Целофиз - Coelophysis
| Целофиз | |
|---|---|
 | |
| Установлен скелет на Кливлендский музей естественной истории | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Семья: | †Coelophysidae |
| Род: | †Целофиз Справиться, 1889 |
| Типовой вид | |
| †Целофиз баури Коп, 1887 г. | |
| Разновидность | |
| |
| Синонимы | |
Синонимия рода
Синонимия видов (К. Баури)
| |
Целофиз (/sɛˈлɒжɪsɪs/ seh-LOF-ih-sis традиционно; /ˌsɛлoʊˈжаɪsɪs/ SEL-о-FY-sis или же /ˌsялoʊˈжаɪsɪs/ ВИДЕТЬ-ло-FY-sis, о чем чаще говорят в последние десятилетия[5]) является вымерший род из целофизид теропод динозавр которые жили примерно от 216 до 196 миллион лет назад во второй половине Триасовый Период на юго-западе США[6] а также в Южной Африке и Зимбабве.[7]
Целофиз был маленьким, стройным, наземным жилищем, двуногий плотоядное животное, который мог вырасти до 3 м (9,8 футов) в длину. Это один из самых ранних известных динозавр роды. Разрозненный материал, представляющий похожих животных, был обнаружен во всем мире в некоторых Поздний триас и Раннеюрский период образования.
В типовой вид К. Баури, первоначально переданный роду Целурус к Эдвард Дринкер Коуп в 1887 г., был описан последним в 1889 г. Имена Лонгозавр и Риоаррибазавр являются синонимами Целофиз. Другой род динозавров, Мегапнозавр, также считается синонимом. Этот примитивный теропод известен тем, что является одним из самых богатых образцами родов динозавров.
Описание

Целофиз известен из ряда полных ископаемых скелетов этого вида К. Баури, который был динозавром легкого телосложения длиной до 3 метров (9,8 футов).[8] и которая была выше метра в бедрах. Пол (1988) оценил вес изящной формы в 15 кг (33 фунта), а вес устойчивой формы в 20 кг (44 фунта).[9] Целофиз был двуногим плотоядным динозавром-теропод, быстрым и ловким бегуном.[10] Несмотря на то, что они были ранним динозавром, эволюция теропод форма тела уже значительно продвинулись по сравнению с такими существами, как Herrerasaurus и Эораптор. Торс Целофиз соответствует основной форме тела теропод, но грудной пояс отображает некоторые интересные особенности: К. Баури имел фуркула (поперечный рычаг), самый ранний известный пример динозавра. Целофиз также сохраняет наследственное состояние обладания четырьмя цифрами на руке (манус). У него было всего три функциональных пальца, четвертый врезался в ладонь.[11]
Целофиз имел узкие бедра, приспособленные для захвата добычи передние конечности и узкие лапы.[12] Его шея и хвост были длинными и тонкими.[6] В таз и задние конечности К. Баури также небольшие вариации в плане тела теропод. Он имеет открытый вертлужная впадина и прямой шарнир на щиколотке, которые определяют Dinosauria. Задняя конечность оканчивалась трехпалой ступней (пес ), с повышенным Hallux. Хвост имел необычную структуру внутри переплетения. презигапофиз позвонков, которые образовывали полужесткую решетку, по-видимому, чтобы не дать хвосту двигаться вверх и вниз.[13]
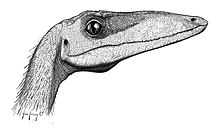
Целофиз имел длинную узкую голову (примерно 270 мм (0,9 фута)) с большими, обращенными вперед глазами, что позволяло стереоскопический зрение и, как следствие, отличное восприятие глубины. Райнхарт и другие. (2004) описали полную склеротическое кольцо найдено для несовершеннолетнего Целофиз баури (образец NMMNH P-4200), и сравнил его с данными по склеротические кольца рептилий и птиц и пришел к выводу, что Целофиз был дневной, зрительно ориентированный хищник.[14] Исследование показало, что видение Целофиз превосходил зрение большинства ящериц и приравнивался к зрению современных хищных птиц. Глаза Целофиз кажутся наиболее близкими к орлам и ястребам, с высокой способностью к аккомодации. Данные также свидетельствуют о плохом ночном зрении, что означает, что у этого динозавра был круглый зрачок, а не разделенный.[10]
Целофиз имел удлиненную морду с большим фенестры что помогло снизить вес черепа, а узкие костные перемычки сохранили структурную целостность черепа. Шея имела ярко выраженный сигмовидный изгиб. Головной мозг известен в Целофиз баури но получить мало данных, потому что череп был раздавлен.[12] В отличие от некоторых других теропод, черепные украшения Целофиз не располагался наверху черепа. На дорсолатеральном крае носовой и слезной костей черепа, прямо над костями, располагались низкие, выступающие с боков костные гребни. анторбитальное окно.[15]
Отличительные анатомические особенности
Диагноз - это изложение анатомических особенностей организма (или группы), которые в совокупности отличают его от всех других организмов. Некоторые, но не все особенности диагноза также являются аутапоморфиями. Аутапоморфия - это отличительная анатомическая особенность, уникальная для данного организма или группы.
