Инбридинг - Inbreeding
Эта статья поднимает множество проблем. Пожалуйста помоги Улучши это или обсудите эти вопросы на страница обсуждения. (Узнайте, как и когда удалить эти сообщения-шаблоны) (Узнайте, как и когда удалить этот шаблон сообщения)
|

Инбридинг это производство потомство от вязка или разведение особей или организмы которые близко генетически связанные.[2] По аналогии этот термин используется в человеческое воспроизводство, но чаще относится к генетическим нарушениям и другим последствиям, которые могут возникнуть в результате выражения вредный или же рецессивный черты характера кровосмесительный сексуальные отношения и кровное родство.
Инбридинг приводит к гомозиготность, что может увеличить вероятность заражения потомства вредными или рецессивными черты.[3] Обычно это приводит, по крайней мере, к временному снижению биологическая пригодность из численность населения[4][5] (называется инбридинговая депрессия ), то есть его способность выживать и воспроизводиться. Человека, унаследовавшего такие вредные черты, в просторечии называют инбредный. Избегание проявления таких вредных рецессивных аллели вызвано инбридингом, через избегание инбридинга механизмы, является основной селективной причиной ауткроссинга.[6][7] Скрещивание между популяциями также часто оказывает положительное влияние на характеристики, связанные с фитнесом,[8] но также иногда приводит к негативным эффектам, известным как аутбредная депрессия. Однако повышенная гомозиготность увеличивает вероятность фиксации полезных аллелей, а также немного снижает вероятность фиксации вредных аллелей в популяции.[9] Инбридинг может привести к удалению вредных аллелей из популяции через очищающий отбор.[10][11][12]
Инбридинг - это метод, используемый в селекция. Например, в животноводство, заводчики могут использовать инбридинг при попытке создать новый и желательный черта в поголовье и для создания отдельных семейств внутри породы, но необходимо будет следить за нежелательными характеристиками у потомства, которые затем могут быть устранены путем дальнейшего селективного разведения или выбраковка. Инбридинг также помогает установить тип действия гена, влияющий на признак. Инбридинг также используется для выявления вредных рецессивных аллелей, которые затем могут быть устранены путем ассортативное разведение или через выбраковку. В селекция растений, инбредные линии используются как запасы для создания гибридный линий, чтобы использовать эффекты гетерозис. Инбридинг у растений также происходит естественным образом в виде самоопыление.
Инбридинг может существенно повлиять на экспрессия гена которые могут предотвратить инбридинговую депрессию.[13]
Обзор
Потомство биологически связанных лиц подвержено возможным последствиям инбридинга, например: врожденные пороки развития. В шансы таких расстройств увеличиваются, когда биологические родители более тесно связаны. Это потому, что такие пары имеют 25% вероятность получения гомозиготный зиготы, в результате чего потомство с двумя рецессивные аллели, которые могут вызывать расстройства когда эти аллели вредны.[14] Поскольку большинство рецессивных аллелей в популяциях встречается редко, маловероятно, что два неродственных брака будут носителями одного и того же вредного аллеля; однако, поскольку близкие родственники имеют большую часть своих аллелей, вероятность того, что любой такой вредный аллель унаследован от общего предка через обоих родителей, резко возрастает. Для каждой сформированной гомозиготной рецессивной особи существует равный шанс произвести гомозиготную доминантную особь, полностью лишенную вредного аллеля. Вопреки распространенному мнению, инбридинг сам по себе не изменяет частоты аллелей, а скорее увеличивает относительное соотношение гомозигот к гетерозиготам; однако, поскольку повышенная доля вредных гомозигот подвергает аллель воздействию естественный отбор, в долгосрочной перспективе его частота снижается быстрее в инбредных популяциях. В краткосрочной перспективе ожидается, что кровосмесительное размножение увеличит количество самопроизвольных абортов зигот, перинатальных смертей и послеродового потомства с врожденными дефектами.[15] Преимущества инбридинга могут быть результатом тенденции к сохранению структур аллелей, взаимодействующих в разных локусах, которые были адаптированы вместе за счет общей селективной истории.[16]
Пороки развития или вредные черты могут оставаться в популяции из-за высокого уровня гомозиготности, и это приведет к тому, что популяция станет фиксированной по определенным признакам, таким как наличие слишком большого количества костей в определенной области, например, позвоночный столб волков на острове Рояль или черепной мозг. аномалии, такие как у северных морских слонов, у которых изменилась длина их черепных костей в нижнем нижнечелюстном ряду зубов. Высокий уровень гомозиготности проблематичен для популяции, потому что он демаскирует рецессивные вредные аллели, генерируемые мутациями, уменьшит преимущество гетерозигот и пагубно сказывается на выживании небольших популяций животных, находящихся под угрозой исчезновения.[17] Когда вредные рецессивные аллели обнаруживаются из-за повышенной гомозиготности, вызванной инбридингом, это может вызвать инбридинговую депрессию.[18]
Помимо рецессивных заболеваний, могут быть и другие вредные эффекты. Таким образом, аналогичные иммунная система может быть более уязвимым к инфекционным заболеваниям (см. Главный комплекс гистосовместимости и половой отбор ).[19]
Историю инбридинга в популяции также следует учитывать при обсуждении различий в степени тяжести депрессии инбридинга между видами и внутри видов. Имеются данные, свидетельствующие о том, что при стойком инбридинге депрессия инбридинга становится менее серьезной. Это связано с обнаружением и устранением крайне вредных рецессивных аллелей. Однако инбридинговая депрессия - это не временное явление, потому что устранение вредных рецессивных аллелей никогда не будет полным. Устранение слегка вредоносных мутаций посредством инбридинга при умеренном отборе не так эффективно. Фиксация аллелей, скорее всего, происходит через Трещотка Мюллера, когда в геноме бесполой популяции накапливаются необратимые вредные мутации.[20]
Несмотря на все свои недостатки, инбридинг может иметь ряд преимуществ, например, снижение рекомбинационная нагрузка,[21] и позволяя выражать рецессивные благоприятные фенотипы. Было высказано предположение, что при обстоятельствах, когда преимущества инбридинга перевешивают недостатки, можно было бы продвигать предпочтительное разведение внутри малых групп, что потенциально могло бы привести к видообразование.[22]
Генетические нарушения
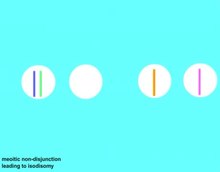
Аутосомно-рецессивный расстройства возникают у людей, которые две копии аллеля для конкретного рецессивного генетического мутация.[23] За исключением некоторых редких обстоятельств, таких как новые мутации или однопородная дисомия, оба родителя человека с таким заболеванием будут носителями гена. Эти носители не проявляют никаких признаков мутации и могут не знать, что они несут мутировавший ген. Поскольку родственники имеют более высокую долю своих генов, чем неродственные люди, более вероятно, что оба родственных родителя будут носителями одного и того же рецессивного аллеля, и, следовательно, их дети подвергаются более высокому риску унаследования аутосомно-рецессивного генетического заболевания. Степень увеличения риска зависит от степени генетического родства между родителями; риск выше, если родители являются близкими родственниками, и ниже для отношений между более дальними родственниками, такими как троюродные братья, хотя все же выше, чем для населения в целом.[24]
Дети союзов родителей и детей или братьев и сестер подвергаются повышенному риску по сравнению с союзами двоюродных братьев и сестер.[25]:3 Инбридинг может привести к большему, чем ожидалось, фенотипическому проявлению вредных рецессивные аллели внутри населения.[26] В результате инбредные особи первого поколения с большей вероятностью будут иметь физические недостатки и дефекты здоровья.[27][28] включая:
- Уменьшенный плодородие как по размеру помета, так и сперма жизнеспособность
- Повысился генетические нарушения
- Колеблющийся асимметрия лица
- Ниже уровень рождаемости
- Выше младенческая смертность и детская смертность[29]
- Меньше взрослый размер
- Утрата иммунная система функция
- Повысился сердечно-сосудистые риски[30]
Изоляция небольшой популяции на период времени может привести к инбридингу внутри этой популяции, что приведет к увеличению генетического родства между размножающимися особями. Инбридинговая депрессия также может возникать в большой популяции, если люди склонны спариваться со своими родственниками, а не случайным образом.
