Генетическая история Европы - Genetic history of Europe
Эта статья поднимает множество проблем. Пожалуйста помоги Улучши это или обсудите эти вопросы на страница обсуждения. (Узнайте, как и когда удалить эти сообщения-шаблоны) (Узнайте, как и когда удалить этот шаблон сообщения)
|

Южная / Западная Европа Северная / Восточная Европа Кавказ
Ближневосточный Южная Азия Восточная Азия Северная Африка / Африка к югу от Сахары [1]
В генетическая история из Европа так как Верхний палеолит неотделимо от более широкого Западная Евразия Примерно 50-40 000 лет назад (50-40 тыс. Лет назад) возникла базальная западно-евразийская линия (наряду с отдельной Восточная Азия родословная) из недифференцированного "неафриканский «родословная 70-50 тыс. лет назад.[3][4][5][6]И базальные восточные, и западные евразийцы приобрели Неандертальская примесь в Европе и Азии.[7]
Европейские ранние современные люди (EEMH) линии от 40 до 26 тыс. Лет назад (Ориньяк ) все еще были частью большой западной евразийской «метапопуляции», связанной с популяциями Центральной и Западной Азии.[3]Дивергенция в генетически обособленные субпопуляции в пределах Западной Евразии является результатом увеличения давление отбора и эффекты основателя вовремя Последний ледниковый максимум (LGM, Gravettian ).[8]К концу LGM, после 20 тыс. Лет назад, западноевропейское происхождение, получившее название Западноевропейский охотник-собиратель (WHG) выходит из Солютрейский рефугиум вовремя Европейский мезолит.[9] Эти мезолитические культуры охотников-собирателей существенно заменены в Неолитическая революция к прибытию Ранние европейские фермеры (EEF) линии, происходящие от мезолитических популяций Западной Азии (Анатолия и Кавказ ).[10]в Европейский бронзовый век в некоторых частях Европы в результате вторжения Древний Север Евразии (ANE) линии от Причерноморско-каспийские степи. Эти замещения населения бронзового века связаны с Стакан культуры археологически и с Индоевропейская экспансия лингвистически.[11]
В результате перемещений населения в период от мезолита до бронзового века современные европейские популяции отличаются различиями в происхождении WHG, EEF и ANE.[12][13][14]Нормы примеси варьировались географически; в позднем неолите происхождение WHG у фермеров в Венгрии составляло около 10%, в Германии - около 25%, а в Иберии - до 50%.[15] Вклад EEF более значителен в Средиземноморской Европе и снижается в направлении северной и северо-восточной Европы, где происхождение WHG сильнее; то Сардинцы считаются ближайшей европейской группой к населению ВЭФ. Происхождение ANE встречается по всей Европе, максимум около 20% встречается в Прибалтики и Финны.
Этногенез современного этнические группы Европы в исторический период связана с многочисленными примесными событиями, в первую очередь связанными с Римский, Германский, Норвежский, Славянский, Араб и турецкий расширения.
Исследования генетической истории Европы стали возможными во второй половине 20-го века, но не дали результатов с высоким разрешением до 1990-х годов. В 1990-х годах стали возможны предварительные результаты, но они остались в основном ограниченными исследованиями митохондриальный и Y-хромосомный родословные. Аутосомная ДНК стали более доступными в 2000-х годах, а с середины 2010-х годов результаты ранее недостижимого решения, многие из которых основаны на полногеномном анализе древней ДНК, публиковались в ускоренном темпе.[16][17]
Предыстория
В предыстория европейских народов можно проследить, исследуя археологический места, лингвистические исследования и путем изучения ДНК людей, живущих в Европе, или из древней ДНК. Исследования продолжаются, и теории возникают и падают. Хотя можно отслеживать миграции людей по Европе с помощью анализа ДНК основателя, большая часть информации об этих перемещениях поступает из археологии.[18]
Важно отметить, что заселение Европы не происходило отдельными миграциями, как можно было бы предположить. Скорее, процесс заселения был сложным и «вероятно, происходил несколькими волнами с востока и впоследствии был затенен тысячелетиями повторяющегося потока генов».[18]
Из-за естественного отбора процент ДНК неандертальцев у древних европейцев со временем постепенно уменьшался. С 45 000 до 7 000 BP процент упал с 3–6% до 2%.[17] Удаление аллелей, происходящих от неандертальцев, происходило чаще вокруг генов, чем других частей генома.[17]
Палеолит
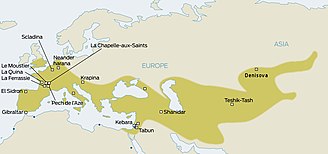
Неандертальцы населяли большую часть Европы и Западной Азии еще 130 000 лет назад. Они существовали в Европе еще 30 000 лет назад. В конечном итоге их заменили анатомически современные люди (AMH; иногда известный как Кроманьонцы ), которые начали появляться в Европе около 40 000 лет назад. Учитывая, что два вида гоминидов, вероятно, сосуществовали в Европе, антропологи давно задавались вопросом, взаимодействовали ли эти два вида.[19] Вопрос был решен только в 2010 г., когда было установлено, что в евразийских популяциях присутствует примесь неандертальцев, оцениваемая в среднем в 1,5–2,1%.[20] Возник вопрос, имело ли место это смешение в Европе или, скорее, в Леванте до миграции AMH в Европу.
Также было предположение о наследовании определенных генов от неандертальцев. Например, один MAPT локус 17q 21.3, который разделен на глубокие генетические линии H1 и H2. Поскольку линия H2, по-видимому, ограничивается европейскими популяциями, несколько авторов выступали за наследование от неандертальцев, начиная с 2005 года.[21][22][23][24][25]Однако предварительные результаты секвенирования полного генома неандертальцев в то время (2009 г.) не смогли выявить доказательств скрещивания между неандертальцами и современными людьми.[26][27]К 2010 году выводы Сванте Пяабо (Институт эволюционной антропологии Макса Планка в Лейпциге, Германия), Ричард Э. Грин (Калифорнийский университет, Санта-Крус) и Дэвид Райх (Гарвардская медицинская школа), сравнивая генетический материал от костей трех неандертальцев с генетическим материалом пяти современных людей, действительно показала связь между неандертальцами и современными людьми за пределами Африки.
Верхний палеолит

Считается, что современные люди начали населять Европу в период верхнего палеолита около 40 000 лет назад. Некоторые свидетельства показывают распространение Ориньяк культура.[28]
С чисто отцовской, Y-хромосома перспектива, возможно, что старые Гаплогруппа C1a2, F и / или E возможно, самые старые в Европе. Они были найдены в некоторых очень старых человеческих останках в Европе. Однако другие гаплогруппы гораздо чаще встречаются среди ныне живущих европейских мужчин.
Гаплогруппа I (M170), который сейчас относительно обычен и широко распространен в Европе, может представлять собой маркер палеолита - его возраст оценивается примерно в 22000 лет назад. Хотя сейчас он сконцентрирован в Европе, он, вероятно, возник у мужчины из Средний Восток или же Кавказ, или их ближайшие потомки, c. 20-25 000 лет назад, когда он отклонился от своего непосредственного предка, гаплогруппа IJ. Примерно в это же время появилась культура верхнего палеолита, известная как Gravettian.[29]
Более ранние исследования Y-ДНК вместо этого были сосредоточены на гаплогруппа R1 (M173): самая многочисленная линия среди ныне живущих европейских мужчин; Считается также, что R1 появился примерно на 40 000 лет назад. Центральная Азия.[30][31] Однако теперь считается, что R1 появился значительно позже: исследование 2008 года датировало самого последнего общего предка гаплогруппы IJ 38 500, а гаплогруппы R1 18 000 лет назад. Это говорит о том, что колонисты гаплогруппы IJ сформировали первую волну, а гаплогруппа R1 появилась намного позже.[32]
Таким образом, генетические данные предполагают, что, по крайней мере, с точки зрения отцовского происхождения, отдельные группы современных людей пошли в Европу двумя путями: с Ближнего Востока через Балканы, а другой из Центральной Азии через Балканы. Евразийская степь, к северу от Черное море.
Мартин Ричардс и другие. обнаружили, что 15-40% существующих клонов мтДНК восходят к миграциям палеолита (в зависимости от того, допускает ли один из них несколько событий основателя).[33] Гаплогруппа U5 мтДНК, датируемая ~ 40–50 кЯ, прибыла во время первой ранней колонизации верхнего палеолита. По отдельности он составляет 5–15% от общего числа линий мтДНК. Средний U.P. движения отмечены гаплогруппами HV, I и U4. HV разделилась на Pre-V (возраст около 26000 лет) и более крупную ветвь H, обе из которых распространились по Европе, возможно, через граветтианские контакты.[29][34]
Гаплогруппа H составляет около половины генных линий в Европе с множеством подгрупп. Вышеупомянутые линии мтДНК или их предшественники, скорее всего, прибыли в Европу через Ближний Восток. Это контрастирует с Y Доказательства ДНК, при этом около 50% плюс мужских линий характеризуются суперсемейством R1, которое, возможно, имеет центральноазиатское происхождение. Орнелла Семино постулирует, что эти различия «могут быть частично связаны с очевидным более поздним молекулярным возрастом Y-хромосом по сравнению с другими локусами, что предполагает более быструю замену предыдущих Y-хромосом. Гендерные различия в миграционном демографическом поведении также будут влиять на наблюдаемые модели мтДНК и Y-вариация ».
Последний ледниковый максимум
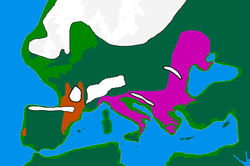
Последний ледниковый максимум («LGM») начался c. 30 тыс. До н. Э., В конце MIS 3, что привело к депопуляции Северной Европы. Согласно классической модели, люди укрывались в климатических заповедниках (или рефугиумах) следующим образом:
- Северная Иберия и Юго-Запад Франция, вместе составляющие "франко-кантабрийский" рефугиум
- Балканы
- Украина и вообще северное побережье Черного моря[29]
- Италия.[35]
Это событие привело к снижению общего генетического разнообразия в Европе в результате «дрейфа, соответствующего предполагаемому узкому месту в популяции во время последнего ледникового максимума».[36] Когда ледники отступили примерно 16 000–13 000 лет назад, Европа начала медленно заселяться людьми из рефугиумов, оставляя генетические следы.[37]
Некоторые клады гаплогруппы Y отошли от своих родительских гаплогрупп где-то во время или вскоре после LGM.[38] Гаплогруппа I2 преобладает на западных Балканах, а также в остальной части юго-восточной и центрально-восточной Европы с более умеренными частотами. Его частота быстро падает в Центральной Европе, что позволяет предположить, что выжившие, несущие линии I2, распространились преимущественно через Юго-Восточную и Центрально-Восточную Европу.[39]
Чинниоглу видит доказательства существования анатолийского убежища, в котором также находилась Hg R1b1b2.[40] Сегодня R1b доминирует в ландшафте y-хромосомы в Западной Европе, включая Британские острова, что указывает на то, что могли быть большие изменения в составе населения, основанные на миграциях после LGM.
Семино, Пассарино и Перичич ставят истоки гаплогруппы R1a внутри украинского Ледниковый период убежище. Его нынешнее распространение в Восточной Европе и некоторых частях Скандинавии частично отражает повторное заселение Европы из южных российских / украинских степей после Поздний ледниковый максимум.[41][42][43]
С точки зрения мтДНК Ричардс и другие. обнаружили, что большая часть разнообразия мтДНК в Европе объясняется послеледниковыми повторными экспансиями во время позднего верхнего палеолита / мезолита. «Региональный анализ в некоторой степени подтверждает предположение о том, что большая часть Западной и Центральной Европы была повторно заселена в основном с юго-запада, когда климат улучшился. Участвующие линии включают большую часть наиболее распространенной гаплогруппы H, а также большую часть K, T, W и X. " В ходе исследования не удалось определить, были ли в этот период новые миграции линий мтДНК с Ближнего Востока; значительный вклад был сочтен маловероятным.[18]
Альтернативная модель увеличения числа беженцев обсуждалась Bilton et al.[44]
Изучив 51 человека, исследователи смогли идентифицировать пять отдельных генетических кластеров древних европейцев во время LGM: Вестоницкий кластер (34 000–26 000 лет назад), связанные с Граветтовская культура; кластер Мальта (24 000–17 000), связанный с Мальта-буретская культура, то Кластер Эль-Мирон (19 000–14 000 лет назад), связанные с Магдаленская культура; то Кластер Виллабруна (14 000–7 000 лет назад) и Кластер Сатсурблия (13 000–10 000 лет назад).[17]
Примерно 37000 лет назад все древние европейцы стали иметь какое-то общее происхождение с современными европейцами.[17] Эта популяция-основательница представлена GoyetQ116-1, 35000-летним экземпляром из Бельгии.[17] Эта линия исчезает из записи и не встречается снова до 19 000 лет назад в Испании в Эль-Мироне, что свидетельствует о сильном родстве с GoyetQ116-1.[17] В этот промежуток времени в Европе преобладает отчетливое скопление Вестонице, даже на Goyet.[17] Повторное расширение скопления Эль-Мирон совпало с потеплением температур после отступления ледников во время Последний ледниковый максимум.[17] От 37000 до 14000 лет назад население Европы состояло из изолированной популяции, происходящей от основной популяции, которая не скрещивалась существенно с другими популяциями.[45]
Мезолит
Мезолит (пост-LGM) популяции значительно разошлись из-за их относительной изоляции в течение нескольких тысячелетий, из-за жесткого давления отбора во время LGM и из-за эффекты основателя вызвано быстрым расширением от LGM Refugia в начале мезолита.[8]К концу LGM, примерно от 19 до 11 тыс. Лет назад, появились знакомые разновидности евразийских фенотипов. Тем не менее, родословная мезолитических охотников-собирателей Западной Европы (WHG) не сохранилась в качестве основного вклада в какое-либо современное население. Скорее всего, они были голубоглазыми и сохранили темную пигментацию кожи, как до LGM EEMH.[46] В HERC2 и OCA2 вариации для голубых глаз происходят от линии WHG, также были обнаружены в Ямный народ.[46][противоречивый ]
Около 14000 лет назад Кластер Виллабруна отошли от сродства GoyetQ116-1 и начали проявлять большую сродство с Ближним Востоком, что совпало с потеплением Bølling-Allerød межстадиальный.[17] Этот генетический сдвиг показывает, что население Ближнего Востока, вероятно, уже начало перемещаться в Европу в конце верхнего палеолита, примерно на 6000 лет раньше, чем предполагалось ранее, до появления земледелия.[45] Несколько экземпляров из кластера Виллабруна также показывают генетическое родство с жителями Восточной Азии, которое происходит из потока генов.[17][45]В HERC2 Вариация голубых глаз впервые появилась около 13–14 тысяч лет назад в Италии и на Кавказе.[17]В светлая кожа пигментация, характерная для современных европейцев, по оценкам, распространилась по Европе «выборочно» во время мезолита (19–11 тыс. лет назад). Связанный TYRP1 SLC24A5 и SLC45A2 аллели появляются около 19 тыс. лет назад, еще во время LGM, скорее всего, на Кавказе.[47][48]
Неолит

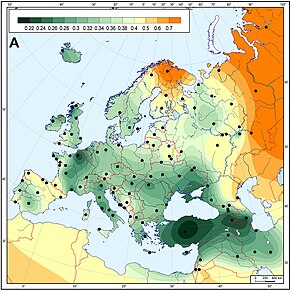
Большая линия генетической изменчивости, которая давно признана в Европе, похоже, демонстрирует важные отклонения со стороны Ближнего Востока. Это часто связывают с распространением сельскохозяйственных технологий в период неолита, который считается одним из самых важных периодов в определении современного европейского генетического разнообразия.
