Система определения пола - Sex-determination system

А система определения пола это биологический система, определяющая развитие половые признаки в организм. Наиболее организмы которые создают свои потомство с помощью половое размножение иметь два пола. Иногда встречаются гермафродиты вместо одного или обоих полов. Есть также некоторые виды, которые имеют только один пол из-за партеногенез, акт воспроизводства самки без оплодотворение.
У многих видов определение пола является генетическим: у самцов и самок разные аллели или даже разные гены которые указывают на их сексуальные морфология. У животных это часто сопровождается хромосомный различия, как правило, через комбинации XY, ZW, XO, ZO хромосомы, или гаплодиплоидия. Половая дифференциация обычно запускается основным геном («половой локус») с множеством других генов, следующих в эффект домино.
В остальных случаях пол плода определяется по экологический переменные (такие как температура ). Детали некоторых систем определения пола еще полностью не изучены. Надежды на будущий анализ биологической системы плода включают в себя инициализированные сигналы системы полного воспроизводства, которые можно измерить во время беременности, чтобы более точно определить, является ли определенный пол плода мужским или женским. Такой анализ биологических систем может также сигнализировать о том, является ли плод гермафродитом, что включает полное или частичное воспроизведение мужских и женских репродуктивных органов.
Некоторые виды, такие как различные растения и рыбы, не имеют фиксированного пола, а вместо этого проходят жизненные циклы и сменить пол основанные на генетических подсказках на соответствующих этапах жизни своего типа. Это может быть связано с факторами окружающей среды, такими как время года и температура. Гениталии человеческого плода могут иногда развиваться во время материнской беременности из-за мутаций в системе детерминизма пола плода, в результате чего плод становится интерсекс.
Открытие
Определение пола было обнаружено в хрущак мучной американского генетика Нетти Стивенс в 1903 г.[1][2][3]
Хромосомные системы
XX / XY половые хромосомы


В Система определения пола XX / XY является наиболее знакомым, поскольку встречается у людей. Система XX / XY встречается в большинстве других млекопитающие, а также некоторые насекомые. В этой системе большинство женщин имеют две половые хромосомы одного типа (XX), в то время как большинство мужчин имеют две различные половые хромосомы (XY). Половые хромосомы X и Y отличаются друг от друга по форме и размеру, в отличие от остальных хромосом (аутосомы ), иногда их называют аллосомы. У некоторых видов, таких как люди, организмы остаются безразличными к полу некоторое время после своего создания; у других же, например у плодовых мух, половая дифференциация происходит сразу после оплодотворения яйца.[4]
Определение пола по оси Y
У некоторых видов (включая человека) есть ген SRY на Y-хромосоме, которая определяет мужественность. Представители SRY-зависимых видов могут иметь необычные хромосомные комбинации XY, такие как XXY и до сих пор живу.[4]Пол человека определяется наличием или отсутствием Y-хромосомы с функциональным геном SRY. После активации гена SRY клетки создают тестостерон и антимюллеров гормон который обычно обеспечивает развитие единой мужской репродуктивной системы.[4] В типичных эмбрионах XX клетки секретируют эстроген, который направляет тело к женскому пути.
При Y-центрированном определении пола ген SRY является основным геном, определяющим мужские характеристики, но для развития семенников требуется несколько генов. У мышей XY отсутствие гена DAX1 на Х-хромосоме приводит к бесплодию, но у людей это вызывает врожденная гипоплазия надпочечников.[5] Однако, когда дополнительный ген DAX1 помещается на Х-хромосому, в результате получается женщина, несмотря на существование SRY.[6] Даже если у женщин XX есть нормальные половые хромосомы, дупликация или экспрессия SOX9 вызывает развитие яичек.[7][8] Постепенный смена пола у развитых мышей также может возникать, когда ген FOXL2 снимается с самок.[9] Хотя ген DMRT1 используется птицами в качестве локуса пола, виды с хромосомами XY также полагаются на DMRT1, содержащийся на хромосоме 9, для половой дифференциации в какой-то момент своего образования.[4]
Определение пола по Х-центру
Некоторые виды, такие как плодовые мошки, используйте наличие двух Х-хромосом для определения женственность.[10] Виды, которые используют число X для определения пола, нежизнеспособны с дополнительной X-хромосомой.
