Армирование (видообразование) - Reinforcement (speciation)


Армирование это процесс видообразование куда естественный отбор увеличивает репродуктивная изоляция (далее делится на предзиготическая изоляция и постзиготическая изоляция ) между двумя популяциями видов. Это происходит в результате отбора, действующего против производства гибридный люди с низким фитнес. Первоначально идея была разработана Альфред Рассел Уоллес и иногда его называют Эффект Уоллеса. Современная концепция армирования берет свое начало от Феодосий Добжанский. Он предвидел разделение видов аллопатрически, где во время вторичный контакт две популяции спариваются, давая гибриды с более низким фитнес. Естественный отбор является результатом неспособности гибрида производить жизнеспособное потомство; таким образом, представители одного вида, не спаривающиеся с представителями другого, имеют больший репродуктивный успех. Это способствует эволюции большей презиготической изоляции (различия в поведении или биологии, которые препятствуют образованию гибридных зигот). Усиление - один из немногих случаев, когда отбор может способствовать увеличению презиготической изоляции, напрямую влияя на процесс видообразования.[1] Этот аспект особенно привлекателен для биологов-эволюционистов.[2]
Поддержка подкрепления колебалась с момента ее появления, а терминологическая путаница и различия в использовании на протяжении истории привели к множеству значений и осложнений. Различные возражения были высказаны эволюционные биологи относительно правдоподобия его возникновения. С 1990-х годов данные теории, экспериментов и природы преодолели многие из прошлых возражений, сделав подкрепление широко принятым,[3]:354 хотя его распространенность в природе остается неизвестной.[4][5]
Для понимания его действия в природе было разработано множество моделей, большинство из которых опирается на несколько аспектов: генетика, популяционные структуры, влияние отбора и вязка поведение. Эмпирический опора для армирования существует как в лаборатории, так и в природе. Задокументированные примеры встречаются у самых разных организмов: у обоих позвоночные и беспозвоночные, грибы, и растения. Вторичный контакт первоначально разделенных зарождающихся видов (начальная стадия видообразования) увеличивается из-за деятельности человека, такой как интродукция инвазивные виды или модификация натурального среда обитания.[6] Это имеет значение для мер биоразнообразие и может стать более актуальным в будущем.[6]
История
Армирование имеет сложную историю, поскольку его популярность среди ученых со временем изменилась.[7][8] Джерри Койн и Х. Аллен Орр утверждают, что теория подкрепления прошла через три фазы исторического развития:[3]:366
- правдоподобие на основе непригодных гибридов
- неправдоподобность на основании пригодности гибридов
- правдоподобие, основанное на эмпирических исследованиях и биологически сложных и реалистичных моделях

Иногда это называется эффектом Уоллеса, усиление было первоначально предложено Альфред Рассел Уоллес в 1889 г.[3]:353 Его гипотеза заметно отличалась от современной концепции тем, что постзиготическая изоляция, усиленный групповой выбор.[9][10][3]:353 Феодосий Добжанский был первым, кто дал подробное описание процесса в 1937 г.,[3]:353 хотя сам термин не был введен до 1955 г. У. Фрэнк Блэр.[11] В 1930 г. Рональд Фишер изложил первое генетическое описание процесса подкрепления в Генетическая теория естественного отбора, а в 1965 и 1970 годах было проведено первое компьютерное моделирование, чтобы проверить его достоверность.[3]:367 Позже популяционно-генетический[12] и количественные генетические[13] Были проведены исследования, показывающие, что совершенно непригодные гибриды однозначно приводят к увеличению презиготной изоляции.[3]:367
Идея Добжанского получила значительную поддержку; он предположил, что это иллюстрирует заключительный этап видообразования, например, после того, как аллопатрическая популяция вступает во вторичный контакт.[3]:353 В 1980-х годах многие биологи-эволюционисты начали сомневаться в правдоподобности этой идеи.[3]:353 основан не на эмпирических данных, а в основном на росте теории, которая считала это маловероятным механизмом репродуктивной изоляции.[2] В то время возник ряд теоретических возражений, которые рассматриваются в разделе «Аргументы против подкрепления» ниже.
