Гаплогруппа R1b - Haplogroup R1b
| Гаплогруппа R1b | |
|---|---|
| Возможное место происхождения | Западная Азия[1]или же Центральная Азия[2] |
| Предок | R1 |
| Потомки |
|
| Определение мутаций | M343 |
Гаплогруппа R1b (R-M343), также известный как Hg1 и Eu18, это гаплогруппа Y-хромосомы человека.
Это наиболее часто встречающаяся отцовская линия в западная Европа, а также некоторые части Россия (например, Башкирский меньшинство) и Центральная Африка (например. Чад и Камерун ). Клада также присутствует на более низких частотах повсюду. Восточная Европа, Западная Азия, а также части Северная Африка и Центральная Азия.
R1b имеет две основные ветви: R1b1a-L754 и R1b1b-PH155. R1b1a1a2-M269, который преобладает в Западной Европе, и R1b1a2-V88, распространенный в Центральной Африке, оба являются субкладами R1b-L754. R1b1b-PH155 настолько редок и широко распространен, что трудно сделать какие-либо выводы о его происхождении. Это было найдено в Бахрейн, Бутан, Ладакх, Таджикистан, Турция и Западный Китай.
Согласно древним исследованиям ДНК, R1a и большая часть R1b расширилась бы из Каспийское море вместе с Индоевропейские языки.[3][4][5][6][7]
Происхождение и распространение

Возраст R1 оценили Татьяна Карафет и др. (2008 г.) от 12 500 до 25 700 BP, и, скорее всего, произошло около 18 500 лет назад.[8] Поскольку самый ранний известный образец датируется примерно 14000 лет назад и принадлежит к группе R1b1a (R-L754),[9] R1b должен был возникнуть относительно вскоре после появления R1.
Ранние человеческие останки, содержащие R1b, включают:
- Виллабруна 1 (индивидуальный I9030), а Западный охотник-собиратель (WHG), найденный в Эпиграветтин культурная обстановка в Cismon долина (современный Венето, Италия), живший около 14000 г. BP и принадлежал R1b1a.[9][10]
- Несколько самцов Железные ворота мезолита в Балканы захоронен между 11200 и 8200 лет назад, содержал R1b1a1a. Было установлено, что эти люди в основном имеют происхождение WHG, с небольшими Восточный охотник-собиратель (EHG) примесь.[11]
- Несколько самцов мезолита Кунда культура и неолит Нарвская культура похоронен в Могильник Звейниеки в наши дни Латвия c. 9500–6000 лет назад несли R1b1b.[12][13] Было установлено, что эти люди в основном происходили из предков WHG с небольшой примесью EHG.[12]
- Несколько мужчин мезолита и неолита похоронены в Дериевка и Васильевка в современной Украине c. 9500-7000 BP несло R1b1a.[14] Эти люди были в основном предками EHG со значительной примесью WHG.[15]
- Мужчина WHG похоронен в Островул Корбули, Румыния c. 8700 BP несло R1b1c.[16][17][18]
- Мужчина похоронен в Лепенски Вир, Сербия гр. 8200-7900 BP несло R1b1a.[19]
- EHG похоронен рядом Самара, Россия 7500 BP несла R1b1a1a.[20]
- An Энеолит мужчина похоронен в Хвалынск, Россия гр. 7200-6000 л.н. несло R1b1a.[21]
- А Неолит мужчина похоронен в Els Trocs, Испания гр. 7178-7066 БП, который, возможно, принадлежал к Эпи-Кардиал культура[22] оказался носителем R1b1.[23][24][25]
- Похоронен мужчина позднего энеолита. Смядово, Болгария c. 6500 BP несло R1b1a.[26]
- Мужчина раннего медного века похоронен в Каннас-ди-Сотто, Карбония, Сардиния c. 6450 лет назад несли R1b1b2.[27]
- Самец Культура Михельсберга или же Вартберг культура похоронен в Blätterhöhle, Германия c. 6000-5300 лет назад несли R1b1 и имели примерно 50% -ное происхождение WHG и 50% Ранний европейский фермер (EFF) родословная.[28]
- Самец Группа Баальберге в Центральной Европе похоронен ок. 5600 BP несли R1b1a.[29]
- Самец Ботайская культура в Центральная Азия похоронен c. 5500 лет назад несли R1b1a1 (R1b-M478).[30]
- Самцы близкородственных Ямная культура[14] (ок. 5300-4800 лет назад) Афанасьевская культура[31][32] (5300-4500 БП), Катакомбная культура (4800-3700 лет назад), Полтавская культура[14] (4700-4100 лет назад) и Культура колокольчиков (4800-3800 лет назад) Евразии в подавляющем большинстве несут R1b1a1a2a2.[14][33]
Считается, что точка происхождения R1b находится в Западная Евразия, скорее всего в Западная Азия.[34] R1b является субкладом в «макро-гаплогруппа " K (M9), самая распространенная группа человеческих мужских линий за пределами Африки. Считается, что K возник в Азии (как и в случае с еще более ранней наследственной гаплогруппой, F (Ф-М89). Карафет Т. и соавт. (2014) «быстрый процесс диверсификации K-M526 вероятно произошло в Юго-Восточная Азия, с последующим расширением на запад предков гаплогрупп р и Q ".[35]
Три генетических исследования в 2015 году подтвердили Курганская гипотеза из Мария Гимбутас взяв во внимание Протоиндоевропейская родина. Согласно этим исследованиям, гаплогруппы R1b-M269 и R1a, наиболее распространенные в настоящее время в Европе (R1a также распространена в Южной Азии), распространились из Западной Евразийской степи вместе с Индоевропейские языки; они также обнаружили аутосомный компонент, присутствующий у современных европейцев, которого не было в Европейцы эпохи неолита, которые были введены с отцовскими линиями R1b и R1a, а также с индоевропейскими языками.[3][4][5]
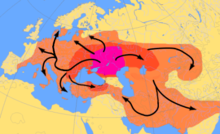
Анализ древней Y-ДНК из останков раннего неолита Центральной и Северной Европы Линейная гончарная культура В поселениях еще не обнаружены самцы, принадлежащие к гаплогруппе R1b-M269.[36][37] Olalde et al. (2017) проследили распространение гаплогруппы R1b-M269 в Западной Европе, особенно в Великобритании, с распространением Стакан культуры, с внезапным появлением многих гаплогрупп R1b-M269 в Западной Европе ок. 5000–4500 лет назад в эпоху ранней бронзы.[38] В 2016 году Природа статья «Генетическая история ледникового периода Европы».[39]
D'Atanasio et al. (2018) предполагают, что R1b-V88 возник в Европе около 12 000 лет назад и перебрался в Северную Африку около 8000 лет назад; возможно, раньше он был обычным явлением в южной Европе, где с тех пор он был заменен волнами других гаплогрупп, оставив остатки субкладов почти исключительно в Сардиния. Впервые он излучился в Африке, вероятно, между 7 и 8000 лет назад - в то же время, что и транссахарские экспансии внутри несвязанных гаплогрупп E-M2 и A-M13 - возможно, из-за роста населения, допущенного влажными условиями и внедрением животноводства. в Сахаре. R1b-V1589, главный подклад в R1b-V88, подвергся дальнейшему расширению около 5500 лет назад, вероятно, в районе бассейна озера Чад, из которого некоторые линии пересекали Сахару в Северную Африку.[40] Секвенирование ДНК древних людей является убедительным доказательством этой предложенной модели транссахарского движения с севера на юг: самые ранние базальные гаплогруппы R1b-V88 обнаружены у нескольких восточноевропейских охотников-собирателей около 10 000 лет назад. Затем гаплогруппа, по-видимому, получила дальнейшее распространение с эпохой неолита. Кардиальная посуда экспансия, которая привела к развитию сельского хозяйства в Западном Средиземноморье около 7500 г. до н.э.: гаплогруппы R1b-V88 были идентифицированы у людей эпохи неолита в центральной Италии, Иберии и, особенно часто, на Сардинии.[41] Часть ветви, ведущей к современным африканским гаплогруппам (V2197), уже произошла от некоторых из этих древних неолитических европейских людей, обеспечивая дополнительную поддержку транссахарскому движению с севера на юг.
Структура
Внешняя филогения R1b
Более широкая гаплогруппа р (M207) - первичный подклад гаплогруппы P1 (M45) сама является основной ветвью п (P295), которая также известна как гаплогруппа K2b2. Таким образом, R-M207 является вторичной ветвью K2b (P331), и прямой потомок K2 (M526).