Другие варианты определения пола XX / XY
У некоторых рыб есть варианты Система определения пола XY, а также штатная система. Например, имея формат XY, Xiphophorus nezahualcoyotl и X. milleri также имеют вторую Y-хромосому, известную как Y ', которая создает XY' женщин и YY 'мужчин.[11]
Хотя бы один монотремный, то утконос, представляет особую схему определения пола, которая в некотором роде напоминает схему ZW половые хромосомы птиц и лишен гена SRY. В утконос имеет десять половых хромосом; у мужчин есть XYXYXYXYXY паттерн, в то время как у женщин десять X-хромосом. Хотя это XY-система, половые хромосомы утконоса не имеют гомологов с евтерский половые хромосомы.[12] Вместо этого гомологи с половыми хромосомами здорового человека лежат на хромосоме 6 утконоса, что означает, что половые хромосомы здорового человека были аутосомами в то время, когда монотремы отделились от терианских млекопитающих (сумчатых и белых млекопитающих). Однако гомологи птичьего DMRT1 Ген на половых хромосомах утконоса X3 и X5 предполагает, что, возможно, ген, определяющий пол утконоса, является тем же геном, который участвует в определении пола птиц. Необходимо провести дополнительные исследования, чтобы определить точный ген, определяющий пол утконоса.[13]

XX / X0 половые хромосомы
В этом варианте системы XY у женщин есть две копии половой хромосомы (XX), а у мужчин - только одна (X0). В 0 означает отсутствие второй половой хромосомы. Обычно в этом методе пол определяется количеством генов, экспрессируемых в двух хромосомах. Эта система наблюдается у ряда насекомых, в том числе у кузнечиков и сверчков отряда. Прямокрылые и у тараканов (порядок Blattodea ). У небольшого числа млекопитающих также отсутствует Y-хромосома. К ним относится колючая крыса Амами (Tokudaia osimensis ) и колючей крысы Токуносима (Tokudaia tokunoshimensis ) и Sorex araneus, а землеройка виды. Закавказские слепушонки (Эллобиус лютесценс ) также имеют форму определения ХО, при которой у обоих полов отсутствует вторая половая хромосома.[6] Механизм определения пола еще не изучен.[14]
В нематода C. elegans мужчина с одной половой хромосомой (X0); с парой хромосом (XX) - гермафродит.[15] Его главный половой ген - XOL, который кодирует XOL-1 а также контролирует экспрессию генов TRA-2 и HER-1. Эти гены соответственно снижают активацию мужских генов и увеличивают ее.[16]
Половые хромосомы ZW / ZZ
В Система определения пола ZW встречается у птиц, некоторых рептилий, некоторых насекомых и других организмов. Система определения пола ZW перевернута по сравнению с системой XY: у женщин есть два разных типа хромосомы (ZW), а у мужчин две хромосомы одного типа (ZZ). Было обнаружено, что у цыплят это зависит от экспрессии DMRT1.[17] У птиц гены FET1 и ASW находятся на W-хромосоме самок, подобно тому, как Y-хромосома содержит SRY.[4] Однако не все виды зависят от W для своего пола. Например, есть бабочки и бабочки ZW, но у некоторых были обнаружены самки с ZO, а также самки с ZZW.[15] Кроме того, в то время как млекопитающие деактивируют одну из своих дополнительных Х-хромосом у самок, оказывается, что в случае Чешуекрылые, самцы производят вдвое больше нормальных ферментов из-за наличия двух Z.[15] Поскольку определение пола ZW используется по-разному, до сих пор неизвестно, как именно большинство видов определяют свой пол.[15] Однако, как сообщается, тутовый шелкопряд Bombyx mori использует одну специфичную для женщин piRNA в качестве основного детерминатора пола.[18] Несмотря на сходство между системами ZW и XY, эти половые хромосомы развивались отдельно. В случае курицы их Z-хромосома больше похожа на человеческую аутосому 9.[19] Z-хромосома курицы также, по-видимому, связана с X-хромосомой утконоса.[20] Когда виды ZW, такие как Дракон Комодо, размножается партеногенетически, обычно производятся только самцы. Это связано с тем, что гаплоидные яйца удваивают свои хромосомы, что приводит к ZZ или WW. ZZ становятся самцами, но WW нежизнеспособны и не доводятся до срока.[21]
