Палеополиплоидия - Paleopolyploidy

Палеополиплоидия это результат дупликации генома который произошел по крайней мере несколько миллионов лет назад (MYA). Такое событие могло либо удвоить геном одного вида (автополиплоидия ) или объединить таковые двух видов (аллополиплоидия ). Из-за функционального избыточность, гены быстро заглушаются или теряются из дублированных геномов. Большинство палеополиплоидов за время эволюции потеряли полиплоид статус через процесс, называемый диплоидизация, и в настоящее время считаются диплоиды например пекарские дрожжи,[1] Arabidopsis thaliana,[2] и, возможно люди.[3][4][5][6]
Палеополиплоидия широко изучается по линиям растений. Было обнаружено, что почти все цветковые растения в какой-то момент своей эволюционной истории претерпели по крайней мере один раунд дупликации генома. Дупликации древнего генома также обнаруживаются у ранних предков позвоночных (включая человеческую родословную) рядом с происхождением костлявые рыбы, и еще один в стволовой линии костистых рыб.[7] Данные свидетельствуют о том, что пекарские дрожжи (Saccharomyces cerevisiae ), имеющий компактный геном, в своей эволюционной истории претерпел полиплоидизацию.
Период, термин мезополиплоид иногда используется для видов, которые подверглись событиям размножения всего генома (дупликация всего генома, триплификация всего генома и т. д.) в более поздней истории, например, в течение последних 17 миллионов лет.[8]
Эукариоты
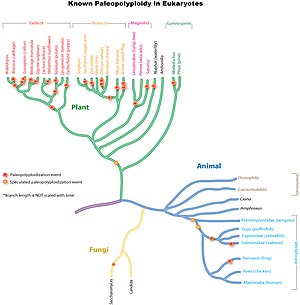
Дупликации древнего генома широко распространены повсюду. эукариотический линии, особенно у растений. Исследования показывают, что общий предок Poaceae, семейство трав, которое включает важные виды сельскохозяйственных культур, такие как кукуруза, рис, пшеница и сахарный тростник, имеет общую дупликацию генома 70 миллион лет назад.[9] В более древних линиях однодольных произошел один или, вероятно, несколько раундов дополнительных дупликаций всего генома, которые, однако, не были общими с предками. эвдикоты.[10] Дальнейшие независимые более поздние дупликации всего генома произошли в линиях, ведущих к кукурузе, сахарному тростнику и пшенице, но не к рису, сорго или просо.
Событие полиплоидии 160 миллион лет назад Предполагается, что он создал линию предков, которая привела к появлению всех современных цветковых растений.[11] Это событие палеополиплоидии было изучено путем секвенирования генома древнего цветкового растения, Amborella trichopoda.[12]
Основные эвдикоты также имели общую трипликацию всего генома (палеогексаплоидию), которая, по оценкам, произошла после однодольные -эвдикот дивергенция, но до расхождения розиды и астериды.[13][14][15] Многие виды эвдикотов испытали дополнительные дупликации или утроения всего генома. Например, модельный завод Arabidopsis thaliana, первое растение, у которого был секвенирован весь свой геном, пережило как минимум два дополнительных цикла дупликации всего генома после дупликации, характерной для основных эвдикотов.[2] Самое последнее событие произошло до расхождения Арабидопсис и Brassica родословные, о 20 миллион лет назад к 45 миллион лет назад. Другие примеры включают секвенированные геномы эвдикотов яблони, сои, томатов, хлопка и т. Д.
По сравнению с растениями палеополиплоидия в животном мире встречается гораздо реже. Он был обнаружен в основном у земноводных и костных рыб. Хотя некоторые исследования предполагают, что одна или несколько общих дупликаций генома характерны для всех позвоночных (включая человека), доказательства не так убедительны, как в других случаях, потому что дупликации, если они существуют, произошли очень давно, и этот вопрос все еще не решен. дебаты. Идея о том, что позвоночные имеют общую дупликацию всего генома, известна как Гипотеза 2R. Многих исследователей интересует причина, по которой линии животных, особенно млекопитающих, имеют гораздо меньше полных дупликаций генома, чем линии растений.
Хорошо подтвержденная палеополиплоидия была обнаружена у пекарских дрожжей (Saccharomyces cerevisiae), несмотря на небольшой компактный геном (~ 13Mbp), после расхождения с обычными дрожжами Kluyveromyces waltii.[16] Благодаря оптимизации генома дрожжи потеряли 90% дублированного генома за время эволюции и теперь признаны диплоидным организмом.