Многие особи первого поколения инбридинга никогда не доживут до воспроизводства.[31] Со временем с изоляцией, такой как узкое место населения вызвано целенаправленным (ассортативный ) племенной или натуральный относящийся к окружающей среде факторов, вредные унаследованные черты отбрасываются.[6][7][32]
Островные виды часто очень инбредны, так как их изоляция от более крупной группы на материке позволяет естественному отбору воздействовать на их популяцию. Этот тип изоляции может привести к образованию раса или даже видообразование, поскольку инбридинг сначала удаляет многие вредоносные гены и позволяет экспрессировать гены, которые позволяют популяции адаптироваться к экосистема. По мере того, как адаптация становится более выраженной, новый вид или раса излучается от входа в новое пространство или вымирает, если не может адаптироваться и, что наиболее важно, воспроизводиться.[33]
Сокращенный генетическое разнообразие, например, из-за узкого места неизбежно приведет к увеличению инбридинга для всей популяции. Это может означать, что разновидность может быть не в состоянии адаптироваться к изменениям условий окружающей среды. У каждого человека будет схожая иммунная система, поскольку иммунная система основана на генетике. Когда вид становится находящихся под угрозой исчезновения, популяция может упасть ниже минимума, в результате чего принудительное скрещивание оставшихся животных приведет к вымирание.
Естественное разведение включает инбридинг по необходимости, и большинство животных мигрируют только при необходимости. Во многих случаях ближайший доступный помощник - это мать, сестра, бабушка, отец, брат или дедушка. Во всех случаях окружающая среда создает стрессовые ситуации для удаления из населения тех людей, которые не могут выжить из-за болезни.[нужна цитата ]
Было предположение[кем? ] что дикие популяции не подвергаются инбридингу; это не то, что в некоторых случаях наблюдается в дикой природе. Однако у таких видов, как лошади, животные в дикий или же дикий условия часто отгоняют детенышей обоих полов, что считается механизмом, с помощью которого вид инстинктивно избегает некоторых генетических последствий инбридинга.[34] В целом, многие виды млекопитающих, в том числе самые близкие к человечеству примат родственники, избегайте близкого инбридинга, возможно, из-за вредных последствий.[25]:6
Примеры
Хотя существует несколько примеров инбредных популяций диких животных, негативные последствия этого инбридинга плохо документированы.[нужна цитата ] в Южноамериканский морской лев высказывались опасения, что недавние популяционные катастрофы уменьшат генетическое разнообразие. Исторический анализ показал, что рост популяции всего по двум матрилинейным линиям был ответственен за большинство людей в популяции. Тем не менее, разнообразие линий позволило значительно разнообразить генофонд, что может помочь защитить южноамериканского морского льва от исчезновения.[35]

Во львах, гордость часто сопровождаются родственными мужчинами в холостяцких группах. Когда один из этих холостяков убивает или изгоняет доминирующего самца, отца может заменить его сын. Не существует механизма предотвращения инбридинга или обеспечения ауткроссинга. В прайдах большинство львиц связаны друг с другом. Если доминирует более одного самца, группа альфа-самцы обычно связаны. Затем проводится «разведение линий» по двум линиям. Кроме того, в некоторых популяциях, таких как Кратер львов, известно, что произошло узкое место популяции. Исследователи обнаружили гораздо большую генетическую гетерозиготность чем ожидалось.[36] Фактически, хищники известны своей низкой генетической изменчивостью, наряду с большей частью верхней части трофических уровней животного. экосистема.[37] Кроме того, альфа-самцы двух соседних прайдов могут быть из одного помета; один брат может прийти к власти над гордостью другого и впоследствии вступить в брак со своими «племянницами» или кузенами. Однако убийство детенышей другого самца при захвате позволяет новому выбранному генному комплексу приходящего альфа-самца преобладать над предыдущим самцом. Есть генетические анализы запланированы львы для определения их генетического разнообразия. Предварительные исследования показывают результаты, несовместимые с парадигмой ауткроссинга, основанной на индивидуальной среде изучаемых групп.[36]
В Центральной Калифорнии, морские выдры считалось, что они вымерли из-за чрезмерной охоты, пока в 1930-х годах в районе Пойнт-Сур не была обнаружена небольшая колония.[38] С тех пор популяция выросла и распространилась вдоль центрального побережья Калифорнии до примерно 2000 особей, и этот уровень оставался стабильным более десяти лет. Рост популяции ограничен тем фактом, что все калифорнийские каланы произошли от изолированной колонии, что привело к инбридингу.[39]
Гепарды - еще один пример инбридинга. Тысячи лет назад гепард пережил узкое место в популяции, которое резко сократило его популяцию, поэтому все животные, живущие сегодня, связаны друг с другом. Следствием инбридинга для этого вида стала высокая смертность молоди, низкая плодовитость и низкий успех размножения.[40]
В исследовании островной популяции певчих воробьев инбредные особи показали значительно более низкую выживаемость, чем аутбредные особи во время резкого падения популяции, связанного с суровой зимней погодой. Эти исследования показывают, что инбридинговая депрессия и экологические факторы влияют на выживаемость.[20]
Меры
Мерой инбридинга особи A является вероятность F(A) что оба аллеля в одном локусе происходят от одного и того же аллеля предка. Эти два идентичных аллеля, оба происходят от общего предка, называются идентичны по происхождению. Эта вероятность F (A) называется "коэффициент инбридинга ".[41]
Еще одна полезная мера, описывающая степень родства двух индивидов (скажем, индивидов A и B), - их коэффициент родства f (A, B), который дает вероятность того, что один случайно выбранный аллель из A и другой случайно выбранный аллель из B являются идентичны по происхождению.[42] Это также обозначается как коэффициент родства между A и B.[43]
Частным случаем является самосогласование индивидуума A с самим собой, f (A, A), что представляет собой вероятность того, что взяв один случайный аллель из A, а затем, независимо и с заменой, другой случайный аллель также из A, оба идентичны по происхождению. Поскольку они могут быть идентичны по происхождению путем выборки одного и того же аллеля или выборки обоих аллелей, которые оказались идентичными по наследству, мы имеем f (A, A) = 1/2 + F (A) / 2.[44]
Коэффициенты инбридинга и coancestry могут быть определены для конкретных особей или как средние значения популяции. Их можно вычислить по генеалогии или оценить по размеру популяции и ее племенным свойствам, но все методы не предполагают отбора и ограничиваются нейтральными аллелями.