Половые хромосомы ZZ / Z0
В Система определения пола ZZ / Z0 встречается у некоторых бабочек. У этих насекомых одна половая хромосома, Z. Самцы имеют две Z-хромосомы, а самки - одну Z. Самцы - ZZ, а самки - Z0.[22][23][24]
УФ-половые хромосомы
В некоторых Мохообразный и немного водоросли виды, гаметофит Стадия жизненного цикла, а не гермафродит, происходит как отдельные мужские и женские особи, которые производят мужские и женские гаметы соответственно. Когда мейоз возникает в спорофит В процессе жизненного цикла половые хромосомы, известные как U и V, сортируются по спорам, которые несут либо U-хромосому и дают начало женским гаметофитам, либо V-хромосому и дают начало мужским гаметофитам.[25][26]
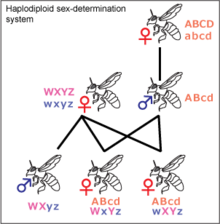
Гаплодиплоидия
Гаплодиплоидия встречается у насекомых, принадлежащих к Перепончатокрылые, такие как муравьи и пчелы. Неоплодотворенные яйца развиваются в гаплоидный особи, которыми являются самцы. Диплоид особи обычно самки, но могут быть и бесплодные самцы. Мужчины не имеют отцов и не производят сыновей, только дочерей. Если пчелиная матка спаривается с одним трутнем, ее дочери разделяют своих генов друг с другом, а не ½, как в системах XY и ZW. Это может иметь значение для развития эусоциальность, так как это увеличивает эффект родственный отбор, но это обсуждается.[27] Большинство самок отряда перепончатокрылых могут определять пол своего потомства, удерживая полученную сперму в своем сперматека и либо выпустить его в свой яйцевод, либо нет. Это позволяет им создавать больше рабочих в зависимости от статуса колонии.[28]
Экологические системы

Зависит от температуры
Существует множество других систем определения пола. У некоторых видов рептилий, в том числе аллигаторы, немного черепахи, а Tuatara, пол определяется температурой, при которой яйцо инкубируется в течение чувствительного к температуре периода. Примеров определения пола в зависимости от температуры (TSD) у птиц нет. Мегаподы ранее считалось, что это явление проявляется, но было обнаружено, что на самом деле у них разные уровни смертности эмбрионов в зависимости от температуры для каждого пола.[29] Для некоторых видов с TSD определение пола достигается воздействием более высоких температур, в результате чего потомство имеет один пол, а более низкие температуры - другого. Этот вид TSD называется Образец I. Для других видов, использующих TSD, это воздействие температуры на обоих крайних значениях, что приводит к потомству одного пола, и воздействие умеренных температур, которое приводит к потомству противоположного пола, называемое Образец II TSD. Конкретные температуры, необходимые для воспроизводства каждого пола, известны как температура, способствующая развитию самок, и температура, способствующая развитию самцов.[30] Когда температура остается около порога в течение периода, чувствительного к температуре, соотношение полов варьируется между двумя полами.[31] Температурные стандарты некоторых видов основаны на том, когда создается определенный фермент. Эти виды, которые зависят от температуры для определения пола, не имеют Ген SRY, но имеют другие гены, такие как DAX1, DMRT1, и SOX9 выраженные или невыраженные в зависимости от температуры.[30] Пол некоторых видов, например Нильская тилапия, Австралийская сцинковая ящерица, и Австралийская ящерица-дракон, изначально определяется хромосомами, но впоследствии может изменяться температурой инкубации.[11]
Неизвестно, как именно развилось определение пола в зависимости от температуры.[32] Он мог развиться благодаря тому, что определенные полы больше подходили к определенным местам, которые соответствовали температурным требованиям. Например, более теплая область может быть более подходящей для гнездования, поэтому будет производиться больше самок, чтобы увеличить количество гнездящихся в следующем сезоне.[32] Определение пола в окружающей среде предшествовало генетически детерминированным системам птиц и млекопитающих; считается, что температурно-зависимый амниот был общий предок амниот с половыми хромосомами.[33]
