Herrerasaurus - Herrerasaurus
| Herrerasaurus | |
|---|---|
 | |
| Навесной каркас литой, Полевой музей | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Семья: | †Herrerasauridae |
| Род: | †Herrerasaurus Рейг, 1963 |
| Разновидность: | †H. ischigualastensis |
| Биномиальное имя | |
| †Herrerasaurus ischigualastensis Рейг, 1963 год. | |
| Синонимы | |
| |
Herrerasaurus был родомзаурисхий динозавр из Поздний триас период. Этот род был одним из первых динозавры из летописи окаменелостей. Его название означает «ящерица Эрреры» в честь владельца ранчо, который обнаружил первый экземпляр в 1958 году в Южной Америке. Все известные окаменелости этого хищника были обнаружены в Формация Исчигуаласто из Карнийский возраст (поздно Триасовый согласно ICS датируется 231,4 миллиона лет назад) на северо-западе Аргентины.[1] В типовой вид, Herrerasaurus ischigualastensis, был описан Освальдо Рейг в 1963 г.[2] и это единственный разновидность назначен на род. Искизавр и Frenguellisaurus находятся синонимы.
На протяжении многих лет классификация Herrerasaurus было неясно, потому что он был известен по очень фрагментарным останкам. Это было выдвинутый быть базальный теропод, базальный завроподоморф, базальный заурисхий или вообще не динозавр, а другой тип архозавр. Однако с открытием почти полного скелета и черепа в 1988 г.[3][4] Herrerasaurus был классифицирован как ранний теропод или ранний заурисхий, по крайней мере, в пяти недавних обзорах эволюции теропод, и многие исследователи рассматривают его, по крайней мере, ориентировочно, как самого примитивного представителя теропод.[5]
Он является членом Herrerasauridae, семейство подобных родов, относящихся к самым ранним из динозавр эволюционное излучение.[6][7]
Описание

Herrerasaurus был легко сложенным двуногим хищником с длинным хвостом и относительно небольшой головой. Взрослые имели черепа до 56 см (22 дюйма) в длину и до 6 м (20 футов) в длину.[4] и весом 350 кг (770 фунтов).[8] Меньшие экземпляры были вдвое меньше, с черепом всего около 30 см (12 дюймов) в длину.[9]
Herrerasaurus был полностью двуногим. У него были сильные задние конечности с короткими бедра и довольно длинные ноги, указывающие на то, что это был, вероятно, быстрый бегун. На ступне было пять пальцев, но только три средних (пальцы II, III и IV) несли вес. Наружные пальцы (I и V) были маленькими; у первого пальца был небольшой коготь. Хвост, частично усиленный перекрывающимися выступами позвонков, уравновешивал тело, а также был приспособлением для скорости.[9] Передние конечности Herrerasaurus были меньше половины длины его задних конечностей. В плечо и предплечье были довольно короткими, а манус (рука) была удлиненной. Первые два пальца и большой палец оканчивались изогнутыми острыми когтями для захвата добычи. Четвертый и пятый пальцы были маленькими окурками без когтей.[4][10]
Herrerasaurus отображает черты, присущие различным группам динозавров, и несколько черт, присущих нединозавровым архозаврам. Хотя он имеет большинство характеристик динозавров, есть несколько отличий, особенно в форме костей бедра и ног. Его таз похож на таз заурисхиевых динозавров, но имеет костлявую форму. вертлужная впадина (где бедренная кость встречает таз ), который был открыт лишь частично. В подвздошная кость, основная бедренная кость, поддерживается только двумя крестцовые, а базальный черта.[9] Тем не менее лобок указывает назад, а полученный черта как видно в дромеозавриды и птицы. Кроме того, конец лобка имеет форму ботинка, как у автероподы; и позвоночный центр имеет песочные часы форма, как найдено в Аллозавр.[8]

Herrerasaurus был длинный, узкий череп в котором отсутствовали почти все специализации, характерные для более поздних динозавров,[11] и больше походили на более примитивные архозавры Такие как Euparkeria. В нем было пять пар фенестры (отверстия в черепе) в его черепе, две пары из которых были для глаз и ноздрей. Между глазами и ноздрями было два анторбитальные отверстия и пара крошечных щелевидных отверстий длиной 1 см (0,39 дюйма), называемых промаксиллярными отверстиями.[12]
Herrerasaurus в нижней челюсти имелся гибкий сустав, который мог скользить вперед и назад для захвата.[11] Эта черепная специализация необычна среди динозавров, но в некоторых странах развивалась независимо. ящерицы.[13] Задняя часть нижней челюсти также имела отверстия. Челюсти были снабжены большими зубчатыми зубами для кусания и поедания мяса, а шея была тонкой и гибкой.[11][14]
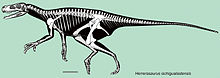
Согласно Новасу (1993), Herrerasaurus можно выделить по следующим признакам:[15] наличие предчелюстная кость -верхняя челюсть фенестра, а дорсальная часть латеротемпорального отверстия менее трети ширины вентральной части; наличие гребня на боковой поверхности скуловой кость и глубоко надрезанный надвисочный ямка который проходит через медиальный заглазничный отросток; субквадратный вентральный чешуйчатый отросток имеет латеральное вдавление, а четырехъядерный кость перекрывает постеродорсальный квадратный лицо; то крыловидный отросток квадратной кости имеет загнутый, желобовидный вентральный край и наличие тонкой ребристой постеродорсальной зубной процесс; то надугловой кость имеет раздвоенный передний отросток для сочленения с постеродорсальным зубным отростком; то плечевая кость ' внутренний бугристость выступает проксимально и отделяется от головки плечевой кости глубокой бороздкой (также имеется у целофизоидов); обладает увеличенными кистями, которые составляют 60% от размера плечевой кости + лучевой кости, а плечевой надмыщелок гребневидный с передним и задним вдавлениями; и задняя граница подвздошный цветонос образует прямой угол с дорсальной границей вала на седалищная кость.
