворота руки) предотвращалось из-за того, что лучевой и локтевой суставы не могли вращаться, и, следовательно, ладони были обращены медиально друг к другу. Неспособность пронировать запястья была наследственной особенностью теропод и других групп динозавров. Запястье имело ограниченную подвижность, пальцы при сгибании расходились и были очень сильно разгибаемы.[57]
Сентер и Салливан пришли к выводу, что Дилофозавр мог захватывать и удерживать предметы двумя руками, захватывать и удерживать мелкие предметы в одной руке, хватать предметы близко под грудью, подносить предмет ко рту, показывать изображение, размахивая руками по дуге вдоль стороны грудной клетки, чтобы почесать грудь, живот или половину другой передней конечности, наиболее удаленную от тела, захватить добычу под грудью или основанием шеи и прижать предметы к груди. Дилофозавр не мог копать царапины, тянуть за крюк, удерживать предметы двумя кончиками пальцев одной руки, поддерживать равновесие, вытягивая руки в стороны, или прощупывать небольшие щели, как современные да да делает. Гиперэксенсибилизация пальцев могла препятствовать жесткой борьбе жертвы с их вывихом, поскольку это позволяло бы больше движения пальцев (без важности для передвижения). Ограниченная подвижность плеча и короткие передние конечности указывают на то, что в первую очередь с жертвой контактировал рот, а не руки. Захват добычи передними конечностями возможен только для животных, достаточно маленьких, чтобы поместиться ниже груди Дилофозавр, или более крупная добыча, которую прижали пастью. Большая длина головы и шеи позволила бы морде вытянуться намного дальше, чем руки.[57]


В Дилофозаврип Следы, о которых сообщил Уэллс в 1971 году, все были на одном уровне и были описаны как «солянка из птичьего двора», состоящая из следов, немногие из которых образуют дорожку. Следы были отпечатаны в грязи, что позволяло ступням опускаться на 5–10 см (2–4 дюйма). Отпечатки были неряшливыми, а разная ширина отпечатков пальцев указывает на то, что к ступням прилипла грязь. Отпечатки варьировались в зависимости от различий в субстрате и способах их изготовления; иногда ступня была поставлена прямо, но часто происходило скольжение вперед или назад, когда ступня опускалась. Положение и углы пальцев ног также значительно различались, что указывает на то, что они должны были быть довольно гибкими. В Дилофозаврип Следы имели смещенный второй палец с толстым основанием и очень длинные прямые когти, которые совпадали с осями подушечек пальцев. На одном из следов не было когтя второго пальца, возможно, из-за травмы.[39] В 1984 году Уэллс истолковал тот факт, что три человека были обнаружены близко друг к другу, и наличие перекрещенных путей поблизости, как указание на то, что Дилофозавр путешествовали группами.[2] Гей согласился с тем, что они могли путешествовать небольшими группами, но отметил, что нет прямых доказательств того, что это подтверждается. паводки мог собрать разбросанные кости у разных людей и сложить их вместе.[12]
Милнер и его коллеги изучили возможные Дилофозавр рельсовый путь SGDS 18.T1 в 2009 году, который состоит из типичных следов с хвостовыми волочениями и более необычного следа покоя, хранящегося в озерный пляж песчаник. Путь начинался с того, что животное сначала ориентировалось примерно параллельно береговой линии, а затем остановилось на берма обеими ногами параллельно, после чего он опустил свое тело и плюсневые кости и мозоль вокруг седалищной кости до земли; это создавало впечатление симметричных «пяток» и круглых отпечатков седалищной кости. Часть хвоста, ближайшая к телу, держалась над землей, в то время как конец, более удаленный от тела, касался земли. Тот факт, что животное отдыхало на склоне, позволил ему опустить обе руки на землю рядом с ногами. Отдохнув, динозавр пополз вперед и оставил новые отпечатки ступнями, плюсневыми костями и седалищной костью, но не руками. Теперь правая ступня наступила на отпечаток правой руки, а второй коготь левой ступни оставил след от первого положения покоя к следующему. Через некоторое время животное встало и двинулось вперед, сначала поставив левую ногу, и после того, как полностью выпрямилось, оно пошло по остальной открытой поверхности, оставляя тонкие следы на конце хвоста.[47]
Приседание - это редко фиксируемое поведение теропод, и SGDS 18.T1 - единственный такой след с однозначными отпечатками рук теропод, который дает ценную информацию о том, как они использовали свои передние конечности. Поза приседания была очень похожа на позу современных птиц и показывает, что ранние тероподы держали ладони ладонями друг к другу медиально. Так как такая поза возникла на ранних этапах линии передачи, она, возможно, была характерна для всех теропод. Их часто изображают ладонями вниз, но исследования их функциональной анатомии показали, что они, как и птицы, не могли пронировать или супинировать руки. . След показал, что ноги держатся симметрично, а вес тела распределяется между ступнями и плюсневыми костями, что также характерно для таких птиц, как ратиты. Милнер и его коллеги также отвергли идею о том, что Kayentapus minor След, о котором сообщил Вимс, показал отпечаток ладони, сделанный четвероногим тероподом. Вимс предположил, что гусеничный мастер мог бы двигаться на четвероногих при медленной ходьбе, в то время как пальцы обычно были бы чрезмерно вытянуты, так что только ладони касались земли. Милнер и его коллеги сочли предполагаемую позу ненужной и предположили, что трек был сделан таким же образом, как SGDS 18.T1, но без следов цифр.[47]
Функция гребня

Уэллс признал, что предположения относительно функции гребней Дилофозавр были предположительными, но считали, что, хотя гребни не имели бороздок, указывающих на васкуляризацию, они могли использоваться для терморегуляция. Он также предположил, что их можно было использовать для признание видов или же украшения.[2]Черки указали, что гребни нельзя было использовать во время боя, так как их хрупкая конструкция была бы легко повреждена. Они предположили, что они визуальный дисплей для привлечения спутника жизни и даже терморегуляции.[53] В 1990 году палеонтолог Уолтер П. Кумбс заявил, что гребни могли быть усилены цветами для демонстрации.[58]