Есть несколько методов вычисления этого процента. Двумя основными способами являются метод пути[45][41] и табличный метод.[46][47]
Типичные половые отношения между родственниками следующие:
- Отец / дочь или мать / сын → 25% (1⁄4)
- Брат / сестра → 25% (1⁄4)
- Дед / внучка или бабушка / внук → 12,5% (1⁄8)
- Сводный брат / сводная сестра, Двойные кузены → 12,5% (1⁄8)
- Дядя / племянница или тетя / племянник → 12,5% (1⁄8)
- Прадед / правнучка или прабабушка / правнук → 6,25% (1⁄16)
- Полудядя / племянница или сводная тетя / племянник → 6,25% (1⁄16)
- Двоюродные братья → 6,25% (1⁄16)
Животные
Дикие животные

- Полосатый мангуст самки регулярно спариваются со своими отцами и братьями.[48]
- Постельные клопы: Университет штата Северная Каролина обнаружили, что клопы, в отличие от большинства других насекомых, хорошо переносят инцест и генетически способны противостоять последствиям инбридинга.[49]
- Обыкновенная плодовая муха самки предпочитают спариваться со своими братьями, нежели с неродственными самцами.[1]
- Весы с хлопковой подушкой «Оказывается, самки этих насекомых-гермафродитов на самом деле не оплодотворяют свои яйца сами, а вместо этого получают это через паразитическую ткань, которая заражает их при рождении», - говорит Лаура Росс из Зоологический факультет Оксфордского университета. «Похоже, что эта инфекционная ткань происходит из оставшейся спермы их отца, который нашел хитрый способ иметь больше детей, спариваясь со своими дочерьми».[50]
- Адактилидиум: Одинокий потомок мужского пола спаривается со всеми дочерьми, когда они все еще находятся в матери. Оплодотворенные самки вырезают отверстия в теле своей матери, чтобы они могли выйти и найти новые яйца трипса. Самец тоже появляется, но не ищет пищи или новых партнеров и умирает через несколько часов. Самки погибают в возрасте 4 дней, когда собственное потомство съесть их живьем изнутри.[51]
Полудомашние животные

Домашние животные

Разведение домашних животных - это прежде всего ассортативный разведение (см. селекция ). Без сортировки особей по признакам невозможно установить породу и удалить плохой генетический материал.Гомозиготность - это случай, когда сходные или идентичные аллели объединяются, чтобы выразить признак, который не выражается иначе (рецессивность). Инбридинг выявляет рецессивные аллели за счет увеличения гомозиготности.[55]
Заводчики должны избегать разведения от особей, которые демонстрируют либо гомозиготность, либо гетерозиготность по аллелям, вызывающим заболевание.[56] Цель предотвращения передачи вредных аллелей может быть достигнута репродуктивной изоляцией. стерилизация, или, в крайнем случае, выбраковка. Выбраковка не является строго необходимой, если единственная проблема - генетика. Мелких животных, таких как кошки и собаки, можно стерилизовать, но в случае крупных сельскохозяйственных животных, таких как крупный рогатый скот, выбраковка обычно является единственным экономичным вариантом.
Вопрос о случайных заводчиках, которые безответственно скрещивают инбрид, обсуждается в следующей цитате по крупному рогатому скоту:
Между тем, производство молока на корову за лактацию увеличилось с 17 444 фунтов до 25 013 фунтов с 1978 по 1998 год для голштинской породы. Средние племенные показатели молока коров голштинской породы увеличились за этот период на 4829 фунтов.[57] Высокопродуктивных коров становится все труднее разводить, и они требуют более высоких затрат на здоровье, чем коровы с более низкой генетической ценностью для производства (Cassell, 2001).
Интенсивный отбор с целью повышения урожайности увеличил взаимоотношения между животными внутри породы и увеличил частоту случайного инбридинга.
Многие признаки, влияющие на прибыльность при скрещивании современных молочных пород, не изучались в запланированных экспериментах. Действительно, все исследования скрещивания с участием североамериканских пород и линий очень устарели (McAllister, 2001), если они вообще существуют.[58]
BBC выпустила два документальных фильма об инбридинге собак под названием Разоблачены породистые собаки и Разоблачение породистых собак: три года спустя документируют негативные последствия для здоровья чрезмерного инбридинга.
