Гаплогруппа E-M215 (Y-ДНК) - Haplogroup E-M215 (Y-DNA)
| Гаплогруппа E-M215 | |
|---|---|
| Возможное время происхождения | 47 500 - 38 600 лет назад[1][2] |
| Возможное место происхождения | Северо-Восточная Африка или северный Африканский рог[1][2] |
| Предок | E-P2 |
| Потомки | E-M35, E-M281 |
| Определение мутаций | M215 |
E-M215, также известный как E1b1b и раньше E3b, является основным гаплогруппа ДНК Y-хромосомы человека. Это подразделение макро-гаплогруппа E-M96, который определяется однонуклеотидный полиморфизм (SNP) мутация M215.[3][4][5] Другими словами, это один из основных родословная из человечество, связывая от отца к сыну с общим предком по мужской линии ("Y-хромосомный Адам "). Это предмет обсуждения и изучения в генетика а также генетическая генеалогия, археология, и историческая лингвистика.
Гаплогруппа E-M215 имеет две древние ветви, которые содержат все известные современные субклады E-M215, E-M35 и E-M281. Из двух последних подгаплогрупп E-M215, единственная ветвь, которая была подтверждена у коренного населения за пределами Эфиопия это E-M35. E-M35, в свою очередь, имеет две известные ветви, E-V68 и E-Z827, которые содержат большинство всех современных носителей субклада E-M215. Субклады E-V68 и E-V257 были обнаружены на самых высоких частотах в Северная Африка и Африканский рог, а также при более низких процентах в части Средний Восток и Европа, а в изолированных популяциях Южная Африка.
Происхождение

Происхождение E-M215 было датировано Кручиани в 2007 году примерно 22 400 лет назад в Африканский рог.[6][Примечание 1] E-M35 был датирован Батини в 2015 году от 15 400 до 20 500 лет назад.[7] В июне 2015 года Тромбетта и др. сообщили о ранее недооцененной большой разнице в возрасте между гаплогруппой E-M215 (38,6 тыс. лет назад; 95% ДИ 31,4–45,9 тыс. лет назад) и ее подгаплогруппой E-M35 (25,0 тыс. лет назад; 95% ДИ 20,0–30,0 тыс. лет назад) и оценили ее происхождение. быть в Северо-Восточная Африка, где узел, разделяющий ветви E-V38 и E-M215, встречается около 47 500 лет назад (95% ДИ: 41,3–56,8 тыс. лет назад).[1]
Считается, что все основные ответвления E-M35 возникли в той же области, что и родительская клада: в Северная Африка, то Африканский рог, или близлежащие районы Ближний Восток. Предполагается, что некоторые ответвления E-M35 покинули Африку тысячи лет назад, тогда как другие могли прибыть с Ближнего Востока. Например, Андерхилл (2002) связывает распространение гаплогруппы с Неолитическая революция, полагая, что структура и региональный образец субкладов E-M35 потенциально дают «реагенты, с помощью которых можно сделать вывод о конкретных эпизодах истории популяции, связанных с Неолит расширение сельского хозяйства ". Battaglia et al. (2007) также подсчитали, что E-M78 (называемый в этой статье E1b1b1a1) находится в Европе более 10 000 лет. Соответственно, человеческие останки, раскопанные в испанской погребальной пещере, датируемые примерно 7000 лет назад, оказались в этой гаплогруппе.[11] Еще два E-M78 были обнаружены в эпоху неолита. Сопот и Lengyel культуры тоже.[12]
По поводу E-M35 в Европа в рамках этой схемы, Андерхилл и Кивисилд (2007) заметили, что E-M215, кажется, представляет собой поздно-Плейстоцен миграция из Северной Африки в Европу через Синайский полуостров в Египет.[Заметка 2] Хотя это предложение не оспаривается, недавно оно было предложено Trombetta et al. (2011) что есть также свидетельства дополнительной миграции E-M215, перевозящего людей прямо из Северной Африки в юго-западную Европу по морскому маршруту (см. ниже).