Произошла «начальная быстрая диверсификация» K-M526, по данным Karafet et al. (2014), что "вероятно, произошло в Юго-Восточная Азия, с последующим расширением на запад предков гаплогрупп R и Q ".
- Филогения внутри K2b
- п P295 / PF5866 / S8 (также известный как K2b2).
Внутренняя структура R1b
Такие имена, как R1b, R1b1 и т. Д., Являются филогенетический (т. е. «генеалогическое древо») имена, которые ясно показывают свое место в ветвлении гаплогрупп или филогенетическом дереве. Альтернативный способ наименования одних и тех же гаплогрупп и субкладов относится к их определению. SNP мутации: например, R-M343 эквивалентен R1b.[42] Филогенетический имена меняются с новыми открытиями и имена на основе SNP, следовательно, реклассифицируются в филогенетическом дереве. В некоторых случаях SNP оказывается ненадежным в качестве определяющей мутации, и имя на основе SNP полностью удаляется. Например, до 2005 года R1b был синонимом R-P25, который позже был переклассифицирован как R1b1; в 2016 году R-P25 был полностью удален как определяющий SNP из-за значительной скорости обратной мутации.[43] (Ниже представлена основная схема R1b в соответствии с деревом ISOGG на 30 января 2017 года.[1])
|
Географическое распределение
R1b * (R-M343 *)
Ни о каких подтвержденных случаях R1b * (R-M343 *) - то есть R1b1 (xR1b1), также известного как R-M343 (xL278) - не сообщалось в рецензируемый литература.
Аналогичным образом не было обнаружено никаких известных примеров R1b1 *, также известного как R-L278 * и R-L278 (xL754, PH155).
- R-M343 (xM73, M269, V88)
В ранних исследованиях, поскольку R-M269, R-M73 и R-V88 на сегодняшний день являются наиболее распространенными формами R1b, иногда предполагалось, что примеры R1b (xM73, xM269) обозначают базальные примеры «R1b *».[43] Однако, хотя парагруппа R-M343 (xM73, M269, V88) встречается редко, она не исключает членства в редких и / или обнаруженных впоследствии относительно базальных субкладах R1b, таких как R-L278 * (R1b1 *), R -L389 * (R1b1a1 *), R-P297 * (R1b1a1a *), R-V1636 (R1b1a1b) или R-PH155 (R1b1b).
Считается, что наибольшая доля R-M343 (xM73, M269, V88) имеет население Курды юго-востока Казахстан с 13%.[34] Однако совсем недавно было проведено большое исследование вариации Y-хромосомы у Иран, выявили, что R-M343 (xV88, M73, M269) достигает 4,3% среди иранских субпопуляций.[45]
Субклады R1b также были обнаружены в Хань китайский из провинций Шаньдун, Хэйлунцзян и Ганьсу.[46]
Остается возможность, что некоторые или даже большинство из этих случаев могут быть R-L278 * (R1b1 *), R-L389 * (R1b1a1 *), R-P297 * (R1b1a1a *), R-V1636 (R1b1a1b), R-PH155 (R1b1b), R1b * (R-M343 *), R1a * (R-M420 *), иначе недокументированная ветвь R1 (R-M173) и / или обратные мутации маркера из положительного к отрицательному наследственному состоянию,[47] и, следовательно, составляют недокументированные субклады R1b.
Сборник предыдущих исследований распределения R1b можно найти в Cruciani et al. (2010).[48] Это кратко изложено в следующей таблице. (Cruciani не включил некоторые исследования, предполагающие даже более высокие частоты R1b1a1a2 [R-M269] в некоторых частях Западной Европы.)
- Распространение R-V88, R-M73 и M269
| Континент | Область, край | Размер образца | Итого R1b | R-P25 (ненадежный маркер для R1b1 *) | R-V88 (R1b1a2) | R-M269 (R1b1a1a2) | R-M73 (R1b1a1a1) |
|---|---|---|---|---|---|---|---|
| Африка | Северная Африка | 691 | 5.9% | 0.0% | 5.2% | 0.7% | 0.0% |
| Африка | Центральный Сахельский регион | 461 | 23.0% | 0.0% | 23.0% | 0.0% | 0.0% |
| Африка | Западная африка | 123 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Африка | Восточная африка | 442 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Африка | Южная Африка | 105 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Европа | Западные европейцы | 465 | 57.8% | 0.0% | 0.0% | 57.8% | 0.0% |
| Европа | Северо-западные европейцы | 43 | 55.8% | 0.0% | 0.0% | 55.8% | 0.0% |
| Европа | Центральные европейцы | 77 | 42.9% | 0.0% | 0.0% | 42.9% | 0.0% |
| Европа | Северо-восточные европейцы | 74 | 1.4% | 0.0% | 0.0% | 1.4% | 0.0% |
| Европа | Россияне | 60 | 6.7% | 0.0% | 0.0% | 6.7% | 0.0% |
| Европа | Восточноевропейцы | 149 | 20.8% | 0.0% | 0.0% | 20.8% | 0.0% |
| Европа | Юго-восточные европейцы | 510 | 13.1% | 0.0% | 0.2% | 12.9% | 0.0% |
| Азия | Жители Западной Азии | 328 | 5.8% | 0.0% | 0.3% | 5.5% | 0.0% |
| Азия | Выходцы из Южной Азии | 288 | 4.8% | 0.0% | 0.0% | 1.7% | 3.1% |
| Азия | Юго-восточные азиаты | 10 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Азия | Северо-восточные азиаты | 30 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Азия | Выходцы из Восточной Азии | 156 | 0.6% | 0.0% | 0.0% | 0.6% | 0.0% |
| ОБЩИЙ | 5326 |
R1b1 (R-L278)
R-L278 среди современных мужчин попадает в субклады R-L754 и R-PH155, хотя возможно, что некоторые очень редкие R-L278 * могут существовать, поскольку не все примеры были протестированы для обеих ветвей. Примеры могут также существовать в древней ДНК, хотя из-за низкого качества часто невозможно сказать, были ли у древних мутации, которые определяют субклады.
Некоторые примеры описаны в более старых статьях, например, два найдены в Турции,[42] в настоящее время считается, что они в основном относятся к недавно обнаруженной подкладе R1b1a2 (R-V88). Таким образом, большинство примеров R1b попадают в субклады R1b1a2 (R-V88) или R1b1a (R-P297). Cruciani et al. в большом исследовании 2010 года было обнаружено 3 случая среди 1173 итальянцев, 1 из 328 жителей Западной Азии и 1 из 156 жителей Восточной Азии.[48] Варзари обнаружил 3 случая в Украина, в исследовании 322 человек из Днестр –Карпаты области, которые были P25 положительными, но M269 отрицательными.[49] Случаи из более ранних исследований в основном относятся к Африке, Ближнему Востоку или Средиземноморью и рассматриваются ниже как вероятные случаи R1b1a2 (R-V88).
R1b1a (R-L754)
R-L754 содержит подавляющее большинство R1b. Единственный известный пример R-L754 * (xL389, V88) также является самым ранним из известных носителей R1b: "Виллабруна 1 ", живший около 14 000 лет назад (северо-восток Италии). Виллабруна 1 принадлежала к Эпиграветтин культура.
R1b1a1 (R-L389)
R-L389, также известный как R1b1a1 (L388 / PF6468, L389 / PF6531), содержит очень распространенный субклад R-P297 и редкий субклад R-V1636. Неизвестно, принадлежат ли все ранее описанные R-L389 * (xP297) к R-V1636 или нет.
R1b1a1a (R-P297)
Маркер SNP P297 был признан в 2008 г. предком значимых субкладов M73 и M269, объединив их в один кластер.[8] Этому было дано филогенетическое название R1b1a1a (а ранее - R1b1a).
Большая часть евразийского R1b попадает в этот субклад, представляя очень большую современную популяцию. Хотя сам по себе P297 еще не подвергался серьезным испытаниям, эта же популяция была относительно хорошо изучена с точки зрения других маркеров. Следовательно, ветвление внутри этой клады может быть объяснено ниже относительно подробно.
R1b1a1a1 (R-M73)
Малярчук и другие. (2011) обнаружили R-M73 у 13,2% (5/38) шорцев, 11,4% (5/44) телеутов, 3,3% (2/60) калмыков, 3,1% (2/64) хакасов, 1,9%. (2/108) тувинцев и 1,1% (1/89) алтайцев.[50] Калмыки, тувинцы и алтайцы принадлежат к кластеру Y-STR, обозначенному DYS390 = 19, DYS389 = 14-16 (или 14-15 в случае алтайской особи) и DYS385 = 13-13.