Линнбридинг
Линнбридинг - это форма инбридинга. Между этими двумя терминами нет четкого различия, но лайнбридинг может включать в себя скрещивание особей и их потомков или двух кузенов.[54][59] Этот метод можно использовать для увеличения вклада конкретного животного в популяцию.[54] Хотя лайнбридинг с меньшей вероятностью вызовет проблемы в первом поколении, чем инбридинг, со временем лайнбридинг может уменьшить генетическое разнообразие популяции и вызвать проблемы, связанные со слишком маленьким генофондом, который может включать повышенную распространенность генетических нарушений и инбридинга. депрессия.[нужна цитата ]
Ауткроссинг
Ауткроссинг - это скрещивание двух неродственных особей для получения потомства. При ауткроссинге, если нет поддающейся проверке генетической информации, можно обнаружить, что все особи отдаленно связаны с древним прародителем. Если эта черта распространяется на всю популяцию, все люди могут иметь эту черту. Это называется эффект основателя. У широко известных пород, которые обычно разводятся, присутствует большой генофонд. Например, в 2004 году было зарегистрировано более 18 000 персидских кошек.[60] Существует возможность полного ауткросса, если между особями не существует препятствий для размножения. Однако это не всегда так, и имеет место разновидность удаленного лайнбридинга. И снова заводчик-ассортативный заводчик должен знать, какие черты, как положительные, так и отрицательные, существуют в разнообразии одного разведения. Это разнообразие генетической экспрессии даже у близких родственников увеличивает изменчивость и разнообразие жизнеспособного стада.
Лабораторные животные
Систематический инбридинг и поддержание инбредных линий лабораторные мыши и крысы имеют большое значение для биомедицинских исследований. Инбридинг гарантирует последовательное и однородное модель животных для экспериментальных целей и позволяет проводить генетические исследования в родственный и нокаутирующие животные. Для того, чтобы получить линию мышей, которая считается инбредной, должно произойти минимум 20 последовательных поколений вязок сиблингов. С каждым последующим поколением разведения гомозиготность во всем геноме увеличивается, устраняя гетерозиготные локусы. При 20 поколениях скрещиваний братьев и сестер гомозиготность наблюдается примерно в 98,7% всех локусов в геноме, что позволяет этим потомкам служить моделями на животных для генетических исследований.[61] Использование инбредных штаммов также важно для генетических исследований на животных моделях, например, чтобы отличить генетические эффекты от воздействия окружающей среды. Инбредные мыши обычно показывают значительно более низкие показатели выживаемости.
Люди
Последствия
Инбридинг увеличивает шансы на экспрессию вредных рецессивных аллелей за счет увеличения гомозиготности и, следовательно, может снизить приспособленность потомства. При непрерывном инбридинге генетическая изменчивость теряется, а гомозиготность увеличивается, что делает возможным экспрессию рецессивных вредных аллелей у гомозигот. В коэффициент инбридинга, или степень инбридинга у индивидуума, является оценкой процента гомозиготных аллелей в общем геноме.[63] Чем более биологически родственны родители, тем выше коэффициент инбридинга, поскольку их геномы уже имеют много общего. Эта общая гомозиготность становится проблемой, когда в генофонде семьи присутствуют вредные рецессивные аллели.[64] Путем спаривания хромосом схожих геномов вероятность спаривания этих рецессивных аллелей и их гомозиготности значительно увеличивается, что приводит к появлению у потомства аутосомно-рецессивных расстройств.[64]
Инбридинг особенно проблематичен в небольших популяциях, где генетическая изменчивость уже ограничена.[65] Путем инбридинга индивиды еще больше уменьшают генетическую изменчивость за счет увеличения гомозиготности в геномах своего потомства.[66] Таким образом, вероятность спаривания вредных рецессивных аллелей значительно выше в небольшой популяции инбридинга, чем в большой популяции инбридинга.[65]
Последствия для приспособленности кровного спаривания изучаются с момента их научного признания Чарльз Дарвин в 1839 г.[67][68] Некоторые из наиболее вредных последствий такого разведения включают в себя его влияние на уровень смертности, а также на общее состояние здоровья потомства.[69] С 1960-х годов было проведено множество исследований, подтверждающих такое изнурительное воздействие на человеческий организм.[66][67][69][70][71] В частности, было обнаружено, что инбридинг снижает фертильность как прямой результат увеличения гомозиготности вредных рецессивных аллелей.[71][72] Плоды, полученные в результате инбридинга, также подвергаются большему риску самопроизвольных абортов из-за присущих им осложнений в развитии.[73] Среди матерей, мертворожденных и умерших в раннем детстве, инбридинговые матери имеют значительно более высокие шансы на получение повторных результатов с будущим потомством.[74] Кроме того, кровные родители имеют высокий риск преждевременных родов и рождения детей с недостаточным весом и ростом.[75] Жизнеспособное инбредное потомство также может иметь физические уродства и генетически унаследованные заболевания.[63] Исследования подтвердили рост ряда генетических нарушений из-за инбридинга, таких как слепота, потеря слуха, неонатальный диабет, пороки развития конечностей, нарушения полового развития, шизофрения и ряд других.[63][76] Более того, существует повышенный риск врожденных пороков сердца в зависимости от коэффициента инбридинга (см. коэффициент инбридинга ) потомства со значительным риском, сопровождающимся F = 0,125 или выше.[27]
Распространенность
Общий негативный настрой и отказ от инбридинга, который преобладает в западный мир сегодня имеет корни более 2000 лет назад. В частности, письменные документы, такие как Библия, показывают, что существовали законы и общественные обычаи, которые призывали воздерживаться от инбридинга. Наряду с культурными табу, образование родителей и осознание последствий инбридинга сыграли большую роль в минимизации частоты инбридинга в таких регионах, как Европа. Таким образом, в мире есть менее урбанизированные и менее населенные регионы, которые продемонстрировали преемственность в практике инбридинга.
Непрерывность инбридинга часто происходит либо по выбору, либо неизбежно из-за ограничений географического района. По выбору, уровень кровного родства сильно зависит от религии и культуры.[65] В западном мире некоторые Анабаптист группы сильно инбредны, потому что они происходят от небольших популяций основателей и до тех пор, пока[требуется разъяснение ] сегодня[когда? ] брак вне группы не допускается для членов.[нужна цитата ] Особенно Меннониты старого порядка Рейденбаха[77] и Гуттериты происходят от очень небольшой популяции основателей. То же верно и для некоторых Хасидский и Евреи харедим группы.
