Эволюция динозавров - Evolution of dinosaurs
- В этой статье дается схема и примеры эволюции динозавров. Подробный список взаимосвязей см. Классификация динозавров.
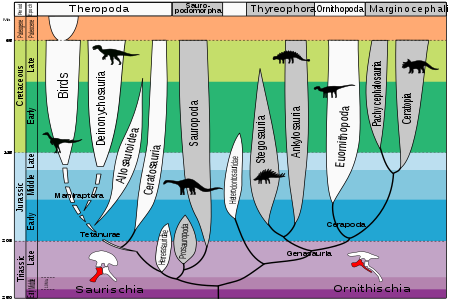
Динозавры развились с единой линией архозавры 243-233 млн лет назад Анисианский к Карнийский возрастов, последняя часть середины Триасовый. Динозаврия хорошо поддерживается клады, присутствует в 98% бутстрепы. Он диагностируется по многим признакам, включая потерю постфронтальной костей черепа и удлиненный дельтопекторальный гребень на черепе. плечевая кость.[1]
В марте 2017 года ученые сообщили о новом способе классификации динозавр генеалогическое древо, основанное на новых и большем количестве данных, чем было доступно ранее Согласно новой классификации, первоначальные динозавры, появившиеся 200 миллионов лет назад, были маленькими, двуногими. всеядный животные с большими хватательными руками. Потомки (для нептичьи динозавры ) длилась до 66 миллионов лет назад.[2][3]
Происхождение среди архозавров
Процесс, ведущий к Динозавроморфа и первых настоящих динозавров можно проследить по окаменелостям ранних Архозавры такой как Proterosuchidae, Erythrosuchidae и Euparkeria окаменелости которых датируются 250 млн лет назад, архозаврами середины триаса, такими как Ticinosuchus 232–236 млн лет. Крокодилы также являются потомками архозавров среднего триаса.[1]
Динозавров можно определить как последнего общего предка птиц (Заурисхия ) и Трицератопс (Орнитишия ) и всех потомков этого предка. С таким определением птерозавры и несколько видов архозавров не попадают в категорию динозавров. Архозавр роды которые также не попадают в категорию динозавров, включая Schleromochlus 220-225 млн лет, Lagerpeton 230-232 млн лет и Марасух 230-232 млн лет.
Древнейшие динозавры
Первые известные динозавры были двуногий хищники которые были 1-2 метра (3,3-6,5 футов) в длину.
Самые ранние подтвержденные окаменелости динозавров включают заурисхий ('ящерицы') динозавры Ньясазавр 243 млн лет,[1] Сатурналии 225-232 млн лет, Herrerasaurus 220-230 млн лет, Стаурикозавр возможно 225-230 млн лет, Эораптор 220-230 млн лет и Alwalkeria 220-230 млн лет. Сатурналии может быть базальным заурисханом или прозауроподом. Остальные - базальные заурисхии.
Среди самых ранних орнитисхий ("птичьи бедра") динозавры - это Пизанозавр 220-230 млн лет. Несмотря на то что Лесотозавр датируется 195-206 млн лет назад, особенности скелета позволяют предположить, что он ответвился от основной линии орнитиший, по крайней мере, еще в Пизанозавр.

Из этого рисунка видно, что ранние заурисхи были похожи на ранних орнитисхов, но не на современных крокодилов. Заурисхи отличаются от орнитисхий тем, что сохраняют изначальную конфигурацию костей таза. Еще одно отличие состоит в черепе, верхний череп орнитиший более прочный, а сустав, соединяющий нижнюю челюсть, более гибкий; оба являются адаптацией к травоядным, и оба уже можно увидеть в Лесотозавр.
Заурисхия
Не считая базальных заурисхий, остальные заурисхии разделяются на Зауроподоморфа и Тероподы.[4] Sauropodomorpha делится на Prosauropoda и Sauropoda. Эволюционные пути теропод очень сложны. Динозаврия (2004), крупный справочник по динозаврам,[1] разделяет Theropoda на группы Ceratosauria, Basal Tetanurae, Tyrannosauroida, Ornithomimosauria, Therizinosauroida, Oviraptorosauria, Troodontidae, Dromaeosauridae и Basal Avialae по очереди. Позже каждая группа ответвляется от основной магистрали. Видеть Классификация динозавров для подробной взаимосвязи между ними.