По данным Lazaridis et al. (2016), Натуфийский скелетные останки древних Левант преимущественно несли гаплогруппу Y-ДНК E1b1b. Из пяти натуфийских экземпляров, проанализированных на предмет отцовских линий, три принадлежали к E1b1b1b2 (xE1b1b1b2a, E1b1b1b2b), E1b1 (xE1b1a1, E1b1b1b1) и E1b1b1b2 (xE1b1b1b1b1b) sub) (E1b1b2b) 60% sub. Гаплогруппа E1b1b также обнаруживалась в умеренных количествах среди окаменелостей последующих поколений. Докерамический неолит B в культуре, причем субклады E1b1b1 и E1b1b1b2 (xE1b1b1b2a, E1b1b1b2b) наблюдались в двух из семи образцов PPNB (~ 29%). Ученые предполагают, что ранние фермеры из Леванта, возможно, распространились на юг, в Восточную Африку, принеся с собой наследственные компоненты Западной Евразии и Базальной Евразии, отдельные от тех, которые позже прибыли в Северную Африку.
Кроме того, гаплогруппа E1b1b1 была обнаружена в древнеегипетский мумия, раскопанная на Абусир эль-Мелек археологические раскопки в Среднем Египте, которые датируются периодом между поздним Новое Королевство и Римская эпоха.[13] Окаменелости на Иберомаврусский сайт Ифри н'Амр или Мусса в Марокко, которые были датированы примерно 5000 г. до н.э., также несли гаплотипы, относящиеся к субкладу E1b1b1b1a (E-M81). Эти древние люди несли автохтонный геномный компонент Магриби, который достигает пика среди современных Североафриканцы, что указывает на то, что они были предками населения в этом районе.[14] Гаплогруппа E1b1b также наблюдалась в древних Гуанчи окаменелости, раскопанные в Гран-Канария и Тенерифе на Канарские острова, которые были датированы радиоуглеродом между VII и XI веками нашей эры. Несущие кладу особи, которые были проанализированы на отцовскую ДНК, были захвачены на сайте Тенерифе, и все эти образцы, как было установлено, принадлежат к субкладу E1b1b1b1a1 или E-M183 (3/3; 100%).[15]
Loosdrecht et al. (2018) проанализировали полногеномные данные семи древних Иберомаврусский лица из Grotte des Pigeons возле Taforalt в восточном Марокко. Окаменелости были непосредственно датированы периодом между 15 100 и 13 900 калиброванными годами до настоящего времени. Ученые обнаружили, что пять мужских особей с достаточной сохранностью ядерной ДНК принадлежали к субкладу E1b1b1a1 (M78), причем один скелет нес родительскую линию E1b1b1a1b1 от E-V13, другой мужской образец принадлежал к E1b1b (M215 *).[16]
Распределение
В Африке E-M215 распространяется с самыми высокими частотами в Африканский рог и Северная Африка, откуда он за последние тысячелетия распространился на юг до Южная Африка, и на север в Западная Азия и Европа (особенно Средиземноморье и Балканы ).[8][17][18][19]
Почти все мужчины E-M215 также находятся в E-M35. В 2004 году было обнаружено, что M215 старше, чем M35, когда были обнаружены люди, которые имеют мутацию M215, но не имеют мутации M35.[8]В 2013 году один человек в Хорасане, Северо-Восточный Иран, был обнаружен Ди Кристофаро и др. (2013) быть положительным для M215, но отрицательным для M35.