Метод обнаружения
Дублированные гены можно идентифицировать с помощью гомология последовательностей на уровне ДНК или белка. Палеополиплоидия может быть идентифицирована как одновременная массивная дупликация генов, используя молекулярные часы. Чтобы различать дупликацию всего генома и набор (более распространенных) одиночных дупликация гена событиях часто применяются следующие правила:

- Дублированные гены располагаются в больших дублированных блоках. Дублирование одного гена - это случайный процесс, который приводит к тому, что дублированные гены разбросаны по всему геному.
- Дублированные блоки не перекрываются, потому что были созданы одновременно. Сегментарное дублирование внутри генома может выполняться первое правило; но множественные независимые сегментарные дупликации могут перекрывать друг друга.
Теоретически два дублированных гена должны иметь одинаковый «возраст»; то есть дивергенция последовательности должна быть одинаковой между двумя генами, дублированными палеополиплоидией (гомеологи ). Синонимичная замена ставка, Ks, часто используются в качестве молекулярных часов для определения времени дупликации гена. Таким образом, палеополиплоидия определяется как «пик» на графике зависимости числа дубликатов от Ks (показан справа).
События дублирования, которые произошли давным-давно в истории различных эволюционных линий, может быть трудно обнаружить из-за последующей диплоидизации (такой, что полиплоид со временем начинает вести себя цитогенетически как диплоид), поскольку мутации и трансляции генов постепенно создают по одной копии каждого хромосома в отличие от своего аналога. Обычно это приводит к низкой уверенности в идентификации очень древней палеополиплоидии.
Эволюционное значение
События палеополиплоидизации приводят к массивным клеточным изменениям, включая удвоение генетического материала, изменения в экспрессии генов и увеличение размера клеток. Потеря генов во время диплоидизации не является полностью случайной, но тщательно отобранной. Дублируются гены из больших семейств генов. С другой стороны, отдельные гены не дублируются.[требуется разъяснение ] В целом палеополиплоидия может иметь как краткосрочные, так и долгосрочные эволюционные эффекты на приспособленность организма к естественной среде.
- Усиленная фенотипическая эволюция
- Дупликация всего генома может повысить скорость и эффективность приобретения микроорганизмами новых биологических свойств. Тем не менее, одна проверка этой гипотезы, которая сравнивала эволюционные темпы инноваций у ранних костистых рыб (с дублированными геномами) и ранних голостовых рыб (без дублированных геномов), обнаружила небольшую разницу между ними.[7]
- Разнообразие генома
- Удвоение генома обеспечило организм избыточными аллелями, которые могут свободно развиваться при небольшом давлении отбора. Дублированные гены могут подвергаться неофункционализация или же субфункционализация которые могут помочь организму адаптироваться к новой среде или пережить различные стрессовые условия.
- Гибридная сила
- Полиплоиды часто имеют более крупные клетки и даже более крупные органы. Многие важные культуры, включая пшеницу, кукурузу и хлопок, являются палеополиплоидами, которые были выбраны для приручения древними народами.
- Видообразование
- Было высказано предположение, что многие события полиплоидизации создали новые виды за счет приобретения адаптивных черт или из-за половой несовместимости с их диплоидными аналогами. Примером может служить недавний видообразование аллополиплоида Спартина — S. anglica; полиплоидное растение настолько успешное, что внесено в список инвазивные виды во многих регионах.
Аллополиплоидия и автополиплоидия
Есть два основных подразделения полиплоидия, аллополиплодия и автополиплоидия. Аллополиплоиды возникают в результате гибридизации двух родственных видов, в то время как автополиплоиды возникают в результате дублирования генома вида в результате гибридизации двух конспецифических родителей,[17] или соматическое удвоение репродуктивной ткани родителя. Считается, что в природе гораздо чаще встречаются аллополиплоидные виды,[17] возможно потому, что аллополиплоиды наследуют разные геномы, что приводит к увеличению гетерозиготность, а значит, и более высокая пригодность Эти разные геномы приводят к повышенной вероятности крупных реорганизаций генома,[17][18] что может быть как вредным, так и полезным. Однако автополиплоидия обычно считается нейтральным процессом.[19] хотя была выдвинута гипотеза, что автополиплоидия может служить полезным механизмом для индукции видообразования и, следовательно, способствовать способности организма быстро колонизироваться в новых средах обитания, не подвергаясь длительному и дорогостоящему периоду геномной реорганизации, который испытывают аллополиплоидные виды. Один из распространенных источников автополиплоидии у растений - это «идеальные цветы», которые способны к самоопылению или «самоопылению». Это, наряду с ошибками в мейоз что приводит к анеуплоидия, может создать среду, в которой очень высока вероятность автополиплоидии. Этот факт можно использовать в лабораторных условиях, используя колхицин подавлять хромосома сегрегация во время мейоза, создавая синтетические автополиплоидные растения.