Другие системы
Есть другие определение пола по окружающей среде системы, включая системы определения местоположения, как показано в морском черве Bonellia viridis - личинки становятся самцами, если они вступают в физический контакт с самкой, и самками, если они оказываются на голом морском дне. Это вызвано наличием химического вещества, вырабатываемого самками, Bonellin.[34] Некоторые виды, например некоторые улитки, практика смена пола: взрослые сначала становятся мужчиной, затем становятся женщиной. В тропическом рыба-клоун, доминирующая особь в группе становится самкой, остальные - самцами, а голубоголовые губаны (Thalassoma bifasciatum ) наоборот. У некоторых видов, однако, нет системы определения пола. Виды гермафродитов включают обыкновенного дождевого червя и некоторые виды улиток. Некоторые виды рыб, рептилий и насекомых размножаются партеногенез и в целом женщины. Есть некоторые рептилии, такие как удав и Дракон Комодо которые могут воспроизводиться как половым, так и бесполым способом, в зависимости от наличия пары.[35]
К другим необычным системам относятся системы рыба-меч[требуется разъяснение ];[11] то Хирономус мошки[требуется разъяснение ][нужна цитата ]; то ювенильный гермафродитизм из данио, с неизвестным триггером;[11] и плоская рыба, который имеет W, X и Y хромосомы. Это позволяет женщинам WY, WX или XX и мужчинам YY или XY.[11]
Эволюция
Происхождение половых хромосом
Принятая гипотеза эволюции половых хромосом XY и ZW состоит в том, что они развивались одновременно, в двух разных ветвях.[36][37] Однако есть некоторые свидетельства того, что могли быть переходы между ZW и XY, например, в Xiphophorus maculatus, которые имеют системы ZW и XY в одной популяции, несмотря на то, что ZW и XY имеют разные местоположения генов.[38][39] Недавняя теоретическая модель поднимает возможность как переходов между системами XY / XX и ZZ / ZW, так и определения пола в окружающей среде.[40] Гены утконоса также подтверждают возможную эволюционную связь между XY и ZW, потому что у них есть DMRT1 ген, которым обладают птицы на их Х-хромосомах.[41] Тем не менее, XY и ZW следуют аналогичным маршрутом. Все половые хромосомы возникли как оригинальная аутосома оригинальной амниоты, которая зависела от температуры для определения пола потомства. После разделения млекопитающих ветвь разделилась на Лепидозаврия и Архозавроморфа. Обе эти две группы развили систему ZW по отдельности, о чем свидетельствует наличие разных половых хромосом.[37] У млекопитающих одна из пары аутосом, теперь Y, мутировала свою SOX3 ген в SRY ген, заставляющий эту хромосому определять пол.[37][41][42] После этой мутации SRY-содержащая хромосома перевернутый и больше не было полностью гомологичный со своим партнером. Регионы Икс и Y-хромосомы которые все еще гомологичны друг другу, известны как псевдоавтосомальная область.[43] После инвертирования Y-хромосома стала неспособной исправить вредные мутации, и, таким образом, выродившийся.[37] Есть некоторые опасения, что Y-хромосома еще больше сократится и перестанет функционировать через десять миллионов лет: но Y-хромосома была строго консервативной после первоначальной быстрой потери гена.[44][45]
Есть некоторые виды, такие как Медака рыбы, у которых половые хромосомы развивались отдельно; их Y-хромосома никогда не инвертируется и все еще может обмениваться генами с X. Половые хромосомы этих видов относительно примитивны и неспециализированы. Поскольку Y не имеет специфичных для мужчин генов и может взаимодействовать с X, XY и YY могут образовываться самки, а также самцы XX.[11] Неинвертированные Y-хромосомы с длинной историей встречаются у питоны и эму, возраст каждой системы превышает 120 миллионов лет, что позволяет предположить, что инверсии не обязательно случаются.[46]
Смотрите также
- Кларенс Эрвин МакКлунг, открывшие роль хромосом в определении пола.