Дулик и другие. (2012) обнаружили R-M73 в 35,3% (6/17) образцов Кумандин Республики Алтай в России.[51] Три из этих шести кумандинов имеют одинаковый гаплотип Y-STR с 15 локусами, а еще два отличаются только по локусу DYS458, имея DYS458 = 18 вместо DYS458 = 17. Эта пара кумандинских гаплотипов R-M73 напоминает гаплотипы двух калмыков, двух тувинцев и одного алтайца, Y-ДНК которых была проанализирована Малярчуком. и другие. (2011). У оставшегося кумандинца R-M73 есть гаплотип Y-STR, который резко отличается от гаплотипов других кумандинцев R-M73, напоминая гаплотипы пяти шорцев, пяти телеутов и двух хакасов.[50]
В то время как ранние исследования R-M73 утверждали, что он был значительно представлен среди Хазара Афганистана и Башкиры Уральских гор, это, очевидно, было отменено. Например, вспомогательные материалы из исследования Бехара 2010 г. и другие. предположил, что Сенгупта и другие. (2006) могли ошибочно идентифицировать хазарейцев, которые вместо этого принадлежали к «PQR2», а не к «R (xR1a)».[52][34][53] Однако отнесение Y-ДНК этих хазарейцев к категории "PQR2" Бехаром и другие. (2010), вероятно, можно приписать привычке, которая какое-то время была популярной, обозначать R-M269 как «R1b» или «R (xR1a)», при этом любые члены R-M343 (xM269) помещаются в полифилетический, улавливающий - все категории "R *" или "P". Myres и другие. (2011), Ди Кристофаро и другие. (2013) и Липпольд и другие. (2014) все согласны с тем, что Y-ДНК 32% (8/25) образца HGDP из пакистанской Хазары должна принадлежать к гаплогруппе R-M478 / M73.[34][54][55] Аналогичным образом было обнаружено, что большинство башкирских мужчин принадлежали к U-152 (R1b1a1a2a1a2b), а некоторые, в основном из юго-восточного Башкортостана, принадлежали к U-152. Гаплогруппа Q-M25 (Q1a1b), а не R1b; против этого, Майрес и другие. (2011) обнаружили высокую частоту R-M73 среди своей выборки башкир из юго-восточного Башкортостана (77/329 = 23,4% R1b-M73), что согласуется с более ранним исследованием башкир.[34] Помимо высокой частоты R-M73 у юго-восточных башкир, Myres и другие. также сообщили о обнаружении R-M73 в следующих образцах: 10,3% (14/136) Балкарцы из северо-западного Кавказа, 9,4% (8/85) образцов HGDP из северного Пакистана (это уже упомянутые пакистанские хазарейцы), 5,8% (4/69) образцов Карачаевцы с северо-запада Кавказа, 2,6% (1/39) татар из Башкортостана, 1,9% (1/54) башкир из юго-запада Башкортостана, 1,5% (1/67) Мегрелы с Южного Кавказа, 1,4% (1/70) башкир из Северного Башкортостана, 1,3% (1/80) татар из Казани, 1,1% (1/89) выборки из Каппадокии, Турция, 0,7% (1 / 141) из Кабардинцы из Северо-Западного Кавказа 0,6% (3/522) пула выборок из Турции и 0,38% (1/263) русских из Центральной России.[34]
Помимо вышеупомянутых пакистанских хазарейцев, Ди Кристофаро и другие. (2013) обнаружили R-M478 / M73 в 11,1% (2/18) Монголы из центральной Монголии, 5,0% (1/20) Кыргызский из юго-запада Кыргызстана, 4,3% (1/23) монголов из юго-восточной Монголии, 4,3% (4/94) Узбеки из Джузджана, Афганистан, 3,7% (1/27) иранцев из Гилан, 2,5% (1/40) киргизов из центрального Кыргызстана, 2,1% (2/97) монголов из северо-западной Монголии и 1,4% (1/74) Туркмены из Джаузджана, Афганистан.[54] Монголы, а также индивид с юго-запада Кыргызстана, индивид из Гиляна и один из узбеков из Джаузджана принадлежат к тому же кластеру гаплотипов Y-STR, что и пять из шести кумандинских членов R-M73, изученных Дуликом. и другие. (2012). Наиболее характерное значение Y-STR для этого кластера - DYS390 = 19.[34]
Карафет и другие. (2018) обнаружили R-M73 в 37,5% (15/40) выборке телеутов из Беково Кемеровской области, 4,5% (3/66) уйгуров из выборки Синьцзян-Уйгурского автономного района, 3,4% (1/29 ) выборки казахов из Казахстана, 2,3% (3/129) выборки селькупов, 2,3% (1/44) выборки туркмен из Туркменистана и 0,7% (1/136) выборки иранцев. из Ирана.[56] Четыре из этих индивидов (один из телеутов, один из уйгуров, казах и иранец), по-видимому, принадлежат к вышеупомянутому кластеру, обозначенному DYS390 = 19 (кумандино-монгольский кластер R-M73); Телеуты и уйгуры также разделяют модальные значения в локусах DYS385 и DYS389. Иранский отличается от модального для этого кластера тем, что имеет 13-16 (или 13-29) на DYS389 вместо 14-16 (или 14-30). Казахский отличается от модального тем, что у DYS385 13–14 вместо 13-13. Остальные четырнадцать Телеутов и три селькупа принадлежат к Телеутско-Шорско-Хакасскому кластеру R-M73 из набора данных Малярчука. и другие. (2011); этот кластер имеет модальные значения DYS390 = 22 (но 21 в случае двух телеутов и одного хакасца), DYS385 = 13-16 и DYS389 = 13-17 (или 13-30, но 14-31 в случае один селькуп).
В казахстанской статье, опубликованной в 2017 году, гаплогруппа R1b-M478 Y-ДНК обнаружена в 3,17% (41/1294) образца Казахи из Казахстана, при этом эта гаплогруппа наблюдается с большей, чем средняя частота, среди членов Кыпшак (12/29 = 41,4%), Ысты (6/57 = 10,5%), Конгырат (8/95 = 8,4%), Ошакты (2 / 29 = 6,9%), Керей (1/28 = 3,6%) и Джетыру (3/86 = 3,5%) племена.[57] В китайской статье, опубликованной в 2018 году, была обнаружена Y-ДНК гаплогруппы R1b-M478 в 9,2% (7/76) образца Долан Уйгуры из поселка Хориколь, Округ Ават, Синьцзян.[58]
R1b1a1a2 (R-M269)
R-M269 или R1b1a1a2 (по состоянию на 2017 год) среди других имен,[59] в настоящее время является наиболее распространенной линией Y-ДНК у европейских мужчин. Его переносят примерно 110 миллионов мужчин в Европе.[60]

R-M269 вызвал значительный научный и популярный интерес из-за его возможного подключения к Индоевропейская экспансия в Европе. В частности, было обнаружено, что субклад R-L23 (R-Z2103) преобладает в древней ДНК, связанной с Ямная культура.[61] Было установлено, что семь человек принадлежат к субкладу R1b-M269.[3]
Более ранние исследования, опубликованные до того, как исследователи смогли изучить ДНК древних останков, предположили, что R-M269, вероятно, возник в Западной Азии и присутствовал в Европе в период неолита.[1][34][62][63] Но результаты, основанные на реальной древней ДНК, показали, что до бронзового века в Европе был дефицит R-M269,[3] а распространение субкладов в Европе в значительной степени связано с различными миграциями Бронза и Железный век. Точно так же самые старые образцы, классифицируемые как принадлежащие R-M269, были обнаружены в Восточной Европе и степях Причерноморско-Каспийского региона, а не в Западной Азии. Западноевропейское население делится на субклады R-P312 / S116 и R-U106 / S21 R-M412 (R-L51).
Распространение R-M269 в Европе увеличивается с востока на запад. На национальном уровне он достигает пика в Уэльс в размере 92%, в 82% в Ирландия, 70% в Шотландия, 68% в Испания, 60% в Франция (76% в Нормандия ), около 60% в Португалия,[34] 45% в Восточной Англия, 50% в Германия, 50% в Нидерланды, 42% в Исландия, и 43% в Дания, 39% в Италия.R-M269 достигает уровня 95% в некоторых частях Ирландии. Он также был обнаружен на более низких частотах по всей центральной Евразия,[64] но с относительно высокой частотой среди Башкиры из Пермь область (84,0%).[65] Этот маркер присутствует в Китае и Индии с частотой менее одного процента. В Северная Африка и прилегающие острова, в то время как R-V88 (R1b1a2) более широко представлен, R-M269, по-видимому, присутствует с древних времен. R-M269 был обнаружен, например, в количестве ~ 44% среди останков, датируемых 11-13 веками. Пунта Азул, в Канарские острова. Эти останки были связаны с Bimbache (или бимапе), подгруппа гуанчей.[66] У живых самцов он достигает пика в некоторых частях Северной Африки, особенно Алжир, в размере 10%.[67] В Африке к югу от Сахары R-M269, по-видимому, достигает пика в Намибия, в размере 8% среди Гереро самцы.[68] В Западной Азии R-M269 был зарегистрирован в 40% случаев. Армянский самцы.[69][требуется проверка ] (В таблице ниже более подробно перечислены частоты M269 в регионах Азии, Европы и Африки.)