После событий полиплоидии существует несколько возможных судеб дублированных гены; обе копии могут быть сохранены как функциональные гены, изменение функции гена может произойти в одной или обеих копиях, подавление гена может замаскировать одну или обе копии, или может произойти полная потеря гена.[17][20] События полиплоидии приведут к более высокому уровню гетерозиготности и со временем могут привести к увеличению общего числа функциональных генов в геноме. По прошествии времени после события дупликации генома многие гены изменят функцию в результате либо изменения функции дублирующих генов как для алло-, так и для автополиплоидных видов, либо будут изменения в экспрессии генов, вызванные геномными перестройками, вызванными дупликацией генома у аллополиплоидов. . Когда сохраняются обе копии гена, и, таким образом, количество копий удваивается, есть вероятность пропорционального увеличения экспрессии этого гена, что приводит к удвоению мРНК стенограмма готовится. Также существует вероятность того, что транскрипция дублированного гена будет подавляться, что приведет к менее чем двукратному увеличению транскрипции этого гена, или что событие дупликации приведет к более чем двукратному увеличению транскрипции.[21] У одного вида Глицин долихокарпа (близкий родственник соя, Глицин макс), было замечено, что после дупликации генома примерно 500 000 лет назад транскрипция увеличилась в 1,4 раза, что указывает на пропорциональное снижение транскрипции по сравнению с числом копий гена после события дупликации.[21]
Позвоночные животные как палеополиплоид
Гипотеза о палеополиплоидии позвоночных возникла еще в 1970-х годах, предложена биологом. Сусуму Оно. Он рассудил, что геном позвоночных не может достичь своей сложности без крупномасштабных дупликаций всего генома. Гипотеза «двух раундов дупликации генома» (Гипотеза 2R ) возникла и приобрела популярность, особенно среди биологов развития.
Некоторые исследователи подвергли сомнению гипотезу 2R, поскольку она предсказывает, что геномы позвоночных должны иметь соотношение генов 4: 1 по сравнению с геномами беспозвоночных, и это не подтверждается результатами 48 проектов генома позвоночных, доступными в середине 2011 года. Например, геном человека состоит из ~ 21 000 генов, кодирующих белок, согласно подсчетам в июне 2011 г. в центрах анализа генома UCSC и Ensembl.[нужна цитата ] в то время как средний размер генома беспозвоночных составляет около 15 000 генов. В амфиокс Последовательность генома подтвердила гипотезу о двух раундах дупликации всего генома с последующей потерей дублирующих копий большинства генов.[22] Дополнительные аргументы против 2R были основаны на отсутствии топологии дерева (AB) (CD) у четырех членов семейства генов у позвоночных. Однако, если бы две дупликации генома произошли близко друг к другу, мы не ожидали бы найти такую топологию.[23] Недавнее исследование привело к морская минога генетическая карта, которая дала сильную поддержку гипотезе о том, что единственная дупликация всего генома произошла в основной линии позвоночных, предшествовавшей и сопровождаемой несколькими эволюционно независимыми сегментными дупликациями, которые произошли в ходе эволюции хордовых.[24]
Смотрите также
Рекомендации
- ^ Келлис М., Биррен Б.В., Лендер Е.С. (апрель 2004 г.). «Доказательство и эволюционный анализ дупликации древнего генома дрожжей Saccharomyces cerevisiae». Природа. 428 (6983): 617–24. Bibcode:2004Натура.428..617K. Дои:10.1038 / природа02424. PMID 15004568. S2CID 4422074.
- ^ а б Бауэрс Дж. Э., Чепмен Б. А., Ронг Дж., Патерсон А. Х. (март 2003 г.). «Раскрытие эволюции генома покрытосеменных с помощью филогенетического анализа событий хромосомной дупликации». Природа. 422 (6930): 433–8. Bibcode:2003Натура.422..433Б. Дои:10.1038 / природа01521. PMID 12660784. S2CID 4423658.
- ^ Смит Дж. Дж., Кураку С., Холт С., Саука-Шпенглер Т., Цзян Н., Кэмпбелл М. С. и др. (Апрель 2013). «Секвенирование генома морской миноги (Petromyzon marinus) дает представление об эволюции позвоночных». Природа Генетика. 45 (4): 415–21, 421e1-2. Дои:10,1038 / нг.2568. ЧВК 3709584. PMID 23435085.
