Арковенатор - Arcovenator
| Арковенатор | |
|---|---|
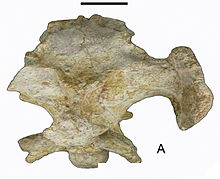 | |
| Мозговая оболочка (MHNAix-PV 2011-12) в дорсальной проекции | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Семья: | †Abelisauridae |
| Подсемейство: | †Majungasaurinae |
| Род: | †Арковенатор Тортоса и другие.., 2013 |
| Разновидность: | †A. escotae |
| Биномиальное имя | |
| †Arcovenator escotae Тортоса и другие.., 2013 | |
Арковенатор («Arc hunter») - это род из Абелизаврид теропод динозавры родом из Поздний мел из Франция.[1] В тип и только описал разновидность является Arcovenator escotae.[1]
Описание

Хотя более мелкий, почти полный черепная коробка из Арковенатор в остальном похож по размеру на Маджунгазавр и Карнотавр; таким образом, первоначально она была оценена как длина около 5–6 м (16–20 футов),[1] но, по оценкам, в 2016 году его длина составила 4,8 м (16 футов).[2] В том же году Молина-Перес и Ларраменди оценили рост в 7,2 м (23,6 фута) в длину и 950 кг (2094 фунта).[3] В крыша черепа экспонаты как уникальный диагностический признак средняя линия отверстие, возможно, размещение шишковидная железа, расположенный на задней поверхности небольшого купола, образованного лобные кости такой же средней толщины, как в Ауказавр, поэтому меньше, чем для Раджасавр, хотя больше, чем у Rugops.[1] Менее характерно, выше орбита низкий ямка с небольшим фенестра граничит с слезный, лобная и заглазничная.[1] В теменный граничащие с надвисочными отверстиями, образуют гребни медиально на соответствующих переднемедиальных краях последнего, которые по мере приближения к теменное возвышение, слиться в сагиттальный гребень.[1] В заглазничный занимает промежуточное положение между плезиоморфный Т-образное состояние Эоабелизавр и производная инвертированная L-образный один из Карнотавр из-за уникальной особенности наличия листа кости, соединяющего его вентральный и задний отростки.[1] У него похожим аутапоморфным образом имеется толстый отросток с шероховатой поверхностью дорсальнее глазницы, который простирается до слезной железы, образуя костный отросток. надбровная дуга и менее заметно - латеральный морщинистый бугорок на конце его вентрального отростка.[1] Затылочные отростки имеют замечательные дополнительные дорсальные и вентральные костные перемычки, которые, таким образом, ограничивают углубления латерально большое затылочное отверстие.[1] В область уха очень похож на Маджунгазавр, хотя наиболее существенно различаются латерально направленным базиптеригоидным отростком, с более короткой crista prootica и меньшей протяженностью борозды впереди 2-й и 3-й черепной нерв foramina являются незначительными отклонениями от типа Majungasaurinae.[1] В чешуйчатый похож на последний, за исключением менее заметного теменного отростка.[1] Как правило, внешний костный орнамент более приглушенный, чем у Маджунгазавр.[1] Высокие зубы (3-5,5 см) имеют зубчики на апикальной части зуба. мезиальный киля и по длине дистального с разной плотностью.[1]
В каудальный позвонки из A. escotae удивительно похожи на таковые из Маджунгазавр, хотя более сжато дорсовентрально.[1] Центры обладают амфицелозными сочленениями с соответствующими гранями промежуточного характера между круговыми. Илокелесия и эллиптической формы в Раджасавр и не имеют ни пневматических выемок, ни принадлежностей гипосфен-гипантрум суставы.[1] В поперечные отростки из нервные дуги не так склонны, как в Брахиростра.[1]
В синемиальный гребень из Арковенатор 's the стройный 51-см большеберцовая кость хорошо развит, что характерно для абелизавроиды.[1] Имеет проксимальный боковой мыщелок более заметный, чем медиальный один, небольшой антеродорсальный изгиб на проксимальной стороне гребня малоберцовой кости, заметный дистальный продольный гребень и конусообразный лодыжки.[1] Почти полметра длиной малоберцовая кость обладает типичными анатомическими особенностями кератозавры.[1]