Согласно Серено (1993), Herrerasaurus можно отличить по следующим признакам, все из которых неизвестны другим герреразавридам:[16] круглая яма присутствует на плечевой ectepicondyle, особенность, также присутствующая в Сатурналии; седловидный локтевой мыщелок плечевая кость, а суставная поверхность для локтевой на локтевая кость выпуклый; суставная поверхность локтевой кости меньше, чем у локтевой кости, что неизвестно Стаурикозавр и Санджуанзавр; центральная часть расположена дистальнее лучевой кости; широкий субнарный отросток предчелюстной кости и широкая надвисочная депрессия (отмечено Sereno и Novas, 1993);[17] базальный клубень и затылочный мыщелок примерно равны по ширине (отмечено Sereno, Novas, 1993).[17]
История

Herrerasaurus был назван палеонтолог Освальдо Рейг после Викторино Эрреры, андского жителя козопас кто первым заметил это окаменелости в обнажениях близ г. Сан-Хуан, Аргентина в 1959 г.[2] Эти породы, которые позже дали Эораптор,[18] являются частью Формация Исчигуаласто и свидание с конца Ладинский рано Карнийский этапы позднего Триасовый период.[19] Рейг назвал второго динозавра из этих камней в той же публикации, что и Herrerasaurus;[2] этот динозавр, Ischisaurus cattoi, теперь считается младший синоним и малолетний из Herrerasaurus.[14]
Рейг верил Herrerasaurus был ранним примером карнозавр,[2] но в течение следующих 30 лет это было предметом многочисленных споров, и в течение этого времени этот род был классифицирован по-разному. В 1970 году сталь классифицирована Herrerasaurus как прозауропод.[20] В 1972 г. Питер Гальтон классифицировал этот род как не диагностируемый за пределами Заурисхия.[21] Позже, используя кладистический анализ, некоторые исследователи полагают Herrerasaurus и Стаурикозавр у основания дерева динозавров до разделения орнитисхий и заурисхий.[22][23][24][25] Несколько исследователей классифицировали останки как нединозавры.[26]
Два других частичных скелета с черепным материалом были названы Frenguellisaurus ischigualastensis к Фернандо Новас в 1986 г.[27] но этот вид теперь тоже считается синонимом.[14] Frenguellisaurus ischigualastensis был открыт в 1975 г. и описан Новасом (1986), который считал его примитивным заурисхом и, возможно, теропод. Новас (1992) и Серено и Новас (1992) исследовали Frenguellisaurus остается и считает их относящимися к Herrerasaurus.[28] Ischisaurus cattoi был обнаружен в 1960 году и описан Рейгом в 1963 году. Новас (1992) и Серено и Новас (1992) рассмотрели его останки и пришли к выводу, что их можно отнести к Herrerasaurus.[28]
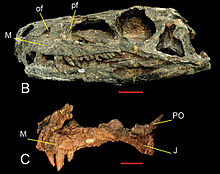
Полный Herrerasaurus череп был найден в 1988 году группой палеонтологов во главе с Пол Серено.[4] На основе новых окаменелостей такие авторы, как Томас Хольц[29] и Хосе Бонапарт[30] классифицированный Herrerasaurus у основания заурисхиевого дерева до расхождения между прозауроподами и теропод. Однако Серено предпочитал классификацию Herrerasaurus (и Herrerasauridae) как примитивные теропод. Эти две классификации стали наиболее устойчивыми, согласно Rauhut (2003)[31] и Биттенкур и Келлнер (2004)[32] в пользу ранних теропод гипотеза, и Макс Лангер (2004),[9] Лангер и Наклонился на (2006),[33] и Рэндалл Ирмис и его соавторы (2007)[34] в пользу гипотезы базального заурисхия. Если Herrerasaurus действительно были тероподом, это означало бы, что тероподы, зауроподоморфы, и орнитисхианы разошлись даже раньше герреразаврид, до середины Карнийский, и что «все три линии независимо друг от друга развили некоторые особенности динозавров, такие как более развитый голеностопный сустав или открытая вертлужная впадина».[35] Это мнение дополнительно подтверждается ихнологический записи, показывающие большие трехпалые (трехпалые) следы, которые можно отнести только к тероподным динозаврам. Эти следы датируются Ладинский (Средний триас) формации Лос-Растрос в Аргентине и до Herrerasaurus от 3 до 5 миллионов лет.[36][37]
Изучение ранних динозавров, таких как Herrerasaurus и Эораптор поэтому имеет важное значение для концепции динозавров как монофилетический группа (группа, произошедшая от общего предка). Монофилия динозавров была явно предложена в 1970-х годах Гальтоном и Роберт Т. Баккер,[38][39] кто составил список черепных и посткраниальных синапоморфии (общие анатомические черты, полученные от общего предка). Позднее авторы предложили дополнительные синапоморфии.[22][23] Обширное исследование Herrerasaurus Серено в 1992 г. предположил, что из этих предложенных синапоморфий только одна краниальная и семь посткраниальных особенностей фактически произошли от общего предка, а остальные были отнесены к конвергентная эволюция. Анализ Серено Herrerasaurus также побудил его предложить несколько новых синапоморфий динозавров.[4]
Классификация

