Полиплоидия - Polyploidy

Полиплоидия это состояние, в котором клетки из организм иметь более двух парных (гомологичный ) наборы хромосомы. Большинство видов, клетки которых имеют ядра (эукариоты ) находятся диплоид, что означает, что у них есть два набора хромосом - по одному, унаследованному от каждого родителя. Однако некоторые организмы полиплоид, а полиплоидия особенно распространена у растений. У большинства эукариот диплоид соматические клетки, но производят гаплоид гаметы (яйца и сперма) мейоз. А моноплоидный имеет только один набор хромосом, и этот термин обычно применяется только к клеткам или организмам, которые обычно являются гаплоидными. Самцы пчелы и другие Перепончатокрылые, например, моноплоидны. В отличие от животных, растения и многоклеточные водоросли имеют жизненные циклы с двумя чередование многоклеточных поколений. В гаметофит поколение гаплоидно и производит гаметы путем митоз, то спорофит поколение диплоидно и производит споры мейозом.
Полиплоидия может возникнуть из-за ненормального деление клеток, либо во время митоз, или обычно во время метафаза I в мейоз. Кроме того, это может быть вызвано у растений и клеточные культуры некоторыми химическими веществами: наиболее известным является колхицин, что может привести к удвоению хромосом, хотя его использование может иметь и другие менее очевидные последствия. Оризалин также удвоит существующее содержание хромосом.
Полиплоидия возникает в высокодифференцированных тканях человека в печени, сердечной мышце, костном мозге и плаценте.[1] Встречается в соматических клетках некоторых животные, Такие как золотая рыбка,[2] лосось, и саламандры, но особенно распространен среди папоротники и цветение растения (видеть Hibiscus rosa-sinensis ), включая как дикие, так и культурные разновидность. Пшеница, например, спустя тысячелетия гибридизация и модификация человеком, имеет штаммы, которые диплоид (два набора хромосом), тетраплоид (четыре набора хромосом) с общим названием твердый или макароны из пшеницы, и гексаплоид (шесть наборов хромосом) с общим названием мягкой пшеницы. Многие важные сельскохозяйственные растения этого рода Brassica тоже тетраплоиды.
Полиплоидизация может быть механизмомсимпатрическое видообразование потому что полиплоиды обычно не могут скрещиваться со своими диплоидными предками. Примером может служить завод Erythranthe peregrina. Секвенирование подтвердило, что этот вид произошел от Е. × Роберции, стерильный триплоидный гибрид между E. guttata и E. lutea, оба были введены и натурализованы в Соединенном Королевстве. Новые популяции Е. перегрина возник на материковой части Шотландии и на Оркнейских островах путем дупликации генома у местных популяций Е. × Роберции.[3] Из-за редкой генетической мутации Е. перегрина не бесплоден.[4]
Терминология
Типы
Полиплоид типы помечаются в соответствии с количеством наборов хромосом в ядро. Письмо Икс используется для обозначения количества хромосом в одном наборе:
- гаплоидный (один комплект; 1x)
- диплоид (два набора; 2x)
- триплоид (три комплекта; 3Икс), например стерильный шафрановый крокус, или же арбузы без косточек, также распространен в филюм Тихоходка[5]
- тетраплоид (четыре комплекта; 4Икс), Например Лососевые рыбы,[6] хлопок Gossypium hirsutum[7]
- пентаплоид (пять сетов; 5Икс), например Kenai Birch (Betula papyrifera var. Kenaica)
- гексаплоид (шесть сетов; 6Икс), Например пшеница, киви[8]
- гептаплоид или же септаплоид (семь сетов; 7Икс)
- октаплоид или же октоплоид, (восемь сетов; 8Икс), Например Acipenser (род осетр рыбы), георгины
- декаплоид (десять комплектов; 10Икс), например некоторые клубника
- додекаплоид (двенадцать наборов; 12Икс), например растения Celosia argentea и Spartina anglica [9] или амфибия Xenopus ruwenzoriensis.
Классификация
Автополиплоидия
Автополиплоиды - это полиплоиды с несколькими наборами хромосом, полученными из одного таксон.
Двумя примерами природных автополиплоидов являются контрейлерные растения, Толмия Мензисии[10] и белый осетр, Acipenser transmontanum.[11] Большинство случаев автополиплоидии является результатом слияния нередуцированных (2п) гаметы, в результате чего либо триплоид (п + 2п = 3п) или тетраплоид (2п + 2п = 4п) потомство.[12] Триплоидное потомство обычно бесплодно (как в случае феномена 'триплоидный блок '), но в некоторых случаях они могут давать высокую долю невосстановленных гамет и, таким образом, способствовать образованию тетраплоидов. Этот путь к тетраплоидии называется «триплоидным мостом».[12] Триплоиды также могут сохраняться через бесполое размножение. Фактически, стабильная аутотриплоидия у растений часто связана с апомиктик системы сопряжения.[13] В сельскохозяйственных системах аутотриплоидия может привести к бессемянности, как в арбузы и бананы.[14] Триплоидия также используется при выращивании лосося и форели для обеспечения бесплодия.[15][16]
Редко автополиплоиды возникают в результате спонтанного соматического удвоения генома, которое наблюдалось у яблони (Malus domesticus) Bud Sports.[17] Это также наиболее распространенный путь искусственно индуцированной полиплоидии, при котором такие методы, как слияние протопластов или лечение с колхицин, Оризалин или же ингибиторы митоза используются для нарушения нормального митотический деление, в результате которого образуются полиплоидные клетки. Этот процесс может быть полезен при селекции растений, особенно при попытках интрогрессии зародышевой плазмы на плоидных уровнях.[18]
Автополиплоиды обладают как минимум тремя гомологичная хромосома наборы, которые могут привести к высокому уровню поливалентного спаривания во время мейоз (особенно в недавно сформированных автополиплоидах, также известных как неополиплоиды) и связанном с этим снижением фертильности из-за производства анеуплоид гаметы.[19] Естественный или искусственный отбор на фертильность может быстро стабилизировать мейоз у автополиплоидов, восстанавливая бивалентные пары во время мейоза, но высокая степень гомология среди дублированных хромосом вызывает отображение автополиплоидов полисомное наследование.[20] Этот признак часто используется в качестве диагностического критерия для отличия автополиплоидов от аллополиплоидов, которые обычно демонстрируют дисомную наследование после того, как они прогрессируют после стадии неополиплоида.[21] Хотя большинство полиплоидных видов однозначно охарактеризованы как автополиплоидные или аллополиплоидные, эти категории представляют собой концы спектра дивергенции между родительскими субгеномами. Полиплоиды, которые находятся между этими двумя крайностями, которые часто называют сегментарными аллополиплоидами, могут иметь промежуточные уровни полисомного наследования, которые варьируются в зависимости от локуса.[22][23]
Считается, что около половины всех полиплоидов являются результатом автополиплоидии,[24][25] хотя многие факторы затрудняют оценку этой доли.[26]
Аллополиплоидия
Аллополиплоиды или амфиполиплоиды или же гетерополиплоиды представляют собой полиплоиды с хромосомами, происходящими от двух или более расходящихся таксонов.