- Фактор, определяющий яичко
- Материнское влияние на определение пола
- Последовательный гермафродитизм
- Определение пола и дифференциация (человек)
- Тетрахимена иметь семь полов
- Schizophyllum commune иметь 23,328 полов
использованная литература
- ^ "Нетти Стивенс: первооткрыватель половых хромосом | Изучение науки в Scitable". www.nature.com. Получено 7 июн 2018.
- ^ Огилви, Мэрилин Бейли; Шокетт, Клиффорд Дж. (1981). "Нетти Мария Стивенс (1861–1912): ее жизнь и вклад в цитогенетику". Труды Американского философского общества. 125 (4): 292–311. JSTOR 986332. PMID 11620765.
- ^ "Нетти Мария Стивенс (1861–1912) | Энциклопедия проекта" Эмбрион ". embryo.asu.edu. Получено 7 июн 2018.
- ^ а б c d е Хейк, Лаура (2008). «Генетические механизмы определения пола». Природное образование. 1 (1). Получено 8 декабря 2011.
- ^ Goodfellow, P.N .; Камерино, Г. (1999). «DAX-1,« антитестический »ген». Клеточные и молекулярные науки о жизни. 55 (6–7): 857–863. Дои:10.1007 / PL00013201. PMID 10412368. S2CID 19764423.
- ^ а б Чандра, Х.С. (25 апреля 1999 г.). «Другой способ взглянуть на загадку определения пола в Ellobius lutescens». Текущая наука. 76 (8): 1072.
- ^ Кокс, Джеймс Дж .; Willatt, L; Homfray, T; Вудс, К. Г. (6 января 2011 г.). «Дупликация SOX9 и семейное 46, XX Расстройство развития яичек». Медицинский журнал Новой Англии. 364 (1): 91–93. Дои:10.1056 / NEJMc1010311. PMID 21208124.
- ^ Хуанг, Бинг; Ван, S; Нин, Й; Lamb, A. N .; Бартли, Дж. (7 декабря 1999 г.). «Аутосомная смена пола XX, вызванная дупликацией SOX9». Американский журнал медицинской генетики. 87 (4): 349–353. Дои:10.1002 / (SICI) 1096-8628 (19991203) 87: 4 <349 :: AID-AJMG13> 3.0.CO; 2-N. PMID 10588843.
- ^ Uhlenhaut, Henriette N .; Якоб, S; Анлаг, К; Eisenberger, T; Секидо, Р; Kress, J; Treier, A.C .; Клугманн, К; Класен, С; Holter, N. I .; Ритмахер, Д; Schütz, G; Куни, А. Дж .; Ловелл-Бэдж, R; Трейер, М. (11 декабря 2009 г.). «Соматическое половое перепрограммирование взрослых яичников в яички с помощью абляции FOXL2». Ячейка. 139 (6): 1130–1142. Дои:10.1016 / j.cell.2009.11.021. PMID 20005806.
- ^ Penalva, Луис О. Ф .; Санчес (сентябрь 2003 г.). «РНК-связывающий белок, смертельный для секса (Sxl) и контроль определения пола дрозофилы и компенсации дозировки». Обзоры микробиологии и молекулярной биологии. 67 (3): 343–359. Дои:10.1128 / MMBR.67.3.343-359.2003. ЧВК 193869. PMID 12966139.
- ^ а б c d е ж Шартль, Манфред (июль 2004 г.). «Сравнительный взгляд на определение пола в медаке». Механизмы развития. 121 (7–8): 639–645. Дои:10.1016 / j.mod.2004.03.001. PMID 15210173. S2CID 17401686.