Согласно Эскурре (2007) и Бристоу и Раат (2004) Целофиз Отличить можно по следующим признакам: отсутствие смещения рострального отростка верхней челюсти; квадратная кость сильно каудально; небольшое наружное отверстие нижней челюсти, составляющее 9–10% длины нижней челюсти;[16] а переднезадняя длина вентрального слезного отростка превышает 30% его высоты.[17]
Некоторые палеонтологи считают Целофиз баури быть таким же динозавром, как Целофиз родезийский (ранее Syntarsus, поочередно Мегапнозавр), однако это было опровергнуто следующим: Даунс (2000) пришел к выводу, что К. Баури отличается от C. rhodesiensis по длине шейки матки, проксимальным и дистальным пропорциям задних конечностей и анатомии проксимального каудального отдела позвоночника;[18] Тыкоски и Роу (2004) пришли к выводу, что К. Баури отличается от C. rhodesiensis в том, что у него нет ямки у основания носового отростка предчелюстной кости;[12] и Бристоу и Раат (2004) пришли к выводу, что К. Баури отличается от C. rhodesiensis в наличии более длинного зубного ряда на верхней челюсти.[17]
История открытия

Названный в 1887 г. типовой вид Целофиз первоначально был назван как вид Целурус.[19] Эдвард Дринкер Коуп первый названный Целофиз в 1889 г., чтобы назвать новый род вне Целурус и Танистрофей которому К. Баури ранее был отнесен к категории К. Баури, C. willistoni и С. longicollis.[20] Любитель ископаемое Коллекционер Дэвид Болдуин, работавший на Копа, нашел первые останки динозавра в 1881 году в формации Чинл на северо-западе Нью-Мексико.[21] В начале 1887 года Коп отнес собранные образцы к двум новым видам: К. Баури и С. longicollis рода Целурус. Позже, в 1887 году, Коуп переназначил материал еще одному роду, Танистрофей. Двумя годами позже Коуп исправил свою классификацию, осознав различия в позвонках, и назвал Целофиз, с К. Баури как типовой вид,[20] который был назван в честь Георг Баур, сравнительный анатом, идеи которого были аналогичны Коупу.[21][22] Название Целофиз исходит из Греческий слова κοῖλος / koilos (что означает «полый») и φύσις / physis (что означает «форма»), таким образом, «полая форма», которая является ссылкой на полые позвонки.[20][23] Однако эти первые находки были слишком плохо сохранились, чтобы дать полное представление об этом новом динозавре. В 1947 году значительное "кладбище" Целофиз окаменелости были найдены Джорджем Уитакером, помощником Эдвина Х. Колберта, в Нью-Мексико, на Призрачное ранчо, близко к оригинальной находке. Американский музей естественной истории, палеонтолог Эдвин Х. Колберт провели комплексное исследование[15] всех окаменелостей, найденных до этой даты, и отнесены к Целофиз. Экземпляры Ghost Ranch были настолько многочисленны, в том числе многие хорошо сохранившиеся и полностью сочлененные экземпляры, что один из них с тех пор стал диагностическим, или типовой образец, для всего рода, заменяя исходный, плохо сохранившийся экземпляр.[24]

"Syntarsus" rhodesiensis был впервые описан Раатом (1969) и отнесен к Podokesauridae.[25] Таксон «Podokesauridae» был заброшен, так как его типовой экземпляр был уничтожен во время пожара и его уже нельзя сравнивать с новыми находками. На протяжении многих лет палеонтологи относили этот род к Ceratosauridae (Welles, 1984), Procompsognathidae (Parrish, Carpenter, 1986) и Ceratosauria (Gauthier, 1986). В 2004 году "Syntarsus" был признан синонимом Целофиз Тыкоски и Роу (2004). Эскурра и Новас (2007) и Эскурра (2007) также пришли к выводу, что «Синтарсус» был синонимом Целофиз, что является текущим научным консенсусом.[26]
Классификация

|
Целофиз является отдельной таксономической единицей (род ), состоящий из двух видов; К. Баури и C. rhodesiensis (последние ранее относились к роду Мегапнозавр).[1] Два дополнительных первоначально описанных вида, С. longicollis и C. willistoni, теперь считаются синонимом К. Баури. C. rhodesiensis вероятно является частью этого родового комплекса и известен из юрского периода на юге Африки.[1] Третий возможный вид - это Целофиз кайентакатае, ранее относившиеся к роду Мегапнозавр.[28][29] На данный момент нет четкого консенсуса.
В начале 1990-х годов велись споры о диагностических характеристиках первых собранных образцов по сравнению с материалом, раскопанным на ранчо Призраков. Целофиз карьер. Некоторые палеонтологи придерживались мнения, что оригинальные образцы не являлись диагностическими, кроме них самих, и поэтому название К. Баури не может применяться ни к каким дополнительным образцам. Поэтому они применили другое имя, Риоаррибазавр, к образцам из карьера Ghost Ranch.[30]
Поскольку многочисленные хорошо сохранившиеся экземпляры Ghost Ranch использовались в качестве Целофиз в большей части научной литературы использование Риоаррибазавр было бы очень неудобно для исследователей, поэтому была подана петиция о выдаче типового образца Целофиз перенесен из плохо сохранившегося оригинального образца в один из хорошо сохранившихся образцов Ghost Ranch. Это сделало бы Риоаррибазавр определенный синоним Целофиз, а именно младший объективный синоним.[19] В конце концов, Международная комиссия по зоологической номенклатуре (ICZN) проголосовал за то, чтобы сделать один из образцов Ghost Ranch настоящим образцом для Целофиз и избавиться от имени Риоаррибазавр в целом (объявив это nomen rejectum, или «отвергнутое имя»), разрешая таким образом путаницу. Название Целофиз поэтому стал nomen conservandum («сохраненное имя»).[24]