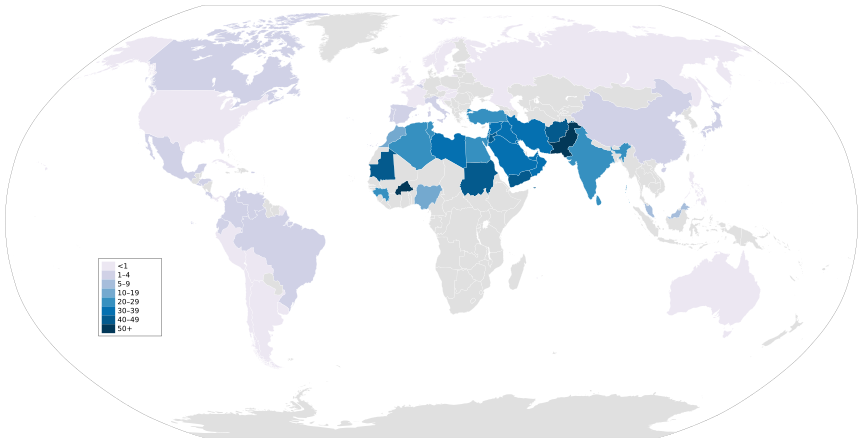
Из практикующих регионов территории Ближнего Востока и Северной Африки демонстрируют наибольшую частоту кровного родства. [65] Связь между высокой частотой и регионом в первую очередь связана с доминированием исламского населения, которое исторически было вовлечено в семейные отношения.[68]
Среди этих популяций с высоким уровнем инбридинга исследователи обнаружили несколько расстройств, распространенных среди инбредных потомков. В Ливан, Саудовская Аравия, Египет, И в Израиль, у потомков кровных родственников повышен риск врожденных пороков развития, врожденных пороков сердца, врожденных гидроцефалия и дефекты нервной трубки.[65] Кроме того, среди инбредных детей в Палестина и Ливана существует положительная связь между кровным родством и заячья губа / нёбо случаи.[65] Исторически сложилось так, что популяции Катар были вовлечены в кровные отношения всех видов, что привело к высокому риску унаследовать генетические заболевания. По состоянию на 2014 год около 5% населения Катара страдали от наследственной потери слуха; большинство из них были потомками кровного родства.[78]
Роялти и дворянство


Меж дворянство брак использовался как метод формирования политические союзы среди элит[нужна цитата ]. Эти связи часто скреплялись только после рождения потомства внутри брак по договоренности. Таким образом, брак рассматривался как союз знатных родов, а не как договор между людьми, как это представляется.[кем? ] сегодня.
Королевский смешанный брак часто практиковалось среди европейских королевских семей, обычно в государственных интересах. Со временем из-за относительно ограниченного числа потенциальных супругов Генофонд многих правящих семей становилось все меньше, пока все европейские королевские семьи не стали родственными. Это также привело к тому, что многие из них произошли от определенного человека по разным линиям происхождения, например, многочисленные европейские члены королевской семьи и дворянство произошли от британских Королева Виктория или же Король Дании Кристиан IX.[79] Дом Габсбург был печально известен своим инбридингом, с Губа Габсбургов цитируется как вредный эффект, хотя никакие генетические доказательства не подтверждают это утверждение. Близкородственные дома Габсбургов, Бурбон, Браганса и Виттельсбах также часто состоит в браках с двоюродными братьями и сестрами, а также иногда двоюродный брат и браки дяди и племянницы. Примеры кровосмесительных браков и влияние инбридинга на королевские семьи включают:
- В древний Египет считалось, что королевские женщины носили родословную, и это было выгодно для фараон жениться на сестре или сводной сестре;[80] в таких случаях особая комбинация между эндогамия и многоженство находится. Обычно старшие сын и дочь старого правителя (которые могли быть либо братьями и сестрами, либо сводными братьями и сестрами) становились новыми правителями. Все правители Династия птолемеев непрерывно из Птолемей IV (Птолемей II женился на своей сестре, но не имел проблем) были женаты на своих братьях и сестрах, чтобы сохранить «чистую» кровь Птолемеев и укрепить линию преемственности. король Тутанхамон мать была сводной сестрой его отцу,[81] Клеопатра VII (также называемый Клеопатрой VI) и Птолемей XIII, которые женились и стали соправителями древний Египет после смерти их отца, являются наиболее широко известным примером.[82]
- В Европе Дом Габсбургов классно обладал нижнечелюстной прогнатизм из Habsburger (Unter) Lippe (иначе известная как «Габсбургская челюсть», «Габсбургская губа» или «Австрийская губа»). Это было типично для многих родственников Габсбургов на протяжении шести веков.[83] Из-за множества кровнородственный браки в семье, состояние прогрессировало из поколения в поколение до последних испанских Габсбургов, Карл II Испании (1661-1700), не мог нормально пережевывать пищу.[84] Карл II имел большое количество генетических, физических, интеллектуальных, сексуальных и эмоциональных проблем. Его коэффициент инбридинга составлял 0,254, что соответствует потомству брата-сестры. инцест.[85] Его импотенция / бесплодие привело к исчезновению (мужчин) испанских Габсбургов после его смерти, что вызвало Война за испанское наследство, что привело к правлению Испании французскими Дом Бурбонов.
Смотрите также
- Дело Альвареса
- Коэффициент родства
- Кровное родство
- Кузен брак
- Кузен брак на Ближнем Востоке
- Эволюция полового размножения
- Экзогамии
- Эффект основателя
- F-статистика
- Футляр Fritzl
- Генетическое разнообразие
- Генетическая чистка
- Генетическое сексуальное влечение
- Преимущество гетерозиготы
- Точка идентичных предков
- Инбридинговая депрессия
- Инцест
- Табу на инцест
- Островная карликовость
- Интеллектуальный инбридинг
- Законность инцеста
- Список парных кузенов
- Махрам
- Аутбридинговая депрессия
- Ауткроссинг
- Близкое родство
- Запрещенная степень родства
- Селекция
- Самонесовместимость растений (как некоторые растения избегают инбридинга)
Рекомендации
- ^ а б Лояу А., Корнуо Дж. Х., Клобер Дж., Данчин Э (2012). «Кровосмесительные сестры: предпочтение партнера братьям неродственным самцам у Drosophila melanogaster». PLOS ONE. 7 (12): e51293. Дои:10.1371 / journal.pone.0051293. ЧВК 3519633. PMID 23251487.
- ^ Инбридинг на Британская энциклопедия
- ^ Набулси М.М., Тамим Х., Саббаг М., Обейд М.Ю., Юнис К.А., Битар Ф.Ф. (февраль 2003 г.). «Родительское кровное родство и врожденные пороки сердца в развивающейся стране». Американский журнал медицинской генетики. Часть А. 116A (4): 342–7. Дои:10.1002 / ajmg.a.10020. PMID 12522788. S2CID 44576506.
- ^ Хименес Дж. А., Хьюз К. А., Алакс Дж., Грэм Л., Лейси Р. К. (октябрь 1994 г.). «Экспериментальное исследование инбредной депрессии в естественной среде обитания». Наука. 266 (5183): 271–3. Дои:10.1126 / science.7939661. PMID 7939661.