Неолит начался с распространения земледелия в юго-восточной Европе примерно в 10–3000 гг. До н.э. и распространился на северо-запад Европы между 4500–1700 гг. До н. Э. В эту эпоху Неолитическая революция привело к резким экономическим, а также социально-культурным изменениям в Европе, и считается, что это оказало большое влияние на генетическое разнообразие Европы, особенно в отношении генетических линий, проникающих в Европу с Ближнего Востока на Балканы. В этот период было несколько фаз:
- В позднеевропейском мезолите, прелюдии к неолиту, кажется, что народы Ближнего Востока из районов, где уже было сельское хозяйство, а также технологии мореплавания, временно пребывали в Греции (например, в Пещера Франчти ).[51][52]
- Есть консенсус, что агротехника и основные породы животных и растений, которые разводятся, попали в Европу откуда-то из Плодородный Полумесяц и особенно Левант регион из Синай к Южная Анатолия.[53][54] (Менее точно это сельскохозяйственная революция иногда утверждают, что это, в свою очередь, было частично вызвано перемещением людей и технологий, пересекающих Синай из Африки.) Плодородный полумесяц: распространение Cosmopolitan.
- Поздний этап неолита, так называемый Керамика эпохи неолита, увидел введение керамика в Левант, Балканы и Южную Италию (он присутствовал на территории современного Судана в течение некоторого времени, прежде чем был обнаружен в Восточное Средиземноморье, но считается, что он развился независимо), и, возможно, это также был период культурного переноса из Леванта на Балканы.
Важным вопросом, касающимся генетического воздействия неолитических технологий в Европе, является то, каким образом они были перенесены в Европу. Земледелие было введено значительной миграцией фермеров с Ближнего Востока (биологические исследования Кавалли-Сфорца). демическая диффузия модель) или "культурное распространение "или их комбинация, и популяционные генетики попытались выяснить, соответствуют ли какие-либо генетические признаки ближневосточного происхождения маршрутам экспансии, постулируемым археологическими свидетельствами.[55]
Мартин Ричардс подсчитал, что только 11% европейской мтДНК связано с иммиграцией в этот период, предполагая, что земледелие было распространено в первую очередь из-за того, что оно было принято коренным мезолитическим населением, а не из-за иммиграции с Ближнего Востока. Поток генов из Юго-Восточной Европы в Северо-Западную Европу, по-видимому, продолжался в неолите, и процентная доля значительно снижалась к Британским островам. Классическая генетика также предположил, что самая большая примесь европейского палеолита / мезолита была связана с Неолитическая революция VII-V тыс. до н. э.[56] Были идентифицированы три основные группы генов мтДНК, внесшие вклад в развитие Европы в эпоху неолита: J, T1 и U3 (в указанном порядке важности). С другими они составляют около 20% от Генофонд.[33]
В 2000 году исследование Semino ДНК Y выявило наличие гаплотипов, принадлежащих к большой кладе. E1b1b1 (E-M35). Они были преимущественно найдены на южных Балканах, юге Италии и некоторых частях Иберии. Семино связал этот паттерн, наряду с субкладами гаплогруппы J, с Y-ДНК компонентом неолитической демической диффузии фермеров с Ближнего Востока Кавалли-Сфорца.[57] Россер и др. скорее рассматривали это как (прямой) «североафриканский компонент» европейской генеалогии, хотя они не предлагали время и механизм для его объяснения.[58] Андерхилл и Кивисилд (2007) также описал E1b1b как представитель позднегоПлейстоцен миграция из Африки в Европу за Синайский полуостров в Египет, свидетельство чего не обнаруживается в митохондриальной ДНК.[59]

Что касается времени распространения и разнообразия V13, однако, Battaglia et al. (2008) предложили более раннее движение, в соответствии с которым линия E-M78 *, являющаяся предком всех современных мужчин E-V13, быстро покинула южную египетскую родину и прибыла в Европу только с Мезолит технологии. Затем они предполагают, что подкласс E-V13 от E-M78 расширился только впоследствии, когда коренные балканские «фуражиры-фермеры» переняли технологии неолита с Ближнего Востока. Они предполагают, что первое крупное рассеяние E-V13 с Балкан могло произойти в направлении Адриатическое море с Неолит Впечатленная посуда культура часто упоминается как Impressa или же Кардиальный. Peričic et al. (2005), скорее предполагают, что основной маршрут распространения E-V13 пролегал по системе «автомагистралей» реки Вардар-Морава-Дунай.
В отличие от Баттальи, Cruciani et al. (2007) предварительно предложил (i) другую точку, где мутация V13 произошла на пути из Египта на Балканы через Ближний Восток, и (ii) более позднее время распространения. Авторы предположили, что мутация V13 впервые появилась в Западной Азии, где она встречается с низкой, но значительной частотой, откуда она проникла на Балканы примерно через 11 тыс. Лет назад. Позже он испытал быстрое распространение, которое он датировал ок. 5300 лет назад в Европе, что совпало с эпохой балканской бронзы. Как Peričic et al. они считают, что «рассеяние гаплогрупп E-V13 и J-M12, по-видимому, в основном произошло по речным водным путям, соединяющим южные Балканы с северной и центральной Европой».
В последнее время, Lacan et al. (2011) объявил, что 7000-летний скелет в контексте неолита в испанской погребальной пещере был мужчиной E-V13. (Остальные образцы, протестированные на том же участке, находились в гаплогруппа G2a, который был обнаружен в контексте неолита по всей Европе.) Используя 7 STR-маркеров, этот образец был идентифицирован как похожий на современных людей, протестированных в Албания, Босния, Греция, Корсика, и Прованс. Поэтому авторы предположили, что, независимо от того, является ли современное распространение E-V13 сегодня результатом более недавних событий, E-V13 уже находился в Европе в эпоху неолита и был перенесен ранними фермерами из Восточного Средиземноморья в Западное Средиземноморье. намного раньше бронзового века. Это поддерживает предложения Battaglia et al. а не Cruciani et al. по крайней мере, в отношении самых ранних европейских разгонов, но E-V13, возможно, разошелся более одного раза. Было высказано предположение, что современное распространение E-V13 в Европе даже позже, чем бронзовый век, по крайней мере частично вызвано перемещениями людей римской эпохи.[60] (Смотри ниже.)
Миграция неолитических фермеров в Европу принесла с собой несколько новых приспособлений.[46] Вариация светлого цвета кожи была введена в Европу неолитические фермеры.[46] После прихода неолитических фермеров SLC22A4 была выбрана мутация, которая, вероятно, возникла для борьбы с эрготионеин дефицит, но увеличивает риск язвенный колит, глютеновая болезнь, и синдром раздраженного кишечника.
Бронзовый век
В Бронзовый век увидел развитие дальней торговые сети, особенно вдоль Атлантического побережья и в долине Дуная. Произошла миграция из Норвегии в Оркнейские острова и Шетландские острова в этот период (и в меньшей степени в материковой Шотландии и Ирландии). Также была миграция из Германии в восточная англия. Мартин Ричардс подсчитал, что в эпоху бронзы иммиграция мтДНК в Европу составляла около 4%.
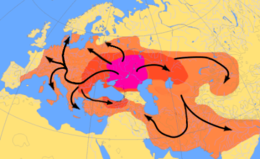
Еще одна теория о происхождении Индоевропейский язык сосредотачивается вокруг гипотетического Протоиндоевропейский люди, которые, по мнению Курганская гипотеза, можно проследить к северу от Черного и Каспийского морей примерно в 4500 году до нашей эры.[61] Они приручил лошадь и, возможно, изобрел деревянный диск колесо, и считается, что они распространили свою культуру и гены по всей Европе.[62] Oни гаплогруппа R1a является предполагаемым маркером этих "курганских" генов, как и Y Гаплогруппа R1b, хотя эти гаплогруппы в целом могут быть намного старше языковой семьи.[63]
На крайнем севере носители Y-гаплогруппа N прибыл в Европу из Сибирь, в конечном итоге расширившись до Финляндия, хотя точное время их прибытия неизвестно. Возраст наиболее распространенного субклада N1c1 в Северной Европе оценивается примерно в 8000 лет. Есть свидетельства поселений людей в Финляндии, относящихся к 8500 г. до н.э., связанных с Кунда культура и его предполагаемый предок, Свидерская культура, но считается, что последний имеет европейское происхождение. Географическое распространение гаплогруппы N в Европе хорошо согласуется с Культура ямочно-гребенчатой посуды, появление которого обычно датируется ок. 4200 г. до н.э., а с распределением Уральские языки. Исследования митохондриальной ДНК Саамский народ, гаплогруппа U5 согласуются с несколькими миграциями в Скандинавия из Волга -Урал области, начиная с 6000 до 7000 лет назад.[64]
Взаимосвязь между ролями европейских и азиатских колонистов в предыстории Финляндии является предметом некоторых споров, и некоторые ученые настаивают на том, что финны «преимущественно восточноевропейские и состоят из людей, которые отправились на север из украинского убежища во время ледникового периода».[65] Дальше на восток вопрос менее спорный. Носители гаплогруппы N составляют значительную часть всех неславянских этнических групп в северных регионах. Россия, в том числе 37% Карелы, 35% Коми народ (65% согласно другому исследованию[66]), 67% Марийский народ, аж 98% Ненцы, 94% Нганасаны, и от 86% до 94% Якуты.[67]
Компонент Ямная содержит частичное происхождение от компонента Древнего Севера Евразии, впервые идентифицированного в Мальта.[68] По словам Иосифа Лазаридиса, «древнее североевразийское происхождение является пропорционально наименьшим компонентом повсюду в Европе, никогда не превышает 20 процентов, но мы находим его почти в каждой европейской группе, которую мы изучили».[69] Этот генетический компонент происходит не напрямую от самой линии передачи Мальта, а из родственной линии, которая отделилась от линии передачи Мальта.[17]
До половины ямной составляющей могло произойти из Кавказский охотник-собиратель прядь.[68] 16 ноября 2015 г. в исследовании, опубликованном в журнале Nature Communications,[68] генетики объявили, что они нашли новое четвертое наследственное «племя» или «цепь», которая внесла свой вклад в современный европейский генофонд. Они проанализировали геномы двух охотников-собирателей из Грузии, которым было 13 300 и 9700 лет, и обнаружили, что эти кавказские охотники-собиратели, вероятно, были источником фермерской ДНК в Ямне. По словам соавтора доктора Андреа Маника из Кембриджского университета: «Вопрос о том, откуда взялись Ямная, до сих пор оставался загадкой ... теперь мы можем ответить на этот вопрос, поскольку мы обнаружили, что их генетическое происхождение. -up представляет собой смесь Восточноевропейские охотники-собиратели и популяция из этого кармана кавказских охотников-собирателей, которые пережили большую часть последнего ледникового периода в очевидной изоляции ».[70]
По данным Lazaridis et al. (2016), население, связанное с народом Энеолит Иран составляли примерно половину предков ямных популяций Понтийско-Каспийской степи. Эти иранские люди эпохи энеолита были смесью «неолитических людей западного Ирана, Леванта и кавказских охотников-собирателей».[71]
Генетические вариации для стойкость лактазы и большая высота пришла с ямным народом.[46] Производный аллель KITLG ген (SNP rs12821256), который связан с - и, вероятно, является причиной - белокурые волосы у европейцев встречается в популяциях с восточными, но не Западные охотники-собиратели происхождение, предполагая, что его происхождение находится в Древний Север Евразии (ANE) и, возможно, были распространены в Европе людьми с степное происхождение. В соответствии с этим, самый ранний известный человек с производным аллелем - это человек ANE из позднего верхнего палеолита. Афонтова гора археологический комплекс в центральной Сибири.[72]
Недавняя история
В период Римская империя, исторические источники показывают, что было много перемещений людей по Европе, как внутри, так и за пределами Империи. В исторических источниках иногда упоминаются случаи геноцида, совершенного римлянами против мятежных провинциальных племен. Если бы это действительно произошло, это было бы ограничено, учитывая, что современные популяции демонстрируют значительную генетическую преемственность в своих соответствующих регионах.[нужна цитата ]. Процесс 'Романизация 'похоже, было достигнуто путем колонизации провинций несколькими латынскими администраторами, военнослужащие, оседлые ветераны и частные лица (купцы, торговцы), которые происходили из различных регионов Империи (а не только из Римская Италия ). Они послужили ядром для аккультурации местной знати.[74]
Учитывая их небольшое количество и различное происхождение, романизация, похоже, не оставила отчетливых генетических признаков в Европе. Действительно, романскоязычное население Балкан, как и Румыны, Арумыны, Молдаване и т. д., как было установлено, генетически напоминают соседних греческих и южнославяноязычных народов, а не современных итальянцев, доказывая, что они генетически говорили, в основном через I2a2 М-423 и E1b1b1, V-13 Гаплогруппы родом из этой области.[75][76]
Стивен Берд предположил, что E1b1b1a распространился во время Римская эпоха через фракийский и Дак популяции с Балкан в остальную Европу.[60]
Что касается позднего римского периода (не только) Германский "Völkerwanderung ", были сделаны некоторые предположения, по крайней мере, для Великобритании, с гаплогруппой Y I1a, связанной с Англосаксонский иммиграция в восточную Англию, а R1a связан со скандинавской иммиграцией в северной Шотландии.[77]
Генетика современных европейских популяций
Патрилинейные исследования
Есть четыре основных ДНК Y-хромосомы гаплогруппы которые составляют большую часть Европы по отцовской линии.[78]
- Гаплогруппа I встречается в виде различных подкладов по всей Европе, а самые высокие частоты встречаются в Скандинавские страны как I1 (Норвегия, Дания, Швеция, Финляндия ) и в Балканский полуостров как I2a (Босния и Герцеговина 65%,[79] Хорватия и Сербия ). I1 также часто встречается в Германия, Великобритания и Нидерланды, а I2a часто встречается и в Сардиния, Румыния /Молдова, Болгария и Украина. Эта клада наиболее ярко выражена далеко в Европе и, возможно, существовала там еще до LGM.[38]
- Гаплогруппа E1b1b (ранее известная как E3b) представляет собой последнюю крупную прямую миграцию из Африки в Европу. Считается, что он впервые появился на Африканском Роге примерно 26000 лет назад и распространился по Северной Африке и Ближнему Востоку в периоды позднего палеолита и мезолита. Клоны E1b1b тесно связаны с распространением Афроазиатские языки. Хотя он присутствует по всей Европе, его пик приходится на западные Балканы среди албанцев и их соседей. Также распространен в Италии и на Пиренейском полуострове. Гаплогруппа E1b1b1, в основном в форме субклада E1b1b1a2 (E-V13), достигает частот выше 47% в области Косово.[80] Считается, что эта клада пришла в Европу из Западной Азии либо в позднем мезолите,[81] или неолит.[82] Субклад E-M81 из Северной Африки также присутствует в Сицилии и Андалусии.