В ситуации, затрагивающей многие таксоны динозавров, некоторые недавно обнаруженные окаменелости первоначально были классифицированы как новые роды, но могут быть видами Целофиз. Например, проф. Миньон Талбот открытие 1911 года[31] которую она назвала Подокезавр Holyokensis, долгое время считалось связанным с Целофиз,[32] и некоторые современные ученые считают Подокезавр синоним Целофиз.[1] Еще один экземпляр из Формация Портленда бассейна Хартфорд, теперь в Бостонский музей науки, также упоминается Целофиз.[33][34] Этот образец состоит из слепков из песчаника лобка, большеберцовой кости, трех ребер и, возможно, позвонка, и, вероятно, был обнаружен в карьере в Мидлтауне, штат Коннектикут. Однако оба типовых образца Подокезавр и особи из Мидлтауна сегодня обычно считаются неопределенными тероподами.[35]

Салливан и Лукас (1999) сослались на один образец из оригинального материала Коупа: Целофиз (AMNH 2706) тому, что они считали недавно обнаруженным тероподом, Эукоэлофиз.[36] Однако последующие исследования показали, что Эукоэлофиз был ошибочно идентифицирован, и на самом деле это примитивный, не динозавр орнитодиран тесно связан с Силезавр.[37]
Род Syntarsus был назван Раатом в 1969 году в честь типового вида Syntarsus rhodesiensis из Африки, а затем применили к Северной Америке Syntarsus kayentakatae.[25] Он был переименован американским энтомологом доктором Майклом Айви (Государственный университет Монтаны из Бозман ), Польский австралийский доктор Адам Слипинский и польский доктор Петр Венгжинович (Muzeum Ewolucji Instytutu Zoologii PAN of Варшава ), трех ученых, которые обнаружили, что род имя Syntarsus был уже занят колидин жук описан в 1869 г.[38] Многим палеонтологам не понравилось наименование Мегапнозавротчасти потому, что обычно ожидается, что таксономисты позволят первоначальным авторам названия исправлять любые ошибки в своей работе. Раат знал об омонимии между динозаврами Syntarsus и жук Syntarsus, но группа, опубликовавшая Мегапнозавр заставили поверить, что Раат умер и, следовательно, не мог исправить свою ошибку, и поступили соответственно. Мортимер (2012) указал, что «палеонтологи могли бы отреагировать более позитивно, если бы заменяющее имя (Мегапнозавр) не был шутливым, что переводится как «большая мертвая ящерица».[39] Йетс (2005) проанализировал Целофиз и Мегапнозавр и пришел к выводу, что эти два рода почти идентичны, и предположил, что Мегапнозавр возможно было синонимом Целофиз.[40] В 2004 году Раат стал соавтором двух статей, в которых утверждал, что Мегапнозавр (ранее Syntarsus) был младшим синонимом Целофиз.[1] Мегапнозавр был расценен Полом (1988) и Даунсом (2000) как родственный с Целофиз.[9][18] Затем в 1993 году Пол предложил Целофиз должен быть помещен в Мегапнозавр (тогда известный как Syntarsus), чтобы обойти вышеупомянутые таксономический путаница.[41] Даунс (2000) исследовал Кампозавр и пришел к выводу, что это младший синоним слова Целофиз, из-за его сходства с некоторыми из Целофиз Образцы Ghost Ranch.[18] Однако переоценка Кампозавр голотип Мартина Эскурры и Стивена Брусатта, опубликованный в 2011 году, показал пару аутапоморфии в голотипе, указывая, что C. arizonensis не был синонимом К. Баури, хотя это был близкий родственник С. rhodesensis.[42]
Палеобиология
Кормление

Зубы Целофиз были типичны для хищных динозавров, лезвийные, загнутые, острые и зазубренные с мелкими зубцами на обоих передний и задний края. Его зубы показывают, что он был плотоядным, вероятно, охотился на маленьких, похожих на ящериц животных, которые были обнаружены вместе с ним.[43] Он также мог охотиться стаями, чтобы поймать более крупную добычу.[9]Целофиз баури имеет примерно 26 зубов на верхнечелюстной кости верхней челюсти и 27 зубов на зубной кости нижней челюсти.[15] Карпентер (2002) изучил биомеханику передних конечностей теропод и попытался оценить их полезность в борьбе с хищниками. Он пришел к выводу, что передняя конечность Целофиз был гибким и имел хороший диапазон движений, но его костная структура предполагала, что он был сравнительно слабым. «Слабые» передние конечности и мелкие зубы у представителей этого рода позволяют предположить, что Целофиз охотились на животных, которые были значительно меньше его. Райнхарт и другие. согласился, что Целофиз был «охотником на мелкую, быстро движущуюся добычу».[10] Карпентер также выделил три различные модели использования передних конечностей тероподов и отметил, что Целофиз был «комбинацией схватывания-хватки» по сравнению с другими динозаврами, которые были «хватателями» или «хватателями с длинными руками».[44]

Было высказано предположение, что К. Баури был каннибал, основанный на предполагаемых молодых особях, обнаруженных «внутри» брюшной полости некоторых особей Ghost Ranch.[23] Тем не мение, Роберт Дж. Гей показали в 2002 году, что эти образцы были неправильно истолкованы. Несколько экземпляров «ювенильных целофизид» действительно были небольшими. Crurotarsan рептилии Такие как Гесперозух.[45] Позиция Гая была подтверждена в исследовании Несбитта в 2006 году. и другие.[46] В 2009 году были обнаружены новые доказательства каннибализма, когда дополнительная подготовка ранее раскопанной матрицы обнаружила отрыгивающий материал во рту и вокруг него. Целофиз образец НММНХ П-44551. В этот материал вошли фрагменты костей зубов и челюстей, которые Райнхарт и другие. считается «морфологически идентичным» несовершеннолетнему Целофиз.[10]
В 2010 году Гей исследовал кости молодых особей, обнаруженных в грудной полости AMNH 7224, и подсчитал, что общий объем этих костей в 17 раз больше, чем максимальный предполагаемый объем желудка Целофиз образец. Гей заметил, что общий объем был бы еще больше, если учесть, что на этих костях была плоть. Этот анализ также отметил отсутствие следов зубов на костях, как можно было бы ожидать при удалении плесени, и отсутствие ожидаемой язвы желудочных кислот. Наконец, Гей продемонстрировал, что предполагаемые каннибализированные ювенильные кости были отложены стратиграфически под более крупным животным, которое предположительно каннибализировало их. Взятые вместе эти данные позволяют предположить, что Целофиз экземпляр AMNH 7224 не был каннибалом, и что кости молодых и взрослых особей были найдены в их окончательном положении в результате «случайного наложения особей разного размера».[47]