- ^ Warren, W.C .; Hillier, Ladeana W .; Маршалл Грейвс, Дженнифер А .; Бирни, Юэн; Понтинг, Крис П .; Грюцнер, Франк; Белов, Катерина; Миллер, Уэбб; и другие. (2008). «Анализ генома утконоса показывает уникальные признаки эволюции». Природа. 453 (7192): 175 – U1. Bibcode:2008 Натур.453..175Вт. Дои:10.1038 / природа06936. ЧВК 2803040. PMID 18464734.
- ^ Gruetzner, F .; Т. Эшли; Д. М. Роуэлл и Дж. А. М. Грейвс (2006). «Анализ утконоса показывает уникальные признаки эволюции». Хромосома. 115 (2): 75–88. Дои:10.1007 / s00412-005-0034-4. PMID 16344965. S2CID 23603889.
- ^ Куроива А., Ханда С., Нишияма С., Чиба Е., Ямада Ф., Абэ С., Мацуда Ю. (8 июня 2011 г.). "Дополнительные копии CBX2 в геномах самцов млекопитающих без SRY, колючей крысы Amami (Tokudaia osimensis) и колючей крысы Токуносима (Tokudaia tokunoshimensis)". Хромосома Res. 19 (5): 635–44. Дои:10.1007 / s10577-011-9223-6. PMID 21656076. S2CID 23311263.
- ^ а б c d (Majerus 2003, п. 60)
- ^ Патрисия Э. Кувабара; Питер Г. Оккема; Джудит Кимбл (апрель 1992 г.). «tra-2 кодирует мембранный белок и может опосредовать клеточную коммуникацию в пути определения пола Caenorhabditis elegans». Молекулярная биология клетки. 3 (4): 461–73. Дои:10.1091 / mbc.3.4.461. ЧВК 275596. PMID 1498366.
- ^ Smith, C.A .; Roeszler, K. N .; Ohnesorg, T .; Cummins, D. M .; Farlie, P.G .; Doran, T. J .; Синклер, А. Х. (сентябрь 2009 г.). «Птичий Z-связанный ген DMRT1 необходим для определения мужского пола у цыплят». Природа. 461 (7261): 267–271. Bibcode:2009Натура.461..267S. Дои:10.1038 / природа08298. PMID 19710650. S2CID 4413389.
- ^ Киучи, Такаши; Кога, Хикару; Кавамото, Мунетака; Сёдзи, Кейсуке; Сакаи, Хироки; Араи, Юджи; Исихара, Генки; Каваока, Синпей; Сугано, Сумио; Шимада, Тору; Судзуки, Ютака; Сузуки, Масатака; Кацума, Сусуму (14 мая 2014 г.). «Единственная специфическая для женщин piRNA является основным детерминатором пола тутового шелкопряда». Природа. 509 (7502): 633–636. Bibcode:2014Натура.509..633K. Дои:10.1038 / природа13315. PMID 24828047. S2CID 205238635.
- ^ Stiglec, R .; Ezaz, T .; Грейвс, Дж. А. (2007). «Новый взгляд на эволюцию половых хромосом птиц». Cytogenet. Genome Res. 117 (1–4): 103–9. Дои:10.1159/000103170. PMID 17675850. S2CID 12932564.
- ^ Grützner, F .; Ренс, Вт .; Ценд-Аюш, Э .; El-Mogharbel, N .; O'Brien, P. C. M .; Jones, R.C .; Фергюсон-Смит, М. А. и Маршалл, Дж. А. (2004). «У утконоса мейотическая цепь из десяти половых хромосом имеет общие гены с Z-хромосомами птиц и X-хромосом млекопитающих». Природа. 432 (7019): 913–917. Bibcode:2004Натура432..913Г. Дои:10.1038 / природа03021. PMID 15502814. S2CID 4379897.
- ^ «Девственные роды для гигантских ящериц». Новости BBC. 20 декабря 2006 г.. Получено 13 марта 2008.
- ^ Traut, W .; Sahara, K .; Марек, Ф. (2007). «Половые хромосомы и определение пола у Lepidoptera». Половое развитие. 1 (6): 332–346. Дои:10.1159/000111765. PMID 18391545. S2CID 6885122.