- ^ Чен X (1993). «Сравнение инбридинга и аутбридинга у гермафродитов. Арианта арбусторум (L.) (наземная улитка) ". Наследственность. 71 (5): 456–461. Дои:10.1038 / хди.1993.163.
- ^ а б Бернштейн Х., Байерли Х.С., Хопф Ф.А., Мичод Р.Э. (сентябрь 1985 г.). «Генетические повреждения, мутации и эволюция пола». Наука. 229 (4719): 1277–81. Дои:10.1126 / science.3898363. PMID 3898363.
- ^ а б Michod RE. Эрос и эволюция: естественная философия секса. (1994) Книги Персея, ISBN 0-201-40754-X
- ^ Линч М. (1991). "Генетическая интерпретация депрессии инбридинга и депрессии аутбридинга". Эволюция; Международный журнал органической эволюции. Орегон: Общество изучения эволюции. 45 (3): 622–629. Дои:10.1111 / j.1558-5646.1991.tb04333.x. PMID 28568822. S2CID 881556.[страница нужна ]
- ^ Whitlock MC (июнь 2003 г.). «Вероятность и время фиксации в разделенных популяциях». Генетика. 164 (2): 767–79. ЧВК 1462574. PMID 12807795.
- ^ Тьен Н.С., Сабелис М.В., Эгас М. (март 2015 г.). «Инбридинговая депрессия и очищение гаплодиплоида: эффекты, связанные с полом». Наследственность. 114 (3): 327–32. Дои:10.1038 / hdy.2014.106. ЧВК 4815584. PMID 25407077.
- ^ Пер К, Таборский М (февраль 2005 г.). «Депрессия аутбридинга, но не депрессия инбридинга у гаплодиплоидных жуков Ambrosia с регулярным спариванием братьев и сестер». Эволюция; Международный журнал органической эволюции. 59 (2): 317–23. Дои:10.1554/04-128. PMID 15807418. S2CID 198156378.
- ^ Гулисия Д., Ворона Дж. Ф. (май 2007 г.). «Вывод чистки по племенным данным». Эволюция; Международный журнал органической эволюции. 61 (5): 1043–51. Дои:10.1111 / j.1558-5646.2007.00088.x. PMID 17492959. S2CID 24302475.
- ^ Гарсия К., Авила В., Кесада Н., Кабальеро А. (2012). «Изменения экспрессии генов, вызванные инбридингом, защищают от депрессии инбридинга у дрозофилы». Генетика. 192 (1): 161–72. Дои:10.1534 / генетика.112.142687. ЧВК 3430533. PMID 22714404.
- ^ Ливингстон Ф. Б. (1969). «Генетика, экология и истоки инцеста и экзогамии». Современная антропология. 10: 45–62. Дои:10.1086/201009. S2CID 84009643.
- ^ Торнхилл Северо-Запад (1993). Естественная история инбридинга и аутбридинга: теоретические и эмпирические перспективы. Чикаго: Издательство Чикагского университета. ISBN 978-0-226-79854-7.
- ^ Шилдс, В. М. 1982. Филопатрия, инбридинг и эволюция пола. Распечатать. 50–69.
- ^ Meagher S, Penn DJ, Potts WK (март 2000 г.). «Соревнование между самцами усугубляет инбридинговую депрессию у диких домашних мышей». Труды Национальной академии наук Соединенных Штатов Америки. 97 (7): 3324–9. Дои:10.1073 / пнас.97.7.3324. ЧВК 16238. PMID 10716731.
- ^ Swindell WR, et al. (2006). «Депрессия селекции и инбридинга: влияние скорости инбридинга и среды инбридинга». Эволюция. 60 (5): 1014–1022. Дои:10.1554/05-493.1. PMID 16817541. S2CID 198156086.
- ^ Либерман Д., Туби Дж., Космидес Л. (апрель 2003 г.). «Имеет ли мораль биологическую основу? Эмпирический тест факторов, определяющих моральные настроения, связанные с инцестом».. Ход работы. Биологические науки. 270 (1517): 819–26. Дои:10.1098 / rspb.2002.2290. ЧВК 1691313. PMID 12737660.
- ^ а б Пусей А., Вольф М. (май 1996 г.). «Предотвращение инбридинга у животных». Тенденции в экологии и эволюции. 11 (5): 201–6. Дои:10.1016/0169-5347(96)10028-8. PMID 21237809.
- ^ Шилдс В.М. (1982). Филопатрия, инбридинг и эволюция пола. Олбани: Государственный университет Нью-Йорка. ISBN 978-0-87395-618-5.
- ^ Joly E (December 2011). "The existence of species rests on a metastable equilibrium between inbreeding and outbreeding. An essay on the close relationship between speciation, inbreeding and recessive mutations". Биология Директ. 6: 62. Дои:10.1186/1745-6150-6-62. ЧВК 3275546. PMID 22152499.
- ^ Hartl, D.L., Jones, E.W. (2000) Genetics: Analysis of Genes and Genomes. Пятое издание. Jones and Bartlett Publishers Inc., pp. 105–106. ISBN 0-7637-1511-5.
- ^ Kingston HM (April 1989). "ABC of clinical genetics. Genetics of common disorders". BMJ. 298 (6678): 949–52. Дои:10.1136/bmj.298.6678.949. ЧВК 1836181. PMID 2497870.
- ^ а б Wolf AP, Durham WH, eds. (2005). Inbreeding, incest, and the incest taboo: the state of knowledge at the turn. Stanford University Press. ISBN 978-0-8047-5141-4.
- ^ Griffiths AJ, Miller JH, Suzuki DT, Lewontin RC, Gelbart WM (1999). An introduction to genetic analysis. Нью-Йорк: У. Х. Фриман. С. 726–727. ISBN 978-0-7167-3771-1.
- ^ а б Bittles AH, Black ML (January 2010). "Evolution in health and medicine Sackler colloquium: Consanguinity, human evolution, and complex diseases". Труды Национальной академии наук Соединенных Штатов Америки. 107 Suppl 1 (suppl 1): 1779–86. Дои:10.1073/pnas.0906079106. ЧВК 2868287. PMID 19805052.
- ^ Fareed M, Afzal M (2014). "Evidence of inbreeding depression on height, weight, and body mass index: a population-based child cohort study". Американский журнал биологии человека. 26 (6): 784–95. Дои:10.1002/ajhb.22599. PMID 25130378. S2CID 6086127.
- ^ Fareed M, Kaisar Ahmad M, Azeem Anwar M, Afzal M (January 2017). "Impact of consanguineous marriages and degrees of inbreeding on fertility, child mortality, secondary sex ratio, selection intensity, and genetic load: a cross-sectional study from Northern India". Педиатрические исследования. 81 (1): 18–26. Дои:10.1038/pr.2016.177. PMID 27632780.