Поведение упаковки

Открытие более 1000 экземпляров Целофиз на карьере Уитакер на Призрачное ранчо, предположил стадное поведение таких исследователей, как Шварц и Джилетт.[48] Существует тенденция рассматривать это массовое скопление животных как свидетельство существования огромных стай животных. Целофиз бродить по земле.[23] Прямых доказательств флокирования не существует; депозиты только указывают на то, что большое количество Целофизвместе с другими животными триаса были похоронены вместе. Некоторые свидетельства из тафономия участка указывает на то, что эти животные могли быть собраны вместе, чтобы кормить или пить из истощенной водяной ямы или кормиться нерест выловили рыбу, а затем погребли в катастрофическом внезапное наводнение[23][48] или засуха.[23]

С 30 экземплярами C. rhodesiensis найденные вместе в Зимбабве, некоторые палеонтологи предположили, что Целофиз действительно был общительным. Опять же, в этом случае нет прямых доказательств стая, и было также высказано предположение, что эти люди также были жертвами внезапных наводнений, поскольку это, кажется, было обычным явлением в тот период.[25][49][50]
Рост и половой диморфизм
Райнхарт (2009) оценил онтогенетический рост этого рода с использованием данных, собранных по длине его верхней кости ноги (бедренная кость ) и пришел к выводу, что Целофиз молодь быстро росла, особенно в течение первого года жизни.[10] Целофиз вероятно, достигли половой зрелости между вторым и третьим годом жизни и достигли своего полного размера, чуть более 10 футов в длину, к восьмому году жизни. Это исследование выявило четыре различных этапа роста: 1 год, 2 года, 4 года и 7+ лет.[10] Также считалось, что как только они вылупятся, им придется заботиться о себе.[23]
Две "морфы" Целофиз были выявлены: более изящный формы, как у экземпляра AMNH 7223, и немного более крепкий формы, как в образцах AMNH 7224 и NMMNH P-42200. Пропорции скелета у этих двух форм были разными;[51] у изящной формы более длинный череп, более длинная шея, более короткие передние конечности и крестцовый сросшиеся нервные отростки; а у крепкой формы более короткий череп, более короткая шея, более длинные передние конечности и несращенные крестцовые нервные отростки.[15] Исторически было выдвинуто множество аргументов в пользу того, что это представляет собой своего рода диморфизм в популяции Целофиз, наверное половой диморфизм.[9][15][52][53] Раат согласился, что диморфизм в Целофиз Об этом свидетельствуют размеры и строение передней конечности.[52] Райнхарт и другие. изучили 15 особей и согласились, что две морфы присутствуют даже у молодых особей, и предположили, что половой диморфизм присутствовал в раннем возрасте, до половой зрелости. Райнхарт пришел к выводу, что изящная форма была женской, а устойчивая - мужской, основываясь на различиях в крестцовых позвонках изящной формы, что позволяло более гибко откладывать яйца.[10] Дальнейшее подтверждение этой позиции было предоставлено анализом, показывающим, что каждая морфа составляла 50% популяции, как и следовало ожидать при соотношении полов 50/50.[54]
Однако более поздние исследования показали, что К. Баури и C. rhodesiensis имели сильно изменчивый рост между особями, при этом некоторые особи были крупнее в незрелой фазе, чем более мелкие взрослые особи в полностью зрелом возрасте; это указывает на то, что предполагаемое присутствие различных морфов - просто результат индивидуальной изменчивости. Этот сильно изменчивый рост, вероятно, был наследником динозавров, но позже утрачен и, возможно, дал таким ранним динозаврам эволюционное преимущество в выживании в суровых экологических условиях.[55]
Размножение
Путем составления и анализа базы данных почти трех десятков птиц и рептилий и сравнения с существующими данными об анатомии Целофиз Райнхарт и другие. (2009) сделали следующие выводы. Было подсчитано, что среднее яйцо Целофиз был 31–33,5 миллиметра в поперечнике, и каждая самка откладывала от 24 до 26 яиц в каждой кладке. Имеющиеся данные свидетельствуют о том, что для выкармливания относительно небольших птенцов в течение первого года жизни необходима определенная родительская забота, когда к концу первой стадии роста они достигают 1,5 метров в длину. Целофиз баури вложили в воспроизводство столько же энергии, сколько и другие вымершие рептилии своего примерного размера.[10][56]
Палеопатология
В исследовании 2001 года, проведенном Брюсом Ротшильдом и другими палеонтологами, 14-футовые кости относились к Целофиз были исследованы на наличие признаков стресс-перелом, но ничего не было найдено.[57]
В C. rhodesiensis, залечил переломы большеберцовая кость и плюсна наблюдались, но очень редко. «[Т] он поддерживает подпорки второго крестцового ребра» в одном образце Syntarsus rhodesiensis проявляли признаки колеблющейся асимметрии. Колеблющийся асимметрия возникает в результате нарушений развития и чаще встречается у людей, находящихся в состоянии стресса, и поэтому может быть информативным о качестве условий, в которых жил динозавр.[58]
Палеоэкология

Распределение
Образцы Целофиз были восстановлены из Формирование Чинле Нью-Мексико и Аризона, более известный в карьере Призрачного ранчо (Уитакер) в члене Рок-Пойнт[10] среди других карьеров в нижележащей пачке Окаменевшего леса, отложения которой датируются примерно 212 миллионов лет назад, что делает их частью среднего Norian этап позднего триаса.[59][23]
C. rhodesiensis был восстановлен в Формация Верхний Эллиотт в мыс и Свободный штат провинции ЮАР, а также карьер с костным слоем реки Читаке на Формирование лесного песчаника в Зимбабве.