- ^ «Генетические механизмы определения пола - изучите науку в Scitable». www.nature.com.
- ^ Handbuch Der Zoologie / Справочник по зоологии. Вальтер де Грюйтер. 1925 г. ISBN 9783110162103 - через Google Книги.
- ^ Бахтрог Д., Киркпатрик М., Манк Дж. Э., МакДэниел С. Ф., Пирес Дж. К., Райс В., Валенсуэла Н. (сентябрь 2011 г.). «Все ли половые хромосомы созданы равными?». Тенденции Genet. 27 (9): 350–7. Дои:10.1016 / j.tig.2011.05.005. PMID 21962970.
- ^ Renner, S. S .; Heinrichs, J .; Соуза, А. (2017). «Половые хромосомы мохообразных: недавние открытия, открытые вопросы и повторные исследования Frullania dilatata и Plagiochila asplenioides». Журнал систематики и эволюции. 55 (4): 333–339. Дои:10.1111 / jse.12266.
- ^ Уилсон, Эдвард О. (12 сентября 2005 г.). «Родственный отбор как ключ к альтруизму: его взлеты и падения». Социальные исследования. 72: 1–8. Получено 25 марта 2011.
- ^ ван Вильгенбург, Эллен; Дриссен, Жерар; Beukeboom, Leow (5 января 2006 г.). "Дополнительное определение пола по одному локусу у перепончатокрылых:" неразумный "замысел?". Границы зоологии. 3 (1): 1. Дои:10.1186/1742-9994-3-1. ЧВК 1360072. PMID 16393347.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Гёт, Энн; Бут, Дэвид Т. (22 марта 2005 г.). «Температурное соотношение полов у птицы». Письма о биологии. 1 (1): 31–33. Дои:10.1098 / rsbl.2004.0247. ЧВК 1629050. PMID 17148121.
- ^ а б Торрес Мальдонадо Л.С., Ланда Пьедра А., Морено Мендоса Н., Мармолехо Валенсия А., Меза Мартинес А., Торговец Лариос Н. (октябрь 2002 г.). «Профили экспрессии Dax1, Dmrt1 и Sox9 во время определения пола температуры в гонадах морской черепахи Lepidochelys olivacea». Gen. Comp. Эндокринол. 129 (1): 20–6. Дои:10.1016 / с0016-6480 (02) 00511-7. PMID 12409092.
- ^ Булл, Дж. Дж. (Март 1980 г.). «Определение пола у рептилий». Ежеквартальный обзор биологии. 55 (1): 3–21. Дои:10.1086/411613. JSTOR 2826077. S2CID 85177125.
- ^ а б Валенсуэла, Николь; Янзен, Фредрик Дж. (2001). «Филопатрия гнездовых участков и эволюция определения пола в зависимости от температуры» (PDF). Исследования эволюционной экологии. 3: 779–794. Получено 7 декабря 2011.
- ^ Janzen, F.J .; Филлипс, П. К. (12 мая 2006 г.). «Изучение эволюции определения пола в окружающей среде, особенно у рептилий». Журнал эволюционной биологии. 19 (6): 1775–1784. Дои:10.1111 / j.1420-9101.2006.01138.x. PMID 17040374. S2CID 15485510.
- ^ Гилберт, Скотт (2006). Биология развития (8-е изд.). Сандерленд, Массачусетс: Издательство Sinauer Associates, Inc. стр.550 –553. ISBN 9780878932504.
- ^ Уоттс, Филип С .; Булей, Кевин Р .; Сандерсон, Стефани; Бордман, Уэйн; Киофи, Клаудио и Гибсон, Ричард (21 декабря 2006 г.). «Партеногенез у драконов Комодо». Природа. 444 (7122): 1021–1022. Bibcode:2006 Натур.444.1021W. Дои:10.1038 / 4441021a. PMID 17183308. S2CID 4311088.