- Гаплогруппа R1b распространен по всей Европе, с R1b1a1a2 особенно часто встречается в западная Европа.[83][84][85] Почти весь этот R1b в Европе находится в форме подклада R1b1a2 (название 2011 г.) (R-M269), в частности, внутри подклада R-L23, тогда как R1b обнаружен в подкладе Центральная Азия, западная азия и Африка имеет тенденцию быть в других кладах. Также указывалось, что выброс типы присутствуют в Европе и особенно заметны в некоторых областях, таких как Сардиния и Армения.[86] Частоты гаплогруппы R1b колеблются от максимумов в Западной Европе и имеют неуклонно уменьшающуюся границу с удалением от Атлантики: 80–90% (валлийский, Баскский, Ирландский, Шотландцы, Бретонцы ) около 70–80% в других регионах Испании, Великобритании и Франции и около 40–60% в большинстве других частей западная Европа подобно восточная Германия, и северо-центральная Италия. За пределами этой области он падает и составляет около 30% или меньше в таких областях, как юг Италии, Швеция, Польша, Балканы и Кипр. R1b остается наиболее распространенной кладой по мере продвижения на восток, в Германию, а дальше на восток, в Польше, R1a более распространен (см. Ниже).[87] В Юго-Восточная Европа, R1b отстает от R1a в районе Венгрии и Сербии и вокруг них, но чаще встречается как на юге, так и на севере этого региона.[88] R1b в Западной Европе преобладает по крайней мере двумя подкладами, R-U106, которые распространяются с восточной стороны Рейн в северную и центральную Европу (с сильным присутствием в Англии) и R-P312, который наиболее распространен к западу от Рейна, включая Британские острова.[84][85] Некоторые утверждают, что присутствие этой гаплогруппы в Европе восходит к LGM, а другие связывают это с распространением Centum филиал Индоевропейские языки.
- Самые старые человеческие останки, содержащие R1b, принадлежат человеку из Эпиграветтин культурный контекст в Италии (Виллабруна), который жил ок. 12000 г. до н.э. и, как сообщается, принадлежали R1b1a (L754), а останки охотника-собирателя возрастом 7000 лет, принадлежащие Самарская культура из района реки Волги, несшие R1b1 * (R-L278 *).
- Гаплогруппа R1a, почти полностью в подкладе R1a1a, преобладает в большинстве Восточная и Центральная Европа (Также в юг и Центральная Азия ). Например, наблюдается резкое увеличение R1a1 и уменьшение R1b1b2 по мере продвижения на восток от Германии в Польшу.[87] Он также имеет значительное присутствие в Скандинавии (особенно в Норвегии).[89][90] В странах Балтии частоты R1a снижаются от Литвы (45%) до Эстонии (около 30%).[91]
Помимо небольших анклавов, есть также несколько гаплогрупп, помимо вышеперечисленных четырех, которые менее заметны или наиболее распространены только в определенных частях Европы.
- Гаплогруппа G, исконно неолитические европейцы (кавказцы), распространены в большинстве частей Европы с низкой частотой, достигая пиков выше 70% около Грузия и среди Маджары (хотя проживая в Азии, они граничат с восточным периметром Европы), до 10% на Сардинии, 12% на Корсике и Упсале (Швеция), 11% на Балканах и Португалии, 10% в Испании и 9% в европейской части России. Эта клада также встречается на Ближнем Востоке.
- Гаплогруппа N, распространен только на северо-востоке Европы и в форме субклада N1c1 достигает частот примерно 60% среди финнов и примерно 40% среди эстонцев, латышей и литовцев.
- Гаплогруппа J2, в различных подкладах (J2a, J2b), обнаруживается на уровнях около 15–30% в некоторых частях Балкан и Италии и распространен по всей Европе и особенно в бассейне Средиземного моря.[92]
Матрилинейные исследования
Был проведен ряд исследований о гаплогруппы митохондриальной ДНК (мтДНК) в Европе. В отличие от гаплогрупп ДНК Y, гаплогруппы мтДНК не демонстрировали такой большой географической структуры, но были более равномерно распространены. За исключением отдаленных саамов, для всех европейцев характерно преобладание гаплогрупп H, U и T. Отсутствие наблюдаемой географической структуры мтДНК может быть связано с социокультурными факторами, а именно с феноменом многоженство и патрилокальность.[93]
Генетические исследования предполагают некоторый поток материнских генов в Восточную Европу из Восточной Азии или южной Сибири 13 000 - 6600 лет. BP.[94] Анализ скелетов эпохи неолита в Великая Венгерская равнина обнаружили высокую частоту восточноазиатских гаплогрупп мтДНК, некоторые из которых выживают в современных популяциях Восточной Европы.[94] Поток материнских генов в Европу из Африки к югу от Сахары начался уже 11000 лет назад, хотя, по оценкам, большинство родословных, примерно 65%, появилось совсем недавно, в том числе в период романизации, арабских завоеваний южной Европы и во время атлантической работорговли.[95]
Подструктура населения Европы
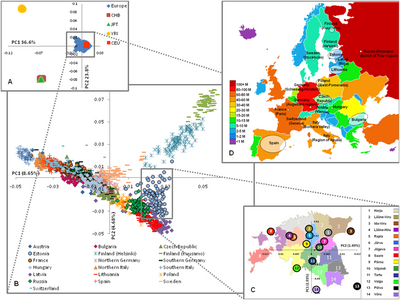
Генетически Европа относительно однородна, но были обнаружены отличные субпопуляционные модели различных типов генетических маркеров,[96] особенно вдоль склона юго-восток-северо-запад.[97] Например, анализ основных компонентов Кавалли-Сфорца выявил пять основных клинических паттернов по всей Европе, и аналогичные паттерны продолжали обнаруживаться в более поздних исследованиях.[98]
- Набор генов с наибольшей частотой в Средний Восток, распространяясь на самые низкие уровни к северо-западу. Кавалли-Сфорца первоначально описал это как верное отражение распространения сельского хозяйства во времена неолита. Это общая тенденция в интерпретации всех генов с этим паттерном.
- Группа генов с наибольшей частотой среди Финский и Саами на крайнем северо-востоке и распространяется до самых низких частот на юго-западе.
- Группа генов с наибольшими частотами в области нижнего Дон и Волга реки в юг россии и распространяется на самые низкие частоты в Испании, Южная Италия, Греция и районы, населенные Говорящие на саами на крайнем севере Скандинавия. Кавалли-Сфорца связал это с распространением индоевропейских языков, которое он, в свою очередь, связывает с «вторичной экспансией» после распространения сельского хозяйства, связанной с выпасом животных.
- Набор генов с наибольшей частотой в Балканы и Южная Италия, доходящая до самых низких уровней в Великобритании и Стране Басков. Кавалли-Сфорца связывает это с «греческой экспансией, которая достигла своего пика в исторические времена около 1000 и 500 гг. До н.э., но, безусловно, началась раньше».
- Набор генов с наибольшей частотой в Страна Басков, и более низкие уровни за пределами Иберии и Южная Франция. Возможно, в наиболее известном заключении Кавалли-Сфорца, этот самый слабый из пяти паттернов был описан как изолированные остатки донеолитического населения Европы, «которое, по крайней мере, частично противостояло экспансии культиваторов». Это примерно соответствует географическому распространению резус отрицательный группы крови. В частности, широко обсуждался вывод о том, что баски являются генетическим изолятом, но также и спорный.
Он также создал филогенетическое древо для анализа внутренних отношений между европейцами. Он обнаружил четыре основных «выброса» - Баски, Саами, Сардинцы и Исландцы;[99] результат, который он объяснил их относительной изоляцией (примечание: исландцы и сардинцы говорят Индоевропейские языки, а две другие группы - нет). Греки и Югославы представляют вторую группу менее экстремальных выбросов. Остальные популяции сгруппированы в несколько групп: "кельтская ", "Германский "," юго-западные европейцы ","Скандинавы »и« восточноевропейцы ».[100]
Исследование в мае 2009 г.[101] из 19 популяций из Европы с использованием 270 000 SNP подчеркнули генетическое разнообразие европейских популяций, соответствующее градиенту с северо-запада на юго-восток, и выделили «четыре отдельных региона» в Европе:
- Финляндия, показав наибольшее расстояние от остальных европейцев.
- то Балтийский регион (Эстония, Латвия и Литва ), западный Россия и восточный Польша.
- Центральная и Западная Европа.
- Италия, "южные итальянцы более далеки".
В этом исследовании анализ барьеров выявил «генетические барьеры» между Финляндией, Италией и другими странами, и эти барьеры также могут быть продемонстрированы внутри Финляндии (между Хельсинки и Куусамо) и Италии (между северной и южной частью, Fst = 0,0050). Fst (Индекс фиксации ) было обнаружено, что она значительно коррелирует с географическими расстояниями от ≤0,0010 для соседних популяций до 0,0200–0,0230 для южной Италии и Финляндии. Для сравнения, попарные Fst неевропейских выборок составили: европейцы - африканцы (йоруба) 0,1530; Европейцы - китайцы 0,1100; Африканцы (йоруба) - китайцы 0,1900.[102]
Исследование, проведенное Чао Тиан в августе 2009 г., расширило анализ генетической структуры европейской популяции, включив в нее дополнительные южноевропейские группы и арабское население (Палестинцы, Друзы...) с Ближнего Востока. Это исследование определило аутосомный Fst между 18 группами населения и пришло к выводу, что в целом генетические расстояния соответствуют географическим отношениям с меньшими значениями между группами населения с происхождением из соседних стран / регионов (например, Греки /Тосканцы: Fst = 0,0010, Греки /Палестинцы: Fst = 0,0057) по сравнению с данными из самых разных регионов Европы (например, Греки /Шведский: Fst = 0,0087, Греки /Россияне: Fst = 0,0108).[103][104]
Аутосомная ДНК
Селдин (2006) использовал более 5000 аутосомных SNP. Он показал «последовательное и воспроизводимое различие между« северным »и« южным » Европейское население группы ". Большинство индивидуальных участников с южноевропейский происхождение (Итальянцы, Греки, португальский, Испанцы ), и Евреи ашкенази имеют> 85% -ную принадлежность к южному населению; и большинство жителей Северной, Западной, Центральной и Восточной Европы (Шведы, английский, Ирландский, Немцы, и Украинцы ) имеют> 90% в северной группе населения. Тем не менее, многие из участников этого исследования на самом деле были американскими гражданами, которые идентифицировали себя с разными европейскими этническими группами на основании собственной семейной родословной.[105]
Аналогичное исследование 2007 года с использованием образцов, преимущественно из Европы, показало, что наиболее важная генетическая дифференциация в Европе происходит на линии с севера на юго-восток (Северная Европа на Балканы), с другой осью дифференциации восток-запад через Европу. Его выводы согласуются с более ранними результатами, основанными на мтДНК и Y-хромосомной ДНК, которые подтверждают теорию о том, что современные иберийцы (испанские и португальские) имеют самую древнюю европейскую генетическую родословную, а также отделяют басков и саамов от других европейских популяций.[106]
Он предположил, что английский и ирландский кластер с другими северными и восточными европейцами, такими как Немцы и Поляки, в то время как некоторые баски и итальянцы также сгруппировались с северными европейцами. Несмотря на эти стратификации, он отметил, что «в Европе наблюдается низкое видимое разнообразие, при этом выборки всего континента лишь незначительно более разбросаны, чем отдельные выборки населения в других странах мира».[106]
В 2008 году две международные исследовательские группы опубликовали результаты анализа крупномасштабного генотипирования больших выборок европейцев с использованием более 300 000 аутосомных SNP. За исключением обычных изолятов, таких как Баски, Финны и Сардинцы, в европейской популяции отсутствовали резкие разрывы (кластеризация), как показали предыдущие исследования (см. Селдин и другие. 2006 и Bauchet и другие. 2007[106]), хотя был заметный градиент с юга на север. В целом, они обнаружили только низкий уровень генетической дифференциации между субпопуляциями, а различия, которые действительно существовали, характеризовались сильной общеконтинентальной корреляцией между географической и генетической дистанцией. Кроме того, они обнаружили, что наибольшее разнообразие наблюдается в южной Европе из-за более крупного эффективного размера популяции и / или рост населения от южной до северной Европы.[107] Исследователи считают, что это наблюдение подразумевает, что генетически европейцы не разделены на отдельные популяции.[108][109]
Исследование северо-восточного населения, опубликованное в марте 2013 года, показало, что Коми народы сформировали полюс генетического разнообразия, отличного от других популяций.[110]
Аутосомные генетические дистанции (Fst) на основе SNP (2009)
Генетическое расстояние между популяциями часто измеряется Индекс фиксации (Fst) на основе данных генетического полиморфизма, таких как однонуклеотидные полиморфизмы (SNP) или же микроспутники. Fst - частный случай F-статистика, концепция, разработанная в 1920-х гг. Сьюэлл Райт. Fst - это просто корреляция случайно выбранных аллелей в одной и той же подгруппе населения по сравнению с аллелями, обнаруженными во всей популяции. Его часто выражают как долю генетического разнообразия из-за различий в частоте аллелей среди популяций.