К началу 1990-х годов популярность подкрепления среди биологов-эволюционистов возродилась; в первую очередь из-за внезапного увеличения количества данных - эмпирических данных исследований в лабораториях и, в основном, примеров, найденных в природе.[3]:354 Кроме того, компьютерное моделирование генетики и моделей миграции популяций обнаружило "кое-что смотрящий как подкрепление ».[3]:372 Самая последняя теоретическая работа по видообразованию была основана на нескольких исследованиях (особенно от Лиу и Прайса, Келли и Нур, и Киркпатрик и Servedio ) с использованием очень сложных компьютерных моделей; все они пришли к схожим выводам: усиление правдоподобно при нескольких условиях и во многих случаях проще, чем считалось ранее.[3]:374
Терминология
Существует путаница в отношении значения термина «подкрепление».[14] Впервые он был использован для описания наблюдаемых различий в брачных криках. Гастрофрин лягушки в гибридной зоне вторичного контакта.[14] Термин вторичный контакт также использовался для описания подкрепления в контексте аллопатрически разделенной популяции, испытывающей контакт после потери географического барьера.[15] Эффект Уоллеса похож на армирование, но используется редко.[14] Роджер Батлин разграничивает неполную постзиготическую изоляцию от полной изоляции, ссылаясь на неполную изоляцию как на подкрепление, а полностью изолированные популяции как на испытывающие смещение репродуктивного характера.[16] Дэниел Дж. Ховард считал смещение репродуктивного характера либо ассортивное спаривание или расхождение черт для распознавания партнера (особенно между симпатрическими популяциями).[14] Подкрепление, по его определению, включало презиготическую дивергенцию и полную постзиготическую изоляцию.[17] Servedio и Noor включают любое обнаруженное увеличение презиготической изоляции в качестве подкрепления, если это является ответом на отбор против спаривания между двумя разными видами.[4] Койн и Орр утверждают, что «истинное подкрепление ограничено случаями, когда изоляция усиливается между таксонами, которые все еще могут обмениваться генами».[3]:352
Модели
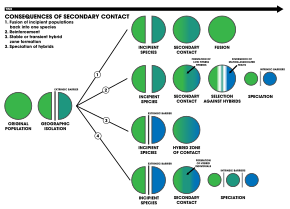
1. An внешний Барьер разделяет популяцию вида на две, но они вступают в контакт до того, как репродуктивная изоляция становится достаточной, чтобы привести к видообразованию. Две популяции сливаются в один вид
2. Видообразование по армированию
3. Две отдельные популяции остаются генетически разными, в то время как гибридные рои форма в зоне контакта
4. Рекомбинация генома приводит к видообразованию двух популяций с дополнительным гибридные виды. Все три вида разделены внутренний репродуктивные барьеры[18]
Одна из сильнейших форм репродуктивная изоляция По своей природе это сексуальная изоляция: особенности организмов, предполагающие спаривание.[19] Этот паттерн привел к мысли, что, поскольку отбор так сильно влияет на черты спаривания, он может быть вовлечен в процесс видообразования.[19] Этот процесс видообразования под влиянием естественного отбора является подкреплением и может происходить при любом способе видообразования.[3]:355 (например географические способы видообразования или экологическое видообразование[20]). Это требует двух сил эволюции, которые действуют на выбор партнера: естественный отбор и поток генов.[21] Отбор действует как главный двигатель усиления, поскольку он выбирает гибрид генотипы которые имеют низкий-фитнес независимо от того, не влияют ли индивидуальные предпочтения на выживание и воспроизводство.[21] Поток генов действует как основная противодействующая сила против подкрепления, поскольку обмен генами между людьми, приводящий к гибридам, вызывает генотипы для гомогенизации.[21]
Бутлин изложил четыре основных критерия подкрепления, которые должны быть обнаружены в естественных или лабораторных популяциях:[16]
- Поток генов между двумя таксонами существует или может быть установлен в какой-то момент.
- Между двумя таксонами наблюдается расхождение признаков, связанных со спариванием.
- Модифицируются модели спаривания, что ограничивает производство гибридов с низкой пригодностью.
- Других факторов давления отбора, ведущих к расхождению в системе распознавания партнера, не произошло.
После того, как происходит видообразование путем подкрепления, изменения после полной репродуктивной изоляции (и последующей изоляции) являются формой репродуктивной изоляции. смещение персонажа.[22] Распространенным признаком появления подкрепления в природе является смещение репродуктивного характера; характеристики населения расходятся по симпатрии, но не по аллопатрии.[6][5] Одна из трудностей в обнаружении заключается в том, что смещение экологического характера может привести к тем же самым образцам.[23] Кроме того, поток генов может уменьшить изоляцию симпатрических популяций.[23] Два важных фактора в исходе процесса зависят от: 1) конкретных механизмов, вызывающих презиготную изоляцию, и 2) количества аллелей, измененных мутациями, влияющими на выбор партнера.[24]