Herrerasaurus изначально считался родом в Карнозаврия, который затем включал формы, похожие на Мегалозавр и Антродем (последнее эквивалентно Аллозавр[40]), хотя Herrerasaurus жили за много миллионов лет до них и сохранили множество примитивный Особенности. Эта классификация карнозавров была дополнена Рождественским и Татариновым в 1964 году, которые классифицировали Herrerasaurus в семье Gryponichidae внутри Carnosauria. В том же году Уокер опубликовал иное мнение, что Herrerasaurus вместо этого был в союзе с Платеозавры, хотя он отличался лобковым ботинком. Уокер также предложил Herrerasaurus вместо этого может быть близко к Попозавр (теперь считается псевдозухий[41]) и безымянный теропод из Докум Групп из Техас (теперь закреплен за Rauisuchian Постозух[42]). В 1985 году Чариг отметил, что Herrerasaurus был неопределенной классификации, показывая сходство с обоими "прозауроподы "и" карнозавры ". Romer (1966) просто отметил, что Herrerasaurus был прозауроподом, возможно, в составе Plateosauridae. В описании Стаурикозавр, Кольбер отметил, что между его таксоном и Herrerasaurus, но классифицировали их в отдельные семейства, с Herrerasaurus в Тератозавриды. В 1970 году Бонапарт также предложил сходство между Herrerasaurus и Стаурикозавр, и четко классифицируя их как в Заурисхия, он заявил, что они выглядели так, как будто они не могли быть помещены в нынешнюю семью. Это было далее поддержано Бенедетто в 1973 году, который назвал в честь таксонов новое семейство Herrerasauridae, которые он классифицировал как заурисхий, возможно, в пределах Тероподы но не в Зауроподоморфа.[43] Однако в 1977 году Гальтон предположил, что Herrerasauridae включают только Herrerasaurus, и обнаружил, что это заурисчианский incertae sedis.[44]
Предложено в 1987 году Бринкманом и Сьюз, Herrerasaurus время от времени считалось базовым для Орнитишия и заурисхию, хотя Бринкманн и Сьюс все еще считали, что она находится внутри Динозаврия. Они поддержали это на том основании, что Herrerasaurus имеет большой палец на педали V и хорошо развитую медиальную стенку вертлужной впадины. Бринкманн и Сьюс считали Стаурикозавр и Herrerasaurus чтобы не образовать настоящую группу под названием Herrerasauridae, и что вместо этого они были последовательно более примитивными формами. Кроме того, они посчитали символы, используемые Бенедетто, недействительными, вместо этого представляя только плезиоморфии состояние, которое было обнаружено в обоих таксонах.[22] В 1992 г. с этим не согласился Новас, заявивший, что многие производные синапоморфии Herrerasauridae, такие как отдельный лобковый ботинок, но все же классифицировали их как базальные к Ornithischia и Saurischia. Новас определил семью как наименее распространенных предков Herrerasaurus и Стаурикозавр и все его потомки.[23] Другое определение Herrerasaurus но нет Прохожий домашний впервые предложен Серено (1998), и более точно следует первоначальному включению, предложенному Бенедетто.[45] Другая группа, Herrerasauria был назван Гальтоном в 1985 году и определен как Herrerasaurus но нет Liliensternus или же Платеозавр Лангером (2004), который использовал определение Herrerasauridae на основе узлов.[46]

В пересмотре основной динозаврии Padian и May (1993) обсудили определение клады и переопределили ее как последнего общего предка Трицератопс и птицы. Они также обсудили, что это определение повлияет на большинство базальных таксонов, таких как Herrerasauridae и Эораптор. Падиан и Мэй считали, что, поскольку и Herrerasauridae, и Эораптор у них отсутствуют многие диагностические признаки Saurischia или Ornithischia, чтобы их нельзя было рассматривать в составе Dinosauria.[47]
Более позднее исследование Новаса 1994 г. Herrerasaurus внутри Dinosauria, и решительно поддерживал свою позицию в Saurischia, а также предоставил синапоморфии, которые он разделял с Theropoda. Новас обнаружил, что примитивные черты отсутствия короткой ямки и наличия только двух крестцовых позвонков были просто обратными чертами, обнаруженными в этом роду.[48] В 1996 году Новас пошел дальше, поддержав позицию тероподов для Herrerasaurus с филогенетический анализ, который поместил его ближе к Neotheropoda чем Эораптор или зауроподоморфа.[49] Лангер (2004) упомянул, что эта гипотеза получила широкое признание, но более поздние авторы вместо этого предпочли поместить Herrerasaurus а также Эораптор базальна к Theropoda и Sauropodomorpha, кладе под названием Euзаурисхия. Лангер (2004) провел филогенетический анализ и обнаружил, что гораздо более вероятно, что Herrerasaurus был базальным заурисхом, чем теропод или нединозавр.[46] Предложение Лангера было подтверждено многочисленными исследованиями до открытия Тава, когда Несбитт и другие. провели более всеобъемлющий анализ, и в результате кладограмма поместил Herrerasauridae базально на Эораптор, но ближе к Дилофозавр чем зауроподоморфа.[50][51] В отличие от Несбитта, Эскурра (2010) провел филогенетический анализ, чтобы разместить свой новый таксон Хромогизавр, и обнаружил, что Herrerasauridae был базальнее Eusaurischia.[52]
В 2010 году Алькосер и Мартинес описали новый таксон герреразаврид, Санджуанзавр. Его можно было отличить от Herrerasaurus на основе нескольких функций. В филогенетическом анализе Herrerasaurus, Санджуанзавр и Стаурикозавр все были в политомия, а Herrerasauridae были самой примитивной группой заурисхий за пределами Евзаурисхии, Эораптор и Гуайбазавр.[1] В 2011 году Мартинес и другие. описанный Эодромей, базальный теропод из той же свиты, что и Herrerasaurus. В филогенетическом анализе Эораптор был помещен в Sauropodomorpha, Herrerasauridae был помещен как самые базальные теропод, и Эодромей был помещен в качестве следующего наиболее базального.[53] Более свежий анализ Bittencourt и другие. (2014) поместили Herrerasauridae в политомию с Theropoda и Sauropodomorpha, с Эораптор также находясь в нерешенном положении. Эта кладограмма представлена ниже.[54]

| Динозаврия |
| |||||||||||||||||||||||||||||||||||||||