Помимо нерасширенных, базальный R-M269 *, существует (по состоянию на 2017 год) две основные ветви R-M269:
- R-L23 (R1b1a1a2a; L23 / PF6534 / S141) и
- R-PF7558 (R1b1a1a2b; PF7558 / PF7562.)
R-L23 (Z2105 / Z2103; a.k.a. R1b1a1a2a) был зарегистрирован среди народов Идель-Урал (Трофимова и др., 2015): 21 из 58 (36,2%) Бурзянский район Башкиры, 11 из 52 (21,2%) Удмуртов, 4 из 50 (8%) из Коми, 4 из 59 (6,8%) Мордвины, 2 из 53 (3,8%) из Бесермян и 1 из 43 (2,3%) Чувашский были R1b-L23.[70]
Субклады в парагруппе R-M269 (xL23), то есть R-M269 * и / или R-PF7558, по всей видимости, наиболее часто встречаются в центральной Балканы, особенно Косово с 7,9%, Македония 5,1% и Сербия 4.4%.[34] В отличие от большинства других областей со значительным процентным содержанием R-L23, Косово, Польша и Башкиры юго-востока Башкортостан примечательны высоким процентным содержанием R-L23 (xM412), также известного как R1b1a1a2a (xR1b1a1a2a1) - 11,4% (Косово), 2,4% (Польша) и 2,4% на юго-востоке Башкортостана.[34] (Эта башкирская популяция также отличается высоким уровнем R-M73 (R1b1a1a1) - 23,4%.[34]) Пять человек из 110 протестированных в Араратская долина из Армения принадлежал R-M269 (xL23) и 36 к R-L23 *, при этом ни один из них не принадлежал к известным субкладам L23.[71]
В 2009 году ДНК, извлеченная из бедренных костей 6 скелетов из раннесредневекового захоронения в г. Эргольдинг (Бавария, Германия), датируемые примерно 670 г. н.э., дали следующие результаты: 4 оказались гаплогруппой R1b с наиболее близкими совпадениями в современных популяциях Германии, Ирландии и США, а 2 были в гаплогруппе R1b. Гаплогруппа G2a.[72]
Ниже приводится краткое изложение большинства исследований, специально протестированных на M269, с указанием его распределения (в процентах от общей численности населения) в Европе. Северная Африка, то Средний Восток и Центральная Азия до Китая и Непал.
Филогения R-M269 согласно ISOGG 2017:
| M269 / PF6517 |
| |||||||||||||||||||||||||||||||||||||||||||||
R1b1a2 (R-V88)
R1b1a2 (PF6279 / V88; ранее R1b1c) определяется наличием SNP-маркера V88, открытие которого было объявлено в 2010 году Cruciani et al.[48] За исключением жителей Южной Европы и Западной Азии, большинство R-V88 было обнаружено в Сахеле, особенно среди населения, говорящего Афроазиатские языки из Чадский ответвляться.
В исследованиях 2005–2008 годов сообщалось о высоком уровне «R1b *» в Иордания, Египет и Судан. Однако последующие исследования показывают, что рассматриваемые образцы, скорее всего, принадлежат субкладу R-V88, который сейчас сосредоточен в К югу от Сахары, после миграции из Азии.[73][34][74][75][68]
- Распространение R1b в Африке
| Область, край | численность населения | Страна | Язык | N | Общий% | R1b1c (R-V88) | R1b1a1a2 (R-M269) | R1b1c * (R-V88 *) | R1b1c3 (R-V69) |
|---|---|---|---|---|---|---|---|---|---|
| Северная Африка | Композитный | Марокко | AA | 338 | 0.0% | 0.3% | 0.6% | 0.3% | 0.0% |
| Северная Африка | Мозабитские берберы | Алжир | AA / берберский | 67 | 3.0% | 3.0% | 0.0% | 3.0% | 0.0% |
| Северная Африка | Северные египтяне | Египет | AA / семитский | 49 | 6.1% | 4.1% | 2.0% | 4.1% | 0.0% |
| Северная Африка | Берберы из Сивы | Египет | AA / берберский | 93 | 28.0% | 26.9% | 1.1% | 23.7% | 3.2% |
| Северная Африка | Бахария | Египет | AA / семитский | 41 | 7.3% | 4.9% | 2.4% | 0.0% | 4.9% |
| Северная Африка | Гурна Оазис | Египет | AA / семитский | 34 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Северная Африка | Южные египтяне | Египет | AA / семитский | 69 | 5.8% | 5.8% | 0.0% | 2.9% | 2.9% |
| C Африка | Сонгай | Нигер | NS / Songhai | 10 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| C Африка | Фульбе | Нигер | Северная Каролина / Атлантика | 7 | 14.3% | 14.3% | 0.0% | 14.3% | 0.0% |
| C Африка | Туареги | Нигер | AA / берберский | 22 | 4.5% | 4.5% | 0.0% | 4.5% | 0.0% |
| C Африка | Нгамбай | Чад | NS / суданский | 11 | 9.1% | 9.1% | 0.0% | 9.1% | 0.0% |
| C Африка | Хауса | Нигерия (север) | AA / Чад | 10 | 20.0% | 20.0% | 0.0% | 20.0% | 0.0% |
| C Африка | Фульбе | Нигерия (север) | Северная Каролина / Атлантика | 32 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| C Африка | Йоруба | Нигерия (юг) | NC / Defoid | 21 | 4.8% | 4.8% | 0.0% | 4.8% | 0.0% |
| C Африка | Ouldeme | Камерун (север) | AA / Чад | 22 | 95.5% | 95.5% | 0.0% | 95.5% | 0.0% |
| C Африка | Мада | Камерун (север) | AA / чадский | 17 | 82.4% | 82.4% | 0.0% | 76.5% | 5.9% |
| C Африка | Мафа | Камерун (север) | AA / чадский | 8 | 87.5% | 87.5% | 0.0% | 25.0% | 62.5% |
| C Африка | Guiziga | Камерун (север) | AA / Чад | 9 | 77.8% | 77.8% | 0.0% | 22.2% | 55.6% |
| C Африка | Даба | Камерун (север) | AA / Чад | 19 | 42.1% | 42.1% | 0.0% | 36.8% | 5.3% |
| C Африка | Guidar | Камерун (север) | AA / чадский | 9 | 66.7% | 66.7% | 0.0% | 22.2% | 44.4% |
| C Африка | Масса | Камерун (север) | AA / Чад | 7 | 28.6% | 28.6% | 0.0% | 14.3% | 14.3% |
| C Африка | Другой чадский | Камерун (север) | AA / Чад | 4 | 75.0% | 75.0% | 0.0% | 25.0% | 50.0% |
| C Африка | Шува арабы | Камерун (север) | AA / семитский | 5 | 40.0% | 40.0% | 0.0% | 40.0% | 0.0% |
| C Африка | Канури | Камерун (север) | NS / Saharan | 7 | 14.3% | 14.3% | 0.0% | 14.3% | 0.0% |
| C Африка | Фульбе | Камерун (север) | Северная Каролина / Атлантика | 18 | 11.1% | 11.1% | 0.0% | 5.6% | 5.6% |
| C Африка | Moundang | Камерун (север) | NC / Adamawa | 21 | 66.7% | 66.7% | 0.0% | 14.3% | 52.4% |
| C Африка | Фали | Камерун (север) | NC / Adamawa | 48 | 20.8% | 20.8% | 0.0% | 10.4% | 10.4% |
| C Африка | Тали | Камерун (север) | NC / Adamawa | 22 | 9.1% | 9.1% | 0.0% | 4.5% | 4.5% |
| C Африка | Мбум | Камерун (север) | NC / Adamawa | 9 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| C Африка | Композитный | Камерун (Sth) | Северная Каролина / Банту | 90 | 0.0% | 1.1% | 0.0% | 1.1% | 0.0% |
| C Африка | Пигмеи Биаки | МАШИНА | Северная Каролина / Банту | 33 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Западная Африка | Композитный | — | 123 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| E Африка | Композитный | — | 442 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| Южная Африка | Композитный | — | 105 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| ОБЩИЙ | 1822 |
| V88 |
| ||||||||||||
Две ветви R-V88, R-M18 и R-V35, встречаются почти исключительно на острове Сардиния.