- ^ Fareed M, Afzal M (April 2016). "Increased cardiovascular risks associated with familial inbreeding: a population-based study of adolescent cohort". Анналы эпидемиологии. 26 (4): 283–92. Дои:10.1016/j.annepidem.2016.03.001. PMID 27084548.
- ^ Bittles AH, Grant JC, Shami SA (June 1993). "Consanguinity as a determinant of reproductive behaviour and mortality in Pakistan". Международный журнал эпидемиологии (Представлена рукопись). 22 (3): 463–7. Дои:10.1093/ije/22.3.463. PMID 8359962.
- ^ Kirkpatrick M, Jarne P (February 2000). "The Effects of a Bottleneck on Inbreeding Depression and the Genetic Load". Американский натуралист. 155 (2): 154–167. Дои:10.1086/303312. PMID 10686158. S2CID 4375158.
- ^ Leck CF (1980). "Establishment of New Population Centers with Changes in Migration Patterns" (PDF). Журнал полевой орнитологии. 51 (2): 168–173. JSTOR 4512538.
- ^ "ADVS 3910 Wild Horses Behavior ", College of Agriculture, Utah State University.
- ^ Freilich S, Hoelzel AR, Choudhury SR. "Genetic diversity and population genetic structure in the South American sea lion (Otaria flavescens)" (PDF). Department of Anthropology and School of Biological & Biomedical Sciences, University of Durham, U.K.
- ^ а б Gilbert DA, Packer C, Pusey AE, Stephens JC, O'Brien SJ (1991-10-01). "Analytical DNA fingerprinting in lions: parentage, genetic diversity, and kinship". Журнал наследственности. 82 (5): 378–86. Дои:10.1093/oxfordjournals.jhered.a111107. PMID 1940281.
- ^ Ramel, C (1998). "Biodiversity and intraspecific genetic variation". Чистая и прикладная химия. 70 (11): 2079–2084. CiteSeerX 10.1.1.484.8521. Дои:10.1351/pac199870112079. S2CID 27867275.
- ^ Kenyon KW (August 1969). "The sea otter in the eastern Pacific Ocean". Фауна Северной Америки. 68: 1–352. Дои:10.3996/nafa.68.0001.
- ^ Bodkin JL, Ballachey BE, Cronin MA, Scribner KT (December 1999). "Population Demographics and Genetic Diversity in Remnant and Translocated Populations of Sea Otters". Биология сохранения. 13 (6): 1378–85. Дои:10.1046/j.1523-1739.1999.98124.x.
- ^ Wielebnowski, Nadja (1996). "Reassessing the relationship between juvenile mortality and genetic monomorphism in captive cheetahs". Зоопарк биологии. 15 (4): 353–369. Дои:10.1002/(SICI)1098-2361(1996)15:4<353::AID-ZOO1>3.0.CO;2-A.
- ^ а б Wright S (1922). "Coefficients of inbreeding and relationship". Американский натуралист. 56 (645): 330–338. Дои:10.1086/279872. S2CID 83865141.
- ^ Reynolds J, Weir BS, Cockerham CC (November 1983). "Estimation of the coancestry coefficient: basis for a short-term genetic distance". Генетика. 105 (3): 767–79. ЧВК 1202185. PMID 17246175.
- ^ Casas AM, Igartua E, Valles MP, Molina-Cano JL (November 1998). "Genetic diversity of barley cultivars grown in Spain, estimated by RFLP, similarity and coancestry coefficients". Селекция растений. 117 (5): 429–35. Дои:10.1111/j.1439-0523.1998.tb01968.x. HDL:10261/121301.
- ^ Malecot G. Les Mathématiques de l'hérédité. Paris: Masson et Cie. p. 1048.
- ^ How to compute and inbreeding coefficient (the path method), Braque du Bourbonnais.
- ^ Christensen K. "4.5 Calculation of inbreeding and relationship, the tabular method". Genetic calculation applets and other programs. Genetics pages.
- ^ García-Cortés LA, Martínez-Ávila JC, Toro MA (2010-05-16). "Fine decomposition of the inbreeding and the coancestry coefficients by using the tabular method". Conservation Genetics. 11 (5): 1945–52. Дои:10.1007/s10592-010-0084-x. S2CID 2636127.
- ^ а б Nichols HJ, Cant MA, Hoffman JI, Sanderson JL (December 2014). "Evidence for frequent incest in a cooperatively breeding mammal". Письма о биологии. 10 (12): 20140898. Дои:10.1098/rsbl.2014.0898. ЧВК 4298196. PMID 25540153.
- ^ "Insect Incest Produces Healthy Offspring". 8 декабря 2011 г.
- ^ Gardner A, Ross L (August 2011). "The evolution of hermaphroditism by an infectious male-derived cell lineage: an inclusive-fitness analysis" (PDF). Американский натуралист. 178 (2): 191–201. Дои:10.1086/660823. HDL:10023/5096. PMID 21750383. S2CID 15361433. Сложить резюме – Живая наука (July 28, 2011).
- ^ Freeman S, Herran JC (2007). "Aging and other life history characters". Evolutionary Analysis (4-е изд.). Pearson Education, Inc. п. 484. ISBN 978-0-13-227584-2.
- ^ "Polycystic kidney disease | International Cat Care". icatcare.org. Получено 2016-07-08.
- ^ "Polycystic Kidney Disease". www.vet.cornell.edu. Получено 2016-07-08.
- ^ а б c Tave D (1999). Inbreeding and brood stock management. Продовольственная и сельскохозяйственная организация Объединенных Наций. п. 50. ISBN 978-92-5-104340-0.
- ^ Bosse, Mirte; Megens, Hendrik‐Jan; Derks, Martijn F. L.; Cara, Ángeles M. R.; Groenen, Martien A. M. (2019). "Deleterious alleles in the context of domestication, inbreeding, and selection". Эволюционные приложения. 12 (1): 6–17. Дои:10.1111/eva.12691. ЧВК 6304688. PMID 30622631.
- ^ G2036 Culling the Commercial Cow Herd: BIF Fact Sheet, MU Extension. Extension.missouri.edu. Retrieved on 2013-03-05.
- ^ "Genetic Evaluation Results". Архивировано из оригинал on August 27, 2001.
- ^ S1008: Genetic Selection and Crossbreeding to Enhance Reproduction and Survival of Dairy Cattle (S-284) В архиве 2006-09-10 на Wayback Machine. Nimss.umd.edu. Retrieved on 2013-03-05.
