Гаплодиплоидия - Haplodiploidy
Гаплодиплоидия это система определения пола в котором самцы развиваются из неоплодотворенных яиц и гаплоидный, а самки развиваются из оплодотворенных яиц и диплоид.[1] Гаплодиплоидию иногда называют арренотоки.
Гаплодиплоидия определяет пол у всех представителей отрядов насекомых. Перепончатокрылые (пчелы, муравьи, и осы )[2] и Thysanoptera ('трипсы').[3] Система также возникает спорадически в некоторых паутинный клещ, Hemiptera, Жесткокрылые (короеды ), и коловратки.
В этой системе пол определяется количеством наборов хромосомы человек получает. Потомство, образовавшееся от союза сперма и яйцо развивается как самка, а неоплодотворенная яйцеклетка развивается как самец. Это означает, что у мужчин половина хромосом меньше, чем у женщин, и они гаплоидный.
Гаплодиплоидная система определения пола имеет ряд особенностей. Например, мужчина не имеет отца и не может иметь сыновей, но у него есть дедушка и могут быть внуки. Кроме того, если эусоциальный -в колонии насекомых только одна матка, и она спаривалась только один раз, тогда родство между рабочими (диплоидными самками) в улье или гнезде3⁄4. Это означает, что рабочие в таких моногамных колониях с одной маткой значительно более тесно связаны между собой, чем в других системах определения пола, где родство братьев и сестер обычно не превышает1⁄2. Именно этот момент движет родственный отбор теория эволюции эусоциальности.[4] Действительно ли гаплодиплоидия проложила путь для эволюции эусоциальности, все еще остается предметом споров.[5][6]
Еще одна особенность системы гаплодиплоидии - рецессивный летальные и вредоносные аллели будут быстро удалены из популяции, потому что они будут автоматически выражаться в самцах (доминантные летальные и вредоносные аллели удаляются из популяции каждый раз, когда они возникают, поскольку они убивают любую особь, в которой возникают).[3]
Гаплодиплоидия - это не то же самое, что Система определения пола X0. При гаплодиплоидии мужчины получают половину хромосом, которые получают женщины, включая аутосомы. В системе определения пола X0 мужчины и женщины получают равное количество аутосом, но когда дело доходит до половых хромосом, женщины получают две X-хромосомы, а мужчины - только одну X-хромосому.
Механизмы
Было предложено несколько моделей генетических механизмов определения пола гаплодиплоидами. Чаще всего упоминается модель модель комплементарного аллеля. Согласно этой модели, если человек гетерозиготный для определенного локус, он превращается в женщину, тогда как гемизиготный и гомозиготный особи развиваются в самцов. Другими словами, диплоид потомство развивается из оплодотворенных яиц и обычно является самками, в то время как гаплоидные потомки развиваются в самцов из неоплодотворенных яиц. диплоидные самцы будут бесплодны, поскольку их клетки не будут подвергаться воздействию мейоз для образования спермы. Следовательно, сперма будет диплоидной, а это значит, что их потомство будет триплоид. Поскольку мать и сыновья перепончатокрылых имеют одни и те же гены, они могут быть особенно чувствительны к инбридинг: Инбридинг снижает количество разнополых аллели присутствует в популяции, следовательно, увеличивается количество диплоидных самцов.