Диапазон значений от 0 до 1. Нулевое значение означает, что две популяции являются панмиктическими, что они свободно скрещиваются. Значение единицы будет означать, что две популяции полностью разделены. Чем больше значение Fst, тем больше генетическая дистанция. По сути, эти низкие значения Fst предполагают, что большая часть генетической изменчивости находится на уровне индивидов в одной и той же группе населения (~ 85%); принадлежность к разным группам населения внутри одной «расы» / континента и даже к разным расовым / континентальным группам добавляла гораздо меньшую степень вариации (3–8%; 6–11%, соответственно).
| Итальянские американцы | Палестинцы | Шведы | Друзы | Испанцы | Немцы | Россияне | Ирландский | Греческие американцы | Евреи ашкенази | Черкесы | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Итальянские американцы | 0.0064 | 0.0064 | 0.0057 | 0.0010 | 0.0029 | 0.0088 | 0.0048 | 0.0000 | 0.0040 | 0.0067 | |
| Палестинцы | 0.0064 | 0.0191 | 0.0064 | 0.0101 | 0.0136 | 0.0202 | 0.0170 | 0.0057 | 0.0093 | 0.0108 | |
| Шведы | 0.0064 | 0.0191 | 0.0167 | 0.0040 | 0.0007 | 0.0030 | 0.0020 | 0.0084 | 0.0120 | 0.0117 | |
| Друзы | 0.0057 | 0.0064 | 0.0167 | 0.0096 | 0.0121 | 0.0194 | 0.0154 | 0.0052 | 0.0088 | 0.0092 | |
| Испанцы | 0.0010 | 0.0101 | 0.0040 | 0.0096 | 0.0015 | 0.0070 | 0.0037 | 0.0035 | 0.0056 | 0.0090 | |
| Немцы | 0.0029 | 0.0136 | 0.0007 | 0.0121 | 0.0015 | 0.0030 | 0.0010 | 0.0039 | 0.0072 | 0.0089 | |
| Россияне | 0.0088 | 0.0202 | 0.0030 | 0.0194 | 0.0070 | 0.0030 | 0.0038 | 0.0108 | 0.0137 | 0.0120 | |
| Ирландский | 0.0048 | 0.0170 | 0.0020 | 0.0154 | 0.0037 | 0.0010 | 0.0038 | 0.0067 | 0.0109 | 0.0110 | |
| Греческие американцы | 0.0000 | 0.0057 | 0.0084 | 0.0052 | 0.0035 | 0.0039 | 0.0108 | 0.0067 | 0.0042 | 0.0054 | |
| Евреи ашкенази | 0.0040 | 0.0093 | 0.0120 | 0.0088 | 0.0056 | 0.0072 | 0.0137 | 0.0109 | 0.0042 | 0.0107 | |
| Черкесы | 0.0067 | 0.0108 | 0.0117 | 0.0092 | 0.0090 | 0.0089 | 0.0120 | 0.0110 | 0.0054 | 0.0107 |
| Австрия | Болгария | Чехия | Эстония | Финляндия (Хельсинки ) | Финляндия (Куусамо ) | Франция | Северная Германия | Южная Германия | Венгрия | Северная Италия | Южная Италия | Латвия | Литва | Польша | Россия | Испания | Швеция | Швейцария | CEU | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Австрия | 1.14 | 1.08 | 1.58 | 2.24 | 3.30 | 1.16 | 1.10 | 1.04 | 1.04 | 1.49 | 1.79 | 1.85 | 1.70 | 1.19 | 1.47 | 1.41 | 1.21 | 1.19 | 1.12 | Австрия | |
| Болгария | 1.14 | 1.21 | 1.70 | 2.19 | 2.91 | 1.22 | 1.32 | 1.19 | 1.10 | 1.32 | 1.38 | 1.86 | 1.73 | 1.29 | 1.53 | 1.30 | 1.47 | 1.13 | 1.29 | Болгария | |
| Чехия | 1.08 | 1.21 | 1.42 | 2.20 | 3.26 | 1.35 | 1.15 | 1.16 | 1.06 | 1.69 | 2.04 | 1.62 | 1.48 | 1.09 | 1.27 | 1.63 | 1.26 | 1.37 | 1.21 | Чехия | |
| Эстония | 1.58 | 1.70 | 1.42 | 1.71 | 2.80 | 2.08 | 1.53 | 1.70 | 1.41 | 2.42 | 2.93 | 1.24 | 1.28 | 1.17 | 1.21 | 2.54 | 1.49 | 2.16 | 1.59 | Эстония | |
| Финляндия (Хельсинки) | 2.24 | 2.19 | 2.20 | 1.71 | 1.86 | 2.69 | 2.17 | 2.35 | 1.87 | 2.82 | 3.37 | 2.31 | 2.33 | 1.75 | 2.10 | 3.14 | 1.89 | 2.77 | 1.99 | Финляндия (Хельсинки) | |
| Финляндия (Куусамо) | 3.30 | 2.91 | 3.26 | 2.80 | 1.86 | 3.72 | 3.27 | 3.46 | 2.68 | 3.64 | 4.18 | 3.33 | 3.37 | 2.49 | 3.16 | 4.21 | 2.87 | 3.83 | 2.89 | Финляндия (Куусамо) | |
| Франция | 1.16 | 1.22 | 1.35 | 2.08 | 2.69 | 3.72 | 1.25 | 1.12 | 1.16 | 1.38 | 1.68 | 2.40 | 2.20 | 1.44 | 1.94 | 1.13 | 1.38 | 1.10 | 1.13 | Франция | |
| Северная Германия | 1.10 | 1.32 | 1.15 | 1.53 | 2.17 | 3.27 | 1.25 | 1.08 | 1.11 | 1.72 | 2.14 | 1.84 | 1.66 | 1.18 | 1.49 | 1.62 | 1.12 | 1.36 | 1.06 | Северная Германия | |
| Южная Германия | 1.04 | 1.19 | 1.16 | 1.70 | 2.35 | 3.46 | 1.12 | 1.08 | 1.08 | 1.53 | 1.85 | 1.20 | 1.84 | 1.23 | 1.58 | 1.40 | 1.21 | 1.17 | 1.07 | Южная Германия | |
| Венгрия | 1.04 | 1.10 | 1.06 | 1.41 | 1.87 | 2.68 | 1.16 | 1.11 | 1.08 | 1.42 | 1.63 | 1.58 | 1.46 | 1.14 | 1.28 | 1.32 | 1.22 | 1.16 | 1.13 | Венгрия | |
| Северная Италия | 1.49 | 1.32 | 1.69 | 2.42 | 2.82 | 3.64 | 1.38 | 1.72 | 1.53 | 1.42 | 1.54 | 2.64 | 2.48 | 1.75 | 2.24 | 1.42 | 1.86 | 1.36 | 1.56 | Северная Италия | |
| Южная Италия | 1.79 | 1.38 | 2.04 | 2.93 | 3.37 | 4.18 | 1.68 | 2.14 | 1.85 | 1.63 | 1.54 | 3.14 | 2.96 | 1.99 | 2.68 | 1.67 | 2.28 | 1.54 | 1.84 | Южная Италия | |
| Латвия | 1.85 | 1.86 | 1.62 | 1.24 | 2.31 | 3.33 | 2.40 | 1.84 | 1.20 | 1.58 | 2.64 | 3.14 | 1.20 | 1.26 | 1.84 | 2.82 | 1.89 | 2.52 | 1.87 | Латвия | |
| Литва | 1.70 | 1.73 | 1.48 | 1.28 | 2.33 | 3.37 | 2.20 | 1.66 | 1.84 | 1.46 | 2.48 | 2.96 | 1.20 | 1.20 | 1.26 | 2.62 | 1.74 | 2.29 | 1.74 | Литва | |
| Польша | 1.19 | 1.29 | 1.09 | 1.17 | 1.75 | 2.49 | 1.44 | 1.18 | 1.23 | 1.14 | 1.75 | 1.99 | 1.26 | 1.20 | 1.18 | 1.66 | 1.30 | 1.46 | 1.28 | Польша | |
| Россия | 1.47 | 1.53 | 1.27 | 1.21 | 2.10 | 3.16 | 1.94 | 1.49 | 1.58 | 1.28 | 2.24 | 2.68 | 1.84 | 1.26 | 1.18 | 2.32 | 1.59 | 1.20 | 1.56 | Россия | |
| Испания | 1.41 | 1.30 | 1.63 | 2.54 | 3.14 | 4.21 | 1.13 | 1.62 | 1.40 | 1.32 | 1.42 | 1.67 | 2.82 | 2.62 | 1.66 | 2.32 | 1.73 | 1.16 | 1.34 | Испания | |
| Швеция | 1.21 | 1.47 | 1.26 | 1.49 | 1.89 | 2.87 | 1.38 | 1.12 | 1.21 | 1.22 | 1.86 | 2.28 | 1.89 | 1.74 | 1.30 | 1.59 | 1.73 | 1.50 | 1.09 | Швеция | |
| Швейцария | 1.19 | 1.13 | 1.37 | 2.16 | 2.77 | 3.83 | 1.10 | 1.36 | 1.17 | 1.16 | 1.36 | 1.54 | 2.52 | 2.29 | 1.46 | 1.20 | 1.16 | 1.50 | 1.21 | Швейцария | |
| CEU | 1.12 | 1.29 | 1.21 | 1.59 | 1.99 | 2.89 | 1.13 | 1.06 | 1.07 | 1.13 | 1.56 | 1.84 | 1.87 | 1.74 | 1.28 | 1.56 | 1.34 | 1.09 | 1.21 | CEU | |
| Австрия | Болгария | Чехия | Эстония | Финляндия (Хельсинки) | Финляндия (Куусамо) | Франция | Северная Германия | Южная Германия | Венгрия | Северная Италия | Южная Италия | Латвия | Литва | Польша | Россия | Испания | Швеция | Швейцария | CEU |
История исследования
Классические генетические маркеры (по доверенности)
Одним из первых ученых, проводивших генетические исследования, был Луиджи Лука Кавалли-Сфорца. Он использовал классические генетические маркеры для прокси-анализа ДНК. Этот метод изучает различия в частотах отдельных аллельных признаков, а именно: полиморфизмы из белков, обнаруженных в человеческая кровь (такой как Группы крови АВО, Антигены крови резуса, HLA локусы, иммуноглобулины, G6PD изоферменты, среди прочего). Впоследствии его команда подсчитала генетическая дистанция между популяциями, основанный на том принципе, что две популяции, которые имеют сходные частоты признака, более тесно связаны, чем популяции, которые имеют более разные частоты признака.[114]
Из этого он построил филогенетические деревья что схематически показало генетические дистанции. Его команда также выступила анализ главных компонентов, который хорошо анализирует многомерные данные с минимальной потерей информации. Утерянную информацию можно частично восстановить, создав второй главный компонент и так далее.[115] В свою очередь, информация от каждого отдельного главного компонента (ПК) могут быть представлены графически в синтетические карты. Эти карты показывают пики и впадины, которые представляют популяции, чьи частоты генов принимают крайние значения по сравнению с другими в изучаемой области.[114]
Пики и впадины, обычно соединенные плавными градиентами, называются клины. Генетические клины могут быть созданы адаптацией к окружающей среде (естественный отбор ), непрерывный поток генов между двумя изначально разными группами населения или демографическая экспансия в малонаселенную среду с небольшим начальным примесь с существующими популяциями.[116] Кавалли-Сфорца связал эти градиенты с постулируемыми доисторическими движениями населения, основанными на археологических и лингвистических теориях. Однако, учитывая, что временная глубина таких моделей неизвестна, «связывание их с конкретными демографическими событиями обычно является умозрительным».[93]
Прямой анализ ДНК
Исследования с использованием прямого анализа ДНК сейчас многочисленны и могут митохондриальная ДНК (мтДНК), нерекомбинирующая часть Y-хромосомы (NRY) или даже аутосомная ДНК. ДНК MtDNA и NRY имеют некоторые общие черты, которые сделали их особенно полезными в генетической антропологии. Эти свойства включают прямое, неизменное наследование мтДНК и ДНК NRY от матери к потомству и от отца к сыну, соответственно, без «скремблирующих» эффектов генетическая рекомбинация. Мы также предполагаем, что эти генетические локусы не подвержены влиянию естественного отбора и что основной процесс, ответственный за изменения в пар оснований произошла мутация (что можно подсчитать).[117]
Меньший эффективная численность населения NRY и мтДНК усиливает последствия дрейфа и эффекта основателя по сравнению с аутосомами, что делает вариацию NRY и мтДНК потенциально чувствительным показателем состава популяции.[93][33][118] Эти биологически правдоподобные предположения не являются конкретными; Россер предполагает, что климатические условия могут влиять на плодовитость определенных линий.[93]
Лежащий в основе скорость мутации используется генетиками более сомнительно. Они часто используют разную частоту мутаций, и исследования часто приходят к совершенно разным выводам.[93] NRY и мтДНК могут быть настолько восприимчивы к дрейфу, что некоторые древние паттерны могут быть скрыты. Другое предположение состоит в том, что генеалогии населения аппроксимируются аллельные генеалогии. Гвидо Барбужани указывает, что это справедливо только в том случае, если группы населения развиваются из генетически мономорфного набора основателей. Барбужани утверждает, что нет оснований полагать, что Европа была колонизирована мономорфными популяциями. Это привело бы к переоценке возраста гаплогруппы, тем самым ошибочно распространяя демографическую историю Европы на Поздний палеолит а не Неолит эпоха.[119] Большую уверенность в хронологии можно получить из исследований древней ДНК (см. Ниже), но до сих пор их было сравнительно немного.
В то время как Y-ДНК гаплогруппы мтДНК представляют собой лишь небольшой компонент пула ДНК человека, аутосомная ДНК имеет то преимущество, что он содержит сотни тысяч исследуемых генетических локусов, что дает более полную картину генетического состава. Родственные отношения могут быть определены только на статистической основе, потому что аутосомная ДНК подвергается рекомбинации. Одна хромосома может записывать историю каждого гена.Аутосомные исследования гораздо более надежны для демонстрации взаимоотношений между существующими популяциями, но не дают возможности раскрыть их историю так, как обещают исследования мтДНК и ДНК NRY, несмотря на их многочисленные сложности.
Генетические исследования основаны на многочисленных предположениях и страдают методологическими ограничениями, такими как критерий отбора и смешивающие явления, такие как генетический дрейф, эффекты основания и узкого места вызывают большие ошибки, особенно в исследованиях гаплогрупп. Независимо от того, насколько точна методология, выводы, сделанные в результате таких исследований, основываются на том, как автор рассматривает свои данные в соответствии с установленными археологическими или лингвистическими теориями.
Смотрите также
- Гаплогруппы Y-ДНК в популяциях Европы
- Археогенетика Ближнего Востока
- Генетическая история коренных народов Америки
- Генетика и археогенетика Южной Азии
- Археогенетика
- Генетическая изменчивость человека
Рекомендации
Встроенные цитаты
- ^ Paschou, Peristera; Ковачевич, Лейла; Тамбец, Кристина; Илумяэ, Анне-Май; Кушняревич Алена; Юнусбаев, Баязит; Сольник, Ану; Бего, Укротитель; Приморак, Драган; Скаро, Ведрана; Лесковац, Андрей; Яковски, Златко; Дробник, Катя; Толк, Хелле-Вийви; Ковачевич, Сандра; Рудан, Павао; Мецпалу, Эне; Марьянович, Дамир (2014). «Стоя у ворот в Европу - генетическая структура населения Западных Балкан, основанная на аутосомных и гаплоидных маркерах». PLOS ONE. 9 (8): e105090. Bibcode:2014PLoSO ... 9j5090K. Дои:10.1371 / journal.pone.0105090. ISSN 1932-6203. ЧВК 4141785. PMID 25148043.
- ^ Флейшер, Роберт С .; Нелис, Мари; Эско, Тыну; Мяги, Ридик; Зимприх, Фриц; Зимприч, Александр; Тончева, Драга; Карачанак, Сена; Пискачкова, Тереза; Балащак, Иван; Пелтонен, Лина; Яккула, Эвелиина; Ренстрём, Карола; Латроп, Марк; Хит, Саймон; Галан, Пилар; Шрайбер, Стефан; Мейтингер, Томас; Пфойфер, Арне; Wichmann, H-Erich; Мелег, Бела; Полгар, Ноэми; Тониоло, Даниэла; Гаспарини, Паоло; Д'Адамо, Пио; Кловинс, Янис; Никитина-Заке, Лиене; Кучинскас, Вайдутис; Каснаускене, Юрате; Любинский, Ян; Дебняк, Тадеуш; Лимборская, Светлана; Хрунин Андрей; Эстивиль, Ксавье; Rabionet, Raquel; Марсал, Сара; Хулиа, Антонио; Антонаракис, Стилианос Э .; Дойч, Самуэль; Борель, Кристель; Аттар, Хома; Гагнебин, Мэрилин; Мацек, Милан; Кравчак, Михаил; Ремм, Майдо; Мецпалу, Андрес (2009). «Генетическая структура европейцев: взгляд с северо-востока». PLOS ONE. 4 (5): e5472. Bibcode:2009PLoSO ... 4.5472N. Дои:10.1371 / journal.pone.0005472. ISSN 1932-6203. ЧВК 2675054. PMID 19424496.
