Аутапоморфия - Autapomorphy
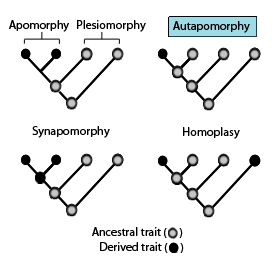
В филогенетика, аутапоморфия отличительная особенность, известная как полученный признак, уникальный для данного таксона. То есть встречается только в одном таксон, но не встречается ни в каких других или аутгруппа таксоны, даже те, которые наиболее тесно связаны с фокальным таксоном (который может быть разновидность, семья или вообще любую кладу).[2] Поэтому его можно считать апоморфия по отношению к одному таксону.[3] Слово аутапоморфия, впервые представленный в 1950 году немецким энтомолог Вилли Хенниг, получается из Греческий слова αὐτός, авто- = "сам"; ἀπό, апо = "далеко от"; и μορφή, морфа = "форма".[4]
Обсуждение
Поскольку аутапоморфии присутствуют только в одном таксоне, они не несут информацию о родстве. Следовательно, аутапоморфии бесполезны для определения филогенетических отношений. Однако аутапоморфия, как и синапоморфия и плезиоморфия - понятие относительное, зависящее от рассматриваемого таксона. Аутапоморфия на данном уровне вполне может быть синапоморфией на менее инклюзивном уровне.[5] Пример аутапоморфии можно описать у современных змей. Змеи лишились двух пар ног, которые характерны для всех Тетрапода, а ближайшие к Офидия - как и их общие предки - у всех по две пары ног. Следовательно, таксон Ophidia представляет собой аутапоморфию в отношении отсутствия ног.[3]
В концепция аутапоморфных видов это один из многих методов, которые ученые могут использовать для определения и различения разновидность от другого. Это определение присваивает виды на основе степени дивергенции, связанной с репродуктивной несовместимостью, которая измеряется в основном числом аутапоморфий.[6] Этот метод группировки часто называют "монофилетический Концепция видов »или концепция« филосовидов », популяризированная Д.Е. Розеном в 1979 году. В рамках этого определения вид рассматривается как« наименее инклюзивная монофилетическая группа, определяемая по крайней мере одной аутапоморфией ».[7] Хотя эта модель видообразования полезна тем, что позволяет избежать немонофилетических группировок, она также имеет свои критические замечания. Н.И. Платник, например, считает концепцию аутапоморфных видов неадекватной, поскольку она допускает возможность репродуктивной изоляции и видообразования при отмене «видового» статуса материнской популяции. Другими словами, если периферическая популяция отделяется и становится репродуктивно изолированной, вероятно, потребуется развить по крайней мере одну аутапоморфию, чтобы ее можно было распознать как другой вид. Если это может произойти без большей материнской популяции также развивая новую аутапоморфию, тогда материнская популяция не может оставаться видом в рамках концепции аутапоморфных видов: у нее больше не будет никаких апоморфий, которые также не разделялись бы дочерними видами.[8]
Филогенетические сходства: Эти филогенетические термины используются для описания различных паттернов наследственных и производных признаков или состояний признаков, как указано на приведенной выше диаграмме в связи с синапоморфиями.[1]
- Гомоплазия в биологическая систематика это когда черта был получен или потерян независимо в отдельных линиях в ходе эволюции. Этот конвергентная эволюция приводит к тому, что виды, независимо друг от друга, обладают признаком, который отличается от признака, который, как предполагается, имел место у их общего предка.[9][10][11]
- Параллельный Гомоплазия - производный признак, присутствующий в двух группах или видах без общего предка из-за конвергентная эволюция.[12]
- Обеспечить регресс Гомоплазия - признак, присутствующий у предка, но не у прямых потомков, который снова появляется у более поздних потомков.[13]
- Апоморфия - производная черта. Апоморфия, разделяемая двумя или более таксонами и унаследованная от общего предка, называется синапоморфией. Апоморфия, уникальная для данного таксона, называется аутапоморфией.[14][15][16][17]
- Синапоморфия /Гомология - производный признак, который встречается в некоторых или всех терминальных группах клады и унаследован от общего предка, для которого он был аутапоморфным (т.е. не присутствует в это непосредственный предок).
- Основная синапоморфия - синапоморфия, которая снова была утеряна у многих членов клады. Если он потеряется во всех, кроме одного, его будет трудно отличить от аутапоморфии.
- Аутапоморфия - отличительный производный признак, который является уникальным для данного таксона или группы.[3]
- Симплезиоморфия - наследственный признак, общий для двух или более таксонов.
- Плезиоморфия - симплезиоморфия, обсуждаемая в отношении более производного состояния.
