Эволюционный класс - Evolutionary grade
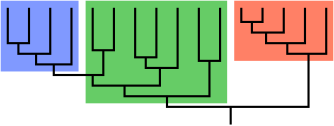
В альфа-таксономия, а оценка таксон, объединенный уровнем морфологический или физиологический сложность. Термин был придуман британским биологом. Джулиан Хаксли, чтобы контрастировать с клады, строго филогенетический Блок.[1]
Определение
Эволюционная ступень - это группа видов, объединенных морфологический или физиологический черты, которые дали начало другой группе, которая имеет серьезные отличия от наследственного состояния и, таким образом, не считается частью наследственной группы, но все же имеет достаточно сходства, чтобы мы могли сгруппировать их в одну кладу. Наследственная группа не будет филогенетически законченной (т.е.не будет формировать клады ), поэтому будет представлять парафилетический таксон.
Чтобы полностью понять эволюционные ступени, нужно сначала лучше понять Филогенетика, определяется как "В биология, это исследование эволюционный история и отношения между людьми или группами организмы (например. виды, или население ). Эти отношения открываются через филогенетический вывод методы оценки наблюдаемых наследственный черты характера, такие как ДНК последовательности или морфология в рамках модели эволюции этих черт »
Чаще всего приводят пример рептилии. В начале 19 века французский натуралист Латрей был первым, кто разделил четвероногие на четыре знакомых класса амфибий, рептилий, птиц и млекопитающих.[2] В этой системе рептилии характеризуются такими чертами, как откладывание перепончатых или очищенных яиц, наличие кожи, покрытой напольные весы или щитки, и имея 'хладнокровный метаболизм. Однако предки млекопитающих и птиц также обладали этими чертами, и поэтому можно сказать, что птицы и млекопитающие «произошли от рептилий», что делает рептилий, если их определять по этим признакам, скорее классом, чем кладой.[3] В микробиология, таксоны, которые, таким образом, рассматриваются как исключенные из родительской группы эволюционного уровня, называются замаскированные таксоны.[4]
Парафилетические таксоны часто, но не всегда, представляют собой эволюционные ступени. В некоторых случаях парафилетические таксоны объединяются просто потому, что не являются частью каких-либо других групп, и дают начало так называемым таксоны корзины для мусора который может даже быть полифилетический.
Оценки по систематике

Традиционный Линнеевский способ определения таксонов - использование анатомических признаков. Когда фактическое филогенетическое родство неизвестно, иногда оказывается, что четко определенные группы определяются признаками, которые являются примитивными, а не производными. По Линнею систематика, эволюционные оценки принимаются в высшие таксономические ранги, хотя обычно избегают семья уровень и ниже. В филогенетическая номенклатура эволюционные оценки (или любая другая форма парафилии) не принимаются.[5]
Если доступна информация о филогенетических отношениях, организмы преимущественно сгруппированы в клады. В случае отсутствия данных или необходимости сравнения групп с неопределенной взаимосвязью, кладистический метод ограничен, а оценка является полезным инструментом для сравнения организмов. Это особенно часто встречается в палеонтология, где окаменелости часто фрагментарны и трудны для интерпретации. Таким образом, традиционные палеонтологические работы часто используют эволюционные степени как формальные или неформальные таксоны, включая такие примеры, как лабиринтодонты, анапсиды, синапсиды, динозавры, аммониты, эвриптериды, лобоподийцы и многие из наиболее известных таксонов эволюция человека. Организация организмов по классам, а не строгим классам, также может быть очень полезной для понимания эволюционной последовательности, лежащей в основе большого разнообразия обоих животных.[6] и растения.[7]
Эволюционные степени, объединенные общими морфологическими признаками, часто в высшей степени узнаваемы в этой области. В то время как таксономия стремится исключить парафилетические таксоны, такие степени иногда сохраняются как формальные или неформальные группы на основе их полезности для неспециалистов и полевых исследователей.[5] В бактериология, переименование видов или групп, которые оказываются эволюционными степенями, сведено к минимуму, чтобы избежать недоразумений, что в случае патогены может иметь фатальные последствия. Когда речь идет о группе организмов, термин «сорт» обычно заключен в кавычки, чтобы обозначить его статус как парафилетического термина.
Сорта и филогенетическая номенклатура
С ростом филогенетическая номенклатура, использование эволюционных оценок как формальных таксоны стал предметом обсуждения. При строгом филогенетическом подходе только монофилетический таксоны признаны.[8] Это отличается от более традиционного подхода эволюционная таксономия.[9] Различие в подходах привело к ожесточенным дебатам между сторонниками двух подходов к таксономии, особенно в таких хорошо известных областях, как палеонтология позвоночных и ботаника.[10] Однако разница между утверждением «B является частью A» (филогенетический подход) и «B произошел от A» (эволюционный подход) является одним из семантика а не филогении. Оба выражают одну и ту же филогению, но первая подчеркивает филогенетический континуум, а вторая подчеркивает отчетливый сдвиг в анатомии или экологии в B по сравнению с A.