Как и в случае автополиплоидии, это происходит прежде всего за счет слияния невосстановленных (2п) гаметы, которые могут иметь место до или после гибридизация. В первом случае нередуцированные гаметы от каждого диплоидного таксона - или редуцированные гаметы от двух автотетраплоидных таксонов - объединяются, образуя аллополиплоидное потомство. В последнем случае один или несколько диплоидов F1 гибриды продуцируют невосстановленные гаметы, которые сливаются с образованием аллополиплоидного потомства.[27] Гибридизация с последующей дупликацией генома может быть более распространенным путем к аллополиплоидии, поскольку F1 гибриды между таксонами часто имеют относительно высокие темпы нередуцированного образования гамет - расхождение между геномами двух таксонов приводит к ненормальному спариванию между гомеологичный хромосомы или нерасхождение во время мейоза.[27] В этом случае аллополиплоидия действительно может восстановить нормальную, двухвалентный мейотическое спаривание путем предоставления каждой гомеологической хромосоме собственного гомолога. Если расхождение между гомеологичными хромосомами наблюдается даже в двух субгеномах, это теоретически может привести к быстрому восстановлению двухвалентного спаривания и дисомной наследования после аллополиплоидизации. Однако поливалентное спаривание является обычным для многих недавно образованных аллополиплоидов, поэтому вполне вероятно, что большая часть мейотической стабилизации происходит постепенно посредством отбора.[19][21]
Поскольку спаривание между гомеологичными хромосомами у устоявшихся аллополиплоидов встречается редко, им может быть полезен фиксированный гетерозиготность гомеологических аллелей.[28] В некоторых случаях такая гетерозиготность может быть полезной. гетеротический эффекты, будь то с точки зрения приспособленности в естественных условиях или желаемых черт в сельскохозяйственных условиях. Это могло частично объяснить преобладание аллополиплоидии среди видов сельскохозяйственных культур. Оба хлеба пшеница и Тритикале являются примерами аллополиплоидов с шестью наборами хромосом. Хлопок, арахис, или же Лебеда являются аллотетраплоидами с множественным происхождением. В Латунный посевы, Треугольник U описывает отношения между тремя распространенными диплоидными Brassicas (B. oleracea, Б. рапа, и Б. нигра ) и трех аллотетраплоидов (B. napus, B. juncea, и B. carinata ), полученный в результате гибридизации диплоидных видов. Подобные отношения существуют между тремя диплоидными видами Трагопогон (T. dubius, T. pratensis, и T. porrifolius ) и два аллотетраплоидных вида (T. mirus и T. miscellus ).[29] Сложные закономерности эволюции аллополиплоидов наблюдались также у животных, например, у рода лягушек. Xenopus.[30]
Анеуплоид
Организмы, в которых определенная хромосома или сегмент хромосомы недопредставлены или представлены в избытке, называются анеуплоид (от греческих слов, означающих «не», «хороший» и «складной»). Анеуплоидия относится к количественному изменению части хромосомного набора, тогда как полиплоидия относится к количественному изменению всего набора хромосом.[31]
Эндополиплоидия
Полиплоидия встречается в некоторых тканях животных, которые в остальном являются диплоидными, например, в тканях человека. мышца ткани.[32] Это известно как эндополиплоидия. Виды, клетки которых не имеют ядер, то есть прокариоты, может быть полиплоидным, как видно на большом бактерия Epulopiscium fishelsoni.[33] Следовательно плоидность определяется относительно ячейки.
Моноплоидный
А моноплоидный имеет только один набор хромосом, и этот термин обычно применяется только к клеткам или организмам, которые обычно являются диплоидными. Более общий термин для таких организмов: гаплоидный.
Временные условия
Неополиплоидия
Новообразованный полиплоид.
Мезополиплоидия
В недавней истории это стало полиплоидным; он не такой новый, как неополиплоид, и не такой старый, как палеополиплоид. Это полиплоид среднего возраста. Часто это относится к дупликации всего генома с последующими промежуточными уровнями диплоидизации.
Палеополиплоидия

Дупликации древних геномов, вероятно, имели место в эволюционной истории всего живого. Дублирование событий, произошедших давно в истории разных эволюционные линии может быть трудно обнаружить из-за последующих диплоидизация (так что полиплоид со временем начинает вести себя цитогенетически как диплоид) как мутации и трансляции генов постепенно делают одну копию каждой хромосомы в отличие от другой копии. Со временем дублированные копии генов также часто накапливают мутации и становятся неактивными псевдогенами.[34]
Во многих случаях об этих событиях можно судить только путем сравнения секвенированные геномы. Примеры неожиданных, но недавно подтвержденных дупликаций древнего генома включают: пекарские дрожжи (Saccharomyces cerevisiae ), горчичный кресс-салат / тале (Arabidopsis thaliana ), рис (Oryza sativa ), и ранний эволюционный предок из позвоночные (который включает человек происхождение) и еще один рядом с источником костистость Рыбы.[35] Покрытосеменные (цветущие растения ) имеют палеополиплоидию в своей родословной. Все эукариоты вероятно, в какой-то момент своей эволюционной истории пережили событие полиплоидии.
Другие похожие термины
Кариотип
Кариотип - это характерный хромосомный набор эукариот разновидность.[36][37] Подготовка и исследование кариотипов является частью цитология и, более конкретно, цитогенетика.
Хотя репликация и транскрипция ДНК очень стандартизированы в эукариоты этого нельзя сказать об их кариотипах, которые сильно различаются между видами по количеству хромосом и по детальной организации, несмотря на то, что они построены из одних и тех же макромолекул. В некоторых случаях существует даже значительное разнообразие внутри видов. Эта вариация обеспечивает основу для ряда исследований в области так называемой эволюционной цитологии.
Гомеологические хромосомы
Гомеологические хромосомы собраны вместе после межвидовая гибридизация и аллополиплоидизация, и чьи отношения были полностью гомологичными у предковых видов. Например, твердая пшеница является результатом межвидовой гибридизации двух диплоидных видов трав. Triticum urartu и Эгилопс спелтоидесный. Оба диплоидных предка имели два набора из 7 хромосом, которые были похожи по размеру и содержащимся в них генам. Твердая пшеница содержит гибридный геном с двумя наборами хромосом, полученных из Triticum urartu и два набора хромосом, полученных из Эгилопс спелтоидесный. Каждая пара хромосом происходит от Triticum urartu родитель гомеологичный к противоположной паре хромосом, полученной из Эгилопс спелтоидесный родитель, хотя каждая пара хромосом сама по себе гомологичный.
Примеры
Животные
Примеры у животных чаще встречаются у беспозвоночных.[38] Такие как плоские черви, пиявки, и рассольная креветка. Среди позвоночных примеры стабильной полиплоидии включают лососевые и много карповые (т.е. карп ).[39] У некоторых рыб насчитывается до 400 хромосом.[39] Полиплоидия также часто встречается у амфибий; например, биомедицинский род Xenopus содержит много разных видов с 12 наборами хромосом (додекаплоид).[40] Полиплоидные ящерицы также довольно обычны, но они бесплодны и должны воспроизводиться партеногенез.[нужна цитата ] Полиплоид крот саламандры (в основном триплоиды) все самки и размножаются клептогенез,[41] "воровство" сперматофоры от диплоидных самцов родственных видов, чтобы вызвать развитие яиц, но не встраивая ДНК самцов в потомство. Хотя клетки печени млекопитающих являются полиплоидными, в редких случаях полиплоидные млекопитающие известны, но чаще всего приводят к пренатальный смерть.
An октодонтид грызун из Аргентина суровый пустыня регионы, известные как равнина вискаша крыса (Tympanoctomys barrerae) рассматривается как исключение из этого «правила».[42] Однако тщательный анализ с использованием хромосомных красок показывает, что в каждой хромосоме всего по две копии. T. barrerae, а не четыре ожидаемых, если бы это был действительно тетраплоид.[43] Этот грызун не крыса, но родственник морские свинки и шиншиллы. Его «новый» диплоид (2п) число равно 102, поэтому его ячейки примерно вдвое превышают нормальный размер. Его ближайшим живым родственником является Octomys mimax, то Андский Вискача-Крыса из той же семьи, у которой 2п = 56. Поэтому было высказано предположение, что Octomys-подобный предок произвел тетраплоид (т. е. 2п = 4Икс = 112) потомство, которое в силу удвоенных хромосом было репродуктивно изолировано от родителей.
Полиплоидия была вызвана у рыб Хар Сваруп (1956), применив холодовой шок для яиц, близкий к моменту оплодотворения, в результате чего были получены успешно созревшие триплоидные эмбрионы.[44][45] Было также показано, что холод или тепловой шок приводят к нередуцированным гаметам амфибий, хотя это чаще встречается в яйцах, чем в сперме.[46] Джон Гэрдон (1958) трансплантировали интактные ядра из соматических клеток для получения диплоидных яиц лягушки, Xenopus (продолжение работы Бриггса и Кинга в 1952 г.), которые смогли развиться до стадии головастика.[47] Британский ученый Дж. Б. С. Холдейн высоко оценил потенциальное медицинское применение этой работы и, описывая результаты, стал одним из первых, кто использовал слово "клон "применительно к животным. Более поздние работы Шинья Яманака показали, как зрелые клетки могут быть перепрограммированы, чтобы стать плюрипотентными, расширяя возможности для не стволовых клеток. Гурдон и Яманака были совместно удостоены Нобелевской премии в 2012 году за эту работу.[47]
Люди
Истинная полиплоидия редко встречается у людей, хотя полиплоидные клетки встречаются в очень высокой степени. дифференцированный ткань, такая как печень паренхима, сердечная мышца, плацента и костный мозг.[1][48] Анеуплоидия встречается чаще.