В случаях перипатрическое видообразование подкрепление вряд ли завершит видообразование в случае, если периферически изолированная популяция вступает во вторичный контакт с основной популяцией.[25] В симпатрическое видообразование, необходима селекция на гибриды; поэтому подкрепление может сыграть свою роль, учитывая эволюцию некоторых форм компромиссов в пригодности.[1] В симпатрии часто наблюдаются модели сильной дискриминации при спаривании, приписываемой подкреплению.[7] Подкрепление считается агентом гаметический изоляция.[26]
Генетика
Основную генетику подкрепления можно понять с помощью идеальной модели двух гаплоидных популяций, испытывающих увеличение нарушение равновесия по сцеплению. Здесь отбор отклоняет низкий фитнес или же аллель комбинации, отдавая предпочтение комбинациям аллели (в первой субпопуляции) и аллели (во второй субпопуляции). Третий локус или же (ассортивные аллели спаривания) влияют на образец спаривания, но не подвергаются прямому отбору. Если выбор на и вызывают изменения частоты аллеля ассортивное спаривание способствует усилению. Необходимы как отбор, так и выборочное спаривание, т. Е. Спаривание и встречаются чаще, чем спаривания и .[27] Ограничение миграции между популяциями может еще больше увеличить шанс подкрепления, так как снижает вероятность обмена разными генотипами.[14]












Альтернативная модель существует для устранения антагонизма рекомбинация, так как это может уменьшить ассоциацию между аллелями, которые связаны с приспособленностью, и ассортивными аллелями спаривания, которые этого не делают.[14] Генетические модели часто различаются по количеству признаков, связанных с места;[28] некоторые полагаются на один локус на каждый признак[25][29][30] и другие на полигенный черты.[22][21][31]
Структура населения
Структура и модели миграции популяции могут влиять на процесс видообразования путем подкрепления. Было показано, что это происходит в рамках островной модели, в которой существуют условия с редкими миграциями, происходящими в одном направлении,[21] и в моделях симметричной миграции, где виды равномерно мигрируют туда и обратно между популяциями.[25][29]
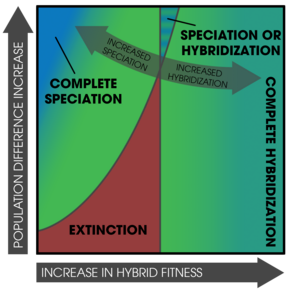
Подкрепление также может происходить в отдельных популяциях,[28][22] мозаика гибридные зоны (неоднородное распределение родительских форм и субпопуляций),[30] И в парапатрический популяции с узкими зонами контакта.[32]
Плотность населения является важным фактором подкрепления, часто в сочетании с вымирание.[22] Возможно, что при вторичном контакте двух видов одна популяция вымрет - в первую очередь из-за низкой гибридности. фитнес сопровождается высокими темпами роста населения.[22] Вымирание менее вероятно, если гибриды нежизнеспособны вместо бесплодный, так как плодородный особи все еще могут выжить достаточно долго, чтобы размножаться.[22]
Выбор
Видообразование путем подкрепления напрямую зависит от отбора, способствующего увеличению презиготической изоляции,[1] и природа роли отбора в подкреплении широко обсуждалась, модели применяли различные подходы.[28] Селекция, действующая на гибриды, может происходить несколькими способами. Все полученные гибриды могут быть равноценны низкой пригодности,[22] что дает большой недостаток. В других случаях выбор может отдавать предпочтение множественным и разным фенотипы[25] например, в случае мозаичной гибридной зоны.[30] Естественный отбор может воздействовать на определенные аллели как на напрямую или косвенно.[28][21][33] При прямом отборе частота выбранного аллеля максимально благоприятна. В случаях, когда аллель выбирается косвенно, его частота увеличивается из-за другого сцепленного аллеля, подвергающегося отбору (нарушение равновесия по сцеплению ).[14]
Состояние селектируемых гибридов может играть роль в постзиготической изоляции, так как гибридная незащищенность (гибрид, неспособный созреть до пригодной взрослой особи) и бесплодие (неспособность полностью произвести потомство) запрещают обмен генами между популяциями.[7] Отбор против гибридов может быть вызван даже любой неудачей в получении партнера, поскольку он практически неотличим от бесплодия - каждое обстоятельство не приводит к появлению потомства.[7]
Брачные отношения и предпочтение партнера
Некоторое начальное расхождение в предпочтениях партнера должно присутствовать, чтобы произошло подкрепление.[7][22][34] Любые черты, способствующие изоляции, могут подвергаться подкреплению, например, сигналам спаривания (например дисплей ухаживания ), сигнал ответы, местонахождение нерестилищ, время вязки (например сезонное разведение ) или даже восприимчивость яиц.[14] Люди также могут дискриминировать партнеров, которые различаются по различным признакам, таким как брачный зов или же морфология.[35] Многие из этих примеров описаны ниже.