E-M215 и E-M35 довольно распространены среди Афроазиатские спикеры. Лингвистическая группа и носители линии E-M35 с большой вероятностью возникли и рассеялись вместе из Афроазиатский урхеймат.[20][21][22] Среди населения с афро-азиатской говорящей историей значительная часть Еврейский мужские линии - E-M35.[23] Гаплогруппа E-M35, на долю которой приходится примерно 18%[17] до 20%[24][25] из Ашкенази и 8,6%[26] до 30%[17] из Сефарды Y-хромосомы, по-видимому, являются одной из основных линий еврейского населения.[27][Заметка 3]
Следующая таблица включает только выборочные популяции с более чем 1% мужчин E-M215 со всеми известными субкладами по состоянию на июнь 2015 года. Она содержит кладу E-V1515, определенную Trombetta et al. 2015, и все субклады E1b1b, распределенные ниже Сахары (E-V42, E-M293, E-V92, E-V6), которые были идентифицированы как базальные клады E-M35 в прежней филогении.[1]
| численность населения | N | Область, край | Язык | Итого E-M215 | E-V2009 | E-M78 * | E-V1477 | E-V1083 * | E-V13 | E-V22 | E-V12 * | E-V32 | E-V259 | E-V65 | E-V257 * | E-M81 | E-M123 * | E-M34 | E-V1515 * | E-V1486 * | E-V2881 * | E-V1792 | E-V92 | E-M293 * | E-V3065 | E-V42 | E-V1785 * | E-V6 | E-V16 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Северная Африка | |||||||||||||||||||||||||||||
| Марокканские арабы | 55 | Марокко | AA / семитский | 70.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 7.3 | 0.0 | 0.0 | 0.0 | 32.7 | 0.0 | 30.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Асни Берберс | 54 | Марокко | AA / берберский | 85.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1.9 | 79.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Бухрия Берберс | 67 | Марокко | AA / берберский | 79.1 | 0.0 | 0.0 | 0.0 | 0.0 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 77.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Берберы Среднего Атласа | 69 | Марокко | AA / берберский | 81.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 10.1 | 0.0 | 71.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Марракешские берберы | 27 | Марокко | AA / берберский | 92.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.7 | 3.7 | 0.0 | 0.0 | 0.0 | 3.7 | 77.8 | 0.0 | 3.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Souss Berbers | 34 | Марокко | AA / берберский | 79.4 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 76.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Уарзазатские берберы | 31 | Марокко | AA / берберский | 54.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 54.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Мозабитские берберы | 67 | Алжир | AA / берберский | 89.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.5 | 0.0 | 86.6 | 0.0 | 1.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Тунисские евреи | 10 | Тунис | AA / семитский | 20.0 | 0.0 | 0.0 | 10.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 10.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ливийские арабы | 10 | Ливия | AA / семитский | 50.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 30.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ливийские евреи | 23 | Ливия | AA / семитский | 26.1 | 0.0 | 0.0 | 0.0 | 0.0 | 4.3 | 0.0 | 0.0 | 0.0 | 0.0 | 4.3 | 0.0 | 0.0 | 0.0 | 17.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Северные египтяне | 49 | Египет | AA / семитский | 42.9 | 0.0 | 0.0 | 0.0 | 0.0 | 2.0 | 16.3 | 4.1 | 2.0 | 0.0 | 0.0 | 0.0 | 4.1 | 4.1 | 10.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Египетские берберы из Сивы | 93 | Египет | AA / семитский | 18.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.2 | 0.0 | 0.0 | 4.3 | 2.2 | 1.1 | 0.0 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.5 | 0.0 |
| Египтяне из Бахарии | 41 | Египет | AA / семитский | 56.1 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 22.0 | 14.6 | 0.0 | 0.0 | 2.4 | 7.3 | 4.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 |
| Египтяне из оазиса Гурна | 34 | Египет | AA / семитский | 17.6 | 0.0 | 5.9 | 0.0 | 0.0 | 0.0 | 0.0 | 8.8 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Южные египтяне | 47 | Египет | AA / семитский | 78.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 74.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.1 | 2.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Западная / Центральная Африка | |||||||||||||||||||||||||||||