Зауроподоморфа
Первыми зауроподоморфами были прозауроподы. Окаменелости просауроподов известны с конца триаса до начала юры 227–180 млн лет.[1] Они могли быть двуногими или четвероногими, имели длинные шеи и хвосты и относительно небольшие головы. Они имели длину от 2,5 (8,2 фута) до 10 м (33 фута) и были в основном травоядными. Самые ранние прозауроподы, такие как Текодонтозавр от 205–220 млн лет назад, все еще сохранял изначальную двуногую стойку и большое соотношение головы и тела.
Они превратились в зауроподов, которые превратились в гигантских четвероногих травоядных, некоторые из которых достигли длины не менее 26 м (85 футов). Особенности, определяющие эту кладу, включают отношение длины передних конечностей к длине задних конечностей более 0,6. У большинства зауроподов все еще были задние конечности больше, чем передние; одно заметное исключение Брахиозавр чьи длинные передние конечности позволяют предположить, что он эволюционировал, чтобы питаться с высоких деревьев, как современный жираф.
Окаменелости зауроподов находят со времен самых ранних динозавров вплоть до Меловое – палеогеновое вымирание, от 227 до 66 млн лет. Большинство зауроподов известны из Юрский, а точнее между 227 и 121 млн лет назад.
В Меловой зауроподы образуют две группы. В Diplodocoidea жил от 121 до 66 млн лет. В Титанозаврообразные жил от 132 до 66 млн лет. Последняя клада состоит из серии вложенных подгрупп, Титанозаврия, то Titanosauridae и Saltasauridae. И Diplodocoidea, и Titanosauriformes произошли от Neosauropoda, самый ранний из которых жил около 169 млн лет назад.
Зауроподы известны тем, что были самыми крупными наземными животными из когда-либо существовавших, и имели относительно небольшие черепа. Увеличение размеров динозавров зауроподов и зауроподов до этих гигантов и изменение длины черепа показано на следующих диаграммах.
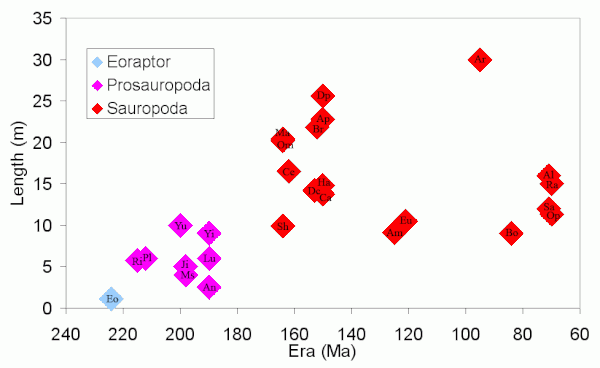

При создании этих диаграмм использовались динозавры (в порядке дат): Эо Эораптор; Прозавроподы Ri Риохазавр, Pl Платеозавр, Ю Юннанозавр, РС Массоспондил, Джи Цзиншанозавр, An Анхизавр, Лу Луфенгозавр, Yi Йименозавр; и зауроподы Sh Шунозавр, Ом Омейзавр, Мм Маменчизавр, Ce Цетиозавр, Округ Колумбия Дикреозавр, Br Брахиозавр, Европа Euhelopus, Ap Апатозавр, Ca Камаразавр, Dp Диплодок,[1] Ха Гаплокантозавр, Являюсь Амаргазавр, Ar Аргентинозавр (приблизительно), Бо Bonitasaura, Q Quaesitosaurus, Al Аламозавр, Сб Сальтазавр, Ра Рапетозавр, Op Опистоцеликаудия, Ne Немегтозавр.
За исключением Аргентинозавр (включены, чтобы заполнить временной промежуток), эти графики показывают только длину зауроподов, для которых известны почти полные ископаемые скелеты. Он не показывает других очень крупных зауроподов (см. Размер динозавров # зауроподы ), потому что они известны только по очень неполным скелетам. Отношение длины черепа к длине тела намного выше у Эораптор чем у зауроподов. Самый длинный череп из представленных на графике Немегтозавр, который не считается особенно большим зауроподом. Череп Немегтозавр был найден возле безголового скелета длиной 11 метров (36 футов). Опистоцеликаудия, и было высказано предположение, что они могут быть одного и того же вида, но см. Nemegtosauridae.