Как видно из приведенной выше таблицы данных, R-V88 находится в северной Камерун в западной и центральной Африке с очень высокой частотой, где считается, что это вызвано доисламским движением людей из Евразия.[68][76] С другой стороны, Gonzalez et al. (2013) обнаружили, что паттерны разнообразия африканских R1b-V88 не соответствовали перемещению говорящих на чадском языке людей с севера через Сахару в Западно-Центральную Африку, но были совместимы с обратным, происхождением линий V88 в Центрально-Западная Африка, затем миграция в Северную Африку.[77]
R1b1a2a (R-M18)
R1b1a2a является подкладой R-V88, которая определяется наличием маркера SNP M18.[8]Он обнаружен только на низких частотах в образцах из Сардиния[64][78] и Ливан.[79]
R1b1b (R-PH155)
Другая первичная ветвь R1b1 - это R-PH155 (R1b1b), которая встречается крайне редко и определяется наличием PH155.[1] Живые самцы, несущие субклады R-PH155, были обнаружены в Бахрейн, Бутан, Ладакх, Таджикистан, Индюк, Синьцзян, и Юньнань. ISOGG (2017) ссылается на две основные ветви: R-M335 (R1b1b1) и R-PH200 (R1b1b2).
Определяющий SNP для R1b1b1, M335, был впервые задокументирован в 2004 году, когда в Турции был обнаружен пример, хотя в то время он был классифицирован как R1b4.[42] Другие примеры R-M335 были зарегистрированы в выборке Хуэй из Юньнань, Китай[80] и в выборке людей из Ладакх, Индия.[81] При коммерческом тестировании Y-ДНК R-M335 был обнаружен у лиц, сообщивших о происхождении от отца в Германии и Италии (включая Арбёрешэ ).[82]
Примеры другого субклада R-PH155, т.е. R1b1b2-PH200 были обнаружены у лиц из Турции (Конья и Газиантеп, по крайней мере, последний отчет Армянский этническая принадлежность), Бахрейн, и Бутан.[82]
Другие примеры R-PH155 с нерешенными точными субкладами были обнаружены в Таджикский в Таджикистане и в Уйгурский в академических исследованиях и у человека, сообщившего о своем отцовском происхождении в Варанаси, Индия в коммерческом тестировании.[82]
Исторические люди R1b
Ниже приведены исторические люди или династии, которые могут принадлежать к гаплогруппе R1b, как предполагают потомки тестирования или другие родственники:
- Самцы Дом Бурбонов и, соответственно, все ветви Династия капетингов. (R1b1b2a1a1b / R-Z381).[83]
- Чарльз Дарвин.[84]
- Найл из девяти заложников.[85]
- Египетский фараон Аменхотеп III, Эхнатон и Тутанхамон[86]
В популярной культуре
- Брайан Сайкс в своей книге 2006 г. Кровь островов, дает членам - и условному патриарху-основателю - R1b имя "Oisín ".
- Стивен Оппенгеймер в своей книге 2007 года Истоки британцев, дает патриарху R1b Баскский название «Руиско» в честь того, что Оппенгеймер считал иберийским происхождением R1b.
- Режиссер Артем Лукичев создал (около 2009 г.) 14-минутный анимационный фильм по мотивам Башкирский эпос из Уральские горы, связывая эпос с появлением и географическим расширением R1a и R1b.[87]
- ДНК-тесты, которые помогли идентифицировать царя Николай II России обнаружил, что он принадлежал R1b.[88] Это может означать, что более поздние цари Дом Романовых, произошедшие по мужской линии от Дом Гольштейн-Готторпский (который возник в Шлезвиг-Гольштейн ) также являются членами R1b.
Здоровье
Исследования показывают, что гаплогруппа R1b может оказывать защитное действие на иммунную систему,[89] и люди, принадлежащие к этой гаплогруппе, могут, скорее всего, не иметь симптомов, если они инфицированы вирусом SARS-2.[90]
Смотрите также
| Филогенетическое древо человека Гаплогруппы ДНК Y-хромосомы [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-хромосомный Адам " | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| грамм | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | ЧАС | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| я | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | Т | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | п [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| НЕТ | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | О | Q | р | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Рекомендации
- ^ а б c d "ISOGG 2017 Y-ДНК гаплогруппа R". isogg.org.
- ^ Чон Си (2018). «Характеризуя генетическую историю примесей во внутренней Евразии». bioRxiv 10.1101/327122. Дои:10.1101/327122. S2CID 90048752. Цитировать журнал требует
| журнал =(помощь) - ^ а б c d Хаак В., Лазаридис И., Паттерсон Н., Роланд Н., Маллик С., Лламас Б. и др. (Июнь 2015 г.). «Массовая миграция из степи была источником индоевропейских языков в Европе». Природа. 522 (7555): 207–11. arXiv:1502.02783. Bibcode:2015Натура.522..207H. bioRxiv 10.1101/013433. Дои:10.1038 / NATURE14317. ЧВК 5048219. PMID 25731166.
- ^ а б Allentoft ME, Sikora M, Sjögren KG, Rasmussen S, Rasmussen M, Stenderup J и др. (Июнь 2015 г.). «Популяционная геномика Евразии бронзового века». Природа. 522 (7555): 167–72. Bibcode:2015Натура.522..167A. Дои:10.1038 / природа14507. PMID 26062507. S2CID 4399103.
- ^ а б Мэтисон И., Лазаридис И., Роланд Н., Маллик С., Паттерсон Н., Роденберг С.А. и др. (2015). «Восемь тысяч лет естественного отбора в Европе». bioRxiv: 016477. Дои:10.1101/016477. S2CID 7866359.
- ^ Кэссиди Л.М., Мартиниано Р., Мерфи Е.М., Тисдейл, доктор медицины, Мэллори Дж., Хартвелл Б., Брэдли Д.Г. (январь 2016 г.). «Миграция эпохи неолита и бронзового века в Ирландию и установление островного атлантического генома». Труды Национальной академии наук Соединенных Штатов Америки. 113 (2): 368–73. Bibcode:2016PNAS..113..368C. Дои:10.1073 / pnas.1518445113. ЧВК 4720318. PMID 26712024.
- ^ Мартиниано Р., Кэссиди Л. М., О'Маолдуин Р., Маклафлин Р., Сильва Н. М., Манко Л. и др. (Июль 2017 г.). «Геномика населения археологического перехода на западе Иберии: исследование древней субструктуры с использованием методов вменения и гаплотипов». PLOS Genetics. 13 (7): e1006852. Дои:10.1371 / journal.pgen.1006852. ЧВК 5531429. PMID 28749934.
- ^ а б c Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (май 2008 г.). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека». Геномные исследования. 18 (5): 830–8. Дои:10.1101 / gr.7172008. ЧВК 2336805. PMID 18385274.
- ^ а б Fu Q, Posth C, Hajdinjak M, Petr M, Mallick S, Fernandes D и др. (Июнь 2016). «Генетическая история ледникового периода Европы». Природа. 534 (7606): 200–5. Bibcode:2016Натура.534..200F. Дои:10.1038 / природа17993. HDL:10211.3/198594. ЧВК 4943878. PMID 27135931.
- ^ Мэтисон 2018, Дополнительная таблица 1, строка 467.
- ^ Мэтисон 2018, Дополнительная таблица 1, строки 251–272.
- ^ а б Джонс 2017.
- ^ Мэтисон 2018, Дополнительная таблица 1, строки 205–245.
- ^ а б c d Мэтисон 2018, Дополнительная таблица 1.
- ^ Мэтисон 2018, стр. 2-3.
- ^ González-Fortes et al. 2017 г., стр. 8-9.
- ^ González-Fortes et al. 2017 г., п. 4, таблица 1, OC1_Meso.
- ^ Санчес-Квинто и др. 2019 г., Расширенный набор данных 1.3, OC1.
- ^ Мэтисон 2018, Дополнительная таблица 1, строка 298, I4666.
- ^ Мэтисон 2018, Дополнительная таблица 1, строка 153, I0124.
- ^ Мэтисон 2018, Дополнительная таблица 1, строка 375, I0122.
- ^ Хаак 2015, Дополнительная информация, стр. 26–27.