| Манденка | 16 | Сенегал | NC / Mande | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Туареги | 22 | Нигер | AA / берберский | 13.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.5 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Даба | 29 | Камерун (Север) | AA / Чад | 3.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Guidar | 9 | Камерун (Север) | AA / Чад | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Мандара | 82 | Камерун (Север) | AA / Чад | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Шува арабы | 5 | Камерун (Север) | AA / семитский | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Фульбе из Камеруна | 76 | Камерун (Север) | Северная Каролина / Атлантика | 1.3 | 1.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Moundang | 21 | Камерун (Север) | NC / Adamawa | 4.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Восточная африка | |||||||||||||||||||||||||||||
| Тигре | 5 | Эритрея | AA / семитский | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 60.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 |
| Нара | 15 | Эритрея | NS / суданский | 60.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.7 | 0.0 | 13.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.3 | 6.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.3 | 0.0 | 6.7 | 0.0 |
| Кунама | 20 | Эритрея | NS / Cunama | 65.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.0 | 0.0 | 0.0 | 5.0 | 0.0 | 0.0 | 0.0 | 0.0 | 15.0 | 10.0 | 5.0 | 0.0 |
| Сахо | 94 | Эритрея | AA / Cushitic | 98.9 | 0.0 | 0.0 | 0.0 | 1.1 | 0.0 | 88.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.5 | 0.0 |
| Tigrai | 32 | Эритрея / Эфиопия | AA / семитский | 71.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.1 | 3.1 | 21.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.1 | 3.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 31.3 | 6.3 | 0.0 |
| Афар | 25 | Джибути | AA / Cushitic | 60.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 56.0 | 0.0 |
| Сомалийский | 40 | Джибути | AA / Cushitic | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Эфиопские евреи | 22 | Эфиопия | AA / Cushitic | 31.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 | 0.0 | 0.0 |
| Амхара | 82 | Эфиопия | AA / семитский | 45.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 | 11.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 13.4 | 0.0 | 0.0 | 2.4 | 0.0 | 1.2 | 0.0 | 0.0 | 1.2 | 0.0 | 8.5 | 4.9 |
| Оромо | 62 | Эфиопия | AA / Cushitic | 53.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 22.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.8 | 0.0 | 0.0 | 17.7 | 0.0 | 0.0 | 1.6 | 0.0 | 3.2 | 0.0 | 1.6 | 1.6 |
| Волайта | 12 | Эфиопия | AA / Omotic | 58.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.3 | 0.0 | 8.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.3 | 0.0 | 0.0 | 8.3 | 0.0 | 0.0 | 8.3 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 |
| Сомалийский | 12 | Эфиопия | AA / Cushitic | 50.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.3 | 0.0 | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Gurage | 7 | Эфиопия | AA / семитский | 42.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 28.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 14.3 | 0.0 |
| Сомалийский | 5 | Сомали | AA / Cushitic | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 80.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Туркана | 6 | Кения | NS / суданский | 50.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 33.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Борана | 9 | Кения | AA / Cushitic | 77.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 66.7 | 0.0 | 0.0 | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Сомалийский | 6 | Кения | AA / Cushitic | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 66.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 16.7 | 0.0 |
| Нилотическая Западная Кения | 11 | Кения | NS / суданский | 45.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.1 | 9.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 18.2 | 0.0 | 0.0 | 0.0 | 9.1 | 0.0 |