Свидетельство
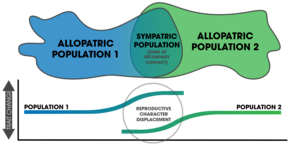
В доказательства для подкрепления исходит из наблюдений в природе, сравнительных исследований и лабораторных экспериментов.[3]:354
Природа
Можно показать, что подкрепление происходит (или происходило в прошлом), путем измерения силы презиготической изоляции в симпатрической популяции по сравнению с аллопатрической популяцией того же вида.[3]:357 Сравнительные исследования этого позволяют определить крупномасштабные закономерности в природе для различных таксонов.[3]:362 Схемы спаривания в гибридные зоны также может использоваться для обнаружения армирования.[17] Смещение репродуктивного характера рассматривается как результат армирования,[7] так много случаев в природе выражают эту закономерность в симпатии. Вездесущность подкрепления неизвестна,[4] но паттерны смещения репродуктивных признаков обнаруживаются во многих таксонах и считаются обычным явлением в природе.[17] Изучение подкрепления в природе часто оказывается трудным, поскольку можно утверждать альтернативные объяснения обнаруженных закономерностей.[3]:358 Тем не менее, существуют эмпирические доказательства того, что подкрепление встречается в различных таксонах.[7] и его роль в ускорении видообразования неоспорима.[14]
Сравнительные исследования
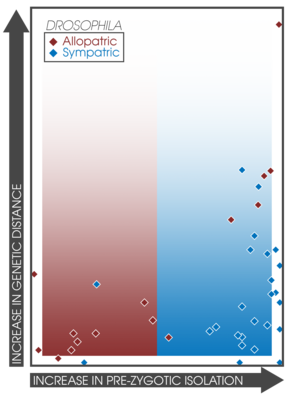
Ожидается, что ассортивное спаривание будет увеличиваться среди симпатрических популяций, испытывающих подкрепление.[14] Этот факт позволяет напрямую сравнивать силу презиготической изоляции в симпатрии и аллопатрии между различными экспериментами и исследованиями.[3]:362 Койн и Орр обследовали 171 пару видов, собирая данные об их географическом характере, генетическая дистанция и сила презиготической и постзиготической изоляции; обнаружение, что презиготическая изоляция была значительно сильнее в симпатрических парах, коррелируя с возрастом вида.[3]:362 Кроме того, сила постзиготической изоляции не различалась между симпатрическими и аллопатрическими парами.[14] Этот вывод поддерживает предсказания видообразования путем подкрепления и хорошо коррелирует с более поздними исследованиями.[17] которые обнаружили 33 исследования, демонстрирующие образцы сильной презиготической изоляции у сочувствие.[3]:363 Исследование скорости видообразования у рыб и связанных с ними гибридных зон обнаружило сходные закономерности в симпатрии, что подтверждает наличие подкрепления.[37]
Лабораторные эксперименты
Лабораторные исследования, явно проверяющие наличие подкрепления, ограничены.[3]:357 причем многие эксперименты проводились на Дрозофила плодовые мошки. В общем, было проведено два типа экспериментов: с использованием искусственного отбора для имитации естественного отбора, который исключает гибриды (часто называемого «уничтожение гибридов»), и с использованием подрывной отбор отбирать по признаку (независимо от его функции в половом размножении).[3]:355–357 Многие эксперименты с использованием техники уничтожения гибридов обычно упоминаются как поддерживающие подкрепление; однако некоторые исследователи, такие как Койн и Орр, Уильям Р. Райс и Эллен Э. Хостерт, утверждают, что они не моделируют подкрепление, поскольку поток генов между двумя популяциями полностью ограничен.[38][3]:356
Альтернативные гипотезы
Были предложены различные альтернативные объяснения закономерностей, наблюдаемых в природе.[3]:375 Не существует единого всеобъемлющего признака подкрепления; однако есть две предлагаемые возможности:[3]:379 асимметрия пола (где женщины в симпатрических популяциях вынуждены становиться разборчивыми перед лицом двух разных мужчин)[39] и что из аллельное доминирование: любой из аллелей, подвергшихся отбору для выделения, должен преобладать.[7] Хотя эта подпись не полностью учитывает вероятности фиксации или же смещение экологического характера.[3]:380 Койн и Орр расширяют сигнатуру половой асимметрии и утверждают, что, независимо от изменений, наблюдаемых у женщин и мужчин в симпатии, изоляция в большей степени определяется самками.[3]:380
Экологические или этологические влияния
Экология может также играть роль в наблюдаемых закономерностях, называемых смещением экологического характера. Естественный отбор может способствовать сокращению перекрытия ниш между видами вместо того, чтобы действовать для уменьшения гибридизации.[3]:377 Хотя один эксперимент в колюшка рыба, которая явно проверила эту гипотезу, не нашла доказательств.[40]
Взаимодействие видов также может приводить к смещению репродуктивного характера (как по предпочтению партнера, так и по сигналу спаривания).[19] Примеры включают давление хищников и конкуренции, паразиты, обманчивый опыление, и мимикрия.[19] Поскольку эти и другие факторы могут привести к смещению репродуктивного характера, Конрад Дж. Хоскин и Меган Хигги дадим пять критериев усиления, чтобы различать экологические и этологический влияет:
(1) брачные признаки определены у основных видов; (2) на брачные признаки влияет взаимодействие видов, так что отбор по брачным признакам вероятен; (3) виды взаимодействия различаются между популяциями (присутствующие и отсутствующие, или различные виды взаимодействия, влияющие на брачные черты в каждой популяции); (4) брачные признаки (сигнал и / или предпочтение) различаются между популяциями из-за различий во взаимодействии видов; (5) видообразование требует демонстрации того, что расхождение брачных признаков приводит к полной или почти полной сексуальной изоляции среди популяций. Результаты будут наиболее информативными в хорошо разрешенной биогеографической обстановке, где известны отношения и история популяций.[19]
Слияние
Возможно, что паттерн усиленной изоляции может быть просто временным результатом вторичного контакта, когда два аллопатрических вида уже имеют различный диапазон презиготической изоляции: одни проявляют больше, чем другие.[41] Те, у кого более слабая презиготическая изоляция, в конечном итоге сливаются, теряя свою отличительность.[7] Эта гипотеза не объясняет того факта, что отдельные виды в аллопатрии, испытывающие постоянный поток генов, не будут различаться по уровням потока генов при вторичном контакте.[7][42] Кроме того, закономерности, обнаруженные в Дрозофила обнаруживают высокий уровень презиготической изоляции в симпатрии, но не в аллопатрии.[43] Гипотеза слияния предсказывает, что сильная изоляция должна быть найдена как в аллопатрии, так и в симпатрии.[43] Считается, что этот процесс слияния происходит в природе, но он не полностью объясняет закономерности, обнаруживаемые при подкреплении.[3]:376
Сочувствие

Возможно, что процесс симпатрическое видообразование сам по себе может привести к наблюдаемым образцам армирования.[3]:378 Один из методов различения этих двух видов - построить филогенетическую историю вида, поскольку сила презиготической изоляции между группой родственных видов должна различаться в зависимости от того, как они видоизменялись в прошлом.[44] Два других способа определить, происходит ли подкрепление (в отличие от симпатрического видообразования):
- если два недавно видоизмененных таксона не обнаруживают признаков постзиготической изоляции как симпатрических, так и аллопатрических популяций (при симпатрическом видообразовании постзиготическая изоляция не является обязательным условием);[45]
- если клина существует между двумя видами по ряду признаков (симпатрическое видообразование вообще не требует существования клины).[46]
Половой отбор
В неконтролируемом процессе (мало чем отличается от Отбор беглых рыбаков ), отбор против гибридов с низкой пригодностью способствует ассортивному спариванию, быстро увеличивая дискриминацию спаривания.[7][43] Кроме того, когда предпочтения самок невысоки, в результате могут происходить изменения мужских фенотипов, выражающие паттерн, идентичный паттерну репродуктивного смещения признаков.[47] Постзиготическая изоляция не нужна, потому что непригодные гибриды не могут найти себе пару.[7]
Аргументы против подкрепления
Был выдвинут ряд возражений, в основном в течение 1980-х годов, аргументирующих неправдоподобие подкрепления.[7][19][3]:369 Большинство полагается на теоретические работы, которые предполагали, что антагонизм между силами естественного отбора и потоком генов был самым большим препятствием на пути к его выполнимости.[3]:369–372 Эти возражения с тех пор в значительной степени опровергаются свидетельствами природы.[17][3]:372
Генетический поток
Обеспокоенность по поводу того, что приспособленность гибридов играет роль в подкреплении, привела к возражениям, основанным на взаимосвязи между отбором и рекомбинацией.[5][3]:369 То есть, если поток генов не равен нулю (если гибриды не полностью непригодны), отбор не может управлять фиксацией аллелей для презиготической изоляции.[27] Например: если население имеет презиготный изолирующий аллель и высокая пригодность, постзиготические аллели и ; и население имеет презиготный аллель а и высокую приспособленность постзиготические аллели и , обе и генотипы будут подвергаться рекомбинации перед лицом потока генов. Каким-то образом необходимо поддерживать популяцию.[3]:369






Кроме того, определенные аллели, которые обладают избирательным преимуществом в перекрывающихся популяциях, полезны только в пределах этой популяции.[48] Однако, если они имеют избирательное преимущество, поток генов должен позволить аллелям распространяться в обеих популяциях.[48] Чтобы предотвратить это, аллели должны быть вредными или нейтральными.[3]:371 Это не без проблем, поскольку поток генов из предположительно больших аллопатрических областей может подавить область, когда две популяции перекрываются.[3]:371 Чтобы подкрепление работало, поток генов должен присутствовать, но очень ограниченный.[25][30]