Классификация и систематика
Арковенатор род теропод, гнездящийся внутри клады Abelisauridae,[1] который в Линнеевская таксономия имеет ранг семья.[4] Эта таксономическая группа имеет как близких родственников ноазавриды в пределах Абелизавройда.[1][5] Последний в свою очередь вместе с Лимузавр и Цератозавр гнезда внутри Цератозаврия.[1][6]
Отличительными чертами абелизаврид являются их короткие высокие черепа с сильно рельефными внешними поверхностями, резко уменьшенные передние конечности и крепкие задние конечности.[7]

Тьерри Тортоса и его коллеги провели филогенетический анализ, который кратко изложен в кладограмма ниже и частично основывается на ранее опубликованных работах, включая как недавно обнаруженные ископаемые останки, так и другие описанные, но неназванные французские абелизавры.[1]
| Majungasaurinae |
| ||||||||||||||||||
Исследование в целом согласуется с предыдущими результатами, а именно с относительно недавним результатом, полученным как Мэтью Каррано, так и Скоттом Сэмпсоном (2008).[8] и Диего Пол и Оливер В. М. Раухут (2012)[6] клады, которая включает не менее Маджунгазавр, Индозавр и Раджасавр, который в более позднем анализе включает Арковенатор.[1] Тортоса и другие.. назовите эту хорошо поддерживаемую кладу Majungasaurinae, оценивая его как подсемейство и определив, что он содержит всех абелизавридов, более тесно связанных с Маджунгазавр чем Карнотавр.[1] Члены этой таксономической группы имеют различные общие черты черепа, включая удлиненное анторбитальное отверстие и теменное с сагиттальным гребнем, расширяющимся кпереди в треугольную поверхность.[1] Также следует отметить, что в частичном согласии с некоторыми анализами более фрагментарные останки французских цератозавров помещены в Abelisauridae, и в отличие от других, Абелизавр восстанавливается как карнотаурин.[1]
Кроме того, понимание палеобиогеография абелизавроидов существуют; просто наличие их на так называемом Европейском архипелаге[9] противоречит гипотезам, которые рассматривают только континенты, полученные из расставаться из Мезозойский Гондвана.[1] Выделяются две линии европейских абелизавров: базальная, включая малую Альбианский Родозавр и ниже Кампанский Тараскозавр, а производный - больший кампанский Арковенатор в союзе с Мадагасканец Маджунгазавр и Индийский Раджасавр в Majungasaurinae.[1] Поскольку предполагаемое распределение признаков, полученное с помощью филогенетического анализа, делает маловероятным, что эти линии более тесно связаны друг с другом, чем с другими абелизавридами, это предполагает более сложную серию событий, касающихся их биогеографии с викарианс применимо к старому и океаническому рассредоточение более вероятен более поздний.[1] Эти результаты подтверждают предполагаемую роль Африки как центра перемещений фауны между Европа и Индия или Мадагаскар[10] и изоляция южноамериканских абелизаврид.[1][8]
Открытие и наименование
Ископаемые останки A. escotae были найдены рядом Pourrières, Вар отделение, Прованс-Альпы-Лазурный берег область, край,[11] во время профилактических палеонтологических и археологических изысканий до начала строительства на участке Автомагистраль A8 между Шатонеф-ле-Руж и Святой Максимин.[1][12][13] Уместное позднее Кампанский страты (от 72 до 76 миллион лет назад )[14][15][16] нижнего Формация Аргилес Рутилантес расположены в Экс-ан-Прованс Бассейн юго-востока Франция.