- ^ Хаак 2015, Таблица расширенных данных 2, I0410.
- ^ Хаак 2015, Дополнительная информация, стр. 44-45.
- ^ Мэтисон 2018, Дополнительная таблица 1, I0410.
- ^ Мэтисон 2018, Дополнительная таблица 1, строка 73, I2430.
- ^ Маркус и др. 2020 г., Дополнительные данные 1, основная таблица, строка 25, MA89.
- ^ Lipson et al. 2017 г., Sup Table 1, Sample Information, Row 121, Bla16.
- ^ Мэтисон 2018, Дополнительная таблица 1, строка 128, I0559.
- ^ де Баррос Дамгаард П., Мартиниано Р., Камм Дж., Морено-Майар СП, Кроонен Г., Пейрот М. и др. (Июнь 2018). «Первые пастухи и влияние степной экспансии ранней бронзы в Азию». Наука. 360 (6396): eaar7711. Дои:10.1126 / science.aar7711. ЧВК 6748862. PMID 29743352.
- ^ Нарасимхан и др. 2019 г., Таблица S1.
- ^ Hollard et al. 2018 г., стр. 6-7.
- ^ Onalde et al. 2019 г., Дополнительная таблица 4.
- ^ а б c d е ж грамм час я j k л м Myres NM, Rootsi S, Lin AA, Järve M, King RJ, Kutuev I, et al. (Январь 2011 г.). «Эффект основателя основной гаплогруппы R1b Y-хромосомы эпохи голоцена в Центральной и Западной Европе». Европейский журнал генетики человека. 19 (1): 95–101. Дои:10.1038 / ejhg.2010.146. ЧВК 3039512. PMID 20736979.
- ^ Karafet TM, Mendez FL, Sudoyo H, Lansing JS, Hammer MF (март 2015 г.). «Улучшенное филогенетическое разрешение и быстрая диверсификация гаплогруппы Y-хромосомы K-M526 в Юго-Восточной Азии». Европейский журнал генетики человека. 23 (3): 369–73. Дои:10.1038 / ejhg.2014.106. ЧВК 4326703. PMID 24896152.
- ^ Lacan M, Keyser C, Ricaut FX, Brucato N, Duranthon F, Guilaine J и др. (Июнь 2011 г.). «Древняя ДНК свидетельствует о распространении мужчин по неолитическому средиземноморскому пути». Труды Национальной академии наук Соединенных Штатов Америки. 108 (24): 9788–91. Bibcode:2011PNAS..108.9788L. Дои:10.1073 / pnas.1100723108. ЧВК 3116412. PMID 21628562.
- ^ Хаак В., Балановский О., Санчес Дж. Дж., Кошель С., Запорожченко В., Адлер С. Дж. И др. (Ноябрь 2010 г.). Пенни Д. (ред.). «Древняя ДНК европейских фермеров раннего неолита показывает их близость к востоку». PLOS Биология. 8 (11): e1000536. Дои:10.1371 / journal.pbio.1000536. ЧВК 2976717. PMID 21085689.
- ^ Olalde I, Brace S, Allentoft ME, Armit I, Kristiansen K, Booth T и др. (Март 2018 г.). «Феномен Стакана и геномная трансформация северо-западной Европы». Природа. 555 (7695): 190–196. Bibcode:2018Натура.555..190O. Дои:10.1038 / природа25738. ЧВК 5973796. PMID 29466337.
- ^ Fu Q, Posth C, Hajdinjak M, Petr M, Mallick S, Fernandes D и др. (Июнь 2016). «Генетическая история ледникового периода Европы». Природа. 534 (7606): 200–5. Bibcode:2016Натура.534..200F. Дои:10.1038 / природа17993. ЧВК 4943878. PMID 27135931.
- ^ Д'Атанасио Э., Тромбетта Б., Бонито М., Финоккио А., Ди Вито Г., Сегицци М. и др. (Февраль 2018). «Население последней Зеленой Сахары выявлено в результате переупорядочения транссахарских патрилиний с большим охватом». Геномная биология. 19 (1): 20. Дои:10.1186 / s13059-018-1393-5. ЧВК 5809971. PMID 29433568.
- ^ Маркус Дж. Х., Пост С., Рингбауэр Х., Лай Л., Скейтс Р., Сидор С. и др. (Февраль 2020 г.). «Генетическая история от среднего неолита до наших дней на средиземноморском острове Сардиния». Nature Communications. 11 (1): 939. Bibcode:2020NatCo..11..939M. Дои:10.1038 / s41467-020-14523-6. ЧВК 7039977. PMID 32094358.
- ^ а б c Cinniolu C, King R, Kivisild T, Kalfolu E, Atasoy S, Cavalleri GL, et al. (Январь 2004 г.). «Раскопки гаплотипов Y-хромосомы в Анатолии» (PDF). Генетика человека. 114 (2): 127–48. Дои:10.1007 / s00439-003-1031-4. PMID 14586639. S2CID 10763736. Архивировано из оригинал (PDF) 19 июня 2006 г.
- ^ а б "ISOGG 2016 Y-ДНК гаплогруппа R". isogg.org.
- ^ "R-V1636 YTree". www.yfull.com.
- ^ Гругни В., Батталья В., Хушиар Кашани Б., Пароло С., Аль-Захери Н., Ахилли А. и др. (18 июля 2012 г.). «Древние миграционные события на Ближнем Востоке: новые ключи к разгадке вариации Y-хромосомы современных иранцев». PLOS ONE. 7 (7): e41252. Bibcode:2012PLoSO ... 741252G. Дои:10.1371 / journal.pone.0041252. ЧВК 3399854. PMID 22815981.
- ^ Лхагвасурен Г., Шин Х., Ли С.Е., Тумэн Д., Ким Дж. Х., Ким К. Я. и др. (14 сентября 2016 г.). "Молекулярная генеалогия семьи монгольской царицы и ее возможное родство с Чингисханом". PLOS ONE. 11 (9): e0161622. Bibcode:2016PLoSO..1161622L. Дои:10.1371 / journal.pone.0161622. ЧВК 5023095. PMID 27627454.
- ^ Адамс С.М., Кинг Т.Э., Бош Э., Джоблинг М.А. (май 2006 г.). «Случай ненадежного SNP: повторяющаяся обратная мутация маркера Y-хромосомы P25 в результате преобразования гена». Международная криминалистическая экспертиза. 159 (1): 14–20. Дои:10.1016 / j.forsciint.2005.06.003. HDL:2381/443. PMID 16026953.
- ^ а б c Cruciani F, Trombetta B, Sellitto D, Massaia A, Destro-Bisol G, Watson E, et al. (Июль 2010 г.). «Гаплогруппа Y-хромосомы человека R-V88: отцовский генетический отчет о транссахарских связях в начале среднего голоцена и распространении чадских языков». Европейский журнал генетики человека. 18 (7): 800–7. Дои:10.1038 / ejhg.2009.231. ЧВК 2987365. PMID 20051990.
- ^ Варзари А (2006). «История популяции Днестровско-Карпатских гор: данные по внедрению Alu и полиморфизму Y-хромосомы» (PDF). Диссертация der Fakultät für Biologie der Ludwig-Maximilians-Universität München.
- ^ а б Малярчук Б., Деренко М., Денисова Г., Максимов А., Возняк М., Гржибовский Т. и др. (Август 2011 г.). «Древние связи между сибиряками и коренными американцами выявлены путем подтипа гаплогруппы Y-хромосомы Q1a». Журнал генетики человека. 56 (8): 583–8. Дои:10.1038 / jhg.2011.64. PMID 21677663. S2CID 12015336.
- ^ Dulik MC, Zhadanov SI, Osipova LP, Askapuli A, Gau L, Gokcumen O, et al. (Февраль 2012 г.). "Mitochondrial DNA and Y chromosome variation provides evidence for a recent common ancestry between Native Americans and Indigenous Altaians". Американский журнал генетики человека. 90 (2): 229–46. Дои:10.1016/j.ajhg.2011.12.014. ЧВК 3276666. PMID 22281367.
- ^ Behar DM, Yunusbayev B, Metspalu M, Metspalu E, Rosset S, Parik J, et al. (Июль 2010 г.). "The genome-wide structure of the Jewish people". Природа. 466 (7303): 238–42. Bibcode:2010Натура.466..238Б. Дои:10.1038 / природа09103. PMID 20531471. S2CID 4307824.