Полиплоидия встречается у человека в виде триплоидия, с 69 хромосомами (иногда называемыми 69, XXX) и тетраплоидией с 92 хромосомами (иногда называемыми 92, XXXX). Триплоидия, обычно из-за полиспермия, происходит примерно в 2–3% всех беременностей у людей и ~ 15% выкидышей.[нужна цитата ] Подавляющее большинство триплоидных концепций заканчивается выкидыш; те, кто доживают до срока, обычно умирают вскоре после рождения. В некоторых случаях продолжительность жизни после рождения может быть увеличена, если миксоплоидия с обоими диплоид и присутствует популяция триплоидных клеток. Было одно сообщение о ребенке, дожившем до семи месяцев с синдромом полной триплоидии. У него не было нормального умственного или физического неонатального развития, и он умер от Pneumocystis carinii инфекция, что свидетельствует о слабой иммунной системе.[49]
Триплоидия может быть результатом либо Digyny (дополнительный гаплоидный набор от матери) или диандри (дополнительный гаплоидный набор от отца). Диандрия в основном вызвана дублированием отцовского гаплоидного набора из одного сперматозоида, но также может быть следствием диспермической (два сперматозоида) оплодотворение яйца.[50] Дигини чаще всего вызывается отказом одного мейотического деления во время оогенеза, что приводит к диплоиду. ооцит или невозможность выдавить один полярное тело от ооцит. Диандрия, кажется, преобладает среди ранних выкидыши, в то время как digyny преобладает среди триплоидных зигот, доживающих до эмбрионального периода.[нужна цитата ] Однако среди ранних выкидышей дигиния также чаще встречается в тех случаях, когда меньше8 1⁄2 неделей гестационного возраста или тех, в которых присутствует эмбрион. Также есть два разных фенотипы в триплоиде плаценты и плоды которые зависят от происхождения дополнительных гаплоидный набор. В дигини обычно бывает асимметричный, плохо выращенный плод, с отмеченным надпочечник гипоплазия и очень маленький плацента.[нужна цитата ] В диандрии частичное пузырный занавес развивается.[50] Эти эффекты родительского происхождения отражают эффекты геномный импринтинг.[нужна цитата ]
Полная тетраплоидия диагностируется реже, чем триплоидия, но наблюдается в 1-2% ранних выкидышей. Однако некоторые тетраплоидные клетки обычно обнаруживаются при анализе хромосом на пренатальная диагностика и они обычно считаются «безвредными». Неясно, возникают ли эти тетраплоидные клетки во время in vitro культура клеток или присутствуют ли они также в клетках плаценты in vivo. Во всяком случае, существует очень мало клинических сообщений о плодах / младенцах с диагнозом тетраплоидный мозаицизм.
Миксоплоидия довольно часто наблюдается у человеческих доимплантационных эмбрионов и включает гаплоидные / диплоидные, а также смешанные диплоидные / тетраплоидные популяции клеток. Неизвестно, не могут ли эти эмбрионы имплантироваться и поэтому редко обнаруживаются при продолжающейся беременности, или это просто избирательный процесс, благоприятствующий диплоидным клеткам.
Рыбы
Событие полиплоидии произошло в пределах стволовой линии костистых рыб.[35]
Растения

Полиплоидия часто встречается у растений, по некоторым оценкам, 30–80% живых видов растений являются полиплоидными, и многие линии показывают доказательства древней полиплоидии (палеополиплоидия ) в своих геномах.[51][52][53][54] Огромные взрывы в покрытосеменные Видовое разнообразие, по-видимому, совпало со временем дупликации древних геномов, общих для многих видов.[55] Установлено, что 15% покрытосеменных и 31% папоротника видообразование события сопровождаются увеличением плоидности.[56]
Полиплоидные растения могут возникать в природе спонтанно по нескольким механизмам, включая мейотические или митотические сбои и слияние невосстановленных (2п) гаметы.[57] Оба автополиплоида (например, картофель[58]) и аллополиплоиды (такие как канола, пшеница и хлопок) можно найти как среди диких, так и среди одомашненных видов растений.
Большинство полиплоидов демонстрируют новые вариации или морфологии по сравнению с их родительскими видами, что может способствовать процессам видообразование и использование эко-ниши.[52][57] Механизмы, приводящие к новым вариациям во вновь образованных аллополиплоидах, могут включать эффекты дозировки генов (в результате более многочисленных копий содержимого генома), воссоединение дивергентных регуляторных иерархий генов, хромосомные перестройки и эпигенетический ремоделирование, все из которых влияют на содержание гена и / или уровни экспрессии.[59][60][61][62] Многие из этих быстрых изменений могут способствовать репродуктивной изоляции и видообразованию. Однако семена, полученные из интерплоидия кресты, например, между полиплоидами и их родительскими видами, обычно страдают аберрантным развитием эндосперма, что снижает их жизнеспособность,[63][64] тем самым способствуя полиплоидное видообразование.
Некоторые растения триплоидны. В качестве мейоз нарушено, эти растения бесплодны, причем все растения имеют одинаковую генетическую конституцию: среди них исключительно вегетативно размножаемые шафрановый крокус (Крокус сативус). Также чрезвычайно редкий тасманский кустарник Ломатия тасманика представляет собой триплоидный стерильный вид.
Есть несколько встречающихся в природе полиплоидов. хвойные породы. Одним из примеров является прибрежный редвуд. Секвойя семпервиренс, который является гексаплоидом (6Икс) с 66 хромосомами (2п = 6Икс = 66), хотя происхождение неясно.[65]
Водные растения, особенно Однодольные, включают большое количество полиплоидов.[66]
Культуры
Индукция полиплоидии - распространенный метод преодоления стерильности гибридных видов во время селекции растений. Например, тритикале это гибрид пшеница (Triticum turgidum) и рожь (Secale cereale). Он сочетает в себе востребованные качества родителей, но исходные гибриды бесплодны. После полиплоидизации гибрид становится фертильным и, таким образом, может быть далее размножен до тритикале.
В некоторых ситуациях предпочтительны полиплоидные культуры, потому что они стерильны. Например, многие сорта плодов без косточек лишены косточек в результате полиплоидии. Такие культуры размножают бесполыми методами, такими как прививка.
Полиплоидия сельскохозяйственных культур чаще всего вызывается обработкой семян химическим веществом. колхицин.
Примеры
- Триплоидные культуры: некоторые яблоко разновидности (такие как Belle de Boskoop, Джонаголд, Муцу, Рибстон Пиппин ), банан, цитрусовые, имбирь, арбуз,[67] шафрановый крокус, белая мякоть кокоса
- Тетраплоидные культуры: очень мало яблоко сорта, твердый или же макароны пшеница, хлопок, картофель, канола /рапс, лук-порей, табак, арахис, кинноу, Пеларгония
- Гексаплоидные культуры: хризантема, хлеб пшеница, тритикале, овес, киви[8]
- Октаплоидные культуры: клубника, георгин, анютины глазки, сахарный тростник, ока (Oxalis tuberosa )[68]
- Додекаплоидные культуры: некоторые сахарный тростник гибриды[69]
Некоторые культуры встречаются в различных плоидностях: тюльпаны и лилии обычно встречаются как диплоидные, так и триплоидные; лилейники (Hemerocallis сорта) доступны как диплоидные, так и тетраплоидные; яблоки и кинноу мандарины может быть диплоидом, триплоидом или тетраплоидом.
Грибы

Помимо растений и животных, эволюционная история различных грибковые виды усеян прошлыми и недавними событиями дупликации всего генома (см. Albertin and Marullo 2012[70] для обзора). Известно несколько примеров полиплоидов:
- автополиплоид: водные грибы рода Allomyces,[71] немного Saccharomyces cerevisiae штаммы, используемые в пекарня,[72] и Т. Д.
- аллополиплоид: широко распространенный Cyathus stercoreus,[73] аллотетраплоидные лагерные дрожжи Saccharomyces pastorianus,[74] аллотриплоидные дрожжи для порчи вина Dekkera bruxellensis,[75] и Т. Д.
- палеополиплоид: патоген человека Rhizopus oryzae,[76] род Сахаромицеты,[77] и Т. Д.