- ^ Vogt D, Swartz HA, Massey J (October 1993). "Inbreeding: Its Meaning, Uses and Effects on Farm Animals". MU Extension. Университет Миссури. Получено 30 апреля, 2011.
- ^ Top Cat Breeds for 2004. Petplace.com. Retrieved on 2013-03-05.
- ^ Taft, Robert et al. “Know thy mouse.” Science Direct. Vol. 22, No. 12, Dec. 2006, pp. 649-653. Trends in Genetics. https://doi.org/10.1016/j.tig.2006.09.010
- ^ Hamamy H (July 2012). "Consanguineous marriages : Preconception consultation in primary health care settings". Journal of Community Genetics. 3 (3): 185–92. Дои:10.1007/s12687-011-0072-y. ЧВК 3419292. PMID 22109912.
- ^ а б c Woodley, Michael A (2009). "Inbreeding depression and IQ in a study of 72 countries". Интеллект. 37 (3): 268–276. Дои:10.1016/j.intell.2008.10.007.
- ^ а б Kamin, Leon J (1980). "Inbreeding depression and IQ". Психологический бюллетень. 87 (3): 469–478. Дои:10.1037/0033-2909.87.3.469. PMID 7384341.
- ^ а б c d е ж Tadmouri GO, Nair P, Obeid T, Al Ali MT, Al Khaja N, Hamamy HA (October 2009). "Consanguinity and reproductive health among Arabs". Репродуктивное здоровье. 6 (1): 17. Дои:10.1186/1742-4755-6-17. ЧВК 2765422. PMID 19811666.
- ^ а б Roberts DF (November 1967). "Incest, inbreeding and mental abilities". Британский медицинский журнал. 4 (5575): 336–7. Дои:10.1136/bmj.4.5575.336. ЧВК 1748728. PMID 6053617.
- ^ а б Van Den Berghe, Pierre L (2010). "Human inbreeding avoidance: Culture in nature". Поведенческие науки и науки о мозге. 6: 91–102. Дои:10.1017/S0140525X00014850.
- ^ а б Speicher MR, Motulsky AG, Antonarakis SE, Bittles AH, eds. (2010). "Consanguinity, Genetic Drift, and Genetic Diseases in Populations with Reduced Numbers of Founders". Vogel and Motulsky's human genetics problems and approaches (4-е изд.). Берлин: Springer-Verlag. pp. 507–528. ISBN 978-3-540-37654-5.
- ^ а б Ober C, Hyslop T, Hauck WW (January 1999). «Влияние инбридинга на фертильность у людей: доказательства репродуктивной компенсации». Американский журнал генетики человека. 64 (1): 225–31. Дои:10.1086/302198. ЧВК 1377721. PMID 9915962.
- ^ Morton NE (August 1978). "Effect of inbreeding on IQ and mental retardation". Труды Национальной академии наук Соединенных Штатов Америки. 75 (8): 3906–8. Дои:10.1073/pnas.75.8.3906. ЧВК 392897. PMID 279005.
- ^ а б Bittles AH, Grant JC, Sullivan SG, Hussain R (2002-01-01). "Does inbreeding lead to decreased human fertility?". Анналы биологии человека. 29 (2): 111–30. Дои:10.1080/03014460110075657. PMID 11874619. S2CID 31317976.
- ^ Ober C, Elias S, Kostyu DD, Hauck WW (January 1992). "Decreased fecundability in Hutterite couples sharing HLA-DR". Американский журнал генетики человека. 50 (1): 6–14. ЧВК 1682532. PMID 1729895.
- ^ Diamond JM (1987). «Причины смерти до рождения». Природа. 329 (6139): 487–8. Дои:10.1038 / 329487a0. PMID 3657971. S2CID 4338257.
- ^ Stoltenberg C, Magnus P, Skrondal A, Lie RT (April 1999). "Consanguinity and recurrence risk of stillbirth and infant death". Американский журнал общественного здравоохранения. 89 (4): 517–23. Дои:10.2105/ajph.89.4.517. ЧВК 1508879. PMID 10191794.
- ^ Khlat M (December 1989). "Inbreeding effects on fetal growth in Beirut, Lebanon". Американский журнал физической антропологии. 80 (4): 481–4. Дои:10.1002/ajpa.1330800407. PMID 2603950.
- ^ Bener A, Dafeeah EE, Samson N (December 2012). "Does consanguinity increase the risk of schizophrenia? Study based on primary health care centre visits". Психическое здоровье в семейной медицине. 9 (4): 241–8. ЧВК 3721918. PMID 24294299.
- ^ Karsten-Gerhard Albertsen: The History & Life of the Reidenbach Mennonites (Thirty Fivers). Morgantown, Pennsylvania 1996, page 443.
- ^ Girotto G, Mezzavilla M, Abdulhadi K, Vuckovic D, Vozzi D, Khalifa Alkowari M, Gasparini P, Badii R (2014-01-01). "Consanguinity and hereditary hearing loss in Qatar". Человеческая наследственность. 77 (1–4): 175–82. Дои:10.1159/000360475. PMID 25060281.
- ^ Beeche A (2009). The Gotha: Still a Continental Royal Family, Vol. 1. Richmond, US: Kensington House Books. С. 1–13. ISBN 978-0-9771961-7-3.
- ^ Seawright C. "Women in Ancient Egypt, Women and Law". thekeep.org.
- ^ King Tut Mysteries Solved: Was Disabled, Malarial, and Inbred
- ^ Bevan ER. "The House of Ptolomey". uchicago.edu.
- ^ "The Habsburg Lip", Topics in the History of Genetics and Molecular Biology, Fall 2000. Msu.edu. Retrieved on 2013-03-05.
- ^ "The Imperial House of Habsburg: Chapter 5". Архивировано из оригинал 27 августа 2007 г.. Получено 23 сентября, 2007.
- ^ Alvarez G, Ceballos FC, Quinteiro C (2009). "The role of inbreeding in the extinction of a European royal dynasty". PLOS ONE. 4 (4): e5174. Дои:10.1371/journal.pone.0005174. ЧВК 2664480. PMID 19367331.
внешняя ссылка
- Dale Vogt, Helen A. Swartz and John Massey, 1993. Inbreeding: Its Meaning, Uses and Effects on Farm Animals. University of Missouri, Extension.
- Consanguineous marriages with global map
- Ингерсолл Э (1920). . Энциклопедия Американа.
| Ключевые компоненты | |
|---|---|
| Поля | |
| Археогенетика из | |
| похожие темы | |
| Списки | |
| |