Другие члены клады может включать Эораптор из того же Формация Исчигуаласто Аргентины как Herrerasaurus,[55] Чиндезавр с Верхнего Окаменелый лес (Формирование Чинле ) Аризоны,[56] и возможно Казеозавр от Формация Тековас из Докум Групп в Техасе,[57] хотя родство этих животных до конца не изучено, и не все палеонтологи соглашаются. Другие возможные базальные тероподы, Alwalkeria из позднего триаса Формация Малери из Индия,[58] и Тейувасу, известные по очень фрагментарным останкам позднего триаса Бразилии, могут быть связаны.[59] Пол (1988) отметил, что было ошибочно предположено, что Стаурикозавр цени был несовершеннолетним Herrerasaurus. Это утверждение было опровергнуто, когда кости таза несовершеннолетнего Herrerasaurus были обнаружены, которые при осмотре не походили на кости таза Стаурикозавр.[8]
Палеобиология

Зубы Herrerasaurus указать, что это был плотоядное животное; его размер указывает на то, что он охотился на мелких и средних поедателей растений. Они могли включать других динозавров, таких как Пизанозавр, а также более многочисленные ринхозавры и синапсиды.[60] Herrerasaurus сам мог стать жертвой гигантского Rauisuchids подобно Saurosuchus; На одном черепе обнаружены колотые раны.[11]
Копролиты (окаменевший навоз), содержащий мелкие кости, но не содержащий следов фрагментов растений, обнаруженный в формации Исчигуаласто, был отнесен к Herrerasaurus основано на изобилии окаменелостей. Минералого-химический анализ этих копролитов свидетельствует о том, что при отнесении к Herrerasaurus был прав, это плотоядное животное могло переваривать кости.[61]

Сравнения между склеральные кольца из Herrerasaurus и современные птицы и рептилии предполагают, что это могло быть катемерный, активен в течение дня с короткими интервалами.[62]
В исследовании 2001 года, проведенном Брюсом Ротшильдом и другими палеонтологами, 12 костей рук и 20 костей стопы относились к Herrerasaurus были исследованы на наличие признаков стресс-перелом, но ничего не было найдено.[63]
ПВСЖ 407, а Herrerasaurus ischigualastensis, имел яму в кости черепа, приписываемую Пол Серено и Новас перекусить. Две дополнительные ямы образовались на блестящий. Области вокруг этих ям набухшие и пористые, что позволяет предположить, что раны были поражены кратковременной нелетальной инфекцией. Из-за размера и угла раны вполне вероятно, что они были получены в драке с другим Herrerasaurus.[64]
Палеоэкология

В голотип из Herrerasaurus (PVL 2566) был обнаружен в члене Канча-де-Бочас Формация Исчигуаласто в Сан-Хуане, Аргентина. Он был собран в 1961 году Викторино Эррерой в отложениях, которые были отложены в Карнийский этап Триасовый период, примерно 235–231 миллион лет назад. С годами формация Исчигуаласто произвела другие окаменелости, в конечном итоге Herrerasaurus. В 1958 году А.С. Ромер обнаружил образец MCZ 7063, первоначально называвшийся Стаурикозавр в карнийских отложениях. Herrerasaurus образцы PVL 2045 и MLP (4) 61, были собраны в 1959 и 1960 годах соответственно в отложениях, которые откладывались в Norian этап триасового периода, примерно 228–208 миллионов лет назад. В 1960 году Скаглиа собрал экземпляр MACN 18.060, первоначально голотип Ischisaurus cattoi, в отложениях карнийского яруса. В 1961 году Скалья собрал Herrerasaurus экз. ПВЛ 2558, в карнийских отложениях этой свиты. В 1990 году участник Cancha de Bochas произвел еще Herrerasaurus образцы, также из карнийских пластов.[65] Экземпляр PVSJ 53, первоначально голотип Frenguellisaurus, был собран Gargiulo & Oñate в 1975 году в отложениях, отложившихся на карнийском этапе.[9]
Несмотря на то что Herrerasaurus имел форму тела крупных хищных динозавров, он жил в те времена, когда динозавры были маленькими и незначительными. Это было время рептилий нединозавров, а не динозавров, и важный поворотный момент в экологии Земли. Фауна позвоночных свиты исчигуаласто и несколько позже Формация Лос-Колорадос состояла в основном из множества голеностопный архозавры и синапсиды.[66][67] В формации Исчигуаласто динозавры составляли лишь около 6% от общего числа окаменелостей,[68] но к концу триасового периода динозавры стали доминирующими крупными наземными животными, а количество и разнообразие других архозавров и синапсидов уменьшилось.[69]
Исследования показывают, что палеосреда формации Исчигуаласто была вулканически активным пойма покрыта лесами и подвержена сильным сезонным дождям. Климат был влажным и теплым,[70] хотя и подвержен сезонным колебаниям.[71] Растительность состояла из папоротники (Cladophlebis ), хвощи, и гигант хвойные породы (Протоюнипроксилон).[67] Эти растения образовали низинные леса по берегам рек.[4] Herrerasaurus Останки, по-видимому, были наиболее распространенными среди хищников формации Исчигуаласто.[19] Он жил в джунглях позднего триаса. Южная Америка рядом с еще одним ранним динозавром, метровой длиной Эораптор, а также Saurosuchus,[72] гигантское земледелие Rauisuchian (четвероногий мясоед с теропод -подобный череп); в целом похожий, но меньший Venaticosuchus, орнитосухид; и хищный терапсид хиникводонтиды.[19] Травоядных было гораздо больше, чем хищников, и они были представлены ринхозавры Такие как Гиперодапедон (клювая рептилия); этозавры (колючие панцирные рептилии); и терапсиды, включая Kannemeyeriid дицинодонты (коренастые, четвероногие животные с тяжелым передним клювом), такие как Искигуаластия и траверсодонтид цинодонты (несколько похожи по форме на дицинодонтов, но без клювов), такие как Exaeretodon.