Кроме того, полиплоидия часто ассоциируется с гибридизация и сетчатая эволюция, которая, по-видимому, широко распространена у нескольких таксонов грибов. В самом деле, гомоплоидное видообразование (гибридное видообразование без изменения хромосома число) было обнаружено для некоторых видов грибов (таких как базидиомицеты Microbotryum violaceum[78]).
Что касается растений и животных, гибриды и полиплоиды грибов демонстрируют структурные и функциональные модификации по сравнению с их предшественниками и диплоидными аналогами. В частности, структурные и функциональные результаты полиплоидного Сахаромицеты геномы ярко отражают эволюционную судьбу полиплоидных растений. Большие хромосомные перестройки[79] ведущий к химерный хромосомы[80] были описаны, а также более точные генетические модификации, такие как потеря гена.[81] Гомоаллели аллотетраплоидных дрожжей S. pastorianus показать неравный вклад в транскриптом.[82] Фенотипический диверсификация также наблюдается после полиплоидизации и / или гибридизации у грибов,[83] производство топлива для естественный отбор и последующие приспособление и видообразование.
Хромальвеолаты
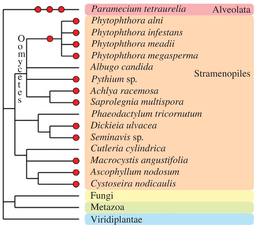
Другие эукариотические таксоны пережили одно или несколько событий полиплоидизации в течение своей эволюционной истории (см. Albertin and Marullo, 2012[70] для обзора). В оомицеты, которые не являются истинными членами грибов, содержат несколько примеров палеополиплоидных и полиплоидных видов, например, в пределах рода Фитофтора.[84] Некоторые виды коричневого водоросли (Fucales, Ламинарии[85] и диатомеи[86]) содержат очевидные полиплоидные геномы. в Альвеолаты группа, замечательный вид Парамеций тетраурелия прошел три последовательных раунда полногеномной дупликации[87] и зарекомендовал себя как основная модель для палеополиплоидных исследований.
Бактерии
Каждый Дейнококк радиодуранс бактерия содержит 4-8 экземпляров его хромосома.[88] Контакт с D. Radiodurans к рентгеновский снимок облучение или высыхание может разрушить его геномы на сотни коротких случайных фрагментов. Тем не менее, D. Radiodurans обладает высокой устойчивостью к таким воздействиям. Механизм точного восстановления генома включает RecA-опосредованный гомологичная рекомбинация и процесс, называемый расширенным зависимый от синтеза отжиг нитей (SDSA).[89]
Azotobacter vinelandii может содержать до 80 копий хромосом на клетку.[90] Однако это наблюдается только в быстрорастущих культурах, тогда как культуры, выращенные в синтетических минимальных средах, не являются полиплоидными.[91]
Археи
В Археон Halobacterium salinarium полиплоидный[92] и, как Дейнококк радиодуранс, обладает высокой устойчивостью к рентгеновскому излучению и высыханию, условиям, которые вызывают ДНК двухниточные разрывы.[93] Хотя хромосомы разбиты на множество фрагментов, полные хромосомы можно регенерировать, используя перекрывающиеся фрагменты. В механизме используется одноцепочечный ДНК-связывающий белок и вероятно гомологичный рекомбинационный ремонт.[94]
Смотрите также
Рекомендации
- ^ а б Velicky, P .; Meinhardt, G .; Plessl, K .; Vondra, S .; Weiss, T .; Haslinger, P .; Lendl, T .; Aumayr, K .; Mairhofer, M .; Чжу, X .; Schütz, B .; Hannibal, R.L .; Lindau, R .; Weil, B .; Ernerudh, J .; Neesen, J .; Egger, G .; Микула, М .; Röhrl, C .; Urban, A.E .; Baker, J .; Knöfler, M .; Поллхаймер, Дж. (2018). «Амплификация генома и клеточное старение - отличительные признаки развития плаценты человека». PLOS Genetics. 14 (10): e1007698. Дои:10.1371 / journal.pgen.1007698. ЧВК 6200260. PMID 30312291.
- ^ Оно, С .; Muramoto, J .; Christian, L .; Аткин, Н. Б. (1967). «Диплоидно-тетраплоидные отношения между старосветскими членами семейства рыб Cyprinidae». Хромосома. 23 (1): 1–9. Дои:10.1007 / BF00293307. S2CID 1181521.
- ^ Vallejo Marín, M .; Buggs, R. J. A .; Кули, А. М .; Пузи, Дж. Р. (2015). "Видообразование путем дупликации генома: повторяющееся происхождение и геномный состав недавно сформированных аллополиплоидных видов. Mimulus peregrinus". Эволюция. 69 (6): 1487–1500. Дои:10.1111 / evo.12678. ЧВК 5033005. PMID 25929999.
- ^ Фессенден, М. "Освободите место для нового цветения: обнаружен новый цветок". Scientific American. Получено 22 февраля 2017.
- ^ Бертолани, Р. (2001). «Эволюция репродуктивных механизмов тихоходок: обзор». Zoologischer Anzeiger. 240 (3–4): 247–252. Дои:10.1078/0044-5231-00032.
- ^ Stouder, D. J .; Bisson, P.A .; Найман, Р. Дж. (1997). Тихоокеанский лосось и его экосистемы: состояние и будущие варианты. Springer. С. 30–31. ISBN 978-0-412-98691-8. Получено 9 июля 2013.
- ^ Adams, K. L .; Вендел, Дж. Ф. (2005). «Полиплоидия и эволюция генома у растений». Текущее мнение в области биологии растений. 8 (2): 135–141. Дои:10.1016 / j.pbi.2005.01.001. PMID 15752992.
- ^ а б Crowhurst, R.N .; Whittaker, D .; Гарднер, Р. «Генетическое происхождение киви».
- ^ Ainouche, M. L .; Fortune, P.M .; Лосось, А .; Parisod, C .; Grandbastien, M.-A .; Фукунага, К .; Ricou, M .; Миссет, М.-Т. (2008). «Гибридизация, полиплоидия и инвазия: уроки Спартина (Poaceae) ". Биологические вторжения. 11 (5): 1159–1173. Дои:10.1007 / s10530-008-9383-2. S2CID 25522023.
- ^ Солтис, Д. Э. (1984-01-01). «Автополиплоидия в Толмия Menziesii (Saxifragaceae) ". Американский журнал ботаники. 71 (9): 1171–1174. Дои:10.2307/2443640. JSTOR 2443640.
- ^ Drauch Schreier, A .; Gille, D .; Махарджа, Б .; Мэй, Б. (2011-11-01). «Нейтральные маркеры подтверждают происхождение октоплоидов и выявляют спонтанную автополиплоидию у белого осетра, Acipenser transmontanus". Журнал прикладной ихтиологии. 27: 24–33. Дои:10.1111 / j.1439-0426.2011.01873.x. ISSN 1439-0426.
- ^ а б Bretagnolle, F .; Томпсон, Дж. Д. (1 января 1995 г.). «Гаметы с соматическим числом хромосом: механизмы их образования и роль в эволюции автополиплоидных растений». Новый Фитолог. 129 (1): 1–22. Дои:10.1111 / j.1469-8137.1995.tb03005.x. ISSN 1469-8137.
- ^ Мюнцинг, Арне (март 1936 г.). «Эволюционное значение автополиплоидии». Наследие. 21 (2–3): 363–378. Дои:10.1111 / j.1601-5223.1936.tb03204.x. ISSN 1601-5223.
- ^ Varoquaux, F .; Blanvillain, R .; Дельсены, М .; Галлуа, П. (июнь 2000 г.). «Лучше меньше, да лучше: новые подходы к производству плодов без косточек». Тенденции в биотехнологии. 18 (6): 233–242. Дои:10.1016 / s0167-7799 (00) 01448-7. ISSN 0167-7799. PMID 10802558.
- ^ Коттер, Д .; О'Донован, В .; Ó Maoiléidigh, N .; Rogan, G .; Roche, N .; Уилкинс, Н. П. (июнь 2000 г.). "Оценка использования триплоидного атлантического лосося (Salmo salar L.) в минимизации воздействия сбежавшего выращенного лосося на дикие популяции ». Аквакультура. 186 (1–2): 61–75. Дои:10.1016 / S0044-8486 (99) 00367-1.
- ^ Lincoln, R. F .; Скотт, А. П. (1983). «Производство полностью самок триплоидной радужной форели». Аквакультура. 30 (1–4): 375–380. Дои:10.1016/0044-8486(83)90179-5.