Недавние исследования показывают, что подкрепление может происходить в более широком диапазоне условий, чем считалось ранее.[28][45][3]:372–373 и что эффект потока генов можно преодолеть путем отбора.[49][50] Например, два вида Drosophila santomea и Д. Якуба на африканском острове Сан-Томе иногда скрещиваются друг с другом, в результате чего получается плодовитое потомство женского пола и бесплодное потомство мужского пола.[49] Эта естественная обстановка была воспроизведена в лаборатории, непосредственно моделируя подкрепление: удаление некоторых гибридов и допуск различных уровней потока генов.[50] Результаты эксперимента убедительно свидетельствуют о том, что подкрепление работает в различных условиях, с эволюцией половой изоляции, возникающей в 5–10 поколениях плодовых мух.[50]
Быстрые требования
В сочетании с гипотезой слияния подкрепление можно рассматривать как гонку против слияния и вымирания.[41] Получение непригодных гибридов фактически то же самое, что и недостаток гетерозиготы; посредством чего отклонение от генетическое равновесие вызывает потерю непригодного аллеля.[51] Этот эффект приведет к исчезновению одной из популяций.[52] Это возражение преодолевается, когда обе популяции не находятся в одинаковых экологических условиях.[3]:370 Тем не менее, вымирание одной популяции все еще возможно, и это было показано в популяционном моделировании.[53] Чтобы произошло подкрепление, презиготическая изоляция должна произойти быстро.[3]:370
Рекомендации
- ^ а б c Ханнес Шулер, Глен Р. Худ, Скотт П. Иган и Джеффри Л. Федер (2016), «Режимы и механизмы видообразования», Обзоры в области клеточной биологии и молекулярной медицины, 2 (3): 60–93, Дои:10.1002 / 3527600906.mcb.201600015 (неактивно 10.10.2020)CS1 maint: несколько имен: список авторов (связь) CS1 maint: DOI неактивен с октября 2020 г. (связь)
- ^ а б Джереми Л. Маршалл, Майкл Л. Арнольд и Дэниел Дж. Ховард (2002), «Укрепление: дорога не пройдена», Тенденции в экологии и эволюции, 17 (12): 558–563, Дои:10.1016 / S0169-5347 (02) 02636-8CS1 maint: несколько имен: список авторов (связь)
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах ай эй ак аль являюсь ан ао Джерри А. Койн; Х. Аллен Орр (2004), Видообразование, Sinauer Associates, стр. 1–545, ISBN 978-0-87893-091-3
- ^ а б c Мария Р. Серведио; Мохамед А. Ф. Нур (2003), "Роль подкрепления в видообразовании: теория и данные", Ежегодный обзор экологии, эволюции и систематики, 34: 339–364, Дои:10.1146 / annurev.ecolsys.34.011802.132412
- ^ а б c Даниэль Ортис-Барриентос, Алисия Грили и Патрик Носил (2009), «Генетика и экология подкрепления: последствия для эволюции презиготической изоляции в симпатрии и за ее пределами», Летопись Нью-Йоркской академии наук, 1168: 156–182, Дои:10.1111 / j.1749-6632.2009.04919.x, PMID 19566707, S2CID 4598270CS1 maint: несколько имен: список авторов (связь)
- ^ а б c Мария Р. Серведио (2004 г.), "Что и почему исследования по армированию", PLOS Биология, 2 (12): 2032–2035, Дои:10.1371 / journal.pbio.0020420, ЧВК 535571, PMID 15597115
- ^ а б c d е ж грамм час я j k л м Мохамед А. Ф. Нур (1999), «Укрепление и другие последствия симпатрии», Наследственность, 83 (5): 503–508, Дои:10.1038 / sj.hdy.6886320, PMID 10620021, S2CID 26625194
- ^ Роджер К. Батлин и Кэрол М. Смаджа (2018), «Соединение, армирование и видообразование» (PDF), Американский натуралист, 191 (2): 155–172, Дои:10.1086/695136, PMID 29351021
- ^ М. Дж. Литтлджон (1981). Репродуктивная изоляция: критический обзор. В У. Р. Атчли и Д. С. Вудрафф (ред.) Эволюция и видообразование, Издательство Кембриджского университета, стр. 298–334.
- ^ Марио А. Фарес (2015), Естественный отбор: методы и приложения, CRC Press, стр. 3, ISBN 9781482263725
- ^ Блэр, В. Франк (1955), «Брачный вызов и стадия видообразования в Microhyla olivacea-M. carolinensis сложный", Эволюция, 9 (4): 469–480, Дои:10.1111 / j.1558-5646.1955.tb01556.x, S2CID 88238743
- ^ Стэнли Сойер и Дэниел Хартл (1981), «Об эволюции поведенческой репродуктивной изоляции: эффект Уоллеса», Теоретическая популяционная биология, 19 (1): 261–273, Дои:10.1016/0040-5809(81)90021-6
- ^ Дж. А. Свед (1981), "Двухполая полигенная модель эволюции преждевременной изоляции. I. Детерминистская теория для естественных популяций", Генетика, 97 (1): 197–215, ЧВК 1214384, PMID 17249073
- ^ а б c d е ж грамм час я j k Гленн-Питер Сэтр (2012). «Армирование». eLS. Дои:10.1002 / 9780470015902.a0001754.pub3. ISBN 978-0470016176. Отсутствует или пусто
| название =(помощь) - ^ Добжанский, Феодосий (1937). Генетика и происхождение видов. Издательство Колумбийского университета.
- ^ а б Бутлин, Роджер К. (1989). «Усиление внутренней изоляции». In Otte, D .; Эндлер, Джон А. (ред.). Видообразование и его последствия. Sinauer Associates. стр.158–179. ISBN 978-0-87893-657-1.
- ^ а б c d е Ховард, Дэниел Дж. (1993). «Подкрепление: происхождение, динамика и судьба эволюционной гипотезы». В Харрисон, Р. Г. (ред.). Гибридные зоны и эволюционный процесс. Издательство Оксфордского университета. С. 46–69. ISBN 978-0-19-506917-4.