Рекомендации
- ^ а б Олкобер, Оскар А .; Мартинес, Рикардо Н. (2010). «Новый герреразаврид (Dinosauria, Saurischia) из формации Исчигуаласто верхнего триаса на северо-западе Аргентины». ZooKeys (63): 55–81. Дои:10.3897 / zookeys.63.550. ЧВК 3088398. PMID 21594020.
- ^ а б c d Рейг, О.А. (1963). "La presencia de dinosaurios saurisquios en los" Estratos de Ischigualasto (Mesotriásico Superior) de las provincias de San Juan y La Rioja (República Argentina) ". Амегиниана (на испанском). 3 (1): 3–20.
- ^ Sereno, P.C .; Novas, F.E .; Arcucci, A.B .; К. Ю (1988). «Новые свидетельства происхождения динозавров и млекопитающих из формации Исчигуаласто (верхний триас, Аргентина)». Журнал палеонтологии позвоночных. 8 (3, приложение): 26А. Дои:10.1080/02724634.1988.10011734.
- ^ а б c d е ж Sereno, P.C .; Новас, Ф.Э. (1992). «Полный череп и скелет раннего динозавра». Наука. 258 (5085): 1137–1140. Bibcode:1992Научный ... 258.1137S. Дои:10.1126 / science.258.5085.1137. PMID 17789086. S2CID 1640394.
- ^ Хольц, Томас Р. младший (2011) Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов, Зима 2010 Приложение.
- ^ Nesbitt, S.J .; Smith, N.D .; Irmis, R. B .; Тернер, А. Х .; Даунс, А. и Норелл, М. А. (2009). «Полный скелет позднетриасового заурисха и ранняя эволюция динозавров». Наука. 326 (5959): 1530–1533. Bibcode:2009Научный ... 326.1530N. Дои:10.1126 / наука.1180350. PMID 20007898. S2CID 8349110..
- ^ Airhart, Марк (10 декабря 2009 г.). "Новый динозавр-мясоед меняет эволюционное древо". Джексонская школа наук о Земле. Архивировано из оригинал 10 марта 2010 г.
- ^ а б c Пол, Г.С. (1988). Хищные динозавры мира. Нью-Йорк: Саймон и Шустер. стр.248–250. ISBN 978-0-671-68733-5.
- ^ а б c d е Лангер, Макс С. (2004). «Базальная заурисхия». В Weishampel, David B .; Додсон, Питер; Osmólska Halszka (ред.). Динозаврия (2-е изд.). Беркли: Калифорнийский университет Press. С. 25–46. ISBN 978-0-520-24209-8.
- ^ Серено, П. (1993). "Грудной пояс и передняя конечность базального теропода Herrerasaurus ischigualastensis". Журнал палеонтологии позвоночных. 13 (4): 425–450. Дои:10.1080/02724634.1994.10011524.
- ^ а б c d Sereno, P.C .; Новас, Ф.Э. (1993). "Череп и шея базального теропода Herrerasaurus ischigualastensis". Журнал палеонтологии позвоночных. 13 (4): 451–476. Дои:10.1080/02724634.1994.10011525.
- ^ Серено, П. (2007). «Филогенетические отношения ранних динозавров: сравнительный отчет». Историческая биология. 19 (1): 145–155. Дои:10.1080/08912960601167435. S2CID 84270944.
- ^ Макдауэлл, С. Jr .; СМ. Богерт (1954). "Системное положение Лантанотус и родство ящериц-ангуиноморфов ". Бюллетень Американского музея естественной истории. 105: 1–142.
- ^ а б c Новас, Ф. Э. (1994). «Новые сведения о систематике и посткраниальном скелете Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) из формации Исчигуаласто (верхний триас) Аргентины ". Журнал палеонтологии позвоночных. 13 (4): 400–423. Дои:10.1080/02724634.1994.10011523.
- ^ Новас (1993). «Новая информация о систематике и посткраниальном скелете Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) из формации Исчигуаласто (верхний триас) Аргентины». Журнал палеонтологии позвоночных. 13 (4): 400–423. Дои:10.1080/02724634.1994.10011523.
- ^ Серено (1993). «Грудной пояс и передняя конечность базального теропод Herrerasaurus ischigualastensis». Журнал палеонтологии позвоночных. 13 (4): 425–450. Дои:10.1080/02724634.1994.10011524.
- ^ а б Серено; Новас (1993).«Череп и шея базального теропод Herrerasaurus ischigualastensis». Журнал палеонтологии позвоночных. 13 (4): 451–476. Дои:10.1080/02724634.1994.10011525.
- ^ Sereno, P.C .; Форстер, К.А.; Rogers, R.R .; Монетта, А. (1993). «Первобытный скелет динозавра из Аргентины и ранняя эволюция динозавров». Природа. 361 (6407): 64–66. Bibcode:1993Натура 361 ... 64С. Дои:10.1038 / 361064a0. S2CID 4270484.