- ^ Sengupta S, Zhivotovsky LA, King R, Mehdi SQ, Edmonds CA, Chow CE, et al. (Февраль 2006 г.). «Полярность и временность распределений Y-хромосомы высокого разрешения в Индии позволяют идентифицировать как местные, так и экзогенные экспансии и выявить незначительное генетическое влияние среднеазиатских скотоводов». Американский журнал генетики человека. 78 (2): 202–21. Дои:10.1086/499411. ЧВК 1380230. PMID 16400607.
- ^ а б Di Cristofaro J, Pennarun E, Mazières S, Myres NM, Lin AA, Temori SA, et al. (2013). "Afghan Hindu Kush: where Eurasian sub-continent gene flows converge". PLOS ONE. 8 (10): e76748. Bibcode:2013PLoSO...876748D. Дои:10.1371/journal.pone.0076748. ЧВК 3799995. PMID 24204668.
- ^ Lippold S, Xu H, Ko A, Li M, Renaud G, Butthof A, et al. (2014). "Human paternal and maternal demographic histories: insights from high-resolution Y chromosome and mtDNA sequences". Следственная генетика. 5: 13. Дои:10.1186/2041-2223-5-13. ЧВК 4174254. PMID 25254093.
- ^ Karafet TM, Osipova LP, Savina OV, Hallmark B, Hammer MF (November 2018). "Siberian genetic diversity reveals complex origins of the Samoyedic-speaking populations". Американский журнал биологии человека. 30 (6): e23194. Дои:10.1002/ajhb.23194. PMID 30408262. S2CID 53238849.
- ^ Ashirbekov EE, Botbaev DM, Belkozhaev AM, Abayldaev AO, Neupokoeva AS, Mukhataev JE, et al. (2017). "Distribution of Y-Chromosome Haplogroups of the Kazakh from the South Kazakhstan, Zhambyl, and Almaty Regions". Reports of the National Academy of Sciences of the Republic of Kazakhstan. 6 (316): 85–95. ISSN 2224-5227.
- ^ Shuhu LI, Yilihamu NI, Bake RA, Bupatima AB, Matyusup DO (2018). "A study of genetic diversity of three isolated populations in Xinjiang using Y-SNP". Acta Anthropologica Sinica. 37 (1): 146–56. Дои:10.16359/j.cnki.cn11-1963/q.2017.0067.
- ^ R1b1a1a2 (R-M269) was previously R1b1a2, From 2003 to 2005, what is now R1b1a2 was designated R1b3. From 2005 to 2008, it was R1b1c. From 2008 to 2011, it was R1b1b2.
- ^ а б Balaresque P, Bowden GR, Adams SM, Leung HY, King TE, Rosser ZH, et al. (January 2010). Penny D (ed.). "A predominantly neolithic origin for European paternal lineages". PLOS Биология. 8 (1): e1000285. Дои:10.1371 / journal.pbio.1000285. ЧВК 2799514. PMID 20087410.
- ^ Haak W, Lazaridis I (February 10, 2015). "Massive migration from the steppe is a source for Indo-European languages in Europe". bioRxiv 10.1101/013433.
- ^ Arredi B, Poloni ES, Tyler-Smith C (2007). «Население Европы». In Crawford MH (ed.). Антропологическая генетика: теория, методы и приложения. Кембридж, Великобритания: Издательство Кембриджского университета. п. 394. ISBN 978-0-521-54697-3.
- ^ Cruciani F, Trombetta B, Antonelli C, Pascone R, Valesini G, Scalzi V, et al. (Июнь 2011 г.). "Strong intra- and inter-continental differentiation revealed by Y chromosome SNPs M269, U106 and U152". Международная криминалистическая экспертиза. Генетика. 5 (3): e49-52. Дои:10.1016/j.fsigen.2010.07.006. PMID 20732840.
- ^ а б Peter A. Underhill, Peidong Shen, Alice A. Lin и другие., "Y chromosome sequence variation and the history of human populations", Природа Генетика, Volume 26, November 2000
- ^ Lobov AS (2009). Structure of the Gene Pool of Bashkir Subpopulations (PDF) (Ph.D. thesis) (in Russian). Institute of Biochemistry and Genetics of the Ufa Scientific Center of the Russian Academy of Sciences. Архивировано из оригинал (PDF) на 16.08.2011.
- ^ Ordóñez AC, Fregel R, Trujillo-Mederos A, Hervella M, de-la-Rúa C, Arnay-de-la-Rosa M (2017). "Genetic studies on the prehispanic population buried in Punta Azul cave (El Hierro, Canary Islands)". Журнал археологической науки. 78: 20–28. Дои:10.1016 / j.jas.2016.11.004.
- ^ Robino C, Crobu F, Di Gaetano C, Bekada A, Benhamamouch S, Cerutti N, et al. (Май 2008 г.). "Analysis of Y-chromosomal SNP haplogroups and STR haplotypes in an Algerian population sample". Международный журнал судебной медицины. 122 (3): 251–5. Дои:10.1007 / s00414-007-0203-5. PMID 17909833. S2CID 11556974.
- ^ а б c Wood ET, Stover DA, Ehret C, Destro-Bisol G, Spedini G, McLeod H, et al. (Июль 2005 г.). «Контрастные модели вариаций Y-хромосомы и мтДНК в Африке: свидетельства предвзятых по полу демографических процессов» (PDF). Европейский журнал генетики человека. 13 (7): 867–76. Дои:10.1038 / sj.ejhg.5201408. PMID 15856073. S2CID 20279122. Архивировано из оригинал (PDF) 26 июня 2008 г.
- ^ Yepiskoposian L, Khudoyan A, Harutyunian A (2006). "Genetic Testing of Language Replacement Hypothesis in Southwest Asia". Иран и Кавказ. 10 (2): 191–208. Дои:10.1163/157338406780345899. JSTOR 4030922. S2CID 162345193.
- ^ Трофимова Натал'я Вадимовна (Feb. 2015), "Изменчивость Митохондриальной ДНК и Y-Хромосомы в Популяциях Волго-Уральского Региона" В архиве 2017-04-02 at the Wayback Machine ("Mitochondrial DNA variation and the Y-chromosome in the population of the Volga-Ural Region"). Автореферат. диссертации на соискание ученой степени кандидата биологических наук. Уфа – 2015.
- ^ Herrera KJ, Lowery RK, Hadden L, Calderon S, Chiou C, Yepiskoposyan L, et al. (Март 2012 г.). «Неолитические патрилинейные признаки указывают на то, что армянское нагорье было заселено земледельцами». Европейский журнал генетики человека. 20 (3): 313–20. Дои:10.1038 / ejhg.2011.192. ЧВК 3286660. PMID 22085901.
- ^ Vanek D, Saskova L, Koch H (June 2009). "Kinship and Y-chromosome analysis of 7th century human remains: novel DNA extraction and typing procedure for ancient material". Хорватский медицинский журнал. 3. 50 (3): 286–95. Дои:10.3325/cmj.2009.50.286. ЧВК 2702742. PMID 19480023.
- ^ Flores et. al. (2005) found that 20 out of all 146 men tested (13.7%) – including 20 out of 45 men tested from the Мертвое море area of Jordan – were positive for M173 (R1), and negative for both the R1a markers SRY10831.2 and M17, as well as P25 (which was later discovered to be an unreliable marker for R1b1), a study [1] indicates that they are mostly R-V88 (later known as R1b1a2). Wood et al. (2005) also reported two Egyptian cases of R1* (R-M173*) that were negative for SRY10831 (R1a1) and the unreliable R1b1 marker P25, out of a sample of 1,122 males from African countries, including 92 from Egypt. Hassan et al. (2008) found an equally surprising 14 out of 26 (54%) of Sudanese Народ фула who were M173+ and P25-
- ^ Флорес К., Мака-Мейер Н., Ларруга Дж. М., Кабрера В. М., Карадшех Н., Гонсалес А. М. (2005). "Isolates in a corridor of migrations: a high-resolution analysis of Y-chromosome variation in Jordan". Журнал генетики человека. 50 (9): 435–441. Дои:10.1007 / s10038-005-0274-4. PMID 16142507. S2CID 6490283.
- ^ Hassan HY, Underhill PA, Cavalli-Sforza LL, Ibrahim ME (November 2008). "Y-chromosome variation among Sudanese: restricted gene flow, concordance with language, geography, and history" (PDF). Американский журнал физической антропологии. 137 (3): 316–23. Дои:10.1002 / ajpa.20876. PMID 18618658. Архивировано из оригинал (PDF) на 2009-03-04.