Примеры
- Рыбы представляют собой степень, поскольку они дали начало наземные позвоночные. В свою очередь, три традиционных классы рыбы (Агната, Chondrichthyes и Osteichthyes ) все представляют собой эволюционные ступени.[11]
- Амфибии в биологическом смысле (включая вымершие Лабиринтодонты ) представляют собой степень, так как они также являются предками амниот.[11]
- Рептилии представляют собой сорт, состоящий из хладнокровных амниот; это исключает птицы и млекопитающие.[3]
- Динозавры были предложены предками птиц еще в 1860-х годах.[12] Тем не менее, этот термин широко используется как эволюционная ступень, за исключением птиц, хотя большинство ученых используют монофилетическую динозаврию.
- Ящерицы представляют собой ступень эволюции, определяемую удержанием конечностей относительно змеи и амфисбайцы.[13] Однако определять ящериц по наличию конечностей неверно, так как существует много видов безногие ящерицы, которые считаются настоящими ящерицами.
- Зеленые водоросли представляют собой степень, поскольку они являются предками наземные растения.
- Прокариоты, к которым относятся клеточные организмы без ядра, представляют собой класс, поскольку они являются предками эукариоты, который включает в себя животные, растения, грибы, и протисты; и последняя из этих четырех групп также представляет собой оценку, так как она исключает предыдущие три группы.
- Ракообразные представляют собой степень, поскольку они являются предками гексаподы, который включает в себя насекомые и родственные таксоны.
- Обезьяны представляют собой степень, так как они включают предков обезьяны (в том числе люди ).
- Точно так же обезьяны представляют собой класс в обычном употреблении, но, если к ним относятся люди, являются кладой.
использованная литература
- ^ Хаксли Дж. 1959. Клады и сорта. В Cain A.J. (ред) Функция и таксономическое значение. Ассоциация систематиков, Лондон.
- ^ Латрей, П.А. (1804). Nouveau Dictionnaire à Histoire Naturelle, xxiv; цитируется по Latreille, P.A. (1825 г.).Familles naturelles du règne animal, exposés succinctement et dans un ordre analytique.
- ^ а б Тадж, Колин (2000). Разнообразие жизни. Издательство Оксфордского университета. ISBN 0198604262.
- ^ Lan, R; Ривз, PR (2002). «Замаскированная Escherichia coli: молекулярное происхождение шигелл». Микробы и инфекции / Institut Pasteur. 4 (11): 1125–32. Дои:10.1016 / S1286-4579 (02) 01637-4. PMID 12361912.
- ^ а б Грант, Верн (1998). «Первичная классификация и филогения Polemoniaceae с комментариями по молекулярной кладистике». Американский журнал ботаники. 85 (6): 741–752. Дои:10.2307/2446408. JSTOR 2446408. PMID 21684957.
- ^ Sperling, E. A .; Pisani, D .; Петерсон, К. Дж. (1 января 2007 г.). «Пориферская парафилия и ее значение для докембрийской палеобиологии» (PDF). Геологическое общество, Лондон, Специальные публикации. 286 (1): 355–368. Bibcode:2007GSLSP.286..355S. Дои:10.1144 / SP286.25. S2CID 34175521. Архивировано из оригинал (PDF) 9 мая 2009 г.. Получено 22 августа 2012.
- ^ Донохью, Майкл Дж. (1 июня 2005 г.). «Ключевые инновации, конвергенция и успех: макроэволюционные уроки филогении растений» (PDF). Палеобиология. 31 (SP5): 77–93. Дои:10.1666 / 0094-8373 (2005) 031 [0077: KICASM] 2.0.CO; 2.
- ^ Кевин де Кейруш и Жак Готье (1990). «Филогения как центральный принцип в таксономии: филогенетические определения названий таксонов». Syst. Zool. 39 (4): 307–322. Дои:10.2307/2992353. JSTOR 2992353.
- ^ Майр, Эрнст И Бок, W.J. (2002). «Классификации и другие системы заказа» (PDF). J. Zool. Syst. Evol. Исследование. 40 (4): 169–94. Дои:10.1046 / j.1439-0469.2002.00211.x.
- ^ Бентон, М. Дж. (2000). «Стебли, узлы, коронные клады и списки без рангов: Линней мертв?». Биологические обзоры. 75 (4): 633–648. CiteSeerX 10.1.1.573.4518. Дои:10.1111 / j.1469-185X.2000.tb00055.x. ISSN 0006-3231. PMID 11117201. S2CID 17851383. Архивировано из оригинал 5 июня 2011 г.
- ^ а б Ромер, А.С.. И Т.С. Парсонс. 1977 г. Тело позвоночного. 5-е изд. Сондерс, Филадельфия. (6-е изд. 1985 г.)
- ^ Хаксли, Томас Х. (1870). «Еще одно доказательство близости динозавров рептилий и птиц». Ежеквартальный журнал Лондонского геологического общества. 26 (1–2): 12–31. Дои:10.1144 / GSL.JGS.1870.026.01-02.08. S2CID 129125487.
- ^ Гиббонс, Дж. Уитфилд; Гиббонс, Уит (1983). Их кровь холодеет: приключения с рептилиями и амфибиями. Алабама: Университет Алабамы Нажмите. стр.164. ISBN 978-0-8173-0135-4.
| Соответствующие поля | ||
|---|---|---|
| Базовые концепты | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||
| ||