Фауна и среда обитания
Призрачное ранчо было расположено недалеко от экватора более 200 миллионов лет назад и имело теплый, похожий на муссон климат с обильными сезонными осадками. Хайден Карьер, новое место раскопок на ранчо Призраков, Нью-Мексико, дало разнообразную коллекцию ископаемых материалов, которая включала первые свидетельства существования динозавров и менее развитых динозавроморфов того же периода. Открытие указывает на то, что две группы жили вместе в начале триасового периода 235 миллионов лет назад.[60]
Терриен и Фастовский (2001) исследовали палеосреду Целофиз и другие ранние тероподы из национального парка «Окаменевший лес» в Аризоне, и определили, что этот род обитал в среде, состоящей из пойм, отмеченных четко выраженными сухими и влажными сезонами. В более засушливые времена, когда животные боролись за воду в высыхающих руслах рек, была большая конкуренция.[61]
В верхних разрезах чинлской свиты, где Целофиз найдено, динозавры были редкостью, пока только Чиндезавр и Демонозавр[62] известны, а в наземной фауне преобладают другие рептилии, такие как ринхоцефальные Whitakersaurus;[63] в псевдозухий Ревуэльтозавр;[64] этозавры Десматозух,[64] Типоторакс[64] и Гелиокантус;[64] крокодиломорф Гесперозух;[64] "рауйсухиане" Шувозавр,[64] Эффигия[65] и Виварон;[66] наряду с другими редкими компонентами, такими как динозавр Эукоэлофиз,[67] и амниота Кратерохейродон.[64] В водных путях мы находим фитозавр Machaeroprosopus;[64] архозавроморф Vancleavea;[64] амфибии Апахезавр[64] и Koskinonodon;[68] и рыбы Ретикулодус,[64] Арганодус[64] и Lasalichthyes.[64]
Считается, что формация Верхний Эллиот была древней поймой. Окаменелости динозавра прозауроподов Массоспондил и Игнавузавр были извлечены из формации Верхний Эллиот, которая может похвастаться самой разнообразной в мире фауной раннеюрских орнитисхийских динозавров, включая Абриктозавр, Фаброзавр, Гетеродонтозавр, и Лесотозавр, среди прочего. Формация Лесного Песчаника была палеосредой протосухидных крокодилов, сфенодонтов, динозавров. Массоспондил и неопределенные останки прозауропода. Пол (1988) отметил, что Мегапнозавр родезийский жили среди пустынных дюн и оазисов, охотились на молодых и взрослых прозауроподов.[69]
Тафономия
Множество образцов, так близко хранящихся в Призрачное ранчо вероятно, был результатом внезапного наводнения, унесшего большое количество Целофиз и похоронил их быстро и одновременно. На самом деле, похоже, что такое наводнение было обычным делом в тот период История Земли и, действительно, Окаменелый лес поблизости Аризона Это результат сохранившегося затора из стволов деревьев, попавших в один такой паводок. Карьер Уитакера на ранчо Призраков считается монотаксическим участком, потому что в нем есть несколько особей одного таксона. Качество сохранности и онтогенный (возрастной) диапазон особей способствовали формированию Целофиз один из самых известных из всех родов.[70] В 2009 году Райнхарт и другие. отметил, что в одном случае Целофиз образцы были «смыты в топографический низменность с небольшим прудом, где они, вероятно, утонули и были захоронены в результате наводнения из близлежащей реки».[10]
30 образцов C. rhodesiensis найденные вместе в Зимбабве также, вероятно, были результатом внезапного наводнения, которое унесло большое количество Целофиз и закопали их быстро и одновременно.[25][49][50]
Технология

Эдвин Х. Колберт предположил, что следы теропод относятся к ихноген Grallator, расположенный в Долина реки Коннектикут через Коннектикут и Массачусетс, возможно, были сделаны Целофиз.[71] Следы относятся к супергруппе Ньюарк, возраст которой от позднего триаса до ранней юры. На них отчетливо видны пальцы II, III и IV, но не I или V. Это странное состояние для следов их возраста. Считалось, что пальцы короткие и неэффективные, они не касались земли, когда динозавр шел или бежал. Считается, что они произошли от неустановленного примитивного заурисха, похожего на Целофиз Дэвид Б. Вейшампель и Л. Янг, совсем недавно.[72] Скелетные останки напоминают Целофиз также были найдены в долине, что подтверждает идею о том, что вид, похожий на Целофиз отвечает за следы.[73]
Следы динозавров, которые позже были приписаны C. rhodesiensis были обнаружены в Родезии в 1915 году. Эти следы были обнаружены в формации песчаников Ньямандхлову в эоловом красном песчанике, который откладывался в Поздний триас, примерно от 235 до 201 миллиона лет назад.[74]
В популярной культуре
Целофиз был вторым динозавром в космосе после Майасаура (СТС-51-Ф ).[75] А Целофиз череп из Музей естественной истории Карнеги был на борту Космический шаттл индевор миссия СТС-89 когда он покинул атмосферу 22 января 1998 года. Его также доставили на космическую станцию. Мир перед возвращением на Землю.[75][76]
Будучи старше 100 лет, Целофиз - один из самых известных динозавров в литературе. Он был обозначен как официальный окаменелость штата Нью-Мексико в 1981 году, а теперь это логотип Музея естественной истории Нью-Мексико.[19][77]
Рекомендации
- ^ а б c d е ж Бристоу, А. и М.А. Раат (2004). "Молодой целофизоидный череп из ранней юры Зимбабве и синонимия Целофиз и Syntarsus.(СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ)". Palaeontologica Africana. 40 (40): 31–41.CS1 maint: использует параметр авторов (связь)
- ^ а б Каррано, Мэтью Т .; Бенсон, Роджер Б. Дж .; Сэмпсон, Скотт Д. (2012). «Филогения столбняка (Dinosauria: Theropoda)». Журнал систематической палеонтологии. 10 (2): 211–300. Дои:10.1080/14772019.2011.630927. S2CID 85354215.
- ^ Nesbitt, Sterling J .; Irmis, Randall B .; Паркер, Уильям Г. (2007). «Критическая переоценка таксонов динозавров позднего триаса Северной Америки» (PDF). Журнал палеонтологии. 5 (2): 209–243. Дои:10.1017 / с1477201907002040.
- ^ Несбитт, Ирмис и Паркер, 2007. Критическая переоценка таксонов динозавров позднего триаса Северной Америки. Журнал систематической палеонтологии. 5 (2), 209–243.
- ^ "RE: Афровенатор произношение". dml.cmnh.org. Получено 26 мая 2019.
- ^ а б Гейнс, Ричард М. (2001). Целофиз. Издательская компания АБДО. п. 4. ISBN 978-1-57765-488-9.
- ^ Weishampel, David B; и другие. (2004). «Распространение динозавров (ранняя юра, Африка)». В: Weishampel, David B .; Додсон, Питер; и Осмольская, Хальска (ред.): Динозаврия, 2-е, Беркли: Калифорнийский университет Press. Стр. 535–536. ISBN 0-520-24209-2.
- ^ Schwartz, Hilde L .; Gillette, Дэвид Д. (1994). "Геология и тафономия Целофиз карьер, формация Чинл в верхнем триасе, Призрачное ранчо, Нью-Мексико ". Журнал палеонтологии. 68 (5): 1118–1130. Дои:10.1017 / S0022336000026718. JSTOR 1306181.
- ^ а б c d Пол, Грегори С. (1988). Хищные динозавры мира. Саймон и Шустер. п.260. ISBN 978-0-671-61946-6.