- ^ а б c Rogers, R. R .; Swisher III, C.C .; Sereno, P.C .; Monetta, A.M .; Forster, C.A .; Мартинес, Р. (1993). «Комплекс четвероногих Исчигуаласто (поздний триас, Аргентина) и датирование происхождения динозавров 40Ar / 39Ar». Наука. 260 (5109): 794–797. Bibcode:1993Наука ... 260..794R. Дои:10.1126 / science.260.5109.794. PMID 17746113. S2CID 35644127.
- ^ Стил Р. (1970). "Часть 14. Заурисхия. Handbuch der Paläoherpetologie / Энциклопедия палеогерпетологии". Густав Фишер Верлаг, Штутгарт: 1–87.
- ^ Гальтон, П. (1973). "Об анатомии и взаимоотношениях Эфразия диагностическая (Huene) n.gen., Прозауроподный динозавр (Reptilia: Saurischia) из верхнего триаса Германии ». Paläontologische Zeitschrift. 47 (3/4): 229–255. Дои:10.1007 / BF02985709. S2CID 84931897.
- ^ а б c Brinkman, D.B .; Сьюз, H.D. (1987). «Стаурикозаврид динозавр из верхнетриасовой формации Исчигуаласто в Аргентине и отношения Staurikosauridae» (PDF). Палеонтология. 30 (3): 493–503. Архивировано из оригинал (PDF) 8 декабря 2015 г.. Получено 20 октября, 2015.
- ^ а б c Новас, Ф.Э. (1992). «Филогенетические отношения основных динозавров, Herrerasauridae» (PDF). Палеонтология. 35 (1): 51–62. Архивировано из оригинал (PDF) 8 декабря 2015 г.. Получено 20 октября, 2015.
- ^ Пол, Г.С. (1984). «Сегнозавры динозавры: остатки прозауроподно-орнитисхиевого перехода?». Журнал палеонтологии позвоночных. 4 (4): 507–515. Дои:10.1080/02724634.1984.10012026.
- ^ Новас, Ф. Э. (1989). «Большеберцовая кость и предплюсна у Herrerasauridae (Dinosauria, incertae sedis), а также эволюция и происхождение предплюсны динозавров». Журнал палеонтологии. 63: 677–690. Дои:10.1017 / S0022336000041317.
- ^ Gauthier, J.A .; Cannatella, D .; Queiroz, K .; Клюге, А.Г. и Роу, Т. (1989). «Филогения четвероногих». В Fernholm, K. Bremer, & Jörnvall, H. (ed.). Иерархия жизни. Амстердам: Издательство Elsevier Science B.V., стр. 337–353.
- ^ Новас, Ф. Э. (1986). "Un Provenable terópodo (Saurischia) de la Formación Ischigualasto (Triásico superior), Сан-Хуан, Аргентина". IV Congreso Argentino de Paleontología y Bioestratigrafía, Мендоса, 23–27 ноября (на испанском). 2: 1–6.
- ^ а б Серено; Новас (1992). «Полный череп и скелет раннего динозавра». Наука. 258 (5085): 1137–1140. Bibcode:1992Научный ... 258.1137S. Дои:10.1126 / science.258.5085.1137. PMID 17789086. S2CID 1640394.
- ^ Хольц, Т. Jr .; Падиан, К. (1995). «Определение и диагностика Theropoda и родственных таксонов». Журнал палеонтологии позвоночных. 15 (3, приложение): 35А.
- ^ Bonaparte, J.F .; Пумарес, Дж. (1995). "Notas sobre el primer cráneo de Риохазавр incertus (Dinosauria, Prosauropoda, Melanorosauridae) del Triásico superior de La Rioja, Аргентина ". Амегиниана (на испанском). 32: 341–349.
- ^ Раухут, О.В.М. (2003). «Взаимоотношения и эволюция базальных динозавров-теропод». Специальные статьи по палеонтологии. 69: 1–213.
- ^ Bittencourt, J .; Келлнер, В.А. (2004). "Филогенетическое положение Стаурикозавр цени из триаса Бразилии ». Журнал палеонтологии позвоночных. 24 (3, приложение): 39А. Дои:10.1080/02724634.2004.10010643. S2CID 220415208.
- ^ Langer, M.C .; Бентон, М.Дж. (2006). «Ранние динозавры: филогенетическое исследование». Журнал систематической палеонтологии. 4 (4): 309–358. Дои:10.1017 / S1477201906001970. S2CID 55723635.
- ^ Irmis, Randall B .; Nesbitt, Sterling J .; Падиан, Кевин; Смит, Натан Д .; Тернер, Алан Х .; Вуди, Дэниел; Даунс, Алекс (2007). «Позднее триасовое скопление динозавроморф из Нью-Мексико и рост динозавров». Наука. 317 (5836): 358–361. Bibcode:2007Наука ... 317..358I. Дои:10.1126 / science.1143325. PMID 17641198. S2CID 6050601.
- ^ White, A.T .; Казлев, М.А. (2003). "Theropoda: Basal Theropods". Наука. 326 (5959): 1530–1533. Bibcode:2009Научный ... 326.1530N. Дои:10.1126 / наука.1180350. PMID 20007898. S2CID 8349110. Архивировано из оригинал 11 июля 2007 г.. Получено 20 июля, 2007.
- ^ Forster, C.A .; Arcucci, A.B .; Marsicano, CA; Abdala, F .; Мэй, К. (1995). «Новый материал позвоночных из формации Лос-Растрос (средний триас), провинция Ла-Риоха, северо-запад Аргентины». Журнал палеонтологии позвоночных. 15 (3, приложение): 29А. Дои:10.1080/02724634.1995.10011277.