- ^ Джон А. Хвала и Трой Э. Вуд (2012). «Видообразование: Введение». eLS. Дои:10.1002 / 9780470015902.a0001709.pub3. ISBN 978-0470016176. Отсутствует или пусто
| название =(помощь) - ^ а б c d е ж Конрад Дж. Хоскин и Меган Хигги (2010), «Видообразование через видовые взаимодействия: расхождение брачных признаков внутри видов», Письма об экологии, 13 (4): 409–420, Дои:10.1111 / j.1461-0248.2010.01448.x, PMID 20455922, S2CID 16175451
- ^ Марк Киркпатрик (2001), «Усиление во время экологического видообразования», Труды Королевского общества B, 268 (1473): 1259–1263, Дои:10.1098 / rspb.2000.1427, ЧВК 1088735, PMID 11410152
- ^ а б c d е ж Марк Киркпатрик и Мария Р. Серведио (1999), «Укрепление брачных предпочтений на острове», Генетика, 151 (2): 865–884, ЧВК 1460501, PMID 9927476
- ^ а б c d е ж грамм час я Лили В. Лиу и Тревор Д. Прайс (1994), "Видообразование путем усиления изоляции перед беременностью", Эволюция, 48 (5): 1451–1459, Дои:10.1111 / j.1558-5646.1994.tb02187.x, PMID 28568419, S2CID 22630822
- ^ а б Сеть Marie Curie SPECIATION (2012), "Что нам нужно знать о видообразовании?", Тенденции в экологии и эволюции, 27 (1): 27–39, Дои:10.1016 / j.tree.2011.09.002, PMID 21978464
- ^ Claudia Bank, Иоахим Хермиссион и Марк Киркпатрик (2012), "Может ли подкрепление завершить видообразование?", Эволюция, 66 (1): 229–239, Дои:10.1111 / j.1558-5646.2011.01423.x, PMID 22220877, S2CID 15602575CS1 maint: несколько имен: список авторов (связь)
- ^ а б c d е Мария Р. Серведио и Марк Киркпатрик (1997), «Влияние потока генов на подкрепление», Эволюция, 51 (6): 1764–1772, Дои:10.1111 / j.1558-5646.1997.tb05100.x, PMID 28565111, S2CID 12269299
- ^ Дэниел Р. Матуте (2010), «Усиление гаметической изоляции в Дрозофила", PLOS Биология, 8 (6): e1000341, Дои:10.1371 / journal.pbio.1000341, ЧВК 2843595, PMID 20351771
- ^ а б Джозеф Фельзенштейн (1981), «Скептицизм по отношению к Санта-Розалии, или почему так мало видов животных?», Эволюция, 35 (1): 124–138, Дои:10.2307/2407946, JSTOR 2407946, PMID 28563447
- ^ а б c d е Майкл Турелли; Николас Х. Бартон; Джерри А. Койн (2001), «Теория и видообразование», Тенденции в экологии и эволюции, 16 (7): 330–343, Дои:10.1016 / S0169-5347 (01) 02177-2, PMID 11403865
- ^ а б Мария Р. Серведио (2000), «Усиление и генетика неслучайного спаривания», Эволюция, 54 (1): 21–29, Дои:10.1111 / j.0014-3820.2000.tb00003.x, PMID 10937179, S2CID 12563023
- ^ а б c d Майкл Л. Кейн, Вигго Андреасен и Дэниел Дж. Ховард (1999), «Усиление отбора эффективно в относительно широком наборе условий в мозаичной гибридной зоне», Эволюция, 53 (5): 1343–1353, Дои:10.1111 / j.1558-5646.1999.tb05399.x, PMID 28565558, S2CID 31107731CS1 maint: несколько имен: список авторов (связь)
- ^ Марк Киркпатрик (2000), «Усиление и расхождение при ассортивном спаривании», Труды Королевского общества B, 267 (1453): 1649–1655, Дои:10.1098 / rspb.2000.1191, ЧВК 1690725, PMID 11467428
- ^ Нил Сандерсон (1989), «Может ли поток генов предотвратить подкрепление?», Эволюция, 43 (6): 1223–1235, Дои:10.2307/2409358, JSTOR 2409358, PMID 28564502
- ^ Мария Р. Серведио (2001), «Помимо подкрепления: эволюция изоляции перед беременностью путем прямого отбора на основе предпочтений и пост-спаривания, презиготической несовместимости», Эволюция, 55 (10): 1909–1920, Дои:10.1111 / j.0014-3820.2001.tb01309.x, PMID 11761053, S2CID 25296147
- ^ Дж. К. Келли и Мохамед А. Ф. Нур (1996), "Видообразование путем подкрепления: модель, полученная на основе исследований дрозофилы", Генетика, 143 (3): 1485–1497, ЧВК 1207414, PMID 8807317
- ^ Конрад Дж. Хоскин, Меган Хигги, Кейт Р. Макдональд и Крейг Мориц (2005), «Укрепление стимулирует быстрое аллопатрическое видообразование», Природа, 437 (7063): 1353–1356, Bibcode:2005Натура 437.1353H, Дои:10.1038 / nature04004, PMID 16251964, S2CID 4417281CS1 maint: несколько имен: список авторов (связь)
- ^ Джерри А. Койн и Х. Аллен Орр (1997), ""Модели видообразования в Дрозофила"Пересмотренный", Эволюция, 51 (1): 295–303, Дои:10.1111 / j.1558-5646.1997.tb02412.x, PMID 28568795, S2CID 40390753
- ^ А. Р. МакКьюн и Н. Р. Лавджой. (1998). Относительная скорость симпатрического и аллопатрического видообразования у рыб. В Д. Дж. Ховарде и С. Х. Берлоше (редакторы) Бесконечные формы: виды и видообразование, Oxford University Press, стр. 172–185.