| Лухья | 51 | Кения | Северная Каролина / Банту | 9.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Другое банту | 17 | Кения | Северная Каролина / Банту | 11.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 11.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Кикую | 9 | Кения | Северная Каролина / Банту | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Масаи | 45 | Кения | NS / суданский | 37.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.7 | 0.0 | 6.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 24.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Тутси | 9 | Бурунди | Северная Каролина / Банту | 22.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 22.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Южная Африка | |||||||||||||||||||||||||||||
| ! Кунг | 64 | Ангола | KS | 10.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 9.4 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 |
| Кхве | 26 | Намибия | KS | 30.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 30.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| банту | 8 | Южная Африка | Северная Каролина / Банту | 12.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 12.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Европа | |||||||||||||||||||||||||||||
| Северный португальский | 50 | Португалия | IE | 10.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 2.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Южный португальский | 49 | Португалия | IE | 16.3 | 0.0 | 0.0 | 0.0 | 0.0 | 4.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 12.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Пасиегос из Кантабрии | 56 | Испания | IE | 42.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.8 | 41.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Астурийцы | 90 | Испания | IE | 12.2 | 0.0 | 0.0 | 0.0 | 0.0 | 5.6 | 4.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Южные испанцы | 62 | Испания | IE | 6.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.2 | 0.0 | 0.0 | 0.0 | 0.0 | 1.6 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Испанские баски | 55 | Испания | Баскский | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Французский | 85 | Франция | IE | 8.2 | 0.0 | 0.0 | 0.0 | 0.0 | 3.5 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 0.0 | 3.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Французские баски | 16 | Франция | Баскский | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 6.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Корсиканцы | 140 | Франция | IE | 6.4 | 0.0 | 0.0 | 0.0 | 0.0 | 4.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.7 | 0.0 | 0.0 | 1.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Датский | 35 | Дания | IE | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Немцы | 77 | Германия | IE | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Северные итальянцы | 80 | Италия | IE | 11.3 | 0.0 | 0.0 | 0.0 | 0.0 | 6.3 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.3 | 0.0 | 1.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Центральные итальянцы | 356 | Италия | IE | 12.9 | 0.0 | 0.0 | 0.0 | 0.0 | 5.3 | 2.0 | 0.3 | 0.0 | 0.0 | 0.3 | 0.3 | 0.8 | 0.0 | 3.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Южные итальянцы | 141 | Италия | IE | 15.6 | 0.7 | 0.0 | 0.0 | 0.0 | 8.5 | 1.4 | 0.7 | 0.0 | 0.0 | 0.0 | 0.0 | 1.4 | 0.0 | 2.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Сицилийцы | 153 | Италия | IE | 20.3 | 0.0 | 0.0 | 0.0 | 0.0 | 7.2 | 4.6 | 0.7 | 0.0 | 0.0 | 0.7 | 0.0 | 0.7 | 0.0 | 6.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Сардинцы | 374 | Италия | IE | 8.3 | 0.8 | 0.0 | 0.0 | 0.3 | 1.1 | 0.8 | 0.3 | 0.0 | 0.0 | 1.1 | 0.3 | 0.3 | 0.0 | 3.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Польский | 40 | Польша | IE | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Словенцы | 104 | Словения | IE | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Эстонцы | 74 | Эстония | U | 5.4 | 0.0 | 0.0 | 0.0 | 0.0 | 4.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Венгры | 106 | Венгрия | U | 10.4 | 0.0 | 0.0 | 0.0 | 0.0 | 9.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Румыны | 30 | Румыния | IE | 26.7 | 0.0 | 0.0 | 0.0 | 0.0 | 26.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Македонцы | 99 | Македония | IE | 18.2 | 0.0 | 0.0 | 0.0 | 0.0 | 18.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Континентальные греки | 32 | Греция | IE | 28.1 | 0.0 | 0.0 | 0.0 | 0.0 | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Болгары | 112 | Болгария | IE | 22.3 | 0.0 | 0.0 | 0.0 | 0.0 | 21.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Сефардские болгары | 20 | Болгария | IE | 5.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Албанцы | 21 | Албания | IE | 33.3 | 0.0 | 0.0 | 0.0 | 0.0 | 33.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ближний Восток | |||||||||||||||||||||||||||||