- ^ Arcucci, A.C .; Forster, C .; Мэй, Абдала, Ф .; Марсикано, К. (1998). "Una nueva icnofauna de la Formación Los Rastros, Triásico Medio, en la Quebrada de Ischichuca Chica (Провинция де Ла Риоха, Аргентина)". Acta Geológica Lilloana (на испанском). 18: 152.CS1 maint: несколько имен: список авторов (связь)
- ^ Баккер, Р.Т .; Гальтон, П. (1974). «Монофилия динозавров и новый класс позвоночных». Природа. 248 (5444): 168–172. Bibcode:1974Натура.248..168Б. Дои:10.1038 / 248168a0. S2CID 4220935.
- ^ Бонапарт, Дж. Ф. (1976). "Пизанозавр mertii Казимикела и происхождение орнитиский ". Журнал палеонтологии. 50: 808–820.
- ^ Гилмор, Чарльз У. (1920). "Остеология хищных динозавров в Национальном музее США, с особым упором на роды Антродем (Аллозавр) и Цератозавр" (PDF). Бюллетень Национального музея США. 110 (110): 1–159. Дои:10.5479 / si.03629236.110.i. HDL:2027 / uiug.30112032536010.
- ^ Gauthier, J.A .; Несбитт, С.Дж .; Schachner, E.R .; Bever, G.S .; Джойс, W.G. (2011). "Двуногий стволовый крокодил Poposaurus gracilis: определение функции окаменелостей и инновации в перемещении архозавра " (PDF). Бюллетень музея естественной истории Пибоди. 52 (1): 107–126. Дои:10.3374/014.052.0102. S2CID 86687464. Архивировано из оригинал (PDF) 24 сентября 2015 г.
- ^ Чаттерджи, Санкар (1985). "Постозух, новая текодонтианская рептилия из триаса Техаса и происхождение тираннозавров ». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 309 (1139): 395–460. Bibcode:1985RSPTB.309..395C. Дои:10.1098 / рстб.1985.0092.
- ^ Бенедетто, Дж. Л. (1973). "Herrerasauridae, nueva familia de saurisquios triasicos" (PDF). Амегиниана. 10 (1): 89–102.
- ^ Гальтон, П. (1977). "На Staurikosaums pricei, ранний заурисхий динозавр из триаса Бразилии, с примечаниями о Herrerasauridae и Poposauridae ". Paläontologische Zeitschrift. 51 (3): 234–245. Дои:10.1007 / BF02986571. ISSN 0031-0220. S2CID 129438498.
- ^ Серено, П. (1998). «Рационал для филогенетических определений в применении к таксономии динозавров более высокого уровня». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 210 (1): 41–83. Дои:10.1127 / njgpa / 210/1998/41.
- ^ а б Лангер, М. (2004). «Базальная заурисхия». В Weishampel, David B .; Осмольска, Хальска; Додсон, Питер (ред.). Динозаврия (2-е изд.). Издательство Индианского университета. С. 40–42. ISBN 978-0-520-25408-4.
- ^ Padian, K .; Мэй, К. (1993). «Древнейшие динозавры». В Lucas, Spencer G .; Моралес, М. (ред.). Неморский триас. Бюллетень Музея естественной истории и науки Нью-Мексико. 3. С. 379–381.
- ^ Новас, Ф.Э. (1994). «Новые сведения о систематике и посткраниальном скелете Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) из формации Исчигуаласто (верхний триас) Аргентины ". Журнал палеонтологии позвоночных. 34 (4): 400–423. Дои:10.1080/02724634.1994.10011523.
- ^ Новас, Ф.Э. (1996). «Монофилия динозавров». Журнал палеонтологии позвоночных. 16 (4): 723–741. Дои:10.1080/02724634.1996.10011361. JSTOR 4523770.
- ^ Langer, M.C .; Ezcurra, MD; Bittencourt, J.S .; Новас, Ф.Э. (2010). «Происхождение и ранняя эволюция динозавров» (PDF). Биологические обзоры. 85 (1): 55–110. Дои:10.1111 / j.1469-185X.2009.00094.x. PMID 19895605. S2CID 34530296.
- ^ Несбитт, С.Дж .; Smith, N.D .; Irmis, R.B .; Тернер, A.H .; Даунс, А .; Норелл, М.А. (2009). «Полный скелет позднетриасового заурисхия и ранняя эволюция динозавров». Наука. 326 (5959): 1530–1533. Bibcode:2009Научный ... 326.1530N. Дои:10.1126 / наука.1180350. ISSN 0036-8075. PMID 20007898. S2CID 8349110.
- ^ Эскурра, доктор медицины (2010). «Новый ранний динозавр (Saurischia: Sauropodomorpha) из позднего триаса Аргентины: переоценка происхождения и филогении динозавров». Журнал систематической палеонтологии. 8 (3): 371–425. Дои:10.1080/14772019.2010.484650. S2CID 129244872.
- ^ Martinez, R.N .; Sereno, P.C .; Alcober, O.A .; Colombi, C.E .; Renne, P.A .; Montañez, I.P .; Карри, Б.С. (2011). "Базальный динозавр на заре эры динозавров в Юго-Западной Пангеи". Наука. 331 (6014): 206–210. Bibcode:2011Наука ... 331..206М. Дои:10.1126 / science.1198467. ISSN 0036-8075. PMID 21233386. S2CID 33506648.
- ^ Bittencourt, J.S .; Arcucci, A.B .; Marsicano, CA; Лангер, М. (2014). «Остеология среднетриасового архозавра. Lewisuchus admixtus Ромера (формация Чаньярес, Аргентина), его инклюзивность и отношения между ранними динозавроморфами » (PDF). Журнал систематической палеонтологии. 0 (3): 189–219. Дои:10.1080/14772019.2013.878758. S2CID 56155733.
- ^ Кольбер, Э. (1970). «Заурисхий динозавр из триаса Бразилии». Американский музей Novitates. 2405: 1–39.