- ^ а б c d е ж грамм час я j k Rinehart, L.F .; Lucas, S.G .; Heckert, A.B .; Шпильманн, Я. И Селески, доктор медицины (2009). "Палеобиология Целофиз баури (Cope) из верхнего триаса (апачей) карьера Уитакер, Нью-Мексико, с подробным анализом отдельного блока карьера ". Музей естественной истории и науки Нью-Мексико, бюллетень отдела по делам культуры. 45: 260.CS1 maint: использует параметр авторов (связь)
- ^ Райнхарт, Ларри Ф .; Лукас, Спенсер Дж .; Хант, Адриан П. (2007). "Фуркулы в позднем триасовом динозавре теропод Целофиз баури". Paläontologische Zeitschrift. 81 (2): 174–180. Дои:10.1007 / bf02988391.
- ^ а б c Тыкоски, Р. И Роу, Т. (2004). «Цератозаврия»: В: Weishampel, D.B .; Dodson, P .; Осмольска, Х., ред. (6 ноября 2004 г.). Динозаврия (Второе изд.). Беркли: Калифорнийский университет Press. С. 47–70. ISBN 978-0-520-24209-8.
- ^ Гей, Р.Дж. (2001). "Необычное приспособление хвостовых позвонков Целофиз баури (Динозавры: Theropoda) ". Палеобиос. 21: 55.
- ^ Rinehart, L.F .; Heckert, A.B .; Лукас, С.Г. и Хант, А.П. (2004). "Склеротическое кольцо тероподового динозавра позднего триаса Целофиз". Весеннее собрание Геологического общества Нью-Мексико. 26: 64.CS1 maint: использует параметр авторов (связь)
- ^ а б c d е Кольбер, Э. (1989). "Триасовый динозавр Целофиз". Бюллетень Музея Северной Аризоны. 57: 160.
- ^ Эскурра, доктор медицины (2007). "Черепная анатомия целофизоидного теропод. Zupaysaurus rougieri из верхнего триаса Аргентины ». Историческая биология. 19 (2): 185–202. Дои:10.1080/08912960600861467. S2CID 129449572.
- ^ а б Бристоу, А. и Раат, М.А. (2004). "Череп ювенильного целофизоида из ранней юры Зимбабве и синонимия Целофиз и Syntarsus" (PDF). Palaeontologia Africana. 40: 31–41. ISSN 0078-8554.CS1 maint: использует параметр авторов (связь)
- ^ а б c Даунс, А. (2000). "Целофиз баури и Syntarsus rhodesiensis по сравнению с комментариями о подготовке и сохранении окаменелостей из карьера Ghost Ranch Coelophysis »: В: Lucas, S.G .; Гекерт, А. (ред.). «Динозавры Нью-Мексико». Бюллетень Музея естественной истории Нью-Мексико. 17: 33–37.
- ^ а б c Colbert, E.H .; Charig, A.J .; Dodson, P .; Gillette, D.D .; Остром, Дж. И Вайшампель, Д. (1992). "Coelurus bauri Коп, 1887 г. (в настоящее время Целофиз баури; Reptilia, Saurischia): Предлагаемая замена лектотипа неотипом » (PDF). Вестник зоологической номенклатуры. 49 (4): 276–279.CS1 maint: использует параметр авторов (связь)
- ^ а б c Коуп, Э. (1889 г.). «О новом роде триасовых динозавров». Американский натуралист. 23 (271): 621–633. Дои:10.1086/274979.
- ^ а б Глют, Д.Ф. (1999). Динозавры, Энциклопедия, Приложение 1. McFarland & Company, Inc. стр. 442. ISBN 978-0-7864-0591-6.
- ^ Коуп, Э. (1887 г.). «Род динозавров Coelurus». Американский натуралист. XXI 5: 367-369.
- ^ а б c d е ж грамм Хейнс, Т. и Чемберс, П. (2007). Полное руководство по доисторической жизни. Книги Светлячка. стр.70 –71. ISBN 978-1-55407-181-4.CS1 maint: использует параметр авторов (связь)
- ^ а б Международная комиссия по зоологической номенклатуре (1996). «Мнение 1842: Coelurus bauri Коп, 1887 г. (в настоящее время Целофиз баури; Reptilia, Saurischia): лектотип заменен неотипом » (PDF). Вестник зоологической номенклатуры. 53 (2): 142–144.
- ^ а б c d Раат (1969). «Новый динозавр целурозавров из лесных песчаников Родезии». Арнольдия Родезия. 4 (28): 1–25.
- ^ Tykoski, R. S., и Rowe, T., 2004, Ceratosauria, Chapter Three: In: The Dinosauria, Second Edition, отредактированный Weishampel, D.B., Dodson, P., and Osmolska, H., California University Press, p. 47-70.
- ^ Сьюз, H.D .; Несбит, S.J .; Berman, D.S .; Хенрици, A.C. (2011). "Выживший поздно базальный динозавр теропод из позднего триаса Северной Америки". Труды Королевского общества B. 278 (1723): 3459–3464. Дои:10.1098 / rspb.2011.0410. ЧВК 3177637. PMID 21490016.
- ^ "Coelophysoidea". theropoddatabase.com. Получено 5 февраля 2016.
- ^ «Вариации лица у Coelophysis bauri и статус мегапнозавра (Syntarsus)». nmstatefossil.org. Получено 5 февраля 2016.
- ^ Хант, Адриан П .; Лукас, Спенсер Г. (1991). "Риоаррибазавр, новое название позднетриасового динозавра из Нью-Мексико (США) ». Paläontologische Zeitschrift. 65 (1/2): 191–198. Дои:10.1007 / bf02985783.
- ^ Талбот, М. (1911). "Подокезавр Holyokensis, новый динозавр из триаса в долине Коннектикута ". Американский журнал науки. 4. 31 (186): 469–479. Bibcode:1911AmJS ... 31..469T. Дои:10.2475 / ajs.s4-31.186.469.
- ^ Кольбер, Э. (1964). "Триасовые роды динозавров Podokesaurus и Coelophysis". Американский музей Novitates. 2168: 1–12.