- ^ Уильям Р. Райс и Эллен Э. Хостерт (1993), "Лабораторные эксперименты по видообразованию: что мы узнали за 40 лет?", Эволюция, 47 (6): 1637–1653, Дои:10.1111 / j.1558-5646.1993.tb01257.x, PMID 28568007, S2CID 42100751
- ^ Л. Партридж и Г. А. Паркер. (1999). Сексуальный конфликт и видообразование. В А. Э. Магурран и Р. М. Мэй (ред.) Эволюция биологического разнообразия. Oxford University Press, стр.130–159.
- ^ Ховард Д. Рандл (1998), «Укрепление партнерских предпочтений колюшки: сочувствие порождает презрение», Дольф Шлютер, 52 (1): 200–208, Дои:10.1111 / j.1558-5646.1998.tb05153.x, HDL:2429/6366, PMID 28568163, S2CID 40648544
- ^ а б Алан Р. Темплтон (1981), "Механизмы видообразования - популяционно-генетический подход", Ежегодный обзор экологии, эволюции и систематики, 12: 23–48, Дои:10.1146 / annurev.es.12.110181.000323
- ^ Мохамед А. Ф. Нур (1995), "Видообразование, вызванное естественным отбором в Дрозофила", Природа, 375 (6533): 674–675, Bibcode:1995Натура.375..674Н, Дои:10.1038 / 375674a0, PMID 7791899, S2CID 4252448
- ^ а б c Джерри А. Койн; Х. Аллен Орр (1989), "Модели видообразования в Дрозофила", Эволюция, 43 (2): 362–381, Дои:10.1111 / j.1558-5646.1989.tb04233.x, PMID 28568554, S2CID 1678429
- ^ а б Мохамед А. Ф. Нур (1997), "Как часто симпатия влияет на сексуальную изоляцию в Дрозофила?", Американский натуралист, 149 (6): 1156–1163, Дои:10.1086/286044, PMID 18811269
- ^ а б Марк Киркпатрик и Виржини Равинье (2002), "Видообразование путем естественного и полового отбора: модели и эксперименты", Американский натуралист, 159: S22–35, Дои:10.1086/338370, PMID 18707367, S2CID 16516804
- ^ Н. Х. Бартон и Г. М. Хьюитт (1989), "Адаптация, видообразование и гибридные зоны", Природа, 341 (6242): 497–503, Bibcode:1989Натура.341..497Б, Дои:10.1038 / 341497a0, PMID 2677747, S2CID 4360057
- ^ Трой Дэй (2000), «Половой отбор и эволюция дорогостоящих женских предпочтений: пространственные эффекты», Эволюция, 54 (3): 715–730, Дои:10.1554 / 0014-3820 (2000) 054 [0715: SSATEO] 2.3.CO; 2, PMID 10937247
- ^ а б Дж. А. Мур. (1957). Взгляд эмбриолога на понятие вида. У Эрнста Майра (ред.) Проблема видов, Американская ассоциация развития науки, стр. 325–338.
- ^ а б Дэниел Р. Матуте (2010), «Усиление может преодолеть поток генов во время видообразования в Дрозофила", Текущая биология, 20 (24): 2229–2233, Дои:10.1016 / j.cub.2010.11.036, ЧВК 3019097, PMID 21129972
- ^ а б c Джерри А. Койн (2010), «Армирование» и происхождение видов, Wordpress
- ^ А. А. Харпер и Д. М. Ламберт (1983), "Популяционная генетика усиливающего отбора", Genetica, 62 (1): 15–23, Дои:10.1007 / BF00123305, S2CID 7947934
- ^ Х. Э. Х. Патерсон (1978), «Дополнительные доказательства против видообразования путем подкрепления», Южноафриканский научный журнал, 74: 369–371
- ^ Хэмиш Г. Спенсер, Брайан Х. Макардл и Дэвид М. Ламберт (1986), «Теоретическое исследование видообразования путем подкрепления», Американский натуралист, 128 (2): 241–262, Дои:10.1086/284557CS1 maint: несколько имен: список авторов (связь)
| Базовые концепты | ||
|---|---|---|
| Географические режимы | ||
| Изолирующие факторы | ||
| Гибридизация | ||
| Видообразование в таксонах | ||