В 2011 году палеонтологи Кевин Падиан и Джон Р. Хорнер предположил, что «причудливые структуры» динозавров в целом (включая гребни, оборки, рога и купола) в основном использовались для распознавания видов, и отклонил другие объяснения как не подтвержденные доказательствами. Они отметили, что слишком мало образцов черепно-орнаментированных теропод, в том числе Дилофозавр, было известно, что они проверяли свою эволюционную функцию статистически, и представляли ли они половой диморфизм или же половая зрелость.[59] В ответ на Падиана и Хорнера в том же году палеонтологи Роб Дж. Нелл и Скотт Д. Сэмпсон утверждал, что распознавание видов было вполне вероятным как вторичная функция для «причудливых структур» у динозавров, но что половой отбор (использовались в демонстрации или в бою, чтобы соревноваться за помощников) было более вероятным объяснением из-за высокой стоимости их разработки и из-за того, что такие структуры, по-видимому, сильно варьируются в пределах вида.[60]
В 2013 году палеонтологи Дэвид Э. Хоун и Даррен Нейш критиковал «гипотезу распознавания видов» и утверждал, что ни одно из ныне живущих животных не использует такие структуры в первую очередь для распознавания видов, и что Падиан и Хорнер проигнорировали возможность взаимного полового отбора (когда оба пола украшены орнаментом).[61] Марш и Роу согласились в 2020 году, что гребни Дилофозавр вероятно, сыграл роль в идентификации видов или интерсексуальном / внутриполом отборе, как у некоторых современных птиц.[6]
Разработка

Первоначально Уэллс интерпретировал меньшее Дилофозавр особи как молодые особи, а более крупные особи как взрослые особи, позже интерпретируя их как разные виды.[2][7] Пол предположил, что различия между образцами, возможно, связаны с половым диморфизмом, что, по-видимому, также очевидно в Целофиз, которые имели «крепкие» и «изящные» формы одинакового размера, которые в противном случае могли бы рассматриваться как отдельные виды. Следуя этой схеме, меньший Дилофозавр образец представляет собой «изящный» пример.[23]
В 2005 году Тыкоски обнаружил, что большинство Дилофозавр Известные экземпляры были молодыми особями, из которых взрослые особи были самыми крупными, судя по степени сросшихся костей.[14] В 2005 году Гей не обнаружил доказательств полового диморфизма, предложенного Полом (но предположительно присутствующего в Целофиз), и объяснил разницу между Дилофозавр образцы в индивидуальную вариацию и онтогенез (меняется в процессе роста). В скелетах не было диморфизма, но он не исключил, что мог быть в гребнях; для определения этого требовалось больше данных.[16] Основываясь на крошечных носовых гребнях у ювенильного экземпляра, Йейтс был предварительно отнесен к родственному роду. Драковенатор, он предположил, что они стали больше, когда животное стало взрослым.[35]
Палеонтолог И.С. Ткач сообщил о гистологический исследование (микроскопическое изучение внутренних особенностей) Дилофозавр в 1996 г. проведено взятием шлифы длинных костей и ребер образца UCMP 37303 (наименее сохранившийся из двух оригинальных скелетов). Костные ткани были хорошо васкуляризированы и имели фиброзно-ламеллярную структуру, аналогичную той, что обнаруживается у других теропод и зауроподоморфа. Массоспондил. Плексообразная (тканая) структура костей предполагала быстрый рост и Дилофозавр могли достигать темпов роста от 30 до 35 кг (от 66 до 77 фунтов) в год в раннем возрасте.[62]
Уэллс обнаружил, что сменные зубы Дилофозавр и другие тероподы возникли глубоко внутри кости, уменьшаясь в размерах по мере удаления от альвеолярного края. В альвеолах обычно было два или три замещающих зуба, самый молодой из которых представлял собой небольшой полый зуб. Корона. Замещающие зубы прорезывались на внешней стороне старых зубов. Когда зуб приблизился к линия десен внутренняя стенка между межзубными пластинами рассосалась и образовала питательную выемку. Когда новый зуб прорезался, он двинулся наружу, чтобы сосредоточиться в альвеолах, и питательная выемка закрылась.[2]
Палеопатология

Уэллс отметил различные палеопатологии (древние признаки болезни, такие как травмы и пороки развития) в Дилофозавр. Голотип имел борозда (борозда или борозда) на нервной дуге шейного позвонка, которая могла возникнуть в результате травмы или раздавливания, и две ямки на правой плечевой кости, которые могли быть абсцессы (коллекции гной ) или артефакты. Уэллс также отметил, что у него левая плечевая кость меньше и более хрупкая, чем правая, но с обратным состоянием предплечий. В 2001 г. палеонтолог Ральф Мольнар предположил, что это было вызвано аномалией развития, называемой колеблющаяся асимметрия. Эта аномалия может быть вызвана стрессом у популяций животных, например, из-за нарушений в их среде, и может указывать на более интенсивную селективное давление. Асимметрия также может быть результатом травмирующих событий на раннем этапе развития животного, которые будут более случайным образом распределены во времени.[2][63] В исследовании 2001 года, проведенном палеонтологом Брюсом Ротшильдом и его коллегами, было изучено 60Дилофозавр кости стопы на предмет признаков стрессовые переломы (которые вызваны напряженными повторяющимися действиями), но не обнаружены. Такие травмы могут быть результатом очень активного хищного образа жизни.[64]
В 2016 году Сентер и Сара Л. Юенгст изучили палеопатологию образца голотипа и обнаружили, что он несет наибольшее и самое разнообразное количество таких заболеваний грудного пояса и передних конечностей среди всех описанных до сих пор тероподных динозавров, некоторые из которых неизвестны. любой другой динозавр. Известно только шесть других теропод с более чем одной палеопатологией на грудном поясе и передних конечностях. У экземпляра голотипа было восемь пораженных костей, тогда как ни у одного другого экземпляра теропод не было больше четырех. С левой стороны у него был перелом лопатки и лучевой кости, а также фибризисы (похожие на абсцессы) в локтевой и внешней сторонах. кость фаланги большого пальца. С правой стороны у него был перекрут плечевого вала, три костные опухоли на его радиусе усеченная суставная поверхность его третьего пястная кость, и деформации на первой фаланге третьего пальца. Этот палец был постоянно деформирован и не мог сгибаться. Деформации плечевой кости и третьего пальца могли быть вызваны остеодисплазия, о котором ранее не сообщалось от нептичьих динозавров, но известно о птицах. У молодых птиц, переживших недоедание, это заболевание может вызывать боль в одной конечности, из-за чего птицы предпочитают использовать другую конечность, что приводит к скручиванию.[65]