[1] В голотип из Arcovenator escotae, расположенный в Muséum d’Histoire Naturelle d'Aix-en-Provence, был обнаружен тесно связанным в единственном слое речного песчаника и состоит из образцов MHNA-PV-2011.12.1, мозговой коробки в сочленении с правой заглазничной частью, MHNA-PV-2011.12.2, левой чешуйчатой кости, MHNA.PV .2011.12.15, зуб, MHNA.PV.2011.12.5, MHNA.PV.2011.12.5, передний хвостовой позвонок, MHNA.PV.2011.12.3, правая большеберцовая кость, и MHNA.PV.2011.12.4, правая малоберцовая кость.[1] Найдено два передних хвостовых позвонка (MHNA.PV.2011.12.198 и MHNA.PV.2011.12.213) и три зуба (MHNA.PV.2011.12.20, MHNA.PV.2011.12.187 и MHNA.PV.2011.12.297). близкие как по расстоянию, так и по глубине также относились к виду, но принадлежали разным особям.[1] Вполне вероятно, что верхняя челюсть, единственное ископаемое, найденное среди так называемых Pourcieux abelisaurid, по крайней мере, относится к этому роду как из-за его непосредственной близости во времени и пространстве, так и из-за результатов филогенетического анализа.[1][8]
Название рода Арковенатор происходит из реки Дуга так как населенный пункт установлен в пределах своего бассейн и латинский слово для охотника, венатор.[1] В специфический эпитет 'escotae' награды Escota, а автомагистраль уступка Компания,[17] который с 2006 года предоставил необходимые средства для раскопок местности.[1]
Палеоэкология
A. escotae жил на Иберо-Армориканском острове,[1] относительно большой массив суши, образованный тем, что сейчас является частью Франции, Испания, и Португалия.[9] В компрессионный проседание бассейн Экс-ан-Прованса имел низкий рельеф эндорейский дело, расположенное в палеоширота 35 ° с.ш. и граничил с севером и югом в виде известняк нагорье соответственно Sainte Victoire и Etoile массивы, а на восток - горы Мор.[14] Осадки из этих источников текли по рекам в многолетние растения. озеро происходящие с переслаиванием озерных, аллювиальных и речных отложений во время Арковенатор, когда климат был теплым, полувлажным с ярко выраженными временами года.[14] Ископаемые останки были найдены в одном из формирование различные уровни речного песчаник,[1] характеристика устье реки или когда это выходит из берегов,[14] вместе с гибодонты, то черепахи Foxemys и Solemys, то крокодиломорфы Musturzabalsuchus и Ищирочампа, аждархид птерозавры, титанозавр зауроподы, то орнитопод Рабдодон и нодозавриды.[1] Обилие фрагментарных останков абелизавров среднего размера, особенно зубов, в этом и других местах региона показывает, что эти животные были относительно обычными в ландшафте.[1]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах ай эй ак аль являюсь ан ао ap Тортоса, Тьерри; Эрик Баффето; Николя Виалле; Ив Дютур; Эрик Турини; Жиль Шейлан (2013). «Новый динозавр абелизаврид из позднего мела на юге Франции: палеобиогеографические последствия». Анналы палеонтологии. 100 (В печати): 63–86. Дои:10.1016 / j.annpal.2013.10.003.
- ^ Grillo, O. N .; Делькур, Р. (2016). «Аллометрия и длина тела абелизавроидных теропод: Pycnonemosaurus nevesi новый король ". Меловые исследования. 69: 71–89. Дои:10.1016 / j.cretres.2016.09.001.
- ^ Молина-Перес и Ларраменди (2016). Records y curiosidades de los dinosaurios Terópodos y otros dinosauromorfos. Барселона, Испания: Ларусс. п. 256. ISBN 9780565094973.
- ^ Краузе, Дэвид В .; Сэмпсон, Скотт Д.; Каррано, Мэтью Т .; О'Коннор, Патрик М. (2007). "Обзор истории открытия, таксономии, филогении и биогеографии Majungasaurus crenatissimus (Theropoda: Abelisauridae) из позднего мела Мадагаскара ". In Sampson, Scott D .; Krause, David W. (eds.). Majungasaurus crenatissimus (Theropoda: Abelisauridae) из позднего мела Мадагаскара. Мемуары Общества палеонтологии позвоночных 8. 27. С. 1–20. Дои:10.1671 / 0272-4634 (2007) 27 [1: OOTHOD] 2.0.CO; 2.