Связь между эволюцией крупных травоядных и крупных растений остается неопределенной. Около 50% растений во времена динозавров были хвойными; они увеличивались в количестве в триасе, пока не стабилизировались примерно к 190 млн лет назад. Цикады составляли вторую по величине группу примерно до 120 млн лет назад. Папоротники все время присутствовали примерно в постоянном количестве. Цветение растений началось около 120 млн лет назад и к концу периода сменилось саговниками. Все травоядные динозавры, по-видимому, пострадали от вымирания в конце юрского периода.
Тероподы
Безусловно, самые ранние окаменелости Theropoda (не считая базальных заурисхий) относятся к Coelophysoidea, включая Целофиз и другие, из позднего триаса и ранней юры 227–180 млн лет.[1] Кладистический анализ иногда связывает их с группой, называемой Цератозаврия. Основные особенности обоих включают изменения в тазовом поясе и задних конечностях, которые различаются между полами. Другие цератозаврии впервые появляются в поздней юре на западе Северной Америки.
За ними следуют базальные Столбняк, окаменелости которых были обнаружены с середины юры до конца раннего мелового периода с 180 до 94 миллионов лет. У них относительно короткий зубной ряд верхней челюсти. Не все они одновременно ответвлялись по эволюционной линии, ведущей к целурозаврам. Базальные столбняки включают: Megalosauridae, спинозавриды, разнообразный клад аллозавры, и несколько родов менее определенного родства, в том числе Компсогнат. За исключением Компсогнат они крупнотелые. Аллозавры образуют отчетливую долгоживущую кладу, у которой есть некоторые черепные признаки. Среди них хорошо известные Аллозавр и Sinraptor среди прочего.
Великое излучение Theropoda на множество различных кладов Целурозаврия должно было произойти в середине и конце юрского периода, потому что Археоптерикс около 152-154 млн лет назад, и кладистический анализ показал, что многие другие группы Целурозаврия перед этим разветвился.[5] Ископаемые остатки из Китая предполагают, что самые ранние перья были обнаружены на примитивной целурозаврии. Самые примитивные из них, например на тираннозавроид Дилонг, были просто волокнами с полой сердцевиной, которые были бы полезны для изоляции, но бесполезны для полета.
Случайные исследования костей и кладистические исследования указывают на то, что тираннозавроиды ответвлялись от других теропод в начале, в среднем юрском периоде, хотя почти полные скелеты еще не появлялись. Эотираннус от 121-127 млн лет назад, и многие близкие родственники Тиранозавр сами по себе не появляются раньше 84 млн лет назад, ближе к концу позднего мелового периода.
Орнитомимозаврия ископаемые известны от 127 до 65 млн лет назад. Считается, что самой ранней ветвью от основной линии орнитомимозавров является Гарпимим.
В Теризинозауроидеа - необычные тероподы, потому что почти все они вегетарианцы. Ископаемые теризинозаавроиды известны от 127 до 65 млн лет назад.
Maniraptorans включают Овирапторозаврия, Дейнонихозавры и птицы. Для них характерна локтевая кость с загнутым стержнем.
Овирапторозавр ископаемые известны от 127 до 65 млн лет назад. У них беззубый череп, который сильно видоизменился. У скелета необычно короткий хвост.
Дейнонихозавры, названные в честь увеличенного серповидного второго пальца стопы, тесно связаны с птицы. У них две разные семьи, Troodontidae и Dromaeosauridae. Ископаемые остатки троодонтид известны от 127 до 65 млн лет назад. У них более стройное телосложение и более длинные конечности. Самая ранняя известная окаменелость троодонтид - Sinornithoides. Окаменелости дромеозаврид известны примерно от 127 до 65 млн лет, за исключением Ютараптор. Остатки скелетов Ютараптор составляют около 127-144 млн лет. Это интересно, потому что согласно недавнему кладистическому анализу,[5] Ютараптор примерно настолько далеко от предковых теропод, насколько это возможно, дальше, чем Археоптерикс. У дромеозавридов второй палец больше; это семейство включает хорошо известных динозавров Дромеозавр, Дейноних и Велоцираптор.
Древние птицы (Avialae ) включают как Авес, которые определены как потомки общего предка современных птиц и Археоптерикс, а тем примитивнее Эпидендрозавр. Ископаемые птицы простираются от 154 млн лет до Меловое – палеогеновое вымирание 65 млн лет до наших дней. Были найдены десятки полных скелетов более поздних Конфуциусорнис, который является одним из первых представителей Орнитура. У всех орнитурцев костлявые пигостиль, к которому прикреплены рулевые перья. Подробнее об эволюции птиц см. Эволюция птиц.