Количество травмирующих событий, которые привели к этим особенностям, не определено, и вполне возможно, что все они были вызваны одной встречей, например, в результате врезания в дерево или камень во время драки с другим животным, что могло вызвать колотые раны. когтями. Поскольку все травмы зажили, несомненно, что Дилофозавр выжили после этих событий долгое время, месяцы, возможно, годы. Использование передних конечностей для захвата добычи должно быть нарушено в процессе заживления. Следовательно, динозавр мог выдержать длительный период голодания или питаться добычей, которая была достаточно маленькой, чтобы ее можно было убить ртом и ногами, или одной передней конечностью. По словам Сентера и Юенгста, высокая степень боли, которую динозавр мог испытывать в разных местах в течение длительного времени, также показывает, что это было выносливое животное. Они отметили, что данные о палеопатологии динозавров занижены, и что, хотя Уэллс полностью описал голотип, он упомянул только одну из обнаруженных ими патологий. Они предположили, что такие особенности могут иногда опускаться, потому что описания видов связаны с их характеристиками, а не аномалиями, или потому что такие особенности трудно распознать.[65] Сентер и Салливан обнаружили, что патологии значительно изменили диапазон движений правого плеча и правого третьего пальца голотипа, и поэтому оценки диапазона движений могут не соответствовать оценкам, сделанным для здоровой передней конечности.[57]
Палеоэкология

Дилофозавр известен по свите Кайента, которая датируется Синемурийский и Плиенсбахский этапы ранней юры, примерно 196–183 миллиона лет назад.[66] Кайентская свита является частью Glen Canyon Group это включает образования в северной Аризоне, некоторых частях юго-востока Юты, западного Колорадо и северо-западного Нью-Мексико. Он состоит в основном из двух фации, один доминирует алевролит отложения, а другой - песчаником. Фации алевролитов встречаются на большей части территории Аризоны, а фации песчаников - в районах северной Аризоны, южной Юты, западного Колорадо и северо-западного Нью-Мексико. Формация в основном отлагалась реками, причем фация алевролитов была более медленной и более инертной частью речной системы. Отложение формации Кайента закончилось вторжением поля дюн, которое впоследствии стало Песчаник Навахо.[67] Окончательный радиометрическое датирование этого образования еще не проводилось, а доступная стратиграфическая корреляция была основана на комбинации радиометрических датировок окаменелостей позвоночных, магнитостратиграфия, и доказательства пыльцы.[66] Дилофозавр судя по положению экземпляров в пределах формации Кайента, похоже, сохранились в течение значительного периода времени.[6]
В формации Кайента появилось небольшое, но постоянно растущее скопление организмов. Большинство окаменелостей относится к фации алевролитов.[68] Большинство известных к настоящему времени организмов - позвоночные. К непозвоночным относятся микробный или «водорослевой» известняк,[69] окаменелое дерево,[70] впечатления от растений,[71] пресноводные двустворчатые моллюски и улитки,[67] остракоды,[72] и беспозвоночные следы окаменелостей.[69] Позвоночные животные известны как по окаменелостям тела, так и по ископаемым следам. Позвоночные, известные по окаменелостям тела, включают:[68] гибодонт акулы, неопределенные костлявая рыба, двоякодышащая рыба,[70] саламандры[73] лягушка Просалирус, то цецилий Eocaecilia, черепаха Кайентачелис, а сфенодонт рептилии, ящерицы,[74] и несколько ранних крокодиломорфы включая Calsoyasuchus, Эопневматозух, Kayentasuchus, и Протосух, а птерозавр Рамфинион. Помимо Дилофозавр, известно несколько динозавров, в том числе теропод Мегапнозавр,[14] и Кайентавенатор,[75] завроподоморф Саразавр,[76] а гетеродонтозаврид, а тиреофор Скутеллозавр. Синапсиды включить тритилодонтиды Диннебитодон, Кайентатерий, и Олигокифус, морганукодонтиды,[74] возможное раннее настоящее млекопитающее Диннетерий, а харамиид млекопитающее. Большинство этих находок происходит из окрестностей Голд-Спринг, штат Аризона.[68] Ископаемые остатки позвоночных включают: копролиты и следы терапсиды, подобные ящерицам животные и несколько видов динозавров.[69][77]
Тафономия
Уэллс обрисовал тафономия оригинальных образцов, изменения, которые произошли во время их распада и окаменения. Скелет голотипа был найден лежащим на правом боку, а его голова и шея были изогнуты - загнуты назад - в "поза смерти ", в которой часто встречаются скелеты динозавров. Считалось, что эта поза опистотонус (из-за смертельных спазмов) в то время, но, возможно, вместо этого это было результатом того, как туша была встроена в отложения. Спина была прямой, а самые задние спинные позвонки повернуты на левую сторону. Хвостовые позвонки неравномерно отходили от таза, а ноги были сочлененными, с небольшим смещением. Уэллс пришел к выводу, что образцы были похоронены на месте их смерти, но их мало транспортировали, но что образец голотипа, похоже, был потревожен падальщиками, на что указывают повернутые спинные позвонки и раздробленный череп.[2][78] Гей отметил, что экземпляры, которые он описал в 2001 году, свидетельствуют о том, что они были перенесены ручьем. Поскольку ни один из образцов не был готов, они могли быть перенесены на некоторое расстояние или лежали на поверхности и некоторое время подвергались атмосферным воздействиям перед транспортировкой. Они могли быть перевезены наводнение, на что указывает разнообразие животных, обнаруженных в виде фрагментов и переломов костей.[12]
Культурное значение