- ^ Дермен, Х. (май 1951 г.). «Тетраплоидные и диплоидные побочные эффекты: из гигантского спорта Apple McIntosh». Журнал наследственности. 42 (3): 145–149. Дои:10.1093 / oxfordjournals.jhered.a106189. ISSN 0022-1503.
- ^ Dwivedi, S.L .; Упадхьяя, Х. Д .; Сталкер, Х. Т .; Blair, M. W .; Бертиоли, Д. Дж .; Nielen, S .; Ортис, Р. (январь 2007 г.). Яник, Жюль (ред.). Отзывы о селекции растений (PDF). Джон Вили и сыновья. С. 179–230. Дои:10.1002 / 9780470380130.ch3. ISBN 9780470380130.
- ^ а б Джастин, Р. (январь 2002 г.). «Неополиплоидия цветковых растений». Ежегодный обзор экологии и систематики. 33 (1): 589–639. Дои:10.1146 / annurev.ecolsys.33.010802.150437. S2CID 41689384.
- ^ Parisod, C .; Holderegger, R .; Брохманн, К. (апрель 2010 г.). «Эволюционные последствия автополиплоидии». Новый Фитолог. 186 (1): 5–17. Дои:10.1111 / j.1469-8137.2009.03142.x. ISSN 1469-8137. PMID 20070540.
- ^ а б Le Comber, S.C .; Ainouche, M. L .; Коварик, А .; Лейтч, А. Р. (апрель 2010 г.). «Создание функционального диплоида: от полисомного к дисомному наследованию». Новый Фитолог. 186 (1): 113–122. Дои:10.1111 / j.1469-8137.2009.03117.x. ISSN 1469-8137. PMID 20028473.
- ^ Стеббинс, Г. Л. (1947). Типы полиплоидов: их классификация и значение. Успехи в генетике. 1. С. 403–429. Дои:10.1016 / с0065-2660 (08) 60490-3. ISBN 9780120176014. PMID 20259289.
- ^ Стеббинс, Г. Л. (1950). Вариации и эволюция растений. Издательство Оксфордского университета.[страница нужна ]
- ^ Ramsey, J .; Schemske, D. W. (1998-01-01). «Пути, механизмы и скорость образования полиплоидов у цветковых растений». Ежегодный обзор экологии и систематики. 29 (1): 467–501. Дои:10.1146 / annurev.ecolsys.29.1.467. S2CID 31637733.
- ^ Barker, M. S .; Arrigo, N .; Baniaga, A.E .; Ли, З .; Левин, Д. А. (01.04.2016). «Об относительной численности автополиплоидов и аллополиплоидов». Новый Фитолог. 210 (2): 391–398. Дои:10.1111 / nph.13698. ISSN 1469-8137. PMID 26439879.
- ^ Дойл, Дж. Дж .; Шерман-Бройлс, С. (01.01.2017). «Двойная беда: систематика и определения полиплоидии». Новый Фитолог. 213 (2): 487–493. Дои:10.1111 / nph.14276. ISSN 1469-8137. PMID 28000935.
- ^ а б Рэмси, Дж. (Январь 1998 г.). «Пути, механизмы и скорость образования полиплоидов у цветковых растений». Ежегодный обзор экологии и систематики. 29 (1): 467–501. Дои:10.1146 / annurev.ecolsys.29.1.467. S2CID 31637733.
- ^ Комай, Л. (ноябрь 2005 г.). «Преимущества и недостатки полиплоидности». Природа Обзоры Генетика. 6 (11): 836–846. Дои:10.1038 / nrg1711. ISSN 1471-0056. PMID 16304599. S2CID 3329282.
- ^ Оунбей, М. (январь 1950 г.). «Природная гибридизация и амфиплоидия в роде Tragopogon». Американский журнал ботаники. 37 (7): 487–499. Дои:10.2307/2438023. JSTOR 2438023.
- ^ Schmid, M .; Evans, B.J .; Богарт, Дж. П. (2015). «Полиплоидия у амфибий». Цитогенетические и геномные исследования. 145 (3–4): 315–330. Дои:10.1159/000431388. PMID 26112701.
- ^ Гриффитс, А. Дж. Ф. (1999). Введение в генетический анализ. Сан-Франциско, Калифорния: W.H. Фримен. ISBN 978-0-7167-3520-5.[страница нужна ]
- ^ Parmacek, M. S .; Эпштейн, Дж. А. (2009). «Обновление кардиомиоцитов». Медицинский журнал Новой Англии. 361 (1): 86–88. Дои:10.1056 / NEJMcibr0903347. ЧВК 4111249. PMID 19571289.
- ^ Mendell, J.E .; Clements, K. D .; Choat, J. H .; Ангерт, Э. Р. (2008). «Экстремальная полиплоидия у крупной бактерии». Труды Национальной академии наук. 105 (18): 6730–6734. Bibcode:2008ПНАС..105.6730М. Дои:10.1073 / pnas.0707522105. ЧВК 2373351. PMID 18445653.
- ^ Edger, P. P .; Пирес, К. Дж. (2009). «Дупликации генов и геномов: влияние чувствительности к дозировке на судьбу ядерных генов». Хромосомные исследования. 17 (5): 699–717. Дои:10.1007 / s10577-009-9055-9. PMID 19802709.
- ^ а б Кларк, Джон Т .; Ллойд, Грэм Т .; Фридман, Мэтт (2016-10-11). «Мало доказательств усиленной фенотипической эволюции ранних костистых насекомых по сравнению с их живой ископаемой сестринской группой». Труды Национальной академии наук. 113 (41): 11531–11536. Дои:10.1073 / pnas.1607237113. ISSN 0027-8424. ЧВК 5068283. PMID 27671652.
- ^ Уайт, М. Дж. Д. (1973). Хромосомы (6-е изд.). Лондон: Чепмен и Холл. п. 28.
- ^ Стеббинс, Г. Л. (1950). «Глава XII: Кариотип». Вариации и эволюция растений. Нью-Йорк, Нью-Йорк: издательство Колумбийского университета.[страница нужна ]
- ^ Отто, С. П .; Уиттон, Дж. (2000). «Заболеваемость и эволюция полиплоидов» (PDF). Ежегодный обзор генетики. 34 (1): 401–437. CiteSeerX 10.1.1.323.1059. Дои:10.1146 / annurev.genet.34.1.401. PMID 11092833.
- ^ а б Leggatt, R.A .; Ивама, Г. К. (2003). «Возникновение полиплоидии у рыб». Обзоры в биологии рыб и рыболовстве. 13 (3): 237–246. Дои:10.1023 / B: RFBF.0000033049.00668.fe. S2CID 11649126.
- ^ Cannatella, D.C .; Де Са, Р. О. (1993). "Xenopus laevis как модельный организм ». Общество систематических биологов. 42 (4): 476–507. Дои:10.1093 / sysbio / 42.4.476.
- ^ Bonen, L .; Bi, J. P .; Fu, K .; Noble, J .; Niedzwiecki, D. W. A .; Недзвецкий, J. (2007). «Однополые саламандры (род Амбистома) представить новый способ размножения эукариот ». Геном. 50 (2): 119–136. Дои:10.1139 / g06-152. PMID 17546077. S2CID 42528911.
- ^ Gallardo, M. H .; González, C.A .; Себриан, И. (2006). «Молекулярная цитогенетика и аллотетраплоидия у красной вискачи крысы. Tympanoctomys barrerae (Rodentia, Octodontidae) ". Геномика. 88 (2): 214–221. Дои:10.1016 / j.ygeno.2006.02.010. PMID 16580173.
- ^ Свартман, М .; Stone, G .; Станьон, Р. (2005). «Молекулярная цитогенетика отбрасывает полиплоидию у млекопитающих». Геномика. 85 (4): 425–430. Дои:10.1016 / j.ygeno.2004.12.004. PMID 15780745.
- ^ Сваруп, Х. (1956). «Производство гетероплоидии у трехиглой колюшки», Gasterosteus aculeatus (Л.) ". Природа. 178 (4542): 1124–1125. Bibcode:1956Натура.178.1124С. Дои:10.1038 / 1781124a0. S2CID 4226624.
- ^ Сваруп, Х. (1959). «Производство триплоидии в Gasterosteus aculeatus (Л.) ". Журнал генетики. 56 (2): 129–142. Дои:10.1007 / BF02984740. S2CID 42913498.