- ^ Wolfe KH (Май 2001 г.). «Вчерашние полиплоиды и загадка диплоидизации». Обзоры природы. Генетика. 2 (5): 333–41. Дои:10.1038/35072009. PMID 11331899. S2CID 20796914.
- ^ Блан Г, Wolfe KH (Июль 2004 г.). «Широко распространенная палеополиплоидия у модельных видов растений, вытекающая из возрастного распределения повторяющихся генов». Растительная клетка. 16 (7): 1667–78. Дои:10.1105 / tpc.021345. ЧВК 514152. PMID 15208399.
- ^ Блан Г, Wolfe KH (Июль 2004 г.). «Функциональная дивергенция дублированных генов, образованных полиплоидией в ходе эволюции Arabidopsis». Растительная клетка. 16 (7): 1679–91. Дои:10.1105 / tpc.021410. ЧВК 514153. PMID 15208398.
- ^ а б Кларк Дж. Т., Ллойд Г. Т., Фридман М. (октябрь 2016 г.). «Мало доказательств усиленной фенотипической эволюции ранних костистых насекомых по сравнению с их живой ископаемой сестринской группой». Труды Национальной академии наук Соединенных Штатов Америки. 113 (41): 11531–11536. Дои:10.1073 / pnas.1607237113. ЧВК 5068283. PMID 27671652.
- ^ Ван Х, Ван Х, Ван Дж, Сан Р., Ву Дж, Лю С. и др. (Август 2011 г.). «Геном мезополиплоидного вида сельскохозяйственных культур Brassica rapa». Природа Генетика. 43 (10): 1035–9. Дои:10,1038 / нг.919. PMID 21873998. S2CID 205358099.
- ^ Патерсон А.Х., Бауэрс Дж. Э., Чепмен Б.А. (июнь 2004 г.). «Древняя полиплоидизация, предшествовавшая дивергенции зерновых, и ее последствия для сравнительной геномики». Труды Национальной академии наук Соединенных Штатов Америки. 101 (26): 9903–8. Bibcode:2004ПНАС..101.9903П. Дои:10.1073 / pnas.0307901101. ЧВК 470771. PMID 15161969.
- ^ Тан Х, Бауэрс Дж., Ван Х, Патерсон А. Х. (январь 2010 г.). «Сравнение генома покрытосеменных позволяет выявить раннюю полиплоидию в линии однодольных». Труды Национальной академии наук Соединенных Штатов Америки. 107 (1): 472–7. Bibcode:2010ПНАС..107..472Т. Дои:10.1073 / pnas.0908007107. ЧВК 2806719. PMID 19966307.
- ^ Callaway E (декабрь 2013 г.). «Геном куста раскрывает секреты силы цветов». Природа. Дои:10.1038 / природа.2013.14426. S2CID 88293665.
- ^ Адамс К. (декабрь 2013 г.). «Геномика. Геномные ключи к исконному цветку». Наука. 342 (6165): 1456–7. Bibcode:2013Научный ... 342.1456А. Дои:10.1126 / science.1248709. PMID 24357306. S2CID 206553839.
- ^ Тан Х, Ван Х, Бауэрс Дж. Э., Мин Р., Алам М., Патерсон А. Х. (декабрь 2008 г.). «Раскрытие древней гексаплоидии через множественные генные карты покрытосеменных». Геномные исследования. 18 (12): 1944–54. Дои:10.1101 / гр.080978.108. ЧВК 2593578. PMID 18832442.
- ^ Джайон О., Оури Дж. М., Ноэль Б., Поликрити А., Клепет С., Касагранде А. и др. (Сентябрь 2007 г.). «Последовательность генома виноградной лозы предполагает наследственную гексаплоидизацию у основных типов покрытосеменных». Природа. 449 (7161): 463–7. Bibcode:2007Натура.449..463J. Дои:10.1038 / природа06148. PMID 17721507.
- ^ Тан Х, Бауэрс Дж. Э., Ван Х, Мин Р., Алам М., Патерсон А. Х. (апрель 2008 г.). «Синтения и коллинеарность в геномах растений». Наука. 320 (5875): 486–8. Bibcode:2008Научный ... 320..486T. Дои:10.1126 / science.1153917. PMID 18436778. S2CID 206510918.
- ^ Вонг С., Батлер Г., Wolfe KH (Июль 2002 г.). «Эволюция порядка генов и палеополиплоидия в дрожжах гемиаскомицетов». Труды Национальной академии наук Соединенных Штатов Америки. 99 (14): 9272–7. Bibcode:2002PNAS ... 99,9272 Вт. Дои:10.1073 / pnas.142101099. ЧВК 123130. PMID 12093907.