Дилофозавр был показан в романе 1990 года парк Юрского периода, автор Майкл Крайтон, а его 1993 экранизация режиссером Стивен Спилберг. В Дилофозавр из парк Юрского периода был признан "единственным серьезным отклонением от научной достоверности" в фильме изготовление книга, и как «самый беллетризованный» из динозавров фильма в книге о Stan Winston Studios, который создал аниматроники последствия. Для романа Крайтон изобрел способность динозавра плевать ядом (объясняя, как он может убивать добычу, несмотря на его, казалось бы, слабые челюсти). Художественный отдел добавил еще одну функцию, капот сложенная на его шее, которая расширялась и вибрировала, когда животное готовилось атаковать, подобно шее ящерица с оборками. Во избежание путаницы с Велоцираптор как показано в фильме, Дилофозавр был представлен как всего 1,2 метра (4 фута) в высоту вместо предполагаемой истинной высоты около 3,0 метра (10 футов). По прозвищу «плевательница», Дилофозавр Фильм был реализован через кукловодство и требовал полного тела с тремя сменными головами для выполнения действий, требуемых сценарием. Отдельные ноги были также сконструированы для кадра, на котором прыгает динозавр. В отличие от большинства других динозавров в фильме, нет компьютерные изображения был использован при показе Дилофозавр.[79][80][81]
Геолог Дж. Брет Беннингтон в 1996 г. отметил, что, хотя Дилофозавр вероятно, не имел оборок и не мог плевать ядом, как в фильме, его укус мог быть ядовитым, как утверждается в отношении Дракон Комодо. Он обнаружил, что добавление яда к динозавру не менее допустимо, чем придание цвета его коже, который также неизвестен. Если бы у динозавра была оборка, доказательства этого были бы в костях, в форме жесткой конструкции, удерживающей оборку, или отметинах в местах прикрепления мышц, используемых для ее движения. Он также добавил, что если бы у него была оборка, он бы использовал его не для устрашения еды, а скорее для конкурента (он предположил, что он мог отреагировать на персонажа фильма, натягивающего капюшон на голову).[82] В обзоре 1997 г. на книгу о науке парк Юрского периода, палеонтолог Питер Додсон также указал на неправильный масштаб фильма Дилофозавр, а также невероятность его яда и оборки.[83] В 2014 году Баккер отметил, что фильм Дилофозавр не хватало заметной выемки на верхней челюсти, и пришел к выводу, что создатели фильма хорошо поработали над созданием устрашающего химера различных животных, но предупредил, что его нельзя использовать для обучения реальным животным.[84] Сам Уэллс был «взволнован», увидев Дилофозавр в парк Юрского периода: Он отметил неточности, но нашел в них мелкие детали, получил удовольствие от фильма и был счастлив найти динозавра «всемирно известным актером».[85]

В соответствии с Миф навахо, туши убитых монстров были «вбиты в землю», но стереть с лица земли было невозможно, а окаменелости традиционно интерпретировались как их останки. Хотя народ навахо помогал палеонтологам находить окаменелости с 19 века, традиционные верования предполагают, что призраки монстров остаются в своих частично погребенных трупах и должны содержаться там с помощью действенных ритуалов. Точно так же некоторые опасаются, что кости их родственников будут выкопаны вместе с останками динозавров, и что удаление окаменелостей свидетельствует о неуважении к прошлым жизням этих существ.[86] В 2005 году историк Эдриенн Майор заявил, что Уэллс отметил, что во время первоначальных раскопок Дилофозавр, Навахо Уильямс исчез из раскопок через несколько дней, и предположил, что это произошло потому, что Уильямс счел детальную работу с тонкими кистями «ниже своего достоинства». Вместо этого мэр указал, что люди навахо действительно занимаются детальной работой, такой как украшения и живопись, и что объяснение ухода Уильямса могло быть вместо этого традиционным беспокойством, поскольку скелеты появились и были потревожены. Мэр также указал на инцидент в 1940-х годах, когда мужчина навахо помогал раскопать Пентацератопс скелет, при условии, что ему не приходилось касаться костей, но покинул это место, когда их покрыло всего несколько дюймов грязи.[86] В книге 1994 года Уэллс сказал, что Уильямс вернулся через несколько дней с двумя женщинами навахо, которые сказали: «Это не мужская работа, это работа скво».[9]
Скалы в Аризоне, содержащие кости Дилофозавр Также есть петроглифы к предки пуэблоанцев вырезанные на них, и пересекающиеся дорожки местности называются Naasho’illbahitsho Biikee навахо, что означает «следы большой ящерицы». По словам мэра, навахо проводили церемонии и делали подношения этим следам монстров. Треугольные треки также использовались в качестве украшений костюмов и наскальное искусство из Хопи и Зуни, вероятно, под влиянием таких следов динозавров.[86] В 2017 г. Дилофозавр был обозначен как государственный динозавр штата США Коннектикут, чтобы стать официальным с новым государственным бюджетом в 2019 году. Дилофозавр был выбран потому, что следы, предположительно оставленные похожими динозаврами, были обнаружены в Rocky Hill в 1966 г. при раскопках Межгосударственное шоссе 91. Шесть треков были назначены ихно-виды Eubrontes giganteus, который был сделан государственное ископаемое Коннектикута в 1991 году. Район, в котором они были обнаружены, был озером триасового периода, и, когда значимость этого района была подтверждена, шоссе было изменено, и район стал государственный парк названный Государственный парк динозавров. В 1981 г. скульптура Дилофозаврпервая реконструкция этого динозавра в натуральную величину была подарена парку.[23][87][88]
Дилофозавр был предложен в качестве государственного динозавра Аризоны 9-летним мальчиком в 1998 году, но законодатели предложили Соноразавр вместо этого, утверждая, что Дилофозавр не был уникальным для Аризоны. Был предложен компромисс, который распознал бы обоих динозавров, но счет умер, когда выяснилось, что Дилофозавр окаменелости были взяты без разрешения резервации навахо, и потому что они больше не проживали в Аризоне. Впоследствии официальные лица навахо обсудили, как вернуть окаменелости.[89][90] По словам мэра, один навахо заявил, что они больше не просят вернуть окаменелости, но поинтересовался, почему не были сделаны слепки, чтобы кости можно было оставить, поскольку было бы лучше оставить их в земле, и построили музей. чтобы люди могли прийти посмотреть на них там.[86] 11-летний мальчик снова предложил Соноразавр как динозавр штата Аризона в 2018 году.[90]
Рекомендации
- ^ «Дилофозавр». Оксфордские словари Британский словарь. Oxford University Press. Получено 21 января, 2016.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа Уэллс, С. (1984). "Дилофозавр ветхерилли (Dinosauria, Theropoda), остеология и сравнения ». Palaeontographica Abteilung A. 185: 85–180.