- ^ Mable, B.K .; Alexandrou, M. A .; Тейлор, М. И. (2011). «Дублирование генома у амфибий и рыб: расширенный синтез». Журнал зоологии. 284 (3): 151–182. Дои:10.1111 / j.1469-7998.2011.00829.x. S2CID 58937352.
- ^ а б «Нобелевская премия по физиологии и медицине 2012 года присуждена за открытие, что зрелые клетки могут быть перепрограммированы, чтобы стать плюрипотентными». ScienceDaily. 8 октября 2012 г.
- ^ Винкельманн, М .; Pfitzer, P .; Шнайдер, В. (1987). «Значение полиплоидии в мегакариоцитах и других клетках для здоровья и опухолевых заболеваний». Klinische Wochenschrift. 65 (23): 1115–1131. Дои:10.1007 / BF01734832. PMID 3323647. S2CID 23496028.
- ^ «Триплоидия». Национальная организация по редким заболеваниям. Получено 2018-12-23.
- ^ а б Baker, P .; Monga, A .; Бейкер, П. (2006). Гинекология от десяти учителей. Лондон: Арнольд. ISBN 978-0-340-81662-2.
- ^ Мейерс, Л. А .; Левин, Д. А. (2006). «Об изобилии полиплоидов у цветковых растений». Эволюция. 60 (6): 1198–1206. Дои:10.1111 / j.0014-3820.2006.tb01198.x. PMID 16892970.
- ^ а б Rieseberg, L.H .; Уиллис, Дж. Х. (2007). «Вид растений». Наука. 317 (5840): 910–914. Bibcode:2007Научный ... 317..910R. Дои:10.1126 / science.1137729. ЧВК 2442920. PMID 17702935.
- ^ Отто, С. П. (2007). «Эволюционные последствия полиплоидии». Клетка. 131 (3): 452–462. Дои:10.1016 / j.cell.2007.10.022. PMID 17981114. S2CID 10054182.
- ^ Инициатива One Thousand Plant Transcriptomes (2019). «Тысяча транскриптомов растений и филогеномика зеленых растений». Природа. 574 (7780): 679–685. Дои:10.1038 / s41586-019-1693-2. ЧВК 6872490. PMID 31645766.
- ^ Debodt, S .; Maere, S .; Вандепир, Ю. (2005). «Дублирование генома и происхождение покрытосеменных растений». Тенденции в экологии и эволюции. 20 (11): 591–597. Дои:10.1016 / j.tree.2005.07.008. PMID 16701441.
- ^ Wood, T. E .; Takebayashi, N .; Barker, M. S .; Mayrose, I .; Greenspoon, P. B .; Ризеберг, Л. Х. (2009). «Частота видообразования полиплоидов у сосудистых растений». Труды Национальной академии наук. 106 (33): 13875–13879. Bibcode:2009PNAS..10613875W. Дои:10.1073 / pnas.0811575106. JSTOR 40484335. ЧВК 2728988. PMID 19667210.
- ^ а б Комай, Л. (2005). «Преимущества и недостатки полиплоидности». Природа Обзоры Генетика. 6 (11): 836–846. Дои:10.1038 / nrg1711. PMID 16304599. S2CID 3329282.
- ^ Сюй, X .; Xu, S .; Pan, S .; Cheng, B .; Zhang, D .; Mu, P .; Ni, G .; Zhang, S .; Yang, R .; Li, J .; Wang, G .; Orjeda, F .; Гусман, М .; Torres, R .; Lozano, O .; Ponce, D .; Martínez, G .; De La Cruz, S.K .; Чакрабарти, В. У .; Патил, К. Г .; Скрябин, Б. Б .; Кузнецов, Н. В .; Равин, Т. В .; Колганова, А.В .; Белецкий, А. В .; Марданов, А .; Ди Генова, Д. М .; Болсер, Д. М. А .; Martin, G .; Ли Ю. (2011). «Последовательность генома и анализ клубней картофеля». Природа. 475 (7355): 189–195. Дои:10.1038 / природа10158. PMID 21743474.
- ^ Osborn, T. C .; Pires, J .; Birchler, J. A .; Auger, D. L .; Chen, Z .; Ли, Х.-С .; Comai, L .; Madlung, A .; Doerge, R.W .; Colot, V .; Мартиенссен, Р. А. (2003). «Понимание механизмов экспрессии новых генов в полиплоидах». Тенденции в генетике. 19 (3): 141–147. Дои:10.1016 / S0168-9525 (03) 00015-5. PMID 12615008.
- ^ Chen, Z. J .; Ни, З. (2006). «Механизмы геномных перестроек и изменений экспрессии генов в полиплоидах растений». BioEssays. 28 (3): 240–252. Дои:10.1002 / bies.20374. ЧВК 1986666. PMID 16479580.
- ^ Чен, З. Дж. (2007). «Генетические и эпигенетические механизмы экспрессии генов и фенотипических вариаций в полиплоидах растений». Ежегодный обзор биологии растений. 58: 377–406. Дои:10.1146 / annurev.arplant.58.032806.103835. ЧВК 1949485. PMID 17280525.
- ^ Альбертин, В .; Balliau, T .; Brabant, P .; Chèvre, A.M .; Eber, F .; Malosse, C .; Тиллеман, Х. (2006). «Многочисленные и быстрые нестохастические модификации генных продуктов в недавно синтезированных Brassica napus Аллотетраплоиды ». Генетика. 173 (2): 1101–1113. Дои:10.1534 / генетика.106.057554. ЧВК 1526534. PMID 16624896.
- ^ Pennington, P.D .; Costa, L.M .; Gutiérrez Marcos, J. F .; Гренландия, A.J .; Дикинсон, Х. Г. (апрель 2008 г.). «Когда геномы сталкиваются: аномальное развитие семян после межплоидных скрещиваний кукурузы». Анналы ботаники. 101 (6): 833–843. Дои:10.1093 / aob / mcn017. ЧВК 2710208. PMID 18276791.
- ^ Von Wangenheim, K.-H .; Петерсон, Х.-П. (2004). «Аберрантное развитие эндосперма при интерплоидных кроссах обнаруживает таймер дифференциации». Биология развития. 270 (2): 277–289. Дои:10.1016 / j.ydbio.2004.03.014. PMID 15183714.
- ^ Ахуджа, М. Р .; Нил, Д. Б. (2002). "Истоки полиплоидии в прибрежном секвойи (Секвойя семпервиренс (Д. Дон) Эндл.) И связь прибрежной секвойи с другими родами Taxodiaceae ». Silvae Genetica. 51: 2–3.
- ^ Les, D. H .; Филбрик, К. Т. (1993). «Исследования гибридизации и изменения числа хромосом у водных покрытосеменных растений: эволюционные последствия». Водная ботаника. 44 (2–3): 181–228. Дои:10.1016/0304-3770(93)90071-4.
- ^ Плоды без косточек делают другие ненужными
- ^ Эмшвиллер, Э. (2006). «Происхождение полиплоидных культур: пример культуры октаплоидных клубней. Oxalis tuberosa". В Zeder, M. A .; Decker-Walters, D .; Emshwiller, E .; Bradley, D .; Smith, B. D. (eds.). Документирование приручения: новые генетические и археологические парадигмы. Беркли, Калифорния: Калифорнийский университет Press. С. 153–168.
- ^ Le Cunff, L .; Garsmeur, O .; Raboin, L.M .; Pauquet, J .; Telismart, H .; Selvi, A .; Grivet, L .; Philippe, R .; Begum, D .; Deu, M .; Costet, L .; Wing, R .; Glaszmann, J.C .; Д'Хонт, А. (2008). «Диплоидное / полиплоидное картирование синтенических челноков и гаплотип-специфическая хромосома, идущая к гену устойчивости к ржавчине (Bru1) в высокополиплоидном сахарном тростнике (2п ∼ 12Икс ∼ 115)". Генетика. 180 (1): 649–660. Дои:10.1534 / генетика.108.091355. ЧВК 2535714. PMID 18757946.
- ^ а б c d Альбертин, В .; Марулло, П. (2012). «Полиплоидия грибов: эволюция после дупликации всего генома». Труды Королевского общества B. 279 (1738): 2497–2509. Дои:10.1098 / rspb.2012.0434. ЧВК 3350714. PMID 22492065.