После вязка, каждая плодовитая самка перепончатокрылых хранит сперма во внутреннем мешочке, называемом сперматека. Спарившаяся самка контролирует выделение накопленной спермы изнутри органа: если она выпускает сперму, когда яйцеклетка проходит по ней яйцевод, яйцо оплодотворяется.[7]Социальные пчелы, осы и муравьи могут изменять соотношение полов в колониях, что максимизирует родство между членами и создает рабочую силу, соответствующую окружающим условиям.[8] У других одиночных перепончатокрылых самки откладывают неоплодотворенные яйца самцов на более бедных источниках пищи, а оплодотворенные самки откладывают на более качественные источники пищи, возможно, потому, что нехватка самок в раннем возрасте отрицательно скажется на их пригодности.[9][10] Манипуляция соотношением полов также практикуется гаплодиплоидами. жуки-амброзии, которые откладывают больше самцов яиц, когда у самцов больше шансов разойтись и спариться с самками в разных местах.[11]
Определение пола у медоносных пчел

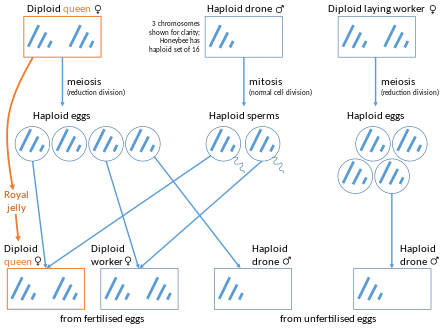
В пчелы, то дроны (самцы) полностью произошли от Королева, их мама. У диплоидной королевы 32 хромосомы, а у гаплоидных трутней - 16 хромосом. Дроны производят сперма клетки, которые содержат весь свой геном, поэтому все сперматозоиды генетически идентичны, за исключением мутаций. Следовательно, генетический состав пчел-самцов полностью происходит от матери, в то время как генетический состав рабочих пчел наполовину происходит от матери, а наполовину от отца.[12] Таким образом, если пчелиная матка спаривается только с одним трутнем, любые две из ее дочерей в среднем разделяют3⁄4 их генов. Геном диплоидной королевы рекомбинированный для ее дочерей, но гаплоидный геном отца наследуется его дочерьми «как есть». Также возможно несушка рабочая пчела откладывать неоплодотворенное яйцо, которое всегда бывает самцом.
Встречаются редкие экземпляры диплоидных личинок трутня. Это явление обычно возникает, когда существует более двух поколений браков между братьями и сестрами.[13] Определение пола у медоносных пчел изначально происходит из-за одного локуса, называемого дополнительным детерминатором пола (csd) ген. В развивающихся пчелах, если условия таковы, что особь гетерозиготна по csd ген, они разовьются в самок. Если условия таковы, что человек гемизиготный или гомозиготный по csd ген, они разовьются в мужчин. Примеры, когда особь гомозиготна по этому гену, - это экземпляры диплоидных самцов.[14] Диплоидные самцы не доживают до взрослой жизни, так как пчелы-медсестры поедают диплоидных самцов при вылуплении.[15]
В то время как рабочие могут откладывать неоплодотворенные яйца, которые становятся их сыновьями, гаплодиплоидная система определения пола увеличивает индивидуальные возможности. фитнес из-за косвенного отбора. Поскольку рабочий больше связан с дочерьми королевы (ее сестрами), чем со своим собственным потомством, помощь потомству королевы в выживании способствует распространению тех же генов, которыми обладает рабочий, более эффективно, чем прямое размножение.[16]
Партии рабочих пчел недолговечны и постоянно заменяются следующей партией, так что это родственный отбор возможно, это стратегия, обеспечивающая правильную работу улья. Однако, поскольку матки обычно спариваются с дюжиной трутней или больше, не все рабочие - полные сестры. Из-за раздельного хранения трутневой спермы определенная партия расплода может быть более тесно связана, чем определенная партия выводка, отложенная позже. Однако многие другие виды пчел, в том числе шмели, такие как Бомб террестрис, находятся чудовищный.[17] Это означает, что сестры почти всегда более связаны друг с другом, чем со своим потомством, что устраняет конфликт переменного родства, присущий пчелам.[18]
Соотношения родства в гаплодиплоидии
Родство используется для расчета прочности родственный отбор (через Правило Гамильтона ).[19] Гипотеза гаплодиплоидии предполагает, что необычный3⁄4 коэффициент родства среди полных гаплодиплоидных сестер отвечает за частоту эволюции эусоциальный поведение у перепончатокрылых.[20] Социальный работник, помогающий своей матери родить больше сестер, передает больше собственных генов, чем она воспроизвела сама.