- ^ Colbert, E .; Бэрд, Д. (1958). «Отливки костей целурозавра из триаса Коннектикутской долины». Американский музей Novitates. 1901: 1–11.
- ^ Getty, P.R .; Буш, А. М. (2011). «Песочные псевдоморфы костей динозавров: последствия для (не) сохранения скелетного материала четвероногих в бассейне Хартфорд, США». Палеогеография, палеоклиматология, палеоэкология. 302 (3–4): 407–414. Bibcode:2011ППП ... 302..407Г. Дои:10.1016 / j.palaeo.2011.01.029.
- ^ Ольшевский, Г. (1991). "Пересмотр параинфракласса Archosauria Cope, 1869, за исключением продвинутых Crocodylia" (PDF). Мезозойские меандры 2: 196.
- ^ Салливан, Роберт М .; Лукас, Спенсер Г. (1999). "Eucoelophysis baldwini, новый динозавр теропод из верхнего триаса штата Нью-Мексико, и статус первоначальных типов Целофиз". Журнал палеонтологии позвоночных. 19 (1): 81–90. Дои:10.1080/02724634.1999.10011124.
- ^ Nesbitt, Sterling J .; Irmis, Randall B .; Паркер, Уильям Г. (2007). «Критическая переоценка таксонов динозавров позднего триаса Северной Америки». Журнал систематической палеонтологии. 5 (2): 209–243. Дои:10.1017 / S1477201907002040. S2CID 28782207.
- ^ Ivie, M. A .; Lipiński, S.A .; Węgrzynowicz, P. (2001). "Общие омонимы в Colydiinae (Coleoptera: Zopheridae)". Insecta Mundi. 15: 63–64.
- ^ Мортимер, Микки (2012). "Coelophysoidea". Архивировано из оригинал 4 мая 2013 г.
- ^ Йейтс, А. (2005). «Новый динозавр теропод из ранней юры Южной Африки и его значение для ранней эволюции теропод». Palaeontologia Africana. 41: 105–122.
- ^ Пол Г.С. (1993). "Находятся Syntarsus и теропод Уайттакер того же рода? ": В: Lucas, S.G .; Моралес, М. (ред.). «Неморский триас». Бюллетень Музея естественной истории Нью-Мексико. 3: 397–402.
- ^ Ezcurra, MD; Брусатте, С. (2011). "Таксономическая и филогенетическая переоценка раннего неотероподного динозавра Camposaurus arizonensis из позднего триаса Северной Америки ». Палеонтология. 54 (4): 763–772. Дои:10.1111 / j.1475-4983.2011.01069.x.
- ^ Бентон, М. (1993). Поиск фактов о динозаврах и других доисторических животных. Kingfisher Books Ldt. п. 256. ISBN 978-0-86272-949-3.
- ^ Карпентер, К. (2002). "Биомеханика передних конечностей нептичьих динозавров-теропод в хищничестве". Senckenbergiana Lethaea. 82: 59–76. Дои:10.1007 / bf03043773.
- ^ Гей, Р.Дж. (2002). "Миф о каннибализме в Целофиз баури". Журнал палеонтологии позвоночных. 22 (3): 57А.
- ^ Nesbitt, S.J .; Тернер, А. Х; Эриксон, Г. М.; Норелл М.А. (2006). "Выбор добычи и каннибалистическое поведение теропод Целофиз". Письма о биологии. 22. 2 (4): 611–614. Дои:10.1098 / rsbl.2006.0524. ЧВК 1834007. PMID 17148302.
- ^ Гей, Р.Дж. (2010a). Заметки о тероподах раннего мезозоя (Первое изд.). Лулу пресс. С. 9–24. ISBN 978-0-557-46616-0.
- ^ а б Шварц, Х.Л. и Жиллетт, Д.Д. (1994). «Геология и тафономия карьера Coelophysis, формация Chinle верхнего триаса, Ghost Ranch, Нью-Мексико». Журнал палеонтологии. 68 (5): 1118–1130. Дои:10.1017 / S0022336000026718. JSTOR 1306181.CS1 maint: использует параметр авторов (связь)
- ^ а б Дж. Бонд, 1965. Некоторые новые ископаемые местонахождения в системе Карроо в Родезии. Арнольдия, Серия разных публикаций, Национальный музей Южной Родезии 2 (11): 1–4
- ^ а б М. А. Раат, 1977. Анатомия триасового теропод Syntarsus rhodesiensis (Saurischia: Podokesauridae) и рассмотрение его биологии. Кафедра зоологии и энтомологии, Родосский университет, Солсбери, Родезия 1–233
- ^ Пол, Грегори С. (2010). Принстонский полевой путеводитель по динозаврам. Издательство Принстонского университета. п.74. ISBN 978-0-691-13720-9.
- ^ а б Карпентер, К .; Карри, П.Дж. (1990). Систематика динозавров: подходы и перспективы. Кембридж: Издательство Кембриджского университета. С. 91–105. ISBN 978-0-521-43810-0.
- ^ Гей Р. (2005). "Половой диморфизм у раннеюрских динозавров теропод Дилофозавр и сравнение с другими родственными формами »: В: Карпентер, К., изд. (2005). Плотоядные динозавры. Блумингтон: Издательство Индианского университета. С. 277–283. ISBN 978-0-253-34539-4.
- ^ Rinehart, L.F .; Lucas, S.G .; Гекерт, А. (2001). "Предварительный статистический анализ, определяющий ювенильные, крепкие и изящные формы триасовых динозавров. Целофиз". Журнал палеонтологии позвоночных. 21: 93A. Дои:10.1080/02724634.2001.10010852. S2CID 220414868.
- ^ Гриффин, C.T .; Несбитт, С.Дж. (2016). «Аномально высокая вариабельность в постнатальном развитии является наследственной для динозавров, но теряется у птиц». Труды Национальной академии наук Соединенных Штатов Америки. 113 (51): 14757–14762. Bibcode:2016PNAS..11314757G. Дои:10.1073 / pnas.1613813113. ЧВК 5187714. PMID 27930315.
- ^ Глют, Д.Ф. (2012). Динозавры, Энциклопедия, Приложение 7. McFarland & Company, Inc. стр. 866. ISBN 978-0-7864-4859-3.