- ^ Emerson, R .; Уилсон, К. М. (1954). «Межвидовые гибриды, цитогенетика и цитотаксономия Euallomyces». Микология. 46 (4): 393–434. Дои:10.1080/00275514.1954.12024382. JSTOR 4547843.
- ^ Альбертин, В .; Marullo, P .; Aigle, M .; Bourgais, A .; Белый, М .; Dillmann, C .; Де Вьенн, Д .; Сикард, Д. (2009). "Доказательства аутотетраплоидии, связанной с репродуктивной изоляцией в Saccharomyces cerevisiae: К новому одомашненному виду ". Журнал эволюционной биологии. 22 (11): 2157–2170. Дои:10.1111 / j.1420-9101.2009.01828.x. PMID 19765175.
- ^ Лу, Б. С. (1964). «Полиплоидия базидиомицетов. Cyathus stercoreus". Американский журнал ботаники. 51 (3): 343–347. Дои:10.2307/2440307. JSTOR 2440307.
- ^ Либкинд, Д .; Hittinger, C.T .; Валерио, Э .; Gonçalves, C .; Dover, J .; Johnston, M .; Gonçalves, P .; Сампайо, Дж. П. (2011). «Одомашнивание микробов и идентификация дикого генетического материала лагерных дрожжей». Труды Национальной академии наук. 108 (35): 14539–14544. Bibcode:2011PNAS..10814539L. Дои:10.1073 / pnas.1105430108. ЧВК 3167505. PMID 21873232.
- ^ Borneman, A. R .; Zeppel, R .; Чемберс, П. Дж .; Куртин, К. Д. (2014). "Взгляд на Dekkera bruxellensis Геномный ландшафт: сравнительная геномика выявила различия в плоидности и потенциале использования питательных веществ среди винных изолятов ». PLOS Genetics. 10 (2): e1004161. Дои:10.1371 / journal.pgen.1004161. ЧВК 3923673. PMID 24550744.
- ^ Ma, L.-J .; Ибрагим, А. С .; Скорый, Ц .; Grabherr, M. G .; Burger, G .; Батлер, М .; Элиас, М .; Idnurm, A .; Lang, B.F .; Соне, Т .; Abe, A .; Calvo, S.E .; Коррочано, Л. М .; Энгельс, Р .; Fu, J .; Hansberg, W .; Kim, J.-M .; Kodira, C.D .; Koehrsen, M. J .; Лю, Б .; Миранда Сааведра, Д .; О'Лири, S .; Ортис Кастелланос, L .; Poulter, R .; Rodríguez Romero, J .; Ruiz Herrera, J .; Shen, Y.-Q .; Zeng, Q .; Galagan, J .; Биррен, Б. В. (2009). Мадхани, Х. Д. (ред.). "Геномный анализ грибов основной линии Rhizopus oryzae Обнаруживает дупликацию всего генома ». PLOS Genetics. 5 (7): e1000549. Дои:10.1371 / journal.pgen.1000549. ЧВК 2699053. PMID 19578406.
- ^ Wong, S .; Батлер, G .; Вулф, К. Х. (2002). «Эволюция порядка генов и палеополиплоидия в дрожжах гемиаскомицетов». Труды Национальной академии наук. 99 (14): 9272–9277. Bibcode:2002PNAS ... 99,9272 Вт. Дои:10.1073 / pnas.142101099. JSTOR 3059188. ЧВК 123130. PMID 12093907.
- ^ Devier, B .; Aguileta, G .; Худ, М. Э .; Жиро, Т. (2009). «Использование филогении генов рецепторов феромонов в комплексе видов Microbotryum violaceum для исследования возможного видообразования путем гибридизации». Микология. 102 (3): 689–696. Дои:10.3852/09-192. PMID 20524600. S2CID 9072505.
- ^ Данн, В .; Шерлок, Г. (2008). «Реконструкция генома и эволюция гибридных лагерных дрожжей. Saccharomyces pastorianus". Геномные исследования. 18 (10): 1610–1623. Дои:10.1101 / гр.076075.108. ЧВК 2556262. PMID 18787083.
- ^ Nakao, Y .; Канамори, Т .; Ито, Т .; Kodama, Y .; Rainieri, S .; Nakamura, N .; Shimonaga, T .; Hattori, M .; Асикари, Т. (2009). "Последовательность генома дрожжей для пивоварения лагера, межвидового гибрида". ДНК исследования. 16 (2): 115–129. Дои:10.1093 / dnares / dsp003. ЧВК 2673734. PMID 19261625.
- ^ Scannell, D. R .; Byrne, K. P .; Gordon, J. L .; Wong, S .; Вулф, К. Х. (2006). «Множественные раунды видообразования, связанные с реципрокной потерей генов у полиплоидных дрожжей». Природа. 440 (7082): 341–345. Bibcode:2006Натура.440..341С. Дои:10.1038 / природа04562. HDL:2262/22660. PMID 16541074. S2CID 94448.
- ^ Минато, Т .; Yoshida, S .; Ishiguro, T .; Shimada, E .; Mizutani, S .; Кобаяши, О .; Ёсимото, Х. (2009). "Профиль экспрессии дрожжей низового брожения" Saccharomyces pastorianus ортологичные гены с использованием олигонуклеотидных микрочипов ». Дрожжи. 26 (3): 147–165. Дои:10.1002 / год.1654. PMID 19243081.
- ^ Лидзбарский, Г. А .; Школьник, Т .; Нево, Э. (2009). Иднурм, А. (ред.). "Адаптивный ответ на агенты, повреждающие ДНК в естественных Saccharomyces cerevisiae Популяции из "Каньона Эволюции", Mt. Кармель, Израиль ". PLOS ONE. 4 (6): e5914. Bibcode:2009PLoSO ... 4.5914L. Дои:10.1371 / journal.pone.0005914. ЧВК 2690839. PMID 19526052.
- ^ Ioos, R .; Andrieux, A .; Marçais, B .; Фрей, П. (2006). «Генетическая характеристика природного гибридного вида Phytophthora alni на основе ядерных и митохондриальных анализов ДНК». Грибковая генетика и биология. 43 (7): 511–529. Дои:10.1016 / j.fgb.2006.02.006. PMID 16626980.
- ^ Phillips, N .; Капраун, Д. Ф .; Gómez Garreta, A .; Ribera Siguan, M.A .; Rull, J. L .; Сальвадор Солер, N .; Lewis, R .; Каваи, Х. (2011). «Оценка содержания ядерной ДНК у 98 видов бурых водорослей (Phaeophyta)». Растения AoB. 2011: plr001. Дои:10.1093 / aobpla / plr001. ЧВК 3064507. PMID 22476472.
- ^ Чепурнов, В. А .; Mann, D.G .; Vyverman, W .; Sabbe, K .; Даниэлидис, Д. Б. (2002). "Половое размножение, система спаривания и динамика протопластов Семинавис (Bacillariophyceae) ". Журнал психологии. 38 (5): 1004–1019. Дои:10.1046 / j.1529-8817.2002.t01-1-01233.x.
- ^ Aury, J.-M .; Jaillon, O .; Duret, L .; Ноэль, Б.; Jubin, C .; Porcel, B.M .; Ségurens, B .; Daubin, V .; Anthouard, V .; Aiach, N .; Arnaiz, O .; Billaut, A .; Beisson, J .; Blanc, I .; Bouhouche, K .; Câmara, F .; Duharcourt, S .; Guigo, R .; Gogendeau, D .; Катинка, М .; Keller, A.-M .; Kissmehl, R .; Klotz, C .; Koll, F .; Le Mouël, A .; Lepère, G .; Малинский, С .; Новацкий, М .; Новак, Дж. К .; Plattner, H .; и другие. (2006). "Глобальные тенденции полногеномных дупликаций инфузорий Paramecium tetraurelia". Природа. 444 (7116): 171–178. Bibcode:2006Натура.444..171А. Дои:10.1038 / природа05230. PMID 17086204.
- ^ Хансен, М. Т. (1978). «Множественность геномных эквивалентов у радиационно-устойчивой бактерии. Микрококк радиодуранс". Журнал бактериологии. 134 (1): 71–75. Дои:10.1128 / JB.134.1.71-75.1978. ЧВК 222219. PMID 649572.