В нормальном половое размножение, у отца два набора хромосом, и пересекая происходит между хроматиды каждой пары во время мейоз который производит сперма. Следовательно, сперма не идентична, потому что в каждой хромосоме пары будут разные аллели во многих места. Но когда отец гаплоидный все сперматозоиды идентичны (за исключением небольшого числа, где генные мутации произошли в линия зародыша ). Итак, все потомство женского пола наследует хромосомы самца на 100% неповрежденными. Пока самка спаривалась только с одним самцом, все ее дочери имеют полный набор хромосом этого мужчины. У перепончатокрылых самцы обычно производят достаточно спермы, чтобы хватило самке на всю ее жизнь после единственного спаривания с этим самцом.[19]
Коэффициенты родства у гаплодиплоидных организмов следующие, предполагая, что самка спаривалась только один раз. Эти соотношения применимы, например, ко всему пчелиному улью, если только некоторые укладчики производят потомство, которое все будут самцами от неоплодотворенных яиц: в этом случае среднее родство будет ниже, чем показано.
Общие пропорции генов в гапло-диплоидной системе определения пола Секс женский Мужской Дочь 1⁄2 1 Сын 1⁄2 Нет данных Мать 1⁄2 1 Отец 1⁄2 Нет данных Сестра 3⁄4 1⁄2 Брат 1⁄4 1⁄2 Тетя по материнской линии 3⁄8 3⁄4 Дядя по матери 1⁄8 1⁄4 По отцовской тетя 1⁄4 Нет данных Дядя по отцу 1⁄4 Нет данных Племянница (дочь сестры) 3⁄8 1⁄4 Племянница (дочь брата) 1⁄4 1⁄2 Племянник (сын сестры) 3⁄8 1⁄4
При этом предположении, что матери спариваются только один раз, сестры более связаны друг с другом, чем со своими собственными дочерьми. Этот факт был использован для объяснения эволюция эусоциальности у многих перепончатокрылых. Однако колонии, в которых есть рабочие от нескольких маток или маток, которые спаривались несколько раз, будут иметь отношение рабочих к рабочим, которое меньше, чем отношение рабочих к дочерям, например, в Melipona scutellaris.
Смотрите также
- Хромосома
- Эффект зеленой бороды
- Плоидность
- Псевдо-арренотоки
- Система определения пола
- Половая дифференциация
- Сотрудник полиции
- Х хромосома
- Y-хромосома
Рекомендации
- ^ King, R.C; Стэнсфилд, W.D .; Маллиган, П. (2006). Словарь генетики (7-е изд.). Издательство Оксфордского университета. п.194. ISBN 978-0-19-530761-0.
- ^ Grimaldi, D .; Энгель М.С. (2005). Эволюция насекомых. Издательство Кембриджского университета. п.408. ISBN 978-0-521-82149-0.
- ^ а б Уайт, Майкл Дж. Д. (1984). «Хромосомные механизмы в репродукции животных». Bolletino di Zoologia. 51 (1–2): 1–23. Дои:10.1080/11250008409439455. ISSN 0373-4137.
- ^ Grimaldi, D .; Энгель М.С. (2005). Эволюция насекомых. Издательство Кембриджского университета. п.465. ISBN 978-0-521-82149-0.
- ^ Hughes, W.O.H .; и другие. (2008). «Моногамия предков показывает, что родственный отбор является ключом к эволюции эусоциальности». Наука. 320 (5880): 1213–1216. Дои:10.1126 / science.1156108. PMID 18511689. S2CID 20388889.
- ^ Эдвард О. Уилсон (2005). «Родственный отбор как ключ к альтруизму: его взлеты и падения». Социальные исследования. 72 (1): 159–166. JSTOR 40972006.
- ^ ван Вильгенбург, Эллен; Дриссен, Джерард и Бёкебум, Лео В. Дополнительное определение пола по одному локусу у перепончатокрылых: «неразумный» замысел? Границы зоологии 2006, 3: 1
- ^ Маховальд, Майкл; фон Веттберг, ЭрикОпределение пола у перепончатокрылых Swarthmore College (1999)
- ^ Чоу, А .; Маккауэр, М. (1996). «Последовательное выделение полов потомства у гиперпаразитоидной осы Dendrocerus carpenteri». Поведение животных. 51 (4): 859–870. Дои:10.1006 / anbe.1996.0090. S2CID 53192398.