- ^ а б c Уэллс, С. (1954). «Новый юрский динозавр из формации Кайента в Аризоне». Бюллетень Геологического общества Америки. 65 (6): 591–598. Bibcode:1954GSAB ... 65..591Вт. Дои:10.1130 / 0016-7606 (1954) 65 [591: NJDFTK] 2.0.CO; 2.
- ^ Уэллс, С.; Гуралник, Р.П. (1994). "Дилофозавр обнаруженный". ucmp.berkeley.edu. Калифорнийский университет в Беркли. В архиве с оригинала 8 ноября 2017 г.. Получено 13 февраля, 2018.
- ^ а б c d е ж Найш, Д. (2009). Великие открытия динозавров. Лондон, Великобритания: A&C Black Publishers Ltd., стр. 94–95. ISBN 978-1-4081-1906-8.
- ^ а б c d е ж грамм час я j k л м п Marsh, A.D .; Роу, Т. (2020). «Комплексная анатомическая и филогенетическая оценка Дилофозавр ветхерилли (Dinosauria, Theropoda) с описаниями новых образцов из формации Кайента в северной Аризоне ». Журнал палеонтологии. 94 (S78): 1–103. Дои:10.1017 / jpa.2020.14. S2CID 220601744.
- ^ а б c d е ж Уэллс, С. (1970). "Дилофозавр (Reptilia: Saurischia), новое название динозавра ». Журнал палеонтологии. 44 (5): 989. JSTOR 1302738.
- ^ а б Уэллс, С.; Гуралник, Р.П. (1994). "Подробности о дилофозавре". ucmp.berkeley.edu. Калифорнийский университет в Беркли. В архиве с оригинала 2 августа 2017 г.. Получено 13 февраля, 2018.
- ^ а б Psihoyos, L .; Кнеббер, Дж. (1994). Охота на динозавров. Лондон, Великобритания: Касселл. С. 86–89. ISBN 978-0679431244.
- ^ а б Раухут, О. (2004). «Взаимоотношения и эволюция базальных динозавров-теропод». Специальные статьи по палеонтологии. 69: 213.
- ^ а б Глют, Д.Ф. (1997). Динозавры: Энциклопедия. Джефферсон: McFarland & Company, Inc., стр. 347–350. ISBN 978-0786472222.
- ^ а б c d Гей, Р. (2001). Новые образцы Дилофозавр ветхерилли (Dinosauria: Theropoda) из раннеюрской формации Кайента в северной Аризоне. Ежегодное собрание Западной ассоциации палеонтологов позвоночных. 1. Меса, Аризона. п. 1.
- ^ а б c d Carrano, M.T .; Benson, R.B.J .; Сэмпсон, С. (2012). «Филогения столбняка (Dinosauria: Theropoda)». Журнал систематической палеонтологии. 10 (2): 211–300. Дои:10.1080/14772019.2011.630927. S2CID 85354215.
- ^ а б c Тыкоски, Р. (2005). Анатомия, онтогенез и филогения целофизоидных теропод (Тезис). Техасский университет. С. 1–232 - через библиотеки UT: Электронные диссертации и диссертации.
- ^ Пикрелл, Дж. (7 июля 2020 г.). "парк Юрского периода почти все неправильно понял об этом культовом динозавре ". Национальная география. Наука. Получено 12 июля, 2020.
- ^ а б Гей, Р. (2005). "Доказательства полового диморфизма у тероподных динозавров ранней юры, Дилофозавр и сравнение с другими родственными формами ». В Carpenter, K. (ed.). Плотоядные динозавры. Блумингтон, Индиана: Издательство Индианского университета. С. 277–283. ISBN 978-0-253-34539-4.
- ^ Мортимер, М. (май 2010 г.). "Таксоны Пикеринга 6: Дилофозавр порода ". Theropoddatabase.blogspot.com. В архиве с оригинала от 29 декабря 2017 г.. Получено 29 декабря, 2017.
- ^ Hendrickx, C .; Mateus, O .; Эванс, Алистер Роберт (2014). "Torvosaurus gurneyi п. sp., крупнейшего наземного хищника из Европы, и предлагаемую терминологию анатомии верхней челюсти у нептичьих теропод ". PLOS ONE. 9 (3): e88905. Bibcode:2014PLoSO ... 988905H. Дои:10.1371 / journal.pone.0088905. ЧВК 3943790. PMID 24598585.
- ^ Ху, С. (1993). "Краткий отчет о возникновении Дилофозавр из округа Цзиньнин провинции Юньнань ". Позвоночные PalAsiatica. 1 (на китайском языке) (1-е изд.). 31: 65–69.
- ^ а б Lamanna, M.C .; Holtz, T.R., Jr .; Додсон, П. (1998). "Переоценка китайского динозавра теропод Дилофозавр китайский". Журнал палеонтологии позвоночных. Тезисы докладов, пятьдесят восьмое ежегодное собрание Общества палеонтологии позвоночных. 18 (3): 57–58. JSTOR 4523942.
- ^ Xing, L .; Bell, P.R .; Rothschild, B.M .; Ran, H .; Zhang, J .; Dong, Z .; Zhang, W .; Карри, П.Дж. (2013). «Потеря зубов и ремоделирование альвеол в Sinosaurus triassicus (Dinosauria: Theropoda) из нижнеюрских слоев бассейна Луфэн, Китай ». Китайский научный бюллетень. 58 (16): 1931. Bibcode:2013ЧСБУ..58.1931Х. Дои:10.1007 / s11434-013-5765-7.
- ^ Ван, Го-Фу; Ты, Хай-Лу; Пан, Ши-Ганг; Ван, Тао (2017). «Новый динозавр-теропод из ранней юры провинции Юньнань, Китай». Позвоночные PalAsiatica. 55 (2): 177–186.
- ^ а б c d е ж грамм Пол, Г.С. (1988). Хищные динозавры мира. Нью-Йорк, Нью-Йорк: Саймон и Шустер. стр.258, 267–271. ISBN 978-0-671-61946-6.
- ^ Хольц, Т. Р., младший (2012). Динозавры: наиболее полная и современная энциклопедия для любителей динозавров всех возрастов. Нью-Йорк, Нью-Йорк: Random House. п.81. ISBN 978-0-375-82419-7.
- ^ Пол, Г.С. (2010). Принстонский полевой путеводитель по динозаврам. Принстон, Нью-Джерси: Издательство Принстонского университета. п.75. ISBN 978-0-691-13720-9.