- ^ а б Сегин-Орландо, Андайн; Korneliussen, Thorfinn S .; Сикора, Мартин; Маласпинас, Анна-Сапфо; Маника, Андреа; Мольтке, Ида; Альбрехтсен, Андерс; Ко, Эми; Маргарян, Ашот; Моисеев, Вячеслав; Гебель, Тед; Вестэвей, Майкл; Ламберт, Дэвид; Хартанович Валерий; Уолл, Джеффри Д .; Nigst, Philip R .; Фоли, Роберт А.; Лар, Марта Миразон; Нильсен, Расмус; Орландо, Людовик; Виллерслев, Эске (2014). «Геномная структура европейцев насчитывает не менее 36 200 лет». Наука. 346 (6213): 1113–1118. Bibcode:2014Научный ... 346.1113С. Дои:10.1126 / science.aaa0114. ISSN 0036-8075. PMID 25378462. S2CID 206632421.
- ^ Posth C, Renaud G, Mittnik M, Drucker DG, Rougier H, Cupillard C, Valentin F, Thevenet C, Furtwängler A, Wißing C, Francken M, Malina M, Bolus M, Lari M, Gigli E, Capecchi G, Crevecoeur I , Beauval C, Flas D, Germonpré M, van der Plicht J, Cottiaux R, Gély B, Ronchitelli A, Wehrberger K, Grigorescu D, Svoboda J, Semal P, Caramelli D, Bocherens H, Harvati K, Conard NJ, Haak W. , Пауэлл А., Краузе Дж. (2016). «Митохондриальные геномы плейстоцена предполагают единственное крупное расселение неафриканцев и позднеледниковую смену населения в Европе». Текущая биология. 26 (6): 827–833. Дои:10.1016 / j.cub.2016.01.037. HDL:2440/114930. PMID 26853362. S2CID 140098861.
- ^ Вай С., Сарно С., Лари М., Луизелли Д., Манци Дж., Галлинаро М., Матайч С., Хюбнер А., Моди А., Пилли Е., Тафури М. А., Карамелли Д., ди Лерния С. (март 2019 г.). «Митохондриальная линия предков N из неолитической« зеленой »Сахары». Научный представитель. 9 (1): 3530. Bibcode:2019НатСР ... 9,35 30В. Дои:10.1038 / s41598-019-39802-1. ЧВК 6401177. PMID 30837540.
- ^ Хабер М., Джонс А.Л., Коннел Б.А., Асан, Арчиеро Э., Хуанмин И., Томас М.Г., Сюэ Ю., Тайлер-Смит К. (июнь 2019 г.). «Редкая африканская гаплогруппа Y-хромосомы с глубокими корнями D0 и ее значение для распространения современного человека за пределы Африки». Генетика. 212 (4): 1421–1428. Дои:10.1534 / генетика.119.302368. ЧВК 6707464. PMID 31196864.
- ^ Вилланья и Шрайбнер (2019). «Множественные эпизоды скрещивания неандертальцев и современных людей». Природа Экология и эволюция. 3 (1): 39–44. Дои:10.1038 / с41559-018-0735-8. ЧВК 6309227. PMID 30478305.
- ^ а б Белеза, Сандра; Santos, António M .; Макэвой, Брайан; Алвес, Изабель; Мартиньо, Клаудиа; Кэмерон, Эмили; Шрайвер, Марк Д .; Парра, Эстебан Дж .; Роча, Хорхе (2013). «Время осветления пигментации у европейцев». Молекулярная биология и эволюция. 30 (1): 24–35. Дои:10.1093 / молбев / mss207. ЧВК 3525146. PMID 22923467.
- ^ Джонс, Эппи Р .; Гонсалес-Фортес, Глория; Коннелл, Сара; Сиска, Вероника; Эрикссон, Андерс; Мартиниано, Руи; McLaughlin, Russell L .; Гальего Льоренте, Маркос; Кэссиди, Лара М .; Гамба, Кристина; Мешвелиани, Тенгиз; Бар-Йосеф, Офер; Мюллер, Вернер; Белфер-Коэн, Анна; Мацкевич, Зиновий; Джакели, Нино; Higham, Thomas F. G .; Куррат, Матиас; Лордкипанидзе, Давид; Хофрайтер, Майкл; Маника, Андреа; Пинхаси, Рон; Брэдли, Дэниел Г. (2015). «Геномы верхнего палеолита раскрывают глубокие корни современных евразийцев». Nature Communications. 6 (1): 8912. Bibcode:2015 НатКо ... 6.8912J. Дои:10.1038 / ncomms9912. ISSN 2041-1723. ЧВК 4660371. PMID 26567969.
- ^ Замена населения в неолите, а затем и в бронзовом веке была почти завершена в Доисторическая Британия, мезолитическая популяция WHG составляет всего около 10% предков современного коренного британского населения.Олальде, Иньиго; Брейс, Селина; Аллентофт, Мортен Э .; Армит, Ян; Кристиансен, Кристиан; Бут, Томас; Роланд, Надин; Маллик, Свапан; Сеченьи-Надь, Анна; Миттник, Алисса; Альтена, Эвелин; Липсон, Марк; Лазаридис, Иосиф; Харпер, Томас К .; Паттерсон, Ник; Брумандхошбахт, Насрин; Дикманн, Йоан; Фалтыскова, Зузана; Фернандес, Даниэль; Ферри, Мэтью; Харни, Иадаоин; де Книжфф, Питер; Мишель, Меган; Оппенгеймер, Йонас; Стюардсон, Кристин; Барклай, Алистер; Альт, Курт Вернер; Лизау, Корина; Риос, Патрисия; и другие. (2018). «Феномен Стакана и геномная трансформация северо-западной Европы». Природа. 555 (7695): 190–196. Bibcode:2018Натура.555..190O. Дои:10.1038 / природа25738. ISSN 0028-0836. ЧВК 5973796. PMID 29466337.
- ^ Лазаридис, Иосиф; Паттерсон, Ник; Миттник, Алисса; Рено, Габриэль; Маллик, Свапан; Кирсанов, Карола; Судмант, Питер Х .; Schraiber, Joshua G .; Кастеллано, Серджи; Липсон, Марк; Бергер, Бонни; Эконому, Христос; Боллонгино, Рут; Фу, Цяомэй; Bos, Кирстен I .; Норденфельт, Сюзанна; Ли, Хэн; де Филиппо, Чезаре; Прюфер, Кей; Сойер, Сюзанна; Пост, Козимо; Хаак, Вольфганг; Халлгрен, Фредрик; Форнандер, Элин; Роланд, Надин; Делсате, Доминик; Франкен, Майкл; Гине, Жан-Мишель; Валь, Иоахим; и другие. (2014). «Древние геномы человека предполагают наличие трех предковых популяций современных европейцев». Природа. 513 (7518): 409–413. arXiv:1312.6639. Bibcode:2014Натура.513..409L. Дои:10.1038 / природа13673. ISSN 0028-0836. ЧВК 4170574. PMID 25230663.
- ^ Поскольку Lazaridis et al. (2014) дальнейшие исследования уточнили картину скрещивания между EEF и WHG. В результате анализа в 2017 году 180 древних наборов данных ДНК эпох энеолита и неолита из Венгрии, Германии и Испании были обнаружены доказательства длительного периода скрещивания EEF-WHG. Смешивание происходило на региональном уровне, из местных популяций охотников-собирателей, так что популяции из трех регионов (Германии, Иберии и Венгрии) были генетически различимы на всех этапах неолита, с постепенно увеличивающимся соотношением предков WHG сельскохозяйственных популяций с течением времени. . Это говорит о том, что после первоначальной экспансии ранних фермеров не было дальнейших миграций на большие расстояния, достаточно существенных для гомогенизации фермерского населения, и что фермерские хозяйства и популяции охотников-собирателей существовали бок о бок на протяжении многих столетий, с продолжающимся постепенным смешением на протяжении 5-го века до 4-го тысячелетия До н.э. (а не единичная примесь при первоначальном контакте).Липсон, Марк; Сеченьи-Надь, Анна; Маллик, Свапан; Поза, Аннамария; Стегмар, Балаж; Кирл, Виктория; Роланд, Надин; Стюардсон, Кристин; Ферри, Мэтью; Мишель, Меган; Оппенгеймер, Йонас; Брумандхошбахт, Насрин; Харни, Иадаоин; Норденфельт, Сюзанна; Ламы, Бастьен; Густав Менде, Балаж; Келер, Китти; Оросс, Кристиан; Бондар, Мария; Мартон, Тибор; Osztás, Anett; Якуч, Янош; Палуч, Тибор; Хорват, Ференц; Ченнери, Пироска; Коос, Юдит; Себек, Каталин; Андерс, Александра; Raczky, Pál; Регенье, Юдит; Барна, Юдит П .; Фабиан, Сильвия; Серлеги, Габор; Толди, Золтан; Дьёндживер Надь, Эмесе; Дани, Янош; Мольнар, Эрика; Палфи, Дьёрдь; Марк, Ласло; Мелег, Бела; Банфаи, Жолт; Домбороцки, Ласло; Фернандес-Эрасо, Хавьер; Антонио Мухика-Алустиса, Хосе; Алонсо Фернандес, Кармен; Хименес Эчеваррия, Хавьер; Боллонгино, Рут; Оршидт, Йорг; Ширхольд, Керстин; Меллер, Харальд; Купер, Алан; Бургер, Иоахим; Банфи, Эстер; Альт, Курт В .; Лалуэса-Фокс, Карлес; Хаак, Вольфганг; Райх, Дэвид (2017). «Параллельные палеогеномные трансекты раскрывают сложную генетическую историю ранних европейских фермеров». Природа. 551 (7680): 368–372. Bibcode:2017Натура.551..368L. Дои:10.1038 / природа24476. ISSN 0028-0836. ЧВК 5973800. PMID 29144465.
- ^ «Не существует такого понятия, как« чистый »европеец или кто-либо еще». Наука | AAAS. 15 мая 2017 года.
- ^ Эндрю Карри (2019). «Генетическое тестирование показывает, что Европа - это плавильный котел, созданный из иммигрантов». Национальная география.
- ^ Lipson et al. (2017), Рис 2.
- ^ Датчен, Стефани (23 ноября 2015 г.). «Фермерство в их ДНК». Гарвардская медицинская школа. Получено 25 ноября 2015.
- ^ а б c d е ж грамм час я j k л м Фу, Цяомэй; Пост, Козимо (2 мая 2016 г.). «Генетическая история ледникового периода Европы». Природа. 534 (7606): 200–05. Bibcode:2016Натура.534..200F. Дои:10.1038 / природа17993. ЧВК 4943878. PMID 27135931.
- ^ а б c Richards et al. 2000 г.
- ^ Еще до появления генетических исследований некоторые антропологи полагали, что они обнаружили скелеты, представляющие «гибриды» неандертальца и современного человека. Эти результаты были сочтены «неоднозначными». Археологические данные указывают на резкий переход от артефактов неандертальцев к артефактам, связанным с AMH, во время верхнего палеолита.Кляйн Р.Г. (март 2003 г.), «Палеоантропология. Куда неандертальцы?», Наука, 299 (5612): 1525–27, Дои:10.1126 / science.1082025, PMID 12624250, S2CID 161836323
- ^ Prüfer, K .; Racimo, F .; Patterson, N .; Джей, Ф .; Sankararaman, S .; Sawyer, S .; и другие. (2014) [Online 2013]. «Полная последовательность генома неандертальца с Горного Алтая». Природа. 505 (7481): 43–49. Bibcode:2014Натура.505 ... 43П. Дои:10.1038 / природа12886. ЧВК 4031459. PMID 24352235.
- ^ Дж. Харди; А. Питтман; А. Майерс; К. Гвинн-Харди; H.C. Гриб; Р. де Сильва; М. Хаттон; Дж. Дакворт (2005), «Свидетельства того, что Homo neanderthalensis внес гаплотип H2 MAPT в Homo sapiens», Сделки Биохимического Общества, 33 (4): 582–85, Дои:10.1042 / bst0330582, PMID 16042549,
Мы предполагаем, что гаплотип H2 произошел от Homo neanderthalensis и вошел в популяции H. sapiens во время сосуществования этих видов в Европе примерно с. 45 000–18 000 лет назад, и с тех пор гаплотип H2 находится под давлением отбора, возможно, из-за роли этого гаплотипа H1 в нейродегенеративном заболевании. «...» Локус тау (MAPT) очень необычен. На территории прибл. 1,8 Mb, в европейских популяциях есть две клады гаплотипов, H1 и H2 [6,7]. В других популяциях встречается только H1 и демонстрирует нормальный паттерн рекомбинации.
- ^ Шоу-Смит, К; Pittman, Am; Willatt, L; Мартин, H; Рикман, L; Гриббл, S; Curley, R; Камминг, S; Данн, К; Калайцопулос, D; Портер, К; Пригмор, Э; Крепищи-Сантос, AC; Варела, MC; Koiffmann, CP; Лис, AJ; Розенберг, К; Ферт, Hv; Де Сильва, Р. Картер, Н. П. (сентябрь 2006 г.), «Микроделеция, охватывающая MAPT на хромосоме 17q21.3, связана с задержкой развития и неспособностью к обучению», Природа Генетика, 38 (9): 1032–37, Дои:10,1038 / ng1858, ISSN 1061-4036, PMID 16906163, S2CID 38047848
- ^ Зоди, Мак; Цзян, З; Fung, Hc; Antonacci, F; Hillier, Lw; Cardone, Mf; Могилы, Та; Kidd, Jm; Cheng, Z; Abouelleil, A; Чен, L; Уоллис, Дж; Гласскок, Дж; Уилсон, РК; Рейли, Ад; Дакворт, Дж; Вентура, М; Харди, Дж; Уоррен, WC; Eichler, EE (август 2008 г.), "Эволюционное переключение области инверсии MAPT 17q21.31", Природа Генетика, 40 (9): 1076–83, Дои:10,1038 / нг.193, ISSN 1061-4036, ЧВК 2684794, PMID 19165922
- ^ Интрогрессия и микроцефалин FAQ Джон Хоукс [1]
- ^ Almos, Pz; Хорват, S; Czibula, A; Раско, я; Сипос, В; Bihari, P; Béres, J; Юхас, А; Янка, З; Калман, Дж. (Ноябрь 2008 г.), «Геномная изменчивость, связанная с гаплотипом тау H1 в 17q21.3, как азиатское наследие европейской цыганской популяции», Наследственность, 101 (5): 416–19, Дои:10.1038 / hdy.2008.70, ISSN 0018-067X, PMID 18648385,
В этом исследовании мы исследуем частоту инверсии 900 kb в 17q21.3 в цыганском и кавказском населении Венгрии, что может отражать азиатское происхождение популяций цыган. Считается, что из двух гаплотипов (H1 и H2) H2 имеет исключительно кавказское происхождение, и его появление в других расовых группах, вероятно, отражает примесь. В нашей выборке гаплотип H1 значительно чаще встречался в популяции цыган (89,8 против 75,5%, P <0,001) и находился в неравновесном состоянии Харди – Вайнберга (P = 0,017). Область 17q21.3 включает ген тау-белка, ассоциированного с микротрубочками, и этот результат может означать более высокую чувствительность к многофакторным таупатиям, связанным с гаплотипом H1, среди цыган.