- ^ Long, R.A .; Мерри, П.А. (1995). «Позднетриасовые (карнийские и норийские) четвероногие животные из Юго-Запада США». Музей естественной истории и науки Нью-Мексико, бюллетень. 4: 1–254.
- ^ Хант, А.П .; Lucas, S.G .; Heckert, A.B .; Sullivan, R.M .; Локли, М. (1998). «Динозавры позднего триаса из Западной части Соединенных Штатов». Geobios. 31 (4): 511–531. Дои:10.1016 / S0016-6995 (98) 80123-X.
- ^ Chatterjee, S .; Крейслер, Б.С. (1994). "Alwalkeria (Theropoda) и Morturneria (Плезиозаврия), новые имена для озабоченных Walkeria Чаттерджи, 1987 и Turneria Чаттерджи и Смолл, 1989 ». Журнал палеонтологии позвоночных. 14 (1): 142. Дои:10.1080/02724634.1994.10011546.
- ^ Кишлат, Э.-Э. (1999). «Спасение» нового динозавра из бразильского триаса: Teyuwasu barberenai, новый таксон ". Paleontologia Em Destaque, Boletim Informativo da Sociedade Brasileira de Paleontologia. 14 (26): 58.
- ^ Сьюз, H.D. (1990). "Стаурикозавр и Herrerasauridae ". In Weishampel, D.B .; Dodson, P .; Osmólska, H. (eds.). Динозаврия. Калифорнийский университет Press. С. 143–47. ISBN 978-0-520-06726-4.
- ^ Hollocher, K.T .; Alcober, O.A .; Colombi, C.E .; Холлохер, Т. (2005). «Копролиты плотоядных животных из формации Исчигуаласто в верхнем триасе, Аргентина: химия, минералогия и свидетельства быстрой начальной минерализации». ПАЛАИ. 20 (1): 51–63. Bibcode:2005Палай..20 ... 51ч. Дои:10.2110 / palo.2003.p03-98. S2CID 131242248.
- ^ Schmitz, L .; Мотани, Р. (2011). «Ночной образ жизни у динозавров на основании морфологии склерального кольца и орбиты». Наука. 332 (6030): 705–8. Bibcode:2011Наука ... 332..705С. Дои:10.1126 / science.1200043. PMID 21493820. S2CID 33253407.
- ^ Ротшильд Б., Танке Д. Х. и Форд Т. Л., 2001, Стрессовые переломы теропод и отрывы сухожилий как ключ к активности: В: Мезозойская жизнь позвоночных, отредактированный Танке Д. Х. и Карпентером К., Indiana University Press, стр. 331–336.
- ^ Мольнар Р. Э., 2001, Палеопатология теропод: обзор литературы: В: Мезозойская жизнь позвоночных, отредактированный Танке Д. Х. и Карпентером К., Indiana University Press, стр. 337–363.
- ^ Sereno, P.C .; Forster, C.A .; Rogers, R. R .; Монетта, А. М. (1993). «Первобытный скелет динозавра из Аргентины и ранняя эволюция динозавров». Природа. 361 (6407): 64–66. Bibcode:1993Натура 361 ... 64С. Дои:10.1038 / 361064a0. S2CID 4270484.
- ^ Бонапарт, Дж. Ф. (1982). «Замена фауны в триасе Южной Америки». Журнал палеонтологии позвоночных. 2 (3): 362–371. Дои:10.1080/02724634.1982.10011938.
- ^ а б Бонапарт, Дж. Ф. (1979). "Фауны и палеобиогеография тетраподос мезозойских Америки". Ameghiniana, Revista de la Asociación Paleontológica Argentina (на испанском). 16 (3–4): 217–238.
- ^ Бентон, Майкл Дж. (1999). «Происхождение и ранняя эволюция динозавров». В Фарлоу, Джеймс О .; Бретт-Сурман, М. (ред.). Полный динозавр. Издательство Индианского университета. С. 204–215. ISBN 978-0-253-21313-6.
- ^ Пэрриш, Дж. Майкл (1999). «Эволюция архозавров». В Фарлоу, Джеймс О .; Бретт-Сурман, М. (ред.). Полный динозавр. Издательство Индианского университета. С. 191–203. ISBN 978-0-253-21313-6.
- ^ Такер, Морис Э.; Бентон, Майкл Дж. (1982). «Триасовая среда, климат и эволюция рептилий» (PDF). Палеогеография, палеоклиматология, палеоэкология. 40 (4): 361–379. Bibcode:1982ППП .... 40..361Т. Дои:10.1016/0031-0182(82)90034-7. Архивировано из оригинал (PDF) 26 января 2009 г.. Получено 23 июля, 2009.
- ^ Колумби, Карина Э. (5 октября 2008 г.). Анализ стабильных изотопов ископаемых растений формации Исчигуаласто верхнего триаса на северо-западе Аргентины. Хьюстон, Техас: Геологическое общество Америки. Архивировано из оригинал 11 января 2012 г.. Получено 23 июля, 2009.
- ^ Подоконник, W.D. (1974). "Анатомия Saurosuchus galilei и взаимоотношения рауизухид-текодонтов ". Вестник Музея сравнительной зоологии. 146: 317–362.
внешняя ссылка
 Wikijunior Динозавры / Herrerasaurus в Викиучебнике
Wikijunior Динозавры / Herrerasaurus в Викиучебнике СМИ, связанные с Herrerasaurus в Wikimedia Commons
СМИ, связанные с Herrerasaurus в Wikimedia Commons Данные, относящиеся к Herrerasaurus в Wikispecies
Данные, относящиеся к Herrerasaurus в Wikispecies- Введение в Herrerasaurus, из Музея палеонтологии Калифорнийского университета