- ^ а б Герлинский, Г. (1996). «Перьевидные отпечатки на остатках теропод из нижней юры Массачусетса». Бюллетень Музея Северной Аризоны. 60: 179–184.
- ^ а б Кундрат, М. (2004). «Когда тероподы стали пернатыми? - Свидетельства до-Археоптерикс перистые придатки ». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития. 302B (4): 355–364. Дои:10.1002 / jez.b.20014. PMID 15287100.
- ^ а б Локли, М .; Мацукава, М .; Цзяньцзюнь, Л. (2003). «Крадущиеся тероподы в таксономических джунглях: технологические и ихнотаксономические исследования следов с плюсневыми и седалищными вдавлениями». Ичнос. 10 (2–4): 169–177. Дои:10.1080/10420940390256249. S2CID 128759174.
- ^ а б Martin, A.J .; Рейнфорт, E.C. (2004). «След покоя тероподов, который также является следом передвижения: тематическое исследование образца Хичкока AC 1/7». Геологическое общество Америки. Рефераты с программами. 36 (2): 96. Архивировано с оригинал 31 мая 2004 г.
- ^ а б c d е ж грамм Tykoski, R.S .; Роу, Т. (2004). «Цератозаврия». In Weishampel, D.B .; Dodson, P .; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли, Калифорния: Калифорнийский университет Press. С. 47–70. ISBN 978-0-520-24209-8.
- ^ Уэллс, С. (1983). «Два центра окостенения у теропод астрагала». Журнал палеонтологии. 57 (2): 401. JSTOR 1304663.
- ^ Уэллс, С.; Лонг, Р.А. (1974). "Плюсна тероподных динозавров" [Летопись Южноафриканского музея]. Музей Annale van die Suid-Afrikaanse. 64: 191–218. ISSN 0303-2515.
- ^ Хольц, Т. Р., младший (1994). «Филогенетическое положение Tyrannosauridae: значение для систематики теропод». Журнал палеонтологии. 68 (5): 1100–1117. Дои:10.1017 / S0022336000026706. JSTOR 1306180.
- ^ Madsen, J.H .; Уэллс, С. (2000). "Ceratosaurus (Dinosauria, Theropoda): пересмотренная остеология". Геологическая служба Юты: 1–89. 41293.
- ^ а б Йейтс, А. (2005). «Новый динозавр-теропод из ранней юры Южной Африки и его значение для ранней эволюции теропод». Palaeontologia Africana. 41: 105–122. ISSN 0078-8554.
- ^ Smith, N.D .; Makovicky, P.J .; Hammer, W.R .; Карри, П.Дж. (2007). «Остеология Криолофозавр эллиоти (Dinosauria: Theropoda) из ранней юры Антарктиды и последствия для ранней эволюции теропод ». Зоологический журнал Линнеевского общества. 151 (2): 377–421. Дои:10.1111 / j.1096-3642.2007.00325.x.
- ^ Hendrickx, C .; Hartman, S.A .; Матеус, О. (2015). «Обзор открытий и классификации нептичьих теропод». Журнал PalArch по палеонтологии позвоночных. 12 (1): 73.
- ^ Zahner, M .; Бринкманн, В. (2019). "Теропод триасового периода авеространской линии из Швейцарии и ранняя эволюция динозавров". Природа Экология и эволюция. 3 (8): 1146–1152. Дои:10.1038 / s41559-019-0941-z. ЧВК 6669044. PMID 31285577.
- ^ а б Уэллс, С. (1971). «Следы динозавров из формации Кайента в северной Аризоне». Плато. 44: 27–38.
- ^ Morales, M .; Балкли, С. (1996). "Палеоихнологические свидетельства существования динозавра теропод крупнее, чем Дилофозавр в нижнеюрской кайентинской свите. Континентальная юра ». Бюллетень Музея Северной Аризоны. 60: 143–145.
- ^ Герлинский, Г. (1991). «Новый ихнотакса динозавра из ранней юры гор Святого Креста, Польша». Палеогеография, палеоклиматология, палеоэкология. 85 (1–2): 137–148. Bibcode:1991ППП .... 85..137Г. Дои:10.1016 / 0031-0182 (91) 90030-У.
- ^ Gierliński, G .; Альберг, А. (1994). «Следы динозавров позднего триаса и раннего юрского периода в формации Хёганас на юге Швеции». Ичнос. 3 (2): 99. Дои:10.1080/10420949409386377.
- ^ Weems, R.E. (2003). "Платеозавр Конструкция стопы предполагает, что для Eubrontes и Гигандипус следы ". In le Tourneau, P.M .; Olsen, P.E. (ред.). Великие рифтовые долины Пангеи в восточной части Северной Америки. 2. Нью-Йорк: издательство Колумбийского университета. стр.293 –313. ISBN 978-0231126762.
- ^ Рейнфорт, E.C. (2003). «Пересмотр и переоценка раннеюрского ихногена динозавров» Отозум". Палеонтология. 46 (4): 803–838. Дои:10.1111/1475-4983.00320.
- ^ а б Weems, R.E. (2006). "Манус печати Kayentapus minor; его влияние на биомеханику и ихнотаксономию раннемезозойских заурисховых динозавров ". Бюллетень Музея естественной истории и науки Нью-Мексико. 37: 369–378.
- ^ Lucas, S.G .; Klein, H .; Lockley, M.G .; Spielmann, J.A .; Герлински, Г.Д .; Хант, А. П .; Таннер, Л.Х. (2006). «Триасово-юрское стратиграфическое распределение тероподного следа ихногена. Eubrontes". Бюллетень Музея естественной истории и науки Нью-Мексико. 37. 265.
- ^ а б c Милнер, Эндрю Р.К .; Harris, J.D .; Lockley, M.G .; Kirkland, J.I .; Matthews, N.A .; Харпендинг, Х. (2009). "Анатомия, поза и поведение птиц, обнаруженные по следам покоя тероподных динозавров ранней юры". PLOS ONE. 4 (3): e4591. Bibcode:2009PLoSO ... 4,4591 млн. Дои:10.1371 / journal.pone.0004591. ЧВК 2645690. PMID 19259260.