- ^ Ротшильд, В .; Танке, Д.Х. и Форд, Т.Л. (2001). «Стресс-переломы теропод и отрывы сухожилий как ключ к активности»: В: Tanke, D. H .; Карпентер, К., ред. (2001). Мезозойская жизнь позвоночных. Издательство Индианского университета. С. 331–336. ISBN 978-0-253-33907-2.
- ^ Мольнар, Р. Э., 2001, Палеопатология теропод: обзор литературы: В: Мезозойская жизнь позвоночных, под редакцией Танке Д. Х. и Карпентером К., Indiana University Press, стр. 337-363.
- ^ Irmis, Randall B .; Мундил, Роланд; Марц, Джеффри В .; Паркер, Уильям Г. (15 сентября 2011 г.). «U – Pb датировки высокого разрешения из формации Чинл в верхнем триасе (Нью-Мексико, США) подтверждают диахронный подъем динозавров». Письма по науке о Земле и планетах. 309 (3–4): 258–267. Bibcode:2011E и PSL.309..258I. Дои:10.1016 / j.epsl.2011.07.015.
- ^ Брагинец, Донна (2007). «Новый вид динозавроморфов (внизу слева) был среди смешанного сообщества динозавров и динозавроморфов, найденных в карьере Хайдена на ранчо призраков, штат Нью-Мексико». Американский музей естественной истории. Получено 31 марта 2013.
- ^ Therrien, F .; Фастовский, Д. Э. (2000). «Палеосреды ранних теропод, формация Чинл (поздний триас), Национальный парк Окаменелого леса, Аризона». ПАЛАИ. 3. 15 (3): 194–211. Bibcode:2000Палай..15..194т. Дои:10.1669 / 0883-1351 (2000) 015 <0194: POETCF> 2.0.CO; 2.
- ^ Сьюз, H. -D .; Nesbitt, S.J .; Берман, Д. С .; Хенрици, А. С. (2011). "Выживший поздно базальный динозавр теропод из позднего триаса Северной Америки". Труды Королевского общества B: биологические науки. 278 (1723): 3459–64. Дои:10.1098 / rspb.2011.0410. ЧВК 3177637. PMID 21490016.
- ^ Heckert, A.B .; Lucas, S.G .; Rinehart, L.F .; Хант, А.П. (2008). "Новый род и вид сфенодонтов с ранчо призраков. Целофиз Карьер (верхний триас: апачей), формация Рок-Пойнт, Нью-Мексико, США ". Палеонтология. 51 (4): 827–845. Дои:10.1111 / j.1475-4983.2008.00786.x.
- ^ а б c d е ж грамм час я j k л м Ирмис, Р. Б. (2005). «Фауна позвоночных чинлской формации верхнего триаса в северной Аризоне»: В: Несбит, S.J .; Паркер, W.G .; Ирмис, Р. Б. (ред.). «Путеводитель по триасовым образованиям плато Колорадо в северной Аризоне: геология, палеонтология и история» (PDF). Бюллетень Юго-Западного музея Меса. 9: 63–88. Архивировано из оригинал (PDF) 4 октября 2012 г.. Получено 2 января 2014.
- ^ Benson, R.J .; Brusatte, S .; Clack, J .; Андерсон, Дж .; Деннис-Брайан, К .; Hone, D .; Duffin, C .; Johanson, Z .; Milner, A .; Naish, D .; Parsons, K .; Prothero, D .; Сюй, X. (2012). Доисторическая жизнь. Лондон: Дорлинг Киндерсли. С. 214–215. ISBN 978-0-7566-9910-9.CS1 maint: использует параметр авторов (связь)
- ^ Эмили Дж. Лесснер; Мишель Р. Стокер; Натан Д. Смит; Алан Х. Тернер; Рэндалл Б. Ирмис; Стерлинг Дж. Несбитт (2016). «Новый rauisuchid (Archosauria, Pseudosuchia) из верхнего триаса (норийский) Нью-Мексико увеличивает разнообразие и временной диапазон клады». PeerJ. 4: e2336. Дои:10.7717 / peerj.2336. ЧВК 5018681. PMID 27651983.
- ^ Weishampel, D.B .; Dodson, P .; И Осмольска, Х. (2004). Динозаврия (второе изд.). Беркли: Калифорнийский университет Press. С. 518–521. ISBN 978-0-520-24209-8.CS1 maint: использует параметр авторов (связь)
- ^ Мюллер, Б. (2007). "Koskinonodon Брэнсон и Мель, 1929, заменяющее название озабоченного темноспондила Buettneria Дело, 1922 год ». Журнал палеонтологии позвоночных. 27 (1): 225. Дои:10.1671 / 0272-4634 (2007) 27 [225: KBAMAR] 2.0.CO; 2.
- ^ Пол, Г.С., 1988, Хищные динозавры мира, полное иллюстрированное руководство: книга Нью-Йоркской академии наук, 464 стр.
- ^ Глют, Д.Ф. (2006). Динозавры, Энциклопедия, Приложение 4. McFarland & Company, Inc. стр. 749. ISBN 978-0-7864-2295-1.
- ^ Колберт, Эдвин Х. (1965). Эпоха рептилий. W. W. Norton & Company. п.97. ISBN 978-0-486-29377-6.
- ^ Weishampel, D.B; Янг, Л. (1998). Динозавры восточного побережья. Издательство Университета Джона Хопкинса. ISBN 978-0-8018-5217-6.
- ^ «Следы динозавров в долине реки Коннектикут». Сайт трека динозавров Нэша и рок-магазин.
- ^ М. А. Раат. 1972 г. Первая запись следов динозавра в Родезии. Арнольдия 5 (37): 1–5
- ^ а б Чуре, Д. (2009). "кости динозавра в космосе - это был пиар". Кливлендский музей естественной истории. Получено 12 ноября 2011.
- ^ Паркер, Стив (2003). Динозавры: полное руководство по динозаврам. Книги Светлячка. ISBN 978-1-55297-772-9.
- ^ https://geoinfo.nmt.edu/publications/periodicals/earthmatters/17/n2/em_v17_n2.pdf
внешняя ссылка
- Целофиз в Справочнике Дино
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||