- ^ Серено, Пол К.; Wilson, JA; Конрад, Дж. Л. (2007). «Новые динозавры связывают южные массивы суши в среднем меловом периоде». Труды Королевского общества B. 271 (1546): 1325–1330. Дои:10.1098 / rspb.2004.2692. ЧВК 1691741. PMID 15306329.
- ^ а б Диего Пол и Оливер В. М. Раухут (2012). «Среднеюрский абелизаврид из Патагонии и раннее разнообразие динозавров-теропод». Труды Королевского общества B: биологические науки. 279 (1804): 3170–5. Дои:10.1098 / rspb.2012.0660. ЧВК 3385738. PMID 22628475.
- ^ Тыкоски, Рональд Б.; Роу, Тимоти (2004). «Цератозаврия». В Weishampel, Дэвид Б.; Додсон, Питер; Osmólska, Halszka (ред.). Динозаврия (Второе изд.). Беркли: Калифорнийский университет Press. С. 47–70. ISBN 0-520-24209-2.
- ^ а б c Carrano, M.T .; S.D. Сэмпсон (2008). «Филогения Ceratosauria (Dinosauria: Theropoda)». Журнал систематической палеонтологии. 6 (2): 183–236. Дои:10.1017 / с1477201907002246. S2CID 30068953.
- ^ а б Далла Веккья, Фабио М. (2006). "Телматозавр и другие гадрозавриды мелового европейского архипелага. Обзор" (PDF). Натура Наскоста (32): 1–55. Архивировано из оригинал (PDF) в 2013-08-29. Получено 2013-12-14.
- ^ Ezcurra, MD; F.L. Аньолин (2012). «Абелизавроидный динозавр из средней юры Лавразии и его влияние на палеобиогеографию и эволюцию теропод». Труды ассоциации геологов. 123 (3): 500–507. Дои:10.1016 / j.pgeola.2011.12.003.
- ^ "Insee - COG - Fiche de la commune de Pourrières". Insee. Получено 13 декабря 2013.
- ^ "VINCI Autoroutes | Quand l'autoroute mène aux dinosaures…" (На французском). VINCI Autoroutes. 4 июня 2012 г.. Получено 13 декабря 2013.
- ^ "VINCI Autoroutes | L'autoroute du temps sur l'A8" (На французском). VINCI Autoroutes. Получено 13 декабря 2013.
- ^ а б c d Кожан, Изабель; Мари-Габриэль Моро (2006). «Корреляция земных климатических колебаний с глобальными сигналами в верхнемеловом и датском периодах в условиях сжатия (Прованс, Франция)». Журнал осадочных исследований. 76 (3): 589–604. Bibcode:2006JSedR..76..589C. Дои:10.2110 / jsr.2006.045.
- ^ Ogg, J.G .; Хиннов Л.А. (2012). «27: Меловой период». У Феликса М. Градштейна; Джеймс Г. Огг; Марк Д. Шмитц; Габи М. Огг (ред.). Шкала геологического времени 2012. п. 810. Дои:10.1016 / B978-0-444-59425-9.00027-5. ISBN 9780444594259.
- ^ Тибо, Николас; Дороти Хассон; Рикке Харлоу; Сильвия Гардин; Бруно Гальбрун; Эмилия Хюре; Фабрис Минолетти (15 июня 2012 г.). «Астрономическая калибровка событий изотопов углерода верхнего кампана – маастрихта и биостратиграфия известкового планктона в Индийском океане (отверстие 762C ODP): влияние на возраст границы кампан – маастрихт». Палеогеография, палеоклиматология, палеоэкология. 337–338 (52–71): 52–71. Bibcode:2012ППП ... 337 ... 52 т. Дои:10.1016 / j.palaeo.2012.03.027.
- ^ "VINCI Autoroutes | Escota". VINCI Autoroutes. Получено 13 декабря 2013.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||