- ^ Weems, R.E. (2019). "Доказательства существования двуногих прозауроподов как вероятного Eubrontes гусеницы ". Ичнос. 26 (3): 187–215. Дои:10.1080/10420940.2018.1532902. S2CID 133770251.
- ^ Баккер, Р. (1986). Ереси динозавров. Нью-Йорк, Нью-Йорк: Уильям Морроу. стр.263 –264. ISBN 978-0-8217-5608-9.
- ^ Therrien, F .; Хендерсон, Д .; Руфф, К. (2005). «Укуси меня - биомеханические модели челюстей теропод и их влияние на пищевое поведение». В Карпентер, К. (ред.). Плотоядные динозавры. Издательство Индианского университета. С. 179–230. ISBN 978-0-253-34539-4.
- ^ Milner, A .; Киркланд, Дж. (2007). «Дело о ловле динозавров на территории открытия динозавров в Сент-Джордж на ферме Джонсон» (PDF). Обзорные заметки Геологической службы штата Юта. 39: 1–3.
- ^ Marsh, A.D .; Роу, Т. (2018). «Анатомия и систематика завроподоморфа. Сарасавр золотистый (Sarahsaurus aurifontanalis) из раннеюрской кайентской свиты ». PLOS ONE. 13 (10): e0204007. Bibcode:2018PLoSO..1304007M. Дои:10.1371 / journal.pone.0204007. ЧВК 6179219. PMID 30304035.
- ^ а б Czerkas, S.J .; Черкас, С.А. (1990). Динозавры: взгляд со всего мира. Лимпсфилд: мир драконов. п. 208. ISBN 978-0-7924-5606-3.
- ^ Пол, Г.С., изд. (2000). «Цветной раздел». Научная американская книга динозавров. Нью-Йорк, Нью-Йорк: Пресса Св. Мартина. п. 216. ISBN 978-0-312-31008-0.
- ^ Senter, P .; Робинс, Дж. (2005). "Диапазон движений передней конечности тероподового динозавра Acrocanthosaurus atokensis, и последствия для хищнического поведения ". Журнал зоологии. 266 (3): 307–318. Дои:10.1017 / S0952836905006989.
- ^ Senter, P .; Робинс, Дж. (2015). «Ориентация лопаток и передних конечностей динозавров в покое: численный анализ с применением для реконструкций и музейных креплений». PLOS ONE. 10 (12): e0144036. Bibcode:2015PLoSO..1044036S. Дои:10.1371 / journal.pone.0144036. ЧВК 4684415. PMID 26675035.
- ^ а б c d Senter, P .; Салливан, К. (2019). "Передние конечности динозавра теропод Дилофозавр ветхерилли: Диапазон движений, влияние палеопатологии и мягких тканей, а также описание дистального отдела запястной кости ». Palaeontologia Electronica. Дои:10.26879/900.
- ^ Кумбс, В. (1990). «Образцы поведения динозавров». In Weishampel, D.B .; Osmolska, H .; Додсон, П. (ред.). Динозаврия (1-е изд.). Беркли, Калифорния: Калифорнийский университет Press. п. 42. ISBN 978-0-520-06727-1.
- ^ Padian, K .; Хорнер, Дж. Р. (2011). «Эволюция« причудливых структур »у динозавров: биомеханика, половой отбор, социальный отбор или распознавание видов?». Журнал зоологии. 283 (1): 3–17. Дои:10.1111 / j.1469-7998.2010.00719.x.
- ^ Knell, R.J .; Сэмпсон, С. (2011). «Причудливые структуры у динозавров: распознавание видов или половой отбор? Ответ Падиану и Хорнеру» (PDF). Журнал зоологии. 283 (1): 18–22. Дои:10.1111 / j.1469-7998.2010.00758.x.
- ^ Hone, D.W.E .; Найш, Д. (2013). «« Гипотеза распознавания видов »не объясняет наличие и эволюцию преувеличенных структур у нептичьих динозавров». Журнал зоологии. 290 (3): 172–180. Дои:10.1111 / jzo.12035.
- ^ Ткач, Ю. (19 октября 1996 г.). Многоэлементный остеогистологический анализ Дильфозавр ветхерилли. Пятьдесят шестое ежегодное собрание Общества палеонтологии позвоночных. Журнал палеонтологии позвоночных. Тезисы докладов. 16 (3). Американский музей естественной истории, Нью-Йорк, Нью-Йорк. стр. 1A – 80A. JSTOR 4523755.
- ^ Мольнар, Р. (2001). «Палеопатология теропод: обзор литературы». In Tanke, D.H .; Карпентер, К. (ред.). Мезозойская жизнь позвоночных. Блумингтон, Индиана: Издательство Индианского университета. стр.337 –363. ISBN 978-0-253-33907-2.
- ^ Ротшильд, В .; Tanke, D.H .; Форд, Т. (2001). «Стресс-переломы теропод и отрывы сухожилий как ключ к активности». In Tanke, D.H .; Карпентер, К. (ред.). Мезозойская жизнь позвоночных. Блумингтон, Индиана: Издательство Индианского университета. стр.331 –336. ISBN 978-0253339072.
- ^ а б Senter, P .; Juengst, S.L .; Хейманн, Д. (2016). «Рекордная боль: самое большое количество и разнообразие заболеваний костей передних конечностей у динозавров-теропод». PLOS ONE. 11 (2): e0149140. Bibcode:2016PLoSO..1149140S. Дои:10.1371 / journal.pone.0149140. ЧВК 4765892. PMID 26909701.
- ^ а б Дж. М., Кларк; Фастовский, Д. (1986). «Биостратиграфия позвоночных группы Глен-Каньон в северной Аризоне». Во Фрейзере, Северная Каролина; Сьюз, Х.-Д. (ред.). Начало эпохи динозавров: изменение фауны на границе триаса и юры. Кембридж, Великобритания: Издательство Кембриджского университета. С. 285–301. ISBN 978-0-521-36779-0.
- ^ а б Harshbarger, J. W .; Repenning, C.A .; Ирвин, Дж. Х. (1957). «Стратиграфия верхов триасовых и юрских отложений страны навахо». Геологическое общество Нью-Мексико. 291: 98–114.
- ^ а б c Lucas, S.G .; Heckert, A.B .; Таннер, Л. Х. (2005). «Юрские окаменелые позвоночные в Аризоне и возраст группы Глен-Каньон». Бюллетени Музея естественной истории Нью-Мексико. 29: 95–104.