- ^ Уэйд, Николас (2009-12-02), "Ученые Германии разрабатывают геном неандертальца", Нью-Йорк Таймс, Нью-Йорк Таймс, получено 2010-05-03
- ^ «Неандертальцы» отличные от нас'", BBC, 2009-12-02
- ^ Милисаускас (2002 г., п. 59)
- ^ а б c Семино 2000
- ^ Семино 2000.
- ^ Уэллс 2001. Евразийский центр.
- ^ Татьяна Михайловна Карафет; и другие. (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека». Genome Res. 18 (5): 830–38. Дои:10.1101 / gr.7172008. ЧВК 2336805. PMID 18385274.
- ^ а б c Ричардс и др. (2000)
- ^ Торрони А., Бандельт Х. Дж., Маколей В. и др. (Октябрь 2001 г.), «Сигнал от мтДНК человека о постледниковой реколонизации в Европе», Являюсь. J. Hum. Genet., 69 (4): 844–52, Дои:10.1086/323485, ЧВК 1226069, PMID 11517423
- ^ Пала, Мария; Ахилли, Алессандро; Оливьери, Анна; Кашани, Бахарак Хушиар; Перего, Уго А; Санна, Дарья; Мецпалу, Эне; Тамбец, Кристина; Тамм, Эрика; и другие. (2009), "Митохондриальная гаплогруппа U5b3: отдаленное эхо эпипалеолита в Италии и наследие ранних сардинцев", Американский журнал генетики человека, 84 (6): 814–21, Дои:10.1016 / j.ajhg.2009.05.004, ЧВК 2694970, PMID 19500771, получено 2014-12-29
- ^ R Wells et al. Евразийский хартленд: континентальный взгляд на разнообразие Y-хромосомы
- ^ Semino et al. (2000)
- ^ а б Рутси С., Магри С., Кивисилд Т., Бенуцци Г., Хелп Х., Бермишева М., Кутуев И., Барак Л., Перичич М. и др. (2004), «Филогеография гаплогруппы I Y-хромосомы выявляет отдельные области доисторического потока генов в Европе» (PDF), Американский журнал генетики человека, 75 (1): 128–37, Дои:10.1086/422196, ЧВК 1181996, PMID 15162323, заархивировано из оригинал (PDF) на 2009-06-24, получено 2009-07-04.
- ^ Pericic et al. 2005 г.
- ^ Cinnioglu et al. Раскопки гаплотипов Y-хромосомы в Анатолии. 2003
- ^ Перичич и др. (2005) Для обсуждения восточноевропейского распространения R1a1
- ^ Пассарино и др. (2001) Для скандинавских данных
- ^ Семино (2000) общее введение
- ^ Билтон Д. Т., Мирол П. М., Маскеретти С., Фредга К., Зима Дж., Сирл Дж. Б. (июль 1998 г.), «Средиземноморская Европа как район эндемизма мелких млекопитающих, а не источник послеледниковой колонизации на север», Труды Королевского общества B, 265 (1402): 1219–26, Дои:10.1098 / rspb.1998.0423, ЧВК 1689182, PMID 9699314
- ^ а б c Датчен, Стефани (2 мая 2016 г.). «История на льду». Гарвардская медицинская школа. Получено 11 мая 2016.
- ^ а б c d е Мэтисон, Иэн; Лазаридис, Иосиф (23 ноября 2015 г.). «Полногеномные модели отбора у 230 древних евразийцев». Природа. 528 (7583): 499–503. Bibcode:2015Натура.528..499М. Дои:10.1038 / природа16152. ЧВК 4918750. PMID 26595274.
- ^ Белеза, Сандра; Santos, António M .; Макэвой, Брайан; Алвес, Изабель; Мартиньо, Клаудиа; Кэмерон, Эмили; Шрайвер, Марк Д .; Парра, Эстебан Дж .; Роча, Хорхе (январь 2013 г.). «Время осветления пигментации у европейцев». Молекулярная биология и эволюция. 30 (1): 24–35. Дои:10.1093 / молбев / mss207. ЧВК 3525146. PMID 22923467.
- ^ Джонс, Эппи Р .; Гонсалес-Фортес, Глория; Коннелл, Сара; Сиска, Вероника; Эрикссон, Андерс; Мартиниано, Руи; McLaughlin, Russell L .; Гальего Льоренте, Маркос; Кэссиди, Лара М .; Гамба, Кристина; Мешвелиани, Тенгиз; Бар-Йосеф, Офер; Мюллер, Вернер; Белфер-Коэн, Анна; Мацкевич, Зиновий; Джакели, Нино; Higham, Thomas F. G .; Куррат, Матиас; Лордкипанидзе, Давид; Хофрайтер, Майкл; Маника, Андреа; Пинхаси, Рон; Брэдли, Дэниел Г. (16 ноября 2015 г.). «Геномы верхнего палеолита раскрывают глубокие корни современных евразийцев». Nature Communications. 6 (1): 8912. Bibcode:2015 НатКо ... 6.8912J. Дои:10.1038 / ncomms9912. ЧВК 4660371. PMID 26567969.
- ^ Bustamante, Carlos D .; Кукка, Франческо (8 мая 2014 г.). «Популяционный геномный анализ древних и современных геномов позволяет по-новому взглянуть на генетическое происхождение тирольского ледяного человека и генетическую структуру Европы». PLOS Genetics. 10 (5): e1004353. Дои:10.1371 / journal.pgen.1004353. ISSN 1553-7404. ЧВК 4014435. PMID 24809476.
- ^ Консорциум, Генографический; Купер, Алан (9 ноября 2010 г.). «Древняя ДНК европейских фермеров эпохи раннего неолита показывает их близость к Ближнему Востоку». PLOS Биология. 8 (11): e1000536. Дои:10.1371 / journal.pbio.1000536. ISSN 1545-7885. ЧВК 2976717. PMID 21085689.
- ^ Perlès C, Monthel G (2001) Ранний неолит в Греции: первые фермерские сообщества в Европе. Издательство Кембриджского университета, Кембридж.
- ^ Руннелс С. (2003) Истоки греческого неолита: личный взгляд, Аммерман и Бьяджи (редакторы 2003 г.).
- ^ Милисаускас (2002 г., с. 1143, 1150).
- ^ Zeder MA (2008), «Одомашнивание и раннее земледелие в Средиземноморском бассейне: происхождение, распространение и влияние», PNAS, 105 (33): 11597–604, Bibcode:2008PNAS..10511597Z, Дои:10.1073 / pnas.0801317105, ЧВК 2575338, PMID 18697943.
- ^ Милисаускас, 2002 г. и Генетики присоединились к дискуссии с исследованиями, касающимися генетических паттернов современных европейских популяций, поскольку они связаны с происхождением неолитических популяций.:146)
- ^ Пьяцца, Альберто; Cavalli-Sforza, L.L .; Меноцци, Паоло (1994), История и география генов человека, Принстон, Нью-Джерси: Издательство Принстонского университета, ISBN 978-0-691-08750-4
- ^ Semino 2000. Здесь кладка E-M35 обозначается как «Eu 4».
- ^ Россер и др. 2000 г.
- ^ Данные по Y-хромосоме показывают сигнал для отдельной миграции позднего плейстоцена из Африки в Европу через Синай, о чем свидетельствует распределение клонов гаплогруппы E3b, которое не проявляется в распределении гаплогрупп мтДНК.
- ^ а б Птица, Стивен (2007), «Гаплогруппа E3b1a2 как возможный индикатор заселения римской Британии солдатами балканского происхождения», Журнал генетической генеалогии, 3 (2)[ISBN отсутствует ]
- ^ Haak, W .; Lazaridis, I .; Patterson, N .; Rohland, N .; Маллик, S .; Llamas, B .; Brandt, G .; Nordenfelt, S .; Harney, E .; Стюардсон, К .; Fu, Q .; Mittnik, A .; Bánffy, E .; Эконому, C .; Francken, M .; Friederich, S .; Pena, R.G .; Hallgren, F .; Хартанович, В .; Хохлов, А .; Кунст, М .; Кузнецов, П .; Meller, H .; Мочалов, О .; Моисеев, В .; Nicklisch, N .; Pichler, S.L .; Risch, R .; Rojo Guerra, M.A .; и другие. (2015). «Массовая миграция из степи была источником индоевропейских языков в Европе». Природа. 522 (7555): 207–11. arXiv:1502.02783. Bibcode:2015Натура.522..207H. Дои:10.1038 / природа14317. ЧВК 5048219. PMID 25731166.
- ^ Каллавей, Юэн (12 февраля 2015 г.). «Европейские языки связаны с миграцией с востока». Природа. Дои:10.1038 / природа.2015.16919. S2CID 184180681.
- ^ Андерхилл, Питер А; Майрес, Натали М; Рутси, Сиири; Мецпалу, Майт; Животовский, Лев А; Кинг, Рой Дж; Лин, Алиса А; Чоу, Шерил-Эмилиан Т; Семино, Орнелла; и другие. (2009), «Разделение постледникового происхождения европейских и азиатских Y-хромосом в гаплогруппе R1a», Европейский журнал генетики человека, 18 (4): 479–84, Дои:10.1038 / ejhg.2009.194, ЧВК 2987245, PMID 19888303
- ^ Макс Ингман; Ульф Гилленстен (2007). «Недавняя генетическая связь между саамами и Волго-Уральским регионом России». Европейский журнал генетики человека. 15 (1): 115–20. Дои:10.1038 / sj.ejhg.5201712. PMID 16985502.
- ^ Калеви Виик. "Кто такие финны?" (PDF). Получено 2016-03-16. Цитировать журнал требует
| журнал =(помощь) - ^ Mirabal et al. 2009 г.
- ^ S Rootsi; и другие. (2006). «Северный путь против часовой стрелки гаплогруппы N Y-хромосомы из Юго-Восточной Азии в Европу». Европейский журнал генетики человека. 15 (2): 204–11. Дои:10.1038 / sj.ejhg.5201748. PMID 17149388., Дополнительная таблица 1
- ^ а б c Эппи Р. Джонс; Глория Гонсалес-Фортес; Сара Коннелл; Вероника Сиска; Андерс Эрикссон; Руи Мартиниано; Рассел Л. Маклафлин; Маркос Гальего Льоренте; Лара М. Кэссиди; Кристина Гамба; Тенгиз Мешвелиани; Офер Бар-Йосеф; Вернер Мюллер; Анна Бельфер-Коэн; Зиновий Мацкевич; Нино Джакели; Томас Ф.Г. Хайэм; Матиас Куррат; Давид Лордкипанидзе; Майкл Хофрайтер; Андреа Маника; Рон Пинхаси; Дэниел Г. Брэдли (2015). «Геномы верхнего палеолита раскрывают глубокие корни современных евразийцев». Nature Communications. 6 (8912): 8912. Bibcode:2015 НатКо ... 6.8912J. Дои:10.1038 / ncomms9912. ЧВК 4660371. PMID 26567969.
- ^ Датчен, Стефани (17 сентября 2014 г.). «В европейское семейное древо добавлена новая ветвь». Гарвардская медицинская школа. Архивировано из оригинал на 2014-10-01. Получено 25 ноября 2015.
- ^ "Раскрыта тайна исконного" племени "". Новости BBC. 16 ноября 2015.
- ^ Иосиф Лазаридис; и другие. (2016). «Генетическая структура первых фермеров мира». bioRxiv 10.1101/059311.
- ^ Иэн Мэтисон; и другие. (2017). «Геномная история Юго-Восточной Европы». bioRxiv 10.1101/135616.
- ^ Басби и др., Текущая биология Том 25, выпуск 19, стр. 2518–26, 5 октября 2015 г.
- ^ "Паннония и Верхняя Мезия. История Среднего Дуная Провинции Римской Империи. Андраш Мочи. Лондон и Бостон, Рутледж и Кеган Пол. ISBN 0-7100-7714-9
- ^ Comas, D .; Schmid, H .; Braeuer, S .; Flaiz, C .; Бускетс, А .; Calafell, F .; Bertranpetit, J .; Scheil, H.-G .; Huckenbeck, W .; Ефремовская, Л .; Шмидт, Х. (март 2004 г.). «Полиморфизмы вставки Alu на Балканах и происхождение аромун». Анналы генетики человека. 68 (2): 120–127. Дои:10.1046 / j.1529-8817.2003.00080.x. PMID 15008791. S2CID 21773796.
- ^ Бош Э., Калафель Ф., Гонсалес-Нейра А. и др. (Июль 2006 г.), «Отцовские и материнские линии на Балканах демонстрируют однородный ландшафт через языковые барьеры, за исключением изолированных аромунов», Анналы генетики человека, 70 (Pt 4): 459–87, Дои:10.1111 / j.1469-1809.2005.00251.x, PMID 16759179, S2CID 23156886
- ^ Capelli et al. (2003).
- ^ Семино О. и др. (Ноябрь 2000 г.), «Генетическое наследие палеолита Homo sapiens sapiens у современных европейцев: перспектива Y-хромосомы», Наука, 290 (5494): 1155–59, Bibcode:2000Sci ... 290.1155S, Дои:10.1126 / наука.290.5494.1155, PMID 11073453 Примечание. Имена гаплогрупп в этой статье отличаются. Например: Гаплогруппа I обозначается как M170
- ^ Мирабал, Шейла; Варлен, Татьяна; Гайден, Тензин; Регейро, Мария; Вуйович, Славица; Попович, Даница; Джурич, Мария; Стойкович, Оливер; Эррера, Рене Дж. (2010). «Короткие тандемные повторы Y-хромосомы человека: рассказ об аккультурации и миграции как механизмах распространения сельского хозяйства на Балканском полуострове». Американский журнал физической антропологии. 142 (3): 380–90. Дои:10.1002 / ajpa.21235. PMID 20091845.
- ^ Перичич, Марияна; Лаук, Ловорка Барак; Кларич, Ирена Мартинович; Рутси, Сиири; Яничиевич, Бранка; Рудан, Игорь; Терзич, Рифет; Чолак, Иванка; Квесич, Анте; Попович, Дан; Шиячки, Ана; Бехлули, Ибрагим; Джордевич, Добривое; Ефремовская Людмила; Bajec, orđe D .; Стефанович, Бранислав Д .; Виллемс, Ричард; Рудан, Павао (октябрь 2005 г.). "Филогенетический анализ с высоким разрешением в Юго-Восточной Европе отслеживает основные эпизоды отцовского потока генов среди славянских народов". Молекулярная биология и эволюция. 22 (10): 1964–1975. Дои:10.1093 / molbev / msi185. PMID 15944443.