- ^ Заградка, К .; Slade, D .; Bailone, A .; Sommer, S .; Averbeck, D .; Петранович, М .; Lindner, A.B .; Радман, М. (2006). "Повторная сборка разрушенных хромосом в Дейнококк радиодуранс". Природа. 443 (7111): 569–573. Bibcode:2006Натура.443..569Z. Дои:10.1038 / природа05160. PMID 17006450. S2CID 4412830.
- ^ Nagpal, P .; Jafri, S .; Редди, М. А .; Дас, Х. К. (1989). "Множественные хромосомы Azotobacter vinelandii". Журнал бактериологии. 171 (6): 3133–3138. Дои:10.1128 / jb.171.6.3133-3138.1989. ЧВК 210026. PMID 2785985.
- ^ Мальдонадо, Р.; Хименес, Дж .; Касадесус, Дж. (1994). «Изменения плоидности во время Azotobacter vinelandii цикл роста ". Журнал бактериологии. 176 (13): 3911–3919. Дои:10.1128 / jb.176.13.3911-3919.1994. ЧВК 205588. PMID 8021173.
- ^ Соппа, Дж. (2011). «Плоидность и конверсия генов в архее». Сделки Биохимического Общества. 39 (1): 150–154. Дои:10.1042 / BST0390150. PMID 21265763. S2CID 31385928.
- ^ Kottemann, M .; Киш, А .; Iloanusi, C .; Bjork, S .; ДиРуджеро, Дж. (2005). "Психологические реакции галофильных археонов. Галобактерии sp. штамм NRC1 к обезвоживанию и гамма-облучению ». Экстремофилов. 9 (3): 219–227. Дои:10.1007 / s00792-005-0437-4. PMID 15844015. S2CID 8391234.
- ^ DeVeaux, L.C .; Müller, J. A .; Smith, J .; Петриско, Дж .; Wells, D. P .; ДасСарма, С. (2007). «Чрезвычайно радиационно-устойчивые мутанты галофильных архей с повышенной экспрессией гена одноцепочечного ДНК-связывающего белка (RPA)». Радиационные исследования. 168 (4): 507–514. Bibcode:2007РадР..168..507Д. Дои:10.1667 / RR0935.1. PMID 17903038. S2CID 22393850.
дальнейшее чтение
- Snustad, D. P .; и другие. (2006). Принципы генетики (4-е изд.). Хобокен, Нью-Джерси: John Wiley & Sons. ISBN 978-0-471-69939-2.
- В Арабидопсис Геномная инициатива (2000). «Анализ последовательности генома цветкового растения. Arabidopsis thaliana". Природа. 408 (6814): 796–815. Bibcode:2000Натура 408..796Т. Дои:10.1038/35048692. PMID 11130711.
- Икин, Г. С .; Берингер, Р. Р. (2003). «Развитие тетраплоида у мыши». Динамика развития. 228 (4): 751–766. Дои:10.1002 / dvdy.10363. PMID 14648853.
- Gaeta, R.T .; Pires, J.C .; Iniguez-Luy, F .; Леон, Э .; Осборн, Т. С. (2007). "Геномные изменения в ресинтезированных Brassica napus и их влияние на экспрессию генов и фенотип ». Растительная клетка онлайн. 19 (11): 3403–3417. Дои:10.1105 / tpc.107.054346. ЧВК 2174891. PMID 18024568.
- Gregory, T. R .; Мейбл, Б. К. (2005). «Полиплоидия у животных». В Грегори, Т. Р. (ред.). Эволюция генома. Сан-Диего, Калифорния: Эльзевир. С. 427–517.
- Jaillon, O .; Aury, J.-M .; Brunet, F .; Petit, J.-L .; Stange-Thomann, N .; Mauceli, E .; Bouneau, L .; Fischer, C .; Ozouf-Costaz, C .; Bernot, A .; Nicaud, S .; Jaffe, D .; Фишер, С .; Lutfalla, G .; Dossat, C .; Segurens, B .; Dasilva, C .; Salanoubat, M .; Леви, М .; Boudet, N .; Castellano, S .; Anthouard, V .; Jubin, C .; Кастелли, В .; Катинка, М .; Vacherie, B .; Biémont, C .; Скалли, З .; Cattolico, L .; Poulain, J .; и другие. (2004). "Дупликация генома костистых рыб Тетраодон нигровиридис выявляет ранний протокариотип позвоночных ». Природа. 431 (7011): 946–957. Bibcode:2004 Натур.431..946J. Дои:10.1038 / природа03025. PMID 15496914.
- Патерсон, А. Х .; Bowers, J. E .; Van De Peer, Y .; Вандепоэле, К. (2005). «Древнее дублирование геномов злаков». Новый Фитолог. 165 (3): 658–661. Дои:10.1111 / j.1469-8137.2005.01347.x. PMID 15720677.
- Raes, J .; Vandepoele, K .; Simillion, C .; Saeys, Y .; Ван Де Пер, Ю. (2003). "Расследование случаев древнего дублирования в Арабидопсис геном ". Журнал структурной и функциональной геномики. 3 (1–4): 117–129. Дои:10.1023 / А: 1022666020026. PMID 12836691. S2CID 9666357.
- Simillion, C .; Vandepoele, K .; Van Montagu, M.C .; Zabeau, M .; Ван Де Пир, Ю. (2002). "Скрытое дублирование прошлого Arabidopsis thaliana". Труды Национальной академии наук. 99 (21): 13627–13632. Bibcode:2002PNAS ... 9913627S. Дои:10.1073 / pnas.212522399. JSTOR 3073458. ЧВК 129725. PMID 12374856.
- Солтис, Д. Э .; Солтис, П.С.; Schemske, D. W .; Hancock, J. F .; Томпсон, Дж. Н .; Муж, Б. С .; Джадд, В. С. (2007). «Автополиплоидия покрытосеменных растений: мы сильно недооценили количество видов?». Таксон. 56 (1): 13–30. JSTOR 25065732.
- Солтис, Д. Э .; Buggs, R. J. A .; Дойл, Дж. Дж .; Солтис, П.С. (2010). «Что мы до сих пор не знаем о полиплоидии». Таксон. 59 (5): 1387–1403. Дои:10.1002 / налог.595006. JSTOR 20774036.
- Taylor, J. S .; Braasch, I .; Frickey, T .; Мейер, А .; Ван Де Пер, Ю. (2003). "Дублирование генома - черта, присущая 22 000 видов лучепёрых рыб". Геномные исследования. 13 (3): 382–390. Дои:10.1101 / гр.640303. ЧВК 430266. PMID 12618368.
- Tate, J. A .; Солтис, Д. Э .; Солтис, П.С. (2005). «Полиплоидия растений». В Грегори, Т. Р. (ред.). Эволюция генома. Сан-Диего, Калифорния: Эльзевир. С. 371–426.
- Van De Peer, Y .; Taylor, J. S .; Мейер, А. (2003). "Все ли рыбы древние полиплоиды?". Журнал структурной и функциональной геномики. 3 (1–4): 65–73. Дои:10.1023 / А: 1022652814749. PMID 12836686. S2CID 14092900.
- Ван Де Пер, Ю. (2004). «Геном тетраодона подтверждает выводы Такифугу: большинство рыб - древние полиплоиды». Геномная биология. 5 (12): 250. Дои:10.1186 / gb-2004-5-12-250. ЧВК 545788. PMID 15575976.
- Van de Peer, Y .; Мейер, А. (2005). «Крупномасштабные гены и дупликации древних геномов». В Грегори, Т. Р. (ред.). Эволюция генома. Сан-Диего, Калифорния: Эльзевир. С. 329–368.
- Wolfe, K. H .; Шилдс, Д. К. (1997). «Молекулярное свидетельство древней дупликации всего генома дрожжей». Природа. 387 (6634): 708–713. Bibcode:1997 Натур.387..708Вт. Дои:10.1038/42711. PMID 9192896. S2CID 4307263.
- Вулф, К. Х. (2001). «Вчерашние полиплоиды и загадка диплоидизации». Природа Обзоры Генетика. 2 (5): 333–341. Дои:10.1038/35072009. PMID 11331899. S2CID 20796914.
внешняя ссылка
- Полиплоидия на страницах биологии Кимбалла
- Портал полиплоидии редактируемый сообществом проект с информацией, исследованиями, образованием и библиографией о полиплоидии.
| Базовые концепты | ||
|---|---|---|
| Географические режимы | ||
| Изолирующие факторы | ||
| Гибридизация | ||
| Видообразование в таксонах | ||