Орнитишия
Орнитишия, как видно из названия, был придуман для птичьего тазового пояса, хотя они не являются предками птиц.
Орнитисхиальный череп и зубной ряд очень рано изменились за счет травоядной диеты.[6] Лесотозавр отделился рано, но череп Лесотозавр уже показывает такие приспособления с широкими пропорциями, менее гибкой верхней челюстью и более подвижным соединением для нижней челюсти.
Heterodontosauridae было показано, что это самая основная группа в пределах орнитиший.[7] Гетеродонтозавриды очень мелкие (длина тела <1 м) и жили с позднего триаса до раннего мела. Помимо Абриктозавр у всех короткий верхний клык и более длинный нижний клык. Передние конечности у известных окаменелостей относительно длинные.
Основные клады были заложены уже в начале юры. Орнитисхианцы делятся на бронированных тиреофораны и без брони орнитоподы и маркоцефалы.
Тиреофораны
Поверхностная бронежилет (щитки ) - самая яркая черта тиреофораны.[6] Скутеллозавр есть они, но в остальном мало отличается от Лесотозавр. У него длинный хвост и комбинированная поза двуногих и четвероногих, что отличает его от всех более поздних тиреофоров, включая Стегозаврия и Анкилозаврия. Эти две клады, хотя и совершенно разные по внешнему виду, имеют много общих черт черепа и скелета.
Стегозавры легко узнать по выступающему ряду пластинок над позвоночником и длинным шипам на хвосте. Большинство стегозавров, но не Стегозавр, также должны иметь позвоночник на каждом плече. Эти шипы и пластины произошли от более ранних поверхностных щитков. Huayangosaurus самый старый и самый примитивный из известных стегозавров.
Анкилозавры легко узнать по их обширной броне. Череп сильно окостенел. В начале своей эволюции анкилозавры разделились на Nodosauridae и Анкилозавриды, отличающийся особенностями черепа.
Орнитопода
Орнитоподы попадают в отдельные клады - Hypsilophodontidae, и Игуанодонтия.[6]
Гипсилофодонтиды больше напоминают своих предков, чем гетеродонтозавриды. Наиболее отличительными чертами являются короткая лопатка и предлобковый отросток палочковидной формы. Самый ранний из них Агилизавр из средней юры Китая.
Игуанодонты - это разнообразный, но морфологически тесно связанный набор родов, известных по окаменелостям в конце мелового периода. К значительным изменениям относятся эволюция зубных батарей, связанная пясть и осанка пальцев рук. Тенонтозавр самый базальный игуанодонт. Другие включают Игуанодон, Камптозавр и Муттабурразавр.
Маргиноцефалия
Маргиноцефалия названы в честь полки, которая выступает над задней частью черепа.[6] Они включают пахицефалозавры и цератопсы.
Пахицефалозавры наиболее известны своей толстой верхней частью черепа. Самый старый из известных - Stenopelix, из раннего мела Европы.
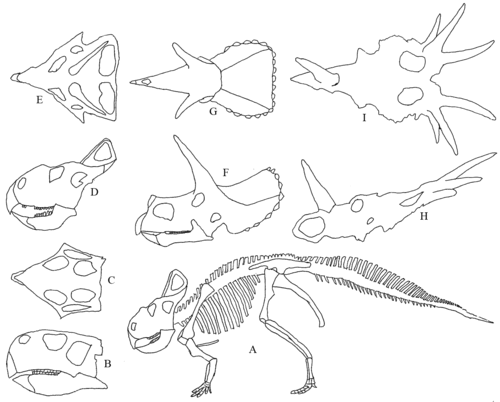
Цератопсы, известен благодаря Протоцератопс, Трицератопс и Стиракозавр иллюстрируют эволюцию черепов с оборками и рогами. Оборки произошли от полочки, общей для всех Marginocephalia. Цератопсы делятся на базальные цератопсы, включая клювов попугая. Пситтакозавр, и неоцератопсы.
Разнообразие цератопсий черепа. А) Скелет Протоцератопс. Б) к I) Черепа. ДО Н.Э) Пситтакозавр сбоку и сверху. D) и E) Протоцератопс сбоку и сверху. F) и G) Трицератопс сбоку и сверху. ЗДРАВСТВУЙ) Стиракозавр сбоку (без нижней челюсти) и сверху.