| Сефардский турецкий | 19 | индюк | А | 10.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.3 | 0.0 | 5.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Стамбул Турецкий | 35 | индюк | А | 17.1 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 5.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5.7 | 0.0 | 2.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Юго-западный турецкий | 40 | индюк | А | 7.5 | 0.0 | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.5 | 0.0 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Северо-Восточный Турецкий | 41 | индюк | А | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Центральная Анатолия | 61 | индюк | А | 9.8 | 0.0 | 0.0 | 0.0 | 0.0 | 4.9 | 0.0 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Юго-Восточный Турецкий | 24 | индюк | А | 8.3 | 0.0 | 0.0 | 0.0 | 0.0 | 4.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Эрзурум Турецкий | 25 | индюк | А | 12.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Турки-киприоты | 46 | индюк | А | 23.9 | 0.0 | 0.0 | 0.0 | 0.0 | 10.9 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 8.7 | 0.0 | 2.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Бедуины | 28 | Израиль | AA / семитский | 14.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 7.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Друзы арабы | 28 | Израиль | AA / семитский | 14.3 | 0.0 | 0.0 | 0.0 | 0.0 | 10.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Палестинцы | 29 | Израиль | AA / семитский | 13.8 | 0.0 | 0.0 | 0.0 | 0.0 | 3.4 | 6.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Арабов | 41 | Объединенные Арабские Эмират | AA / семитский | 7.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 4.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Оманит | 13 | Оман | AA / семитский | 15.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 7.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 7.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Йеменцы | 94 | Йемен | AA / семитский | 14.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.1 | 3.2 | 0.0 | 0.0 | 0.0 | 1.1 | 0.0 | 7.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.1 |
E-M215 ассоциация с выносливостью
Moran et al. (2004) наблюдали, что среди групп Y-ДНК (отцовской), принадлежащих элитным спортсменам на выносливость в Эфиопии, гаплогруппа E3b1 отрицательно коррелировала с показателями элитных спортсменов на выносливость,[28] тогда как гаплогруппы E *, E3 *, К * (xP),[28] и J * (xJ2) были значительно чаще среди элитных спортсменов на выносливость.[28]
Субклады E-M215
Семейное древо
Следующее филогенетическое дерево основан на дереве YCC 2008 и последующих опубликованных исследованиях, резюмированных ISOGG. Он включает все известные субклады по состоянию на июнь 2015 г. (Trombetta et al.2015)[29][4][5]
- E-M215 (E1b1b)
- E-M215 *. Редко или не существует.
- E-M35 (E1b1b1)
- E-V68 (E1b1b1a)
- E-V2009. Встречается у людей на Сардинии и Марокко.
- E-M78 (E1b1b1a1). Северная Африка, Африканский Рог, Западная Азия, Сицилия. (Ранее «E1b1b1a».)
- E-M78 *
- E-V1477. Встречается у тунисских евреев.
- E-V1083.
- E-V1083 *. Встречается только в Эритрее (1,1%) и Сардинии (0,3%).
- E-V13
- E-V22
- E-V1129
- E-V12
- E-V12 *
- E-V32
- E-V264
- E-V259. Найден в Северном Камеруне.
- E-V65
- E-CTS194
- E-V12
- E-Z827 (E1b1b1b)[30]
- E-V257 / L19 (L19, V257) - E1b1b1b1[30]
- E-PF2431
- E-M81 (M81)
- E-PF2546
- E-PF2546 *
- E-CTS12227
- E-MZ11
- E-MZ12
- E-MZ11
- E-A929
- E-Z5009
- E-Z5009 *
- E-Z5010
- E-Z5013
- E-Z5013 *
- E-A1152
- E-A2227
- E-A428
- E-MZ16
- E-PF6794
- E-PF6794 *
- E-PF6789
- E-MZ21
- E-MZ23
- E-MZ80
- E-A930
- E-Z2198 / E-MZ46
- E-A601
- E-L351
- E-Z5009
- E-PF2546
- E-Z830 (Z830) - E1b1b1b2[30]
- E-M123 (M123)
- E-M34 (M34)
- E-M84 (M84)
- E-M136 (M136)
- E-M290 (M290)
- E-V23 (V23)
- E-L791 (L791, L792)
- E-M84 (M84)
- E-M34 (M34)
- E-V1515. E-V1515 и его субклады в основном ограничены Восточной Африкой.
- E-V1515 *
- E-V1486
- E-V1486 *
- E-V2881
- E-V2881 *
- E-V1792
- E-V92
- E-M293 (M293)
- E-M293 *
- E-P72 (P72)
- E-V3065 *
- E-V1700
- E-V42 (V42)
- E-V1785
- E-V1785 *
- E-V6 (V6)
- E-M123 (M123)
- E-V257 / L19 (L19, V257) - E1b1b1b1[