- ^ Батталья, Винченца; Форнарино, Симона; Аль-Захери, Надия; Оливьери, Анна; Пала, Мария; Майрес, Натали М; Кинг, Рой Дж; Рутси, Сиири; Марьянович, Дамир; и другие. (2008), «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе», Европейский журнал генетики человека, 17 (6): 820–30, Дои:10.1038 / ejhg.2008.249, ЧВК 2947100, PMID 19107149
- ^ Кручиани, Фульвио; Ла Фратта, Роберта; Сантоламацца, Пьеро; Селлитто, Даниэле; Пасконе, Роберто; Мораль, Педро; Уотсон, Элизабет; Гуида, Валентина; Коломб, Элиан Беро; Захарова, Бориана; Лавинья, Жуан; Вона, Джузеппе; Аман, Рашид; Кали, Франческо; Акар, Неджат; Ричардс, Мартин; Торрони, Антонио; Новеллетто, Андреа; Скоццари, Розария (май 2004 г.). «Филогеографический анализ хромосом гаплогруппы E3b (E-M215) Y выявил множественные миграционные события в Африке и за ее пределами». Американский журнал генетики человека. 74 (5): 1014–1022. Дои:10.1086/386294. ЧВК 1181964. PMID 15042509.
- ^ Балареска, Патрисия; Bowden, Georgina R .; Адамс, Сьюзан М .; Люн, Хо-Йи; Король, Тури Э .; Россер, Зои Х .; Гудвин, Джейн; Мойзан, Жан-Поль; Ричард, Кристель; Миллуорд, Энн; Demaine, Andrew G .; Барбуджани, Гвидо; Превидере, Карло; Уилсон, Ян Дж .; Тайлер-Смит, Крис; Джоблинг, Марк А .; Пенни, Дэвид (19 января 2010 г.). "Преимущественно неолитическое происхождение европейских отцовских линий". PLOS Биология. 8 (1): e1000285. Дои:10.1371 / journal.pbio.1000285. ЧВК 2799514. PMID 20087410.
- ^ а б Майрес, Натали; Рутси, Сиири; Лин, Алиса А; Ярве, Мари; Кинг, Рой Дж; Кутуев, Ильдус; Кабрера, Висенте М; Хуснутдинова, Эльза К; Пшеничнов Андрей; Юнусбаев, Баязит; Балановский, Олег; Балановская, Елена; Рудан, Павао; Балдович, Мариан; Эррера, Рене Дж; Кьярони, Жак; Ди Кристофаро, Джули; Виллемс, Ричард; Кивисилд, Тоомас; Андерхилл, Питер А. (2010), «Главный эффект голоцена гаплогруппы R1b Y-хромосомы в Центральной и Западной Европе», Европейский журнал генетики человека, 19 (1): 95–101, Дои:10.1038 / ejhg.2010.146, ЧВК 3039512, PMID 20736979
- ^ а б Кручиани, Фульвио; Тромбетта, Бениамино; Антонелли, Шайенн; Пасконе, Роберто; Валезини, Гвидо; Скальци, Валентина; Вона, Джузеппе; Мелег, Бела; Заградисник, Борис; и другие. (2010), «Сильная внутри- и межконтинентальная дифференциация, выявленная SNP M269, U106 и U152 Y-хромосомы», Forensic Science International: генетика, 5 (3): e49 – e52, Дои:10.1016 / j.fsigen.2010.07.006, PMID 20732840
- ^ Морелли; Contu, D; Santoni, F; Whalen, M. B .; Francalacci, P; Кукка, Ф. (2010), «Сравнение вариации Y-хромосомы в Сардинии и Анатолии больше соответствует культурному, а не демическому распространению сельского хозяйства», PLOS ONE, 5 (4): e10419, Bibcode:2010PLoSO ... 510419M, Дои:10.1371 / journal.pone.0010419, ЧВК 2861676, PMID 20454687
- ^ а б Кайзер, М; Лао, О; Анслингер, К; Огюстен, C; Баргель, G; Эдельманн, Дж; Элиас, S; Генрих, М; Хенке, Дж; и другие. (2005), «Значительная генетическая дифференциация между Польшей и Германией следует современным политическим границам, как показал анализ Y-хромосомы», Генетика человека, 117 (5): 428–43, Дои:10.1007 / s00439-005-1333-9, PMID 15959808, S2CID 11066186 Копию можно найти здесь «Значительная генетическая дифференциация между Польшей и Германией следует за современными политическими границами, как показал анализ Y-хромосомы» (PDF). Архивировано из оригинал (PDF) на 2009-03-04. Получено 2008-11-02..
- ^ Pericic et al. (2005)
- ^ Bowden et al. (2008)
- ^ Dupuy et al. (2005)
- ^ Kasperaviciūte et al. (2005)
- ^ Semino et al. (2004)
- ^ а б c d е Россер и др. (2000)
- ^ а б Деренко, Мирослава; Малярчук, Борис; Денисова, Галина; Перкова, Мария; Рогалла, Уршула; Гжибовски, Томаш; Хуснутдинова Эльза; Дамбуева Ирина; Захаров, Илья (2012). «Полный анализ митохондриальной ДНК гаплогрупп Восточной Евразии, редко встречающихся в популяциях Северной Азии и Восточной Европы». PLOS ONE. 7 (2): e32179. Bibcode:2012PLoSO ... 732179D. Дои:10.1371 / journal.pone.0032179. ЧВК 3283723. PMID 22363811.
- ^ Сересо, Мария; Ахилли, Алессандро; Olivieri, A .; Perego, U.A .; Gomez-Carballa, A .; Brisighelli, F .; Lancioni, H .; Woodward, S.R .; Лопес-Сото, М .; Карраседо, А .; Capelli, C .; Torroni, A .; Салас, А. (2012). «Восстановление древних связей митохондриальной ДНК между Африкой и Европой». Геномные исследования. 22 (5): 821–26. Дои:10.1101 / гр.134452.111. ЧВК 3337428. PMID 22454235.
- ^ Кавалли-Сфорца (1993)
- ^ Лао и др. (2008)
- ^ Кавалли-Сфорца (1993 г., стр. 291–96).
- ^ Кавалли-Сфорца, Лука; Меноцци, Паоло; Пьяцца, Альберто (1994). История и география генов человека. Princeton University Press, стр. 272
- ^ Кавалли-Сфорца (1993 г., п. 268)
- ^ Генетическая структура европейцев: взгляд с северо-востока, Нелис и др. 2009 г.
- ^ «Парный Fst между европейскими образцами». Дои:10.1371 / journal.pone.0005472.s003. Цитировать журнал требует
| журнал =(помощь) - ^ Тиан, С; Косой, Р; Nassir, R; и другие. (2009). «Генетическая субструктура европейской популяции: дальнейшее определение информативных маркеров происхождения для различения различных европейских этнических групп». Мол. Med. 15 (11–12): 371–83. Дои:10.2119 / молмед.2009.00094. ЧВК 2730349. PMID 19707526.
- ^ Тиан, С; Косой, Р; Nassir, R; и другие. (2009). «Генетическая субструктура европейской популяции: дальнейшее определение информативных маркеров происхождения для различения различных европейских этнических групп». Мол. Med. 15 (11–12): 371–83. Дои:10.2119 / молмед.2009.00094. ЧВК 2730349. PMID 19707526.
- ^ Селдин, Майкл Ф .; Шигета, Рассел; Виллослада, Пабло; Селми, Карло; Туомилехто, Яакко; Сильва, Габриэль; Бельмонт, Джон В .; Клареског, Ларс; Грегерсен, Питер К. (2006). «Европейская субструктура населения: кластеризация северных и южных популяций». PLOS Genetics. 2 (9): e143. Дои:10.1371 / journal.pgen.0020143. ЧВК 1564423. PMID 17044734.
- ^ а б c Бауше, Марк; Макэвой, Брайан; Пирсон, Лорел Н .; Quillen, Ellen E .; Саркисян, Тамара; Оганесян, Кристине; Дека, Ранджан; Брэдли, Дэниел Дж .; Шрайвер, Марк Д. (май 2007 г.). «Измерение стратификации населения Европы с помощью данных генотипа микрочипов». Американский журнал генетики человека. 80 (5): 948–956. Дои:10.1086/513477. ЧВК 1852743. PMID 17436249.
- ^ Лаос 2008
- ^ Novembre, J; и другие. (Ноябрь 2008 г.), «Гены отражают географию в Европе», Природа, 456 (7218): 98–101, Bibcode:2008Натура.456 ... 98N, Дои:10.1038 / природа07331, ЧВК 2735096, PMID 18758442
- ^ Лао, Оскар; Лу, Тимоти Т .; Нотнагель, Майкл; Юнге, Олаф; Фрейтаг-Вольф, Сандра; Калибе, Амке; Баласкакова, Мирослава; Бертранпетит, Жауме; Биндофф, Лоуренс А .; Комас, Дэвид; Холмлунд, Гунилла; Куваци Анастасия; Мацек, Милан; Молле, Изабель; Парсон, Вальтер; Пало, Юкка; Плоски, Рафаль; Саджантила, Антти; Тальабраччи, Адриано; Гетер, Ульрик; Верге, Томас; Риваденейра, Фернандо; Хофман, Альберт; Uitterlinden, André G .; Гигер, Кристиан; Вихман, Хайнц-Эрих; Рютер, Андреас; Шрайбер, Стефан; Беккер, Кристиан; Нюрнберг, Питер; Нельсон, Мэтью Р .; Кравчак, Михаил; Кайзер, Манфред (август 2008 г.). «Соотношение генетической и географической структуры в Европе». Текущая биология. 18 (16): 1241–1248. Дои:10.1016 / j.cub.2008.07.049. PMID 18691889. S2CID 16945780.
- ^ Хрунин, Андрей В .; Хохрин, Денис В .; Филиппова Ирина Н .; Эско, Тыну; Нелис, Мари; Бебякова Наталья А .; Болотова Наталья Л .; Кловинс, Янис; Никитина-Заке, Лиене; Ренстрём, Карола; Рипатти, Самули; Шрайбер, Стефан; Франке, Андре; Мацек, Милан; Крулишова, Вероника; Любинский, Ян; Мецпалу, Андрес; Лимборская, Светлана А. (7 марта 2013 г.). «Полногеномный анализ популяций европейской части России выявил новый полюс генетического разнообразия в Северной Европе». PLOS ONE. 8 (3): e58552. Bibcode:2013PLoSO ... 858552K. Дои:10.1371 / journal.pone.0058552. ЧВК 3591355. PMID 23505534.
- ^ Тиан, С; Косой, Р; Nassir, R; и другие. (2009). "PubMed Central, Таблица 1: Мол Мед". Мол. Med. 15 (11–12): 371–83. Дои:10.2119 / молмед.2009.00094. ЧВК 2730349. PMID 19707526.
- ^ Нелис, Мари; Эско, Тыну; Мяги, Ридик; Зимприх, Фриц; Зимприч, Александр; Тончева, Драга; Карачанак, Сена; Пискачкова, Тереза; Балащак, Иван; Пелтонен, Лина; Яккула, Эвелиина; Ренстрём, Карола; Латроп, Марк; Хит, Саймон; Галан, Пилар; Шрайбер, Стефан; Мейтингер, Томас; Пфойфер, Арне; Wichmann, H-Erich; Мелег, Бела; Полгар, Ноэми; Тониоло, Даниэла; Гаспарини, Паоло; Д'Адамо, Пио; Кловинс, Янис; Никитина-Заке, Лиене; Кучинскас, Вайдутис; Каснаускене, Юрате; Любинский, Ян; Дебняк, Тадеуш; Лимборская, Светлана; Хрунин Андрей; Эстивиль, Ксавье; Rabionet, Raquel; Марсал, Сара; Хулиа, Антонио; Антонаракис, Стилианос Э .; Дойч, Самуэль; Борель, Кристель; Аттар, Хома; Гагнебин, Мэрилин; Мацек, Милан; Кравчак, Михаил; Ремм, Майдо; Мецпалу, Андрес; Флейшер, Роберт К. (8 мая 2009 г.). «Генетическая структура европейцев: взгляд с северо-востока». PLOS ONE. 4 (5): e5472. Bibcode:2009PLoSO ... 4.5472N. Дои:10.1371 / journal.pone.0005472. ЧВК 2675054. PMID 19424496.