Эволюция цератопсидных динозавров имеет общие черты с эволюцией некоторых групп млекопитающих, оба были «геологически короткими» событиями, ускоряющими одновременную эволюцию большого размера тела, производных структур питания и «разнообразных роговидных органов».[8]
Последовательность эволюции цератопсов в меловом периоде примерно от Пситтакозавр (121-99 млн лет) до Протоцератопс (83 млн лет) до (Трицератопс 67 млн лет и Стиракозавр 72 млн лет). При виде сбоку череп Пситтакозавр очень мало похож на Стиракозавр но на виде сверху можно увидеть аналогичное пятиугольное расположение.
Окаменелости
Первые несколько линий примитивных динозавров быстро изменились. Триасовый период; виды динозавров быстро развили специальные особенности и диапазон размеров, необходимые для эксплуатации почти всех наземных экологическая ниша. В период преобладания динозавров, охвативший последующие Юрский и Меловой периоды, почти все известные наземные животные более 1 метра в длину были динозаврами.
Одним из показателей качества летописи окаменелостей является сравнение даты первого появления с порядком ветвления кладограмма на основе формы ископаемых элементов. Существует тесная переписка между орнитизиками, заурисхами и подгруппами.[6] Связь кладограмм между целофизиды и кератозавры является исключением, так как это привело бы к слишком позднему происхождению целофизид. Самое простое объяснение - конвергентная эволюция: кости цератозавров независимо развились и приобрели форму, напоминающую форму более ранних целофизид. Другая возможность заключается в том, что цератозавры эволюционировали намного раньше, чем предполагает летопись окаменелостей.
Большинство окаменелостей динозавров было найдено в Norian -Синемурийский, Кимериджский -Титонский, и Кампанский -Маастрихтский периоды. Непрерывность родословных через промежуточные промежутки показывает, что эти промежутки являются артефактами сохранения, а не сокращением разнообразия или изобилия.
Во многих случаях кладистический анализ показывает, что наследственные линии разной продолжительности попадают в эти пробелы. Продолжительность отсутствующих наследственных линий в 1997 г.[6] диапазон от 25 млн лет (Лесотозавр, Genasauria, Hadrosauroida, Зауропода, Неоцератопсия, Целурозаврия ) до 85 млн лет (Carcharodontosauridae ). Поскольку излучение динозавров началось с малых размеров тела, незарегистрированная ранняя история может быть связана с менее надежной окаменелостью более мелких видов. Однако некоторые недостающие линии передачи, особенно Carcharodontosauridae и Abelisauridae, требуют альтернативных объяснений, поскольку отсутствующий диапазон распространяется на стадии, богатые ископаемыми материалами.
Эволюционные тенденции
Размер тела
Размер тела важен из-за его корреляции с метаболизмом, диетой, историей жизни, географическим ареалом и скоростью исчезновения.[6] Модальная масса тела динозавров составляет от 1 до 10 тонн в мезозое и во всех основных континентальных регионах. Наблюдалась тенденция к увеличению размеров тела во многих кладах динозавров, включая Thyreophora, Ornithopoda, Pachycephalosauria, Ceratopsia, Sauropomorpha и базальные тероподы. Заметное уменьшение размера тела также произошло в некоторых линиях, но носит более спорадический характер. Самый известный пример - уменьшение размера тела до появления первых птиц; Археоптерикс был ниже 10 кг веса, а позже птицы Конфуциусорнис и Sinornis размером со скворца до голубя. Это произошло для облегчения полета.
Мобильность
Предки динозавра были двуногими.[9] Эволюция осанки на четвероногих произошла четыре раза, у предков Euornithopoda, Тиреофора, Цератопсия и Зауроподоморфа.[6] Во всех четырех случаях это было связано с увеличением размера тела, и во всех четырех случаях тенденция является однонаправленной, без разворота.
У динозавров наблюдается картина сокращения и потери пальцев на боковой стороне кисти (пальцы III, IV и V). Примитивная функция руки динозавра - хватать частично противоположным большим пальцем, а не удерживать вес. Уменьшение цифр - одна из определяющих черт тираннозавриды с двумя функциональными пальцами на очень коротких передних конечностях.