- ^ а б c Luttrell, P.R .; Моралес, М. (1993). «Преодоление разрыва в Моенкопи-Уош: литостратиграфическая корреляция». Бюллетень Музея Северной Аризоны. 59: 111–127.
- ^ а б Jenkins, F.A .; Шубин, Н. Х. (1998). "Просалирус битис и анурановый каудопазовый механизм ". Журнал палеонтологии позвоночных. 18 (3): 495–510. Дои:10.1080/02724634.1998.10011077.
- ^ Rigby, J.K .; Hamblin, W.K .; Matheny, R .; Валлийский, С.Л. (1971). "Путеводитель по реке Колорадо: Часть 3, Моав и Хайт, Юта через национальный парк Каньонлендс" (PDF). Исследования Университета Бригама Янга, серия по геологии. 18 (2): 7.
- ^ Lucas, S.G .; Таннер, Л. Х. (2007). «Биостратиграфия и биохронология тетрапод триасово-юрского перехода на южном плато Колорадо, США». Палеогеография, палеоклиматология, палеоэкология. 244 (1–4): 242–256. Bibcode:2007ППП ... 244..242л. Дои:10.1016 / j.palaeo.2006.06.030.
- ^ Curtis, K .; Падиан, К. (1999). «Раннеюрская фауна микропозвоночных из формации Кайента на северо-востоке Аризоны: изменение микрофауны на границе триаса и юры». Палеобиос. 19 (2): 19–37.
- ^ а б Jenkins, F.A .; Crompton, A.W .; Даунс, В. Р. (1983). «Мезозойские млекопитающие из Аризоны: новые свидетельства эволюции млекопитающих». Наука. 222 (4629): 1233–1235. Bibcode:1983Sci ... 222.1233J. Дои:10.1126 / science.222.4629.1233. PMID 17806725. S2CID 35607107.
- ^ Гей, Р. (2010). "Kayentavenator elysiae, новый тетануран из ранней юры Аризоны ». Гей, Р. (ред.). Заметки о тероподах раннего мезозоя. Лулу Пресс. С. 27–43. ISBN 978-0557466160.
- ^ Rowe, T.B .; Сьюз, Х.-Д .; Рейс, Р.Р. (2010). «Распространение и разнообразие первых североамериканских динозавров зауроподоморфа с описанием нового таксона». Труды Королевского общества B: биологические науки. 278 (1708): 1044–1053. Дои:10.1098 / rspb.2010.1867. ЧВК 3049036. PMID 20926438.
- ^ Hamblin, A.H .; Фостер, Дж. Р. (2000). «Древние следы животных и следы в Национальном памятнике Гранд-Лестница - Эскаланте, юг центральной части штата Юта». In Sprinkel, D.A .; Chidsey, T.C., Jr .; Андерсон, П. (ред.). Геология парков и памятников Юты. Солт-Лейк-Сити: Геологическая ассоциация Юты. С. 557–568. ISBN 978-1882054107.
- ^ Reisdorf, A.G .; Вуттке, М. (2012). "Переоценка гипотезы опистотонической осанки Муди у ископаемых позвоночных, Часть I: Рептилии - тафономия двуногих динозавров. Compsognathus longipes и Юравенатор старки с архипелага Зольнхофен (юрский период, Германия) ». Палеобиоразнообразие и палеоокружение. 92 (1): 119–168. Дои:10.1007 / s12549-011-0068-у. S2CID 129785393.
- ^ Крайтон, М. (1990). парк Юрского периода. Лондон: Random Century Group. стр.76–78. ISBN 978-0-394-58816-2.
- ^ Shay, D .; Дункан, Дж. (1993). Создание Парка Юрского периода. Нью-Йорк: Boxtree Ltd., стр. 24, 35–36, 113. ISBN 978-1-85283-774-7.
- ^ Дункан, Дж. (2007). Эффект Уинстона: искусство и история студии Стэна Уинстона. Лондон, Великобритания: Titan Books. С. 177–178. ISBN 978-1-84576-150-9.
- ^ Беннингтон, Дж. Б. (1996). "Ошибки в фильме парк Юрского периода". Американский палеонтолог. 4 (2): 4–7. Архивировано из оригинал 2 октября 2018 г.. Получено 28 декабря, 2017.
- ^ Додсон, П. (1997). «Воскрешение мертвых». Наука. 277 (5326): 644–645. Дои:10.1126 / science.277.5326.644b. JSTOR 2893254. S2CID 178195275.
- ^ Баккер, Р. (2014). "Сказка двух компов: Что парк Юрского периода понял - и ошибся - в анатомии динозавров ". blog.hmns.org. Хьюстонский музей естествознания. Получено 18 августа, 2020.
- ^ Уэллс, С.; Гуралник, Р.П. (1994). "Дилофозавр, актер". ucmp.berkeley.edu. Калифорнийский университет в Беркли. В архиве с оригинала 4 января 2018 г.. Получено 13 февраля, 2018.
- ^ а б c d Мэр А. (2005). «Юго-Запад: ископаемые фетиши и убийцы монстров». Ископаемые легенды первых американцев. Принстон, Нью-Джерси: Издательство Принстонского университета. С. 130–143. ISBN 978-0691130491. JSTOR j.ctt4cgcs9.10.
- ^ Стоун, М. (2017). «Коннектикут приветствует своего нового государственного динозавра». CTBoom.com. Получено 13 февраля, 2018.
- ^ Роджерс, О. (2016). «Обнаруженные следы динозавров изменяют маршрут шоссе и ведут в государственный парк». История Коннектикута. Получено 3 января, 2018.
- ^ Мозер, К. (26 апреля 1998 г.). «Споры о динозаврах в Аризоне становятся настоящей Годзиллой». Республика Аризона. Deseret News. Получено 12 мая, 2020.
- ^ а б Геберс, С. (19 февраля 2018 г.). "В Аризоне нет официального государственного динозавра, поэтому одиннадцатилетний мальчик предложил одного - Соноразавр". Республика. Получено 20 февраля, 2018 - через AZ Central.
внешняя ссылка
- "Дилофозавр!". Рассказанная выставка "Экскурсия". рассказанный Сэмюэл П. Уэллс.CS1 maint: другие (связь)
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||