- ^ Нелис, М; Эско, Т; Mägi, R; Зимприч, Ф; Зимприч, А; Тончева Д; Карачанак, С; Piskácková, T; Баласкак, I; Пелтонен, L; Jakkula, E; Ренстрём, К; Латроп, М; Хит, S; Галан, П; Шрайбер, S; Мейтингер, Т; Пфойфер, А; Wichmann, H.E .; Мелег, Б; Polgár, N; Тониоло, D; Гаспарини, П; д'Адамо, П; Кловинс, Дж; Никитина-Заке, Л; Кучинскас, В; Kasnauskiene, J; Любинский, Дж; и другие. (2009), "Генетическая субструктура европейского населения: дальнейшее определение информативных маркеров происхождения для различения различных европейских этнических групп", PLOS ONE, 4 (5): e5472, таблица 2, Bibcode:2009PLoSO ... 4.5472N, Дои:10.1371 / journal.pone.0005472, ЧВК 2675054, PMID 19424496
- ^ а б Кавалли-Сфорца (1993 г., п. 51)
- ^ Кавалли-Сфорца (1993 г., п. 39)
- ^ Арреди, Полони и Тайлер-Смит (2007), п. 390)
- ^ Милисаускас (2002 г., п. 58)
- ^ Семино (2000)
- ^ Барбужани и Берторель (2001):22–25)
Ссылки на источники
- Адамс, Сьюзан М .; Босх, Елена; Balaresque, Patricia L .; Ballereau, StéPhane J .; Ли, Эндрю С .; Арройо, Эдуардо; Лопес-Парра, Ана М .; Aler, Mercedes; Грифо, Марина С. Гисберт; и другие. (2008), «Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, евреев и мусульман на Пиренейском полуострове», Американский журнал генетики человека, 83 (6): 725–36, Дои:10.1016 / j.ajhg.2008.11.007, ЧВК 2668061, PMID 19061982, получено 2009-07-22
- Арреди Б., Полони Э.С., Параккини С. и др. (Август 2004 г.), «Преимущественно неолитическое происхождение вариации ДНК Y-хромосомы в Северной Африке», Американский журнал генетики человека, 75 (2): 338–45, Дои:10.1086/423147, ЧВК 1216069, PMID 15202071
- Арреди, Б; Poloni, E; Тайлер-Смит, К. (2007), «Население Европы», Антропологическая генетика: теория, методы и приложения, Издательство Кембриджского университета, ISBN 978-0-521-54697-3
- Auton, А; Bryc, K; Бойко А.Р .; Lohmueller, KE; Novembre, J; Рейнольдс, А; Индап, А; Райт, MH; Дегенхардт, JD; и другие. (2009), «Глобальное распределение геномного разнообразия подчеркивает богатую сложную историю континентальных популяций людей», Геномные исследования, 19 (5): 795–803, Дои:10.1101 / гр.088898.108, ЧВК 2675968, PMID 19218534
- Барбуджани, Гвидо; Берторелле, Джорджио (2001), "Генетика и история населения Европы", PNAS, 98 (1): 22–25, Bibcode:2001PNAS ... 98 ... 22B, Дои:10.1073 / pnas.98.1.22, ЧВК 33353, PMID 11136246
- Батталья, Винченца; Форнарино, Симона; Аль-Захери, Надия; Оливьери, Анна; Пала, Мария; Майрес, Натали М; Кинг, Рой Дж; Рутси, Сиири; Марьянович, Дамир; и другие. (2008), «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе», Европейский журнал генетики человека, 17 (6): 820–30, Дои:10.1038 / ejhg.2008.249, ЧВК 2947100, PMID 19107149
- Бауше, Марк; Макэвой, Брайан; Pearson, L.N .; Quillen, E.E .; Саркисян, Т; Оганесян, К; Дека, Р; Bradley, D.G .; Шрайвер, доктор медицины (2007), "Измерение стратификации населения Европы с использованием данных генотипа микрочипов", Американский журнал генетики человека, 80 (5): 948–56, Дои:10.1086/513477, ЧВК 1852743, PMID 17436249
- Белеза, С; Gusmão, L; Лопес, А; Алвес, С; Gomes, I; Giouzeli, M; Калафель, Франция; Карраседо, А; Аморим, А (2005), "Микрофилогеографическая и демографическая история португальских мужских линий", Анналы генетики человека, 70 (Pt 2): 181–94, Дои:10.1111 / j.1529-8817.2005.00221.x, PMID 16626329, S2CID 4652154[мертвая ссылка ]
- Bosch E, Calafell F, Comas D, Oefner PJ, Underhill PA, Bertranpetit J (апрель 2001 г.), «Анализ вариаций Y-хромосомы человека с высоким разрешением показывает резкий разрыв и ограниченный поток генов между северо-западом Африки и Пиренейским полуостровом», Американский журнал генетики человека, 68 (4): 1019–29, Дои:10.1086/319521, ЧВК 1275654, PMID 11254456
- Brace, C.L .; и другие. (2005), «Сомнительный вклад неолита и бронзового века в европейскую черепно-лицевую форму», Труды Национальной академии наук, 103 (1): 242–47, Bibcode:2006ПНАС..103..242Б, Дои:10.1073 / pnas.0509801102, ЧВК 1325007, PMID 16371462
- Капелли, C; Рыжая, N; Абернети, Дж. К.; Gratrix, F; Wilson, JF; Moen, T; Хервиг, Т; Ричардс, М; Штумпф, депутат; и другие. (2003), «Перепись хромосомы Y на Британских островах», Текущая биология, 13 (11): 979–84, Дои:10.1016 / S0960-9822 (03) 00373-7, PMID 12781138 также в "030705U491" (PDF). Получено 2011-06-01. Цитировать журнал требует
| журнал =(помощь) - Капелли, Кристиан; Онофри, Валерио; Бризигелли, Франческа; Боски, Илария; Скарниччи, Франческа; Масулло, Мара; Ферри, Джанмарко; Тофанелли, Серджио; Тальабраччи, Адриано; и другие. (2009), «Мавры и сарацины в Европе: оценка средневекового наследия мужчин Северной Африки в южной Европе», Европейский журнал генетики человека, 17 (6): 848–52, Дои:10.1038 / ejhg.2008.258, ЧВК 2947089, PMID 19156170
- Кавалли-Сфорца, Луиджи Л.; Меноцци, Паоло; Пьяцца, Альберто (1993), История и география генов человека, Princeton University Press, ISBN 978-0-691-08750-4, получено 2009-07-22
- Кавалли-Сфорца, Луиджи Л. (1997), «Гены, народы и языки», PNAS, 94 (15): 7719–24, Bibcode:1997PNAS ... 94.7719C, Дои:10.1073 / пнас.94.15.7719, ЧВК 33682, PMID 9223254
- Cruciani, F; Santolamazza, P; Шен, П; Маколей, V; Мораль, P; Olckers, A; Модиано, D; Холмс, S; Дестро-Бизоль, Г; и другие. (2002), "Обратная миграция из Азии в Африку к югу от Сахары подтверждается анализом гаплотипов Y-хромосомы человека с высоким разрешением", Американский журнал генетики человека, 70 (5): 1197–214, Дои:10.1086/340257, ЧВК 447595, PMID 11910562
- Cruciani F, La Fratta R, Santolamazza P и др. (Май 2004 г.), «Филогеографический анализ хромосом гаплогруппы E3b (E-M215) y выявляет множественные миграционные события в Африке и за ее пределами», Американский журнал генетики человека, 74 (5): 1014–22, Дои:10.1086/386294, ЧВК 1181964, PMID 15042509
- Кручиани, Фульвио; и другие. (2006), «Молекулярное рассечение гаплогруппы Y-хромосомы E-M78 (E3b1a): апостериорная оценка подхода на основе микросателлитных сетей с помощью шести новых двуаллельных маркеров», Человеческая мутация, 27 (8): 831–32, Дои:10.1002 / humu.9445, PMID 16835895, S2CID 26886757
- Cruciani, F; Ла Фратта, Р; Тромбетта, B; Santolamazza, P; Селлитто, Д; Colomb, EB; Dugoujon, JM; Crivellaro, F; Benincasa, T; и другие. (2007), «Отслеживание прошлых движений мужчин в Северной / Восточной Африке и Западной Евразии: новые ключи к разгадке на основе гаплогрупп Y-хромосомы E-M78 и J-M12», Молекулярная биология и эволюция, 24 (6): 1300–11, Дои:10.1093 / молбев / msm049, PMID 17351267, получено 2009-07-22 Также см Дополнительные данные.
- Ди Гаэтано; и другие. (2008), «Дифференциальные миграции греков и северной Африки на Сицилию подтверждаются генетическими данными по Y-хромосоме», Европейский журнал генетики человека, 17 (1): 91–99, Дои:10.1038 / ejhg.2008.120, ЧВК 2985948, PMID 18685561
- Флорес, Карлос; Maca-Meyer, N; Гонсалес, AM; Oefner, PJ; Шен, П; Pérez, JA; Рохас, А; Ларруга, JM; Андерхилл, Пенсильвания (2004), «Уменьшение генетической структуры Пиренейского полуострова, выявленное анализом Y-хромосомы: последствия для демографии населения» (PDF), Европейский журнал генетики человека, 12 (10): 855–63, Дои:10.1038 / sj.ejhg.5201225, PMID 15280900, S2CID 16765118, заархивировано из оригинал (PDF) на 2008-04-06, получено 2009-07-22
- Francalacci, P .; Morelli, L .; Андерхилл, П.А.; Lillie, A.S .; Пассарино, G .; Усели, А .; Madeddu, R .; Paoli, G .; Tofanelli, S .; и другие. (2003), «Население трех средиземноморских островов (Корсика, Сардиния и Сицилия), обусловленное двуаллельной изменчивостью Y-хромосомы», Американский журнал физической антропологии, 121 (3): 270–79, Дои:10.1002 / ajpa.10265, PMID 12772214
- Гонсалвес, Р. Фрейтас, А; Бранко, М; Роза, А; Fernandes, AT; Животовский, Л.А.; Андерхилл, Пенсильвания; Кивисилд, Т; Брем, А (2005), «Происхождение Y-хромосомы из Португалии, Мадейры и Акорес, записывающих элементы сефардского и берберского происхождения», Анналы генетики человека, 69 (Pt 4): 443–54, Дои:10.1111 / j.1529-8817.2005.00161.x, PMID 15996172, S2CID 3229760[мертвая ссылка ]
- Гальдер, я; и другие. (2007), «Панель информативных маркеров происхождения для оценки индивидуального биогеографического происхождения и примесей с четырех континентов: полезность и применение», Человеческая мутация, 29 (5): 648–58, Дои:10.1002 / humu.20695, PMID 18286470, S2CID 12489449, заархивировано из оригинал на 2012-10-16
- Кинг и Андерхилл (2002), «Конгруэнтное распределение неолитической расписной керамики и керамических фигурок с линиями передачи Y-хромосомы», Античность, 76 (293): 707–14, Дои:10.1017 / с0003598x00091158
- Лао, О; Лу, ТТ; Нотнагель, М; Юнге, О; Freitag-Wolf, S; Калибе, А; Баласкакова, М; Bertranpetit, J; Биндофф, Луизиана; и другие. (Август 2008 г.), «Корреляция между генетической и географической структурой в Европе», Текущая биология, 18 (16): 1241–48, Дои:10.1016 / j.cub.2008.07.049, PMID 18691889, S2CID 16945780
- Пассарино, Джузеппе; и другие. (2001), «Гаплотип 11 49a, f является новым маркером линии происхождения EU19, который отслеживает миграции из северных регионов Черного моря», Гм. Иммунол., 62 (9), стр. 922–32, Дои:10.1016 / S0198-8859 (01) 00291-9, PMID 11543894.
- Pericic, M; Lauc, LB; Кларич, ИМ; Рутси, S; Яничевич, B; Рудан, я; Terzić, R; Чолак, I; Квесич, А; и другие. (Октябрь 2005 г.), «Филогенетический анализ с высоким разрешением в юго-восточной Европе отслеживает основные эпизоды отцовского потока генов среди славянского населения», Мол. Биол. Evol., 22 (10): 1964–75, Дои:10.1093 / molbev / msi185, PMID 15944443, получено 2009-07-22
- Милисаускас, Шарунас (2002), Европейская предыстория: обзор, Birkhauser, ISBN 978-0-306-46793-6
- Ricaut, F.X .; и другие. (2008), "Дискретные черепные особенности византийского населения и перемещения населения Восточного Средиземноморья", Человеческая биология, 80 (5): 535–64, Дои:10.3378/1534-6617-80.5.535, PMID 19341322, S2CID 25142338
- Ричардс, М; Маколей, V; Hickey, E; Vega, E; Сайкс, Б; Guida, V; Ренго, C; Селлитто, Д; Cruciani, F; и другие. (Ноябрь 2000 г.), "Отслеживание родословных европейских основателей в ближневосточном пуле мтДНК" Американский журнал генетики человека, 67 (5): 1251–76, Дои:10.1016 / S0002-9297 (07) 62954-1, ЧВК 1288566, PMID 11032788
- Россер, ZH; Зерджал, Т; Hurles, ME; Адохаан, М; Алавантик, Д; Аморим, А; Амос, Вт; Арментерос, М; Арройо, Э; и другие. (2000), «Разнообразие Y-хромосомы в Европе является клинальным и определяется в первую очередь географией, а не языком», Американский журнал генетики человека, 67 (6): 1526–43, Дои:10.1086/316890, ЧВК 1287948, PMID 11078479, заархивировано из оригинал на 2008-05-06
- Скоццари, Розария; Cruciani, F; Панграцио, А; Santolamazza, P; Вона, G; Мораль, P; Латини, V; Варези, L; Мемми, ММ; и другие. (2001), "Изменения Y-хромосомы человека в районе Западного Средиземноморья: последствия для населения региона" (PDF), Иммунология человека, 62 (9): 871–84, CiteSeerX 10.1.1.408.4857, Дои:10.1016 / S0198-8859 (01) 00286-5, PMID 11543889, получено 2009-07-22
- Семино, Орнелла; Пассарино, G; Oefner, PJ; Линь А.А.; Арбузова, С; Бекман, LE; Де Бенедиктис, G; Francalacci, P; Куваци, А; и другие. (Ноябрь 2000 г.), «Генетическое наследие палеолита Homo sapiens sapiens у современных европейцев: перспектива Y-хромосомы» (PDF), Наука, 290 (5494): 1155–59, Bibcode:2000Sci ... 290.1155S, Дои:10.1126 / наука.290.5494.1155, PMID 11073453, заархивировано из оригинал (PDF) на 2008-03-07, получено 2009-07-22.
- Семино, О; Santachiarabenerecetti, A; Фаласки, Ф; Cavallisforza, L; Андерхилл, П. (2002), «Эфиопы и койсаны разделяют самые глубокие клады филогении Y-хромосомы человека» (PDF), Am J Hum Genet, 70 (1), стр. 265–68, Дои:10.1086/338306, ЧВК 384897, PMID 11719903, заархивировано из оригинал (PDF) на 2006-03-15, получено 2009-07-22
- Семино О, Магри С., Бенуцци Г. и др. (Май 2004 г.), «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в районе Средиземноморья», Американский журнал генетики человека, 74 (5): 1023–34, Дои:10.1086/386295, ЧВК 1181965, PMID 15069642
- Сайкс, Брайан (2006), Кровь островов: исследование генетических корней нашей племенной истории, Бантам, ISBN 978-0-593-05652-3, получено 2009-07-22
- Андерхилл, Питер А .; Шен, Пейдун; Лин, Алиса А .; Джин, Ли; Пассарино, Джузеппе; Ян, Вэй Х .; Кауфман, Эрин; Бонне-Тамир, Батшева; Бертранпетит, Жауме; и другие. (2000), "Вариации последовательности Y-хромосомы и история человеческих популяций", Нат Жене, 26 (3), стр. 358–61, Дои:10.1038/81685, PMID 11062480, S2CID 12893406
- Андерхилл, П.А.; Пассарино, G .; Lin, A.A .; Shen, P .; Mirazon Lahr, M .; Foley, R.A .; Oefner, P.J .; Кавалли-Сфорца, Л.Л. (2001), «Филогеография бинарных гаплотипов Y-хромосомы и происхождение современных человеческих популяций» (PDF), Энн Хам Жене, 65 (Pt 1), pp. 43–62, Дои:10.1046 / j.1469-1809.2001.6510043.x, PMID 11415522, S2CID 9441236, заархивировано из оригинал (PDF) на 2011-07-21, получено 2009-07-22
- Андерхилл (2002), Беллвуд и Ренфрю (ред.), Заключение неолитической истории населения с использованием гаплотипов Y-хромосомы, Кембридж: Институт археологических исследований Макдональда, ISBN 978-1-902937-20-5
- Андерхилл и Кивисилд; Кивисилд, Т. (2007), «Использование структуры популяции Y-хромосомы и митохондриальной ДНК в отслеживании миграций человека», Анну. Преподобный Жене., 41: 539–64, Дои:10.1146 / annurev.genet.41.110306.130407, PMID 18076332
- Perlès C, Monthel G (2001) Ранний неолит в Греции: первые фермерские сообщества в Европе. Издательство Кембриджского университета, Кембридж.
- Руннелс С. (2003) Истоки греческого неолита: личный взгляд, Аммерман и Бьяджи (редакторы 2003 г.).
внешняя ссылка
| Подтемы | |
|---|---|
| Генетическая история по регионам | |
| Популяционная генетика по группе |
|
| |