Влияние источников питания
Древний динозавр был хищником. Травоядность среди динозавров возникала трижды, у истоков клад орнитисхиев, зауроподоморфов и теризинозаврид. Отдельные теризинозавриды растительноядны или всеядны. Травоядность орнитисхийцев и зауроподоморфов никогда не менялась.[6]
Потенциальная совместная эволюция растений и травоядных динозавров была предметом обширных предположений. Появление прозауропод в конце триаса было предположительно связано либо с исчезновением, либо с диверсификацией типов флоры того времени. Подъем цератопсиды и игуанодонт и гадрозаврид орнитоподы в меловом периоде были предварительно связаны с радиацией покрытосеменных. К сожалению, до сих пор нет достоверных данных о диетических предпочтениях травоядных динозавров, кроме данных о технике жевания и гастролиты.
Биогеография
Фауна динозавров, которая была относительно однородной по своему характеру, когда Пангея начал распадаться, стал заметно дифференцироваться к концу мелового периода. Биогеография основан на разделении древних видов путем создания географического барьера. Интерпретация ограничена отсутствием свидетельств окаменелостей восточной части Северной Америки, Мадагаскара, Индии, Антарктиды и Австралии. Не было получено однозначных доказательств биогеографического воздействия на виды динозавров,[6] но некоторые авторы обозначили центры происхождения многих групп динозавров, множественные маршруты расселения и интервалы географической изоляции.
Динозавры, которые были приведены в качестве доказательства биогеографии, включают тероподов абелизаврид из Южной Америки и, возможно, из других мест. Гондвана.
Взаимоотношения между динозаврами демонстрируют множество доказательств их расселения из одного региона земного шара в другой. Тероподы-тетанураны широко путешествовали по западу Северной Америки, Азии, Южной Америке, Африке и Антарктиде. Пахицефалозавры и цератопсы демонстрируют явное свидетельство множественных событий двунаправленной дисперсии в Берингия.
Вымирание
Хотя некоторые ученые утверждают, что массивные извержения вулканов могли стать причиной исчезновения нептичьих динозавров,[10] текущий консенсус - это Меловое – палеогеновое вымирание, который произошел 66 миллионов лет назад в конце мелового периода, вызвал вымирание всех динозавров, кроме той линии, которая уже дала начало первым птицы.[11][12]
Смотрите также
Сноски
- ^ а б c d е ж грамм Вайшампель, Додсон и Осмольска, 2004, Динозаврия
- ^ Уэйд, Николас (22 марта 2017 г.). "Встряхивая генеалогическое древо динозавров". Нью-Йорк Таймс. Получено 22 марта 2017.
- ^ Барон, Мэтью Дж .; Норман, Дэвид Б.; Барретт, Пол М. (22 марта 2017 г.). «Новая гипотеза взаимоотношений динозавров и ранней эволюции динозавров». Природа. 543 (7646): 501–506. Дои:10.1038 / природа21700. PMID 28332513. S2CID 205254710.
- ^ Дэйв Смит. "Заурисханские динозавры". UCMP. Получено 31 июля 2011.
- ^ а б Сентер, П. (2007). «Новый взгляд на филогению целурозаврии (Dinosauria: Theropoda)». Журнал систематической палеонтологии, (Дои:10.1017 / S1477201907002143 )
- ^ а б c d е ж грамм час я j PC Sereno (1997) "Происхождение и эволюция динозавров" Анну. Преподобный "Планета Земля". Sci. 25: 435-489
- ^ Ричард Дж. Батлер, Пол Апчерч и Дэвид Б. Норман (2008). Филогения орнитисхиевых динозавров. Журнал систематической палеонтологии, 6, стр. 1-40 DOI: 10.1017 / S1477201907002271
- ^ «Введение», Сэмпсон (2001); стр.264.
- ^ Левин, Сара. "Динозавры когда-то ходили на кончиках пальцев?". Получено 2015-06-01.
- ^ https://www.theatlantic.com/magazine/archive/2018/09/dinosaur-extinction-debate/565769
- ^ Атлас динозавров, изд. J. Finch & S. Brown, DK, Лондон-Нью-Йорк-Мюнхен-Мельбурн-Дели, ç2006, стр.86.
- ^ К. Пим. Динозавры - Гранд Тур, The Experiment LLC, Нью-Йорк, ç2014, стр.453.
Рекомендации
- Сэмпсон, С. Д., 2001, Размышления о социоэкологии цератопсидных динозавров (Orinthischia: Neoceratopsia): In: Mesozoic Vertebrate Life, под редакцией Танке Д. Х. и Карпентера К., Indiana University Press, стр. 263–276.
- Пол С. Серено (1999) Эволюция динозавров, Science, Vol 284, pp. 2137–2146. http://www.sciencemag.org/cgi/content/abstract/284/5423/2137