- ^ а б c d Солтис PS, Soltis DE (июнь 2000 г.). «Роль генетических и геномных атрибутов в успехе полиплоидов». Труды Национальной академии наук Соединенных Штатов Америки. 97 (13): 7051–7. Bibcode:2000PNAS ... 97.7051S. Дои:10.1073 / pnas.97.13.7051. ЧВК 34383. PMID 10860970.
- ^ Parisod C, Holderegger R, Brochmann C (апрель 2010 г.). «Эволюционные последствия автополиплоидии». Новый Фитолог. 186 (1): 5–17. Дои:10.1111 / j.1469-8137.2009.03142.x. PMID 20070540.
- ^ Parisod C, Holderegger R, Brochmann C (апрель 2010 г.). «Эволюционные последствия автополиплоидии». Новый Фитолог. 186 (1): 5–17. Дои:10.1111 / j.1469-8137.2009.03142.x. PMID 20070540.
- ^ Вендель Дж. Ф. (2000). Эволюция генома полиплоидов. Молекулярная биология растений. 42. С. 225–249. Дои:10.1007/978-94-011-4221-2_12. ISBN 978-94-010-5833-9. PMID 10688139.
- ^ а б Коут Дж. Э., Дойл Дж. Дж. (2010). «Количественное определение размера всего транскриптома, необходимое условие для понимания эволюции транскриптома у разных видов: пример аллополиплоида растения». Геномная биология и эволюция. 2: 534–46. Дои:10.1093 / gbe / evq038. ЧВК 2997557. PMID 20671102.
- ^ Putnam NH, Butts T, Ferrier DE, Furlong RF, Hellsten U, Kawashima T. и др. (Июнь 2008 г.). «Геном амфиоксуса и эволюция хордового кариотипа». Природа. 453 (7198): 1064–71. Bibcode:2008 Натур.453.1064P. Дои:10.1038 / природа06967. PMID 18563158.
- ^ Ферлонг РФ, Голландия PW (апрель 2002 г.). "Были ли позвоночные осьтоплоиды?". Философские труды Лондонского королевского общества. Серия B, Биологические науки. 357 (1420): 531–44. Дои:10.1098 / rstb.2001.1035. ЧВК 1692965. PMID 12028790.
- ^ Смит Дж. Дж., Кейнат МС (август 2015 г.). «Карта мейоза морской миноги улучшает разрешение дупликаций генома древних позвоночных». Геномные исследования. 25 (8): 1081–90. Дои:10.1101 / гр.184135.114. ЧВК 4509993. PMID 26048246.
дальнейшее чтение
- Адамс К.Л., Вендель Дж.Ф. (апрель 2005 г.). «Полиплоидия и эволюция генома у растений». Текущее мнение в области биологии растений. 8 (2): 135–41. Дои:10.1016 / j.pbi.2005.01.001. PMID 15752992.
- Цуй Л., Уолл П.К., Либенс-Мак Дж. Х., Линдси Б. Г., Солтис Д. Е., Дойл Дж. Дж. И др. (Июнь 2006 г.). «Широко распространенные дупликации генома на протяжении всей истории цветковых растений». Геномные исследования. 16 (6): 738–49. Дои:10.1101 / гр.4825606. ЧВК 1479859. PMID 16702410.
- Comai L (ноябрь 2005 г.). «Преимущества и недостатки полиплоидности». Обзоры природы. Генетика. 6 (11): 836–46. Дои:10.1038 / nrg1711. PMID 16304599. S2CID 3329282.
- Отто С.П., Уиттон Дж. (2000). «Заболеваемость и эволюция полиплоидов». Ежегодный обзор генетики. 34 (1): 401–437. CiteSeerX 10.1.1.323.1059. Дои:10.1146 / annurev.genet.34.1.401. PMID 11092833.
- Макаловски В. (май 2001 г.). «Мы полиплоиды? Краткая история одной гипотезы». Геномные исследования. 11 (5): 667–70. Дои:10.1101 / гр.188801. PMID 11337465.
- Келлис М., Биррен Б.В., Лендер Е.С. (апрель 2004 г.). «Доказательство и эволюционный анализ дупликации древнего генома дрожжей Saccharomyces cerevisiae». Природа. 428 (6983): 617–24. Bibcode:2004Натура.428..617K. Дои:10.1038 / природа02424. PMID 15004568. S2CID 4422074.
| Базовые концепты | ||
|---|---|---|
| Географические режимы | ||
| Изолирующие факторы | ||
| Гибридизация | ||
| Видообразование в таксонах | ||
| Геномика | |
|---|---|
| Биоинформатика | |
| Структурная биология | |
| Инструменты исследования | |
| Организации |
|
| |