- ^ Van Alphen, J. J. M .; Тунниссен, И. (1982). «Выбор хозяев и определение пола Pachycrepoideus Vindemiae Rondani (Pteromalidae) как факультативный гиперпаразитоид Asobara Tabida Nees (Braconidae; Alysiinae) и Leptopilina Heterotoma (Cynipoidea; Eucoilidae)». Нидерландский зоологический журнал. 33 (4): 497–514. Дои:10.1163 / 002829683X00228.
- ^ Peer, K .; Таборский, М. (2004). «Самки жуков-амброзий регулируют соотношение полов в потомстве в соответствии с возможностями аутбридинга своих сыновей». Журнал эволюционной биологии. 17 (2): 257–264. Дои:10.1111 / j.1420-9101.2003.00687.x. PMID 15009259.
- ^ Синерво, Барри Родственный отбор и гаплодиплоидия у социальных перепончатокрылых В архиве 2011-07-16 на Wayback Machine 1997
- ^ Woyka, J .; Pszczelnictwa, Zaklad; Личинки трутня из оплодотворенных яиц медоносной пчелы Журнал исследований пчеловодства, (1963), страницы 19-24
- ^ Weinstock, George M .; Робинсон, Джин Э. и Консорциум по секвенированию генома медоносных пчел Понимание социальных насекомых из генома пчелы Apis mellifera Природа, том «443» (2006 г.), страницы 931-949
- ^ Сантомауро, Джулия; Олдхэм, Нил Дж .; Боланд, Вильгельм; Энгельс Вольф; Каннибализм диплоидных трутневых личинок у медоносной пчелы (Apis mellifera) высвобождается из-за необычной структуры круглого вещества Журнал исследований пчеловодства, том «43» (2004 г.), стр. 69-74
- ^ Фостер, Кевин Р .; Ратниекс, Фрэнсис Л. В. (2001). «Влияние предвзятости, связанной с распределением пола, на эволюцию работы полиции в сообществах перепончатокрылых» (PDF). Американский натуралист. 158 (6): 615–623. Дои:10.1086/323588. PMID 18707355.
- ^ Baer, B .; П. Шмид-Хемпель (2001). «Неожиданные последствия полиандрии для паразитизма и приспособленности шмелей, Бомб террестрис". Эволюция. 55 (8): 1639–1643. Дои:10.1554 / 0014-3820 (2001) 055 [1639: ucopfp] 2.0.co; 2. PMID 11580023.
- ^ Дэвис, Николас Б., Джон Р. Кребс и Стюарт А. Вест. (2012). Введение в поведенческую экологию. Вили-Блэквелл. С. 371–375.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Гамильтон, В. Д. (1996). Узкие дороги генной земли: сборник статей У.Д. Гамильтона. Оксфорд, Нью-Йорк: W.H. Freeman / Spektrum. ISBN 978-0-7167-4530-3.
- ^ Кевин Р. Фостер; Том Венселирс; Фрэнсис Л.В. Ратниекс (2006). «Родственный отбор - ключ к альтруизму». Тенденции в экологии и эволюции. 21 (2): 57–60. Дои:10.1016 / j.tree.2005.11.020. PMID 16701471.
Библиография
- Бей, Мартин; и другие. (1999). «Необычно высокая скорость рекомбинации обнаружена в области локуса пола медоносной пчелы (Apis mellifera)". Генетика. 153 (4): 1701–1708. ЧВК 1460844. PMID 10581277.
- Wu, Z .; и другие. (2005). «Дополнительное определение пола по одному локусу отсутствует у Heterospilus prosopidis (Hymenoptera: Braconidae)». Наследственность. 95 (3): 228–234. Дои:10.1038 / sj.hdy.6800720. PMID 16077738.
- Ратниекс, Франциск (1988). «Репродуктивная гармония посредством взаимного контроля со стороны рабочих эусоциальных перепончатокрылых». Американский натуралист. 132 (2): 217–236. Дои:10.1086/284846. JSTOR 2461867.
| Темы |  | |
|---|---|---|
| Группы | ||
| В культуре | ||
| Пионеры, работает |
| |