- ^ Намекава, Сатоши; Ли, Джинни Т. (2009). "XY и ZW: Является ли инактивация мейотической половой хромосомы правилом эволюции?". PLOS Genetics. 5 (5): 3. Дои:10.1371 / journal.pgen.1000493. ЧВК 2679206. PMID 19461890.
- ^ а б c d Валлендер, Эрик; Лан, Б. Т. (28 ноября 2006 г.). «Множественное независимое происхождение половых хромосом у амниот». Труды Национальной академии наук. 103 (5): 18031–2. Bibcode:2006ПНАС..10318031В. Дои:10.1073 / pnas.0608879103. ЧВК 1838700. PMID 17116892.
- ^ Грейвс, Дженнифер (1 сентября 2000 г.). "Y-хромосома человека, определение пола и сперматогенез - взгляд феминисток". Биология размножения. 63 (3): 667–676. Дои:10.1095 / биолрепрод63.3.667б. PMID 10952906.
- ^ Эзаз, Тарик; Стиглек, Рами; Вейрунес, Фредерик; Маршалл Грейвс, Дженнифер А. (5 сентября 2006 г.). «Взаимосвязь между ZW позвоночных и системой XY половой хромосомы». Текущая биология. 16 (17): R736–43. Дои:10.1016 / j.cub.2006.08.021. PMID 16950100. S2CID 18864471.
- ^ Quinn, A.E .; Стивен Д. Сарре; Дженнифер А. Маршалл Грейвс; Артур Жорж; Жорж А. (6 января 2011 г.). «Эволюционные переходы между механизмами определения пола у позвоночных». Письма о биологии. 7 (3): 443–8. Дои:10.1098 / rsbl.2010.1126. ЧВК 3097877. PMID 21212104.
- ^ а б Грейвс, Дженнифер (10 марта 2006 г.). «Специализация и дегенерация половых хромосом у млекопитающих». Ячейка. 124 (5): 901–914. Дои:10.1016 / j.cell.2006.02.024. PMID 16530039. S2CID 8379688.
- ^ «Эволюция половых хромосом: шаг за шагом» (Пресс-релиз). Медицинский центр Чикагского университета. 28 октября 1999 г.. Получено 23 октября 2011.
- ^ Чарльзуорт, Брайан (14 августа 2003 г.). «Организация и эволюция Y-хромосомы человека». Геномная биология. 4 (9): 226. Дои:10.1186 / gb-2003-4-9-226. ЧВК 193647. PMID 12952526.
- ^ Грейвс, Дженнифер (22 июля 2004 г.). «Вырожденная Y-хромосома - может ли конверсия спасти ее?». Размножение, фертильность и развитие. 16 (5): 527–34. Дои:10.1071 / RD03096. PMID 15367368. S2CID 23740483.
- ^ Хьюз Дж. Ф. и др. (22 февраля 2012 г.). «Строгая эволюционная консервация последовала за быстрой потерей генов на Y-хромосомах человека и резуса». Природа. 483 (7387): 82–86. Bibcode:2012Натура 483 ... 82H. Дои:10.1038 / природа10843. ЧВК 3292678. PMID 22367542.
- ^ Bachtrog D, Mank JE, Peichel CL, Kirkpatrick M, Otto SP, Ashman TL, Hahn MW, Kitano J, Mayrose I, Ming R, Perrin N, Ross L, Valenzuela N, Vamosi JC, Tree of Sex Consortium (июль 2014 г.) . «Определение пола: почему так много способов сделать это?». PLOS Биология. 12 (7): e1001899. Дои:10.1371 / journal.pbio.1001899. ЧВК 4077654. PMID 24983465.
Список используемой литературы
- Майерус, М. Э. Н. (2003). Секс-войны: гены, бактерии и предвзятое соотношение полов. Издательство Принстонского университета. п. 250. ISBN 978-0-691-00981-0. Получено 4 ноября 2011.CS1 maint: ref = harv (ссылка на сайт)
- Бёкебум, Л. и Перрин, Н. (2014). Эволюция определения пола. Издательство Оксфордского университета. Интернет-ресурсы.
| Биологические термины | |
|---|---|
| Сексуальный воспроизведение | |
| Сексуальность | |
| |