- Псевдоплезиоморфия - это черта, которая не может быть идентифицирована ни как плезиоморфия, ни как апоморфия, являющаяся разворотом.[18]
- Реверсирование - это потеря производного признака, присутствующего в предке, и восстановление плезиоморфного признака.
- Конвергенция - независимая эволюция сходного признака у двух или более таксонов.
- Гемиплазия[19][20]
Рекомендации
- ^ а б Пейдж РД, Холмс ЕС (14 июля 2009 г.). Молекулярная эволюция: филогенетический подход. Джон Вили и сыновья. ISBN 978-1-4443-1336-9. OCLC 609843839.
- ^ Футуйма DJ (1998). Эволюционная биология (3-е изд.). Sinauer Associates, Inc. стр. 95.
- ^ а б c Аппель Р.Д., Фейтманс Э. (2009). «Глава 3: Введение в филогенетику и ее молекулярные аспекты». Биоинформатика: взгляд из Швейцарии (1-е изд.). Всемирная научная издательская компания.
- ^ Calow PP (2009). Энциклопедия экологии и природопользования. Джон Вили и сыновья. ISBN 978-1-4443-1324-6. OCLC 1039167559.
- ^ Forey PL (1997). История рыб латимерии (1-е изд.). Спринтер.
- ^ Говард DJ, Берлохер SH (1998). Бесконечные формы: виды и видообразование (1-е изд.). США: Издательство Оксфордского университета. ISBN 978-0-19-510901-6. OCLC 60181901.
- ^ Бык АТ (2004). Микробное разнообразие и биоразведка. ASM Press.
- ^ Платник Н.И. (2001). "От кладограмм к классификациям: путь к дефилокоду." (PDF). Ассоциация систематиков.
- ^ Gauger A (17 апреля 2012 г.). «Сходство бывает! Проблема гомоплазии». Evolution Today и новости науки.
- ^ Сандерсон М.Дж., Хаффорд Л. (21 октября 1996 г.). Гомоплазия: повторение сходства в эволюции. Эльзевир. ISBN 978-0-08-053411-4. OCLC 173520205.
- ^ Брэндли М.С., Уоррен Д.Л., Лич А.Д., Макгуайр Дж. А. (апрель 2009 г.). «Гомоплазия и кладовая поддержка». Систематическая биология. 58 (2): 184–98. Дои:10.1093 / sysbio / syp019. PMID 20525577.
- ^ Арчи JW (сентябрь 1989 г.). «Коэффициенты избытка гомоплазии: новые индексы для измерения уровней гомоплазии в филогенетической систематике и критика индекса согласованности». Систематическая зоология. 38 (3): 253–269. Дои:10.2307/2992286. JSTOR 2992286.
- ^ Wake DB, Wake MH, Specht CD (февраль 2011 г.). «Гомоплазия: от выявления закономерностей к определению процесса и механизма эволюции». Наука. 331 (6020): 1032–5. Дои:10.1126 / science.1188545. PMID 21350170. S2CID 26845473. Сложить резюме – Science Daily.
- ^ Simpson MG (9 августа 2011 г.). Систематика растений. Эльзевир. Амстердам: Эльзевир. ISBN 9780080514048.
- ^ Рассел П.Дж., Герц П.Е., Макмиллан Б. (2013). Биология: динамическая наука. Cengage Learning. ISBN 978-1-285-41534-5.
- ^ Липскомб Д. (1998). «Основы кладистического анализа» (PDF). Вашингтон, округ Колумбия: Университет Джорджа Вашингтона.
- ^ Чоудхури С (9 мая 2014 г.). Биоинформатика для начинающих: гены, геномы, молекулярная эволюция, базы данных и аналитические инструменты (1-е изд.). Академическая пресса. п. 51. ISBN 978-0-12-410471-6. OCLC 950546876.
- ^ Уильямс Д., Шмитт М., Уиллер К. (21.07.2016). Будущее филогенетической систематики: наследие Вилли Хеннига. ISBN 978-1-107-11764-8. OCLC 951563305.
- ^ Avise JC, Робинсон TJ (июнь 2008 г.). «Гемиплазия: новый термин в лексиконе филогенетики». Систематическая биология. 57 (3): 503–7. Дои:10.1080/10635150802164587. PMID 18570042.
- ^ Копетти Д., Беркес А., Бустаманте Е., Шарбоно Дж. Л., Чайлдс К. Л., Эгиарте Л. Е. и др. (Ноябрь 2017 г.). «Обширные несоответствия между генами и гемиплазия сформировали геномы североамериканских столбчатых кактусов». Труды Национальной академии наук Соединенных Штатов Америки. 114 (45): 12003–12008. Дои:10.1073 / pnas.1706367114. ЧВК 5692538. PMID 29078296.
| Соответствующие поля | ||
|---|---|---|
| Базовые концепты | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||
| ||