13/32
- ^ Cruciani F, Santolamazza P, Shen P, Macaulay V, Moral P, Olckers A, et al. (Май 2002 г.). «Обратная миграция из Азии в Африку к югу от Сахары подтверждается анализом гаплотипов Y-хромосомы человека с высоким разрешением». Американский журнал генетики человека. 70 (5): 1197–214. Дои:10.1086/340257. ЧВК 447595. PMID 11910562., стр. 13–14
- ^ González M, Gomes V, López-Parra AM, Amorim A, Carracedo A, Sánchez-Diz P, et al. (Март 2013 г.). "The genetic landscape of Equatorial Guinea and the origin and migration routes of the Y chromosome haplogroup R-V88". Европейский журнал генетики человека. 21 (3): 324–31. Дои:10.1038/ejhg.2012.167. ЧВК 3573200. PMID 22892526.
- ^ Contu D, Morelli L, Santoni F, Foster JW, Francalacci P, Cucca F (January 2008). "Y-chromosome based evidence for pre-neolithic origin of the genetically homogeneous but diverse Sardinian population: inference for association scans". PLOS ONE. 3 (1): e1430. Bibcode:2008PLoSO ... 3.1430C. Дои:10.1371 / journal.pone.0001430. ЧВК 2174525. PMID 18183308.
- ^ Zalloua PA, Xue Y, Khalife J, Makhoul N, Debiane L, Platt DE, et al. (Апрель 2008 г.). "Y-chromosomal diversity in Lebanon is structured by recent historical events". Американский журнал генетики человека. 82 (4): 873–82. Дои:10.1016 / j.ajhg.2008.01.020. ЧВК 2427286. PMID 18374297.
- ^ Zhong H, Shi H, Qi XB, Duan ZY, Tan PP, Jin L, et al. (Январь 2011 г.). "Extended Y chromosome investigation suggests postglacial migrations of modern humans into East Asia via the northern route". Молекулярная биология и эволюция. 28 (1): 717–27. Дои:10.1093 / molbev / msq247. PMID 20837606.
- ^ Rowold DJ, Perez Benedico D, Garcia-Bertrand R, Chennakrishnaiah S, Alfonso-Sanchez MA, Gayden T, Herrera RJ (March 2016). "Ladakh, India: the land of high passes and genetic heterogeneity reveals a confluence of migrations". Европейский журнал генетики человека. 24 (3): 442–9. Дои:10.1038/ejhg.2015.80. ЧВК 4755386. PMID 25966630.
- ^ а б c "FamilyTreeDNA - R1b Basal Subclades".
- ^ Larmuseau MH, Delorme P, Germain P, Vanderheyden N, Gilissen A, Van Geystelen A, et al. (Май 2014 г.). "Genetic genealogy reveals true Y haplogroup of House of Bourbon contradicting recent identification of the presumed remains of two French Kings". Европейский журнал генетики человека. 22 (5): 681–7. Дои:10.1038/ejhg.2013.211. ЧВК 3992573. PMID 24105374.
- ^ Marks K (2010-02-04). "Darwin family DNA shows African origin". NZ Herald. ISSN 1170-0777. Получено 2020-07-16.
- ^ Мур Л.Т., Макэвой Б., Мыс Е., Симмс К., Брэдли Д.Г. (февраль 2006 г.). "A Y-chromosome signature of hegemony in Gaelic Ireland". Американский журнал генетики человека. 78 (2): 334–8. Дои:10.1086/500055. ЧВК 1380239. PMID 16358217.
- ^ Half of European men share King Tut's DNA.
- ^ "About R1a and R1b from Ural epic story. Artem Lukichev (c)" - через www.youtube.com.
- ^ Coble MD, Loreille OM, Wadhams MJ, Edson SM, Maynard K, Meyer CE, et al. (2009). "Mystery solved: the identification of the two missing Romanov children using DNA analysis". PLOS ONE. 4 (3): e4838. Bibcode:2009PLoSO ... 4.4838C. Дои:10.1371 / journal.pone.0004838. ЧВК 2652717. PMID 19277206.
- ^ Maan, A. A., Eales, J., Akbarov, A., et al. The Y chromosome: a blueprint for men’s health? . European Journal of Human Genetics, 25(11), 1181–1188. August 30, 2017. doi: https://doi.org/10.1038/ejhg.2017.128
- ^ Schillaci S (2020) Possible correlation between COVID-19 contagion and Y-DNA Haplogroup R1b. 10.31219/osf.io/yv8kc
Библиография
- Jones ER, Zarina G, Moiseyev V, Lightfoot E, Nigst PR, Manica A, et al. (February 2017). "The Neolithic Transition in the Baltic Was Not Driven by Admixture with Early European Farmers". Текущая биология. 27 (4): 576–582. Дои:10.1016/j.cub.2016.12.060. ЧВК 5321670. PMID 28162894.
- González-Fortes G, Jones ER, Lightfoot E, Bonsall C, Lazar C, Grandal-d'Anglade A, et al. (Июнь 2017 г.). "Paleogenomic Evidence for Multi-generational Mixing between Neolithic Farmers and Mesolithic Hunter-Gatherers in the Lower Danube Basin". Текущая биология. 27 (12): 1801–1810.e10. Дои:10.1016/j.cub.2017.05.023. ЧВК 5483232. PMID 28552360.
- Haak W, Lazaridis I, Patterson N, Rohland N, Mallick S, Llamas B, et al. (Июнь 2015 г.). «Массовая миграция из степи была источником индоевропейских языков в Европе». Природа. 522 (7555): 207–11. arXiv:1502.02783. Bibcode:2015Натура.522..207H. Дои:10.1038 / природа14317. ЧВК 5048219. PMID 25731166.
- Hollard C, Zvénigorosky V, Kovalev A, Kiryushin Y, Tishkin A, Lazaretov I, et al. (Сентябрь 2018 г.). "New genetic evidence of affinities and discontinuities between bronze age Siberian populations". Американский журнал физической антропологии. 167 (1): 97–107. Дои:10.1002/ajpa.23607. PMID 29900529.
- Lipson M, Szécsényi-Nagy A, Mallick S, Pósa A, Stégmár B, Keerl V, et al. (November 2017). «Параллельные палеогеномные трансекты раскрывают сложную генетическую историю ранних европейских фермеров». Природа. 551 (7680): 368–372. Bibcode:2017Натура.551..368L. Дои:10.1038 / природа24476. ЧВК 5973800. PMID 29144465.
- Marcus JH, Posth C, Ringbauer H, Lai L, Skeates R, Sidore C, et al. (February 2020). "Genetic history from the Middle Neolithic to present on the Mediterranean island of Sardinia". Nature Communications. 11 (1): 939. Bibcode:2020NatCo..11..939M. Дои:10.1038/s41467-020-14523-6. ЧВК 7039977. PMID 32094358.
- Mathieson I, Alpaslan-Roodenberg S, Posth C, Szécsényi-Nagy A, Rohland N, Mallick S, et al. (Март 2018 г.). «Геномная история Юго-Восточной Европы». Природа. 555 (7695): 197–203. Bibcode:2018Natur.555..197M. Дои:10.1038 / природа25778. ЧВК 6091220. PMID 29466330.
- Narasimhan VM, Patterson N, Moorjani P, Rohland N, Bernardos R, Mallick S, et al. (Сентябрь 2019 г.). «Формирование человеческих популяций в Южной и Центральной Азии». Наука. 365 (6457): eaat7487. Дои:10.1126 / science.aat7487. ЧВК 6822619. PMID 31488661.
- Olalde I, Brace S, Allentoft ME, Armit I, Kristiansen K, Booth T, et al. (Март 2018 г.). "The Beaker phenomenon and the genomic transformation of northwest Europe". Природа. 555 (7695): 190–196. Bibcode:2018Натура.555..190O. Дои:10.1038 / природа25738. ЧВК 5973796. PMID 29466337.
- Sánchez-Quinto F, Malmström H, Fraser M, Girdland-Flink L, Svensson EM, Simões LG, et al. (Май 2019). "Megalithic tombs in western and northern Neolithic Europe were linked to a kindred society". Труды Национальной академии наук Соединенных Штатов Америки. 116 (19): 9469–9474. Дои:10.1073/pnas.1818037116. ЧВК 6511028. PMID 30988179.
- Fu Q, Posth C, Hajdinjak M, Petr M, Mallick S, Fernandes D, et al. (Июнь 2016). «Генетическая история ледникового периода Европы». Природа. 534 (7606): 200–5. Bibcode:2016Натура.534..200F. Дои:10.1038 / природа17993. ЧВК 4943878. PMID 27135931.
внешняя ссылка
- R1b-YDNA discussion group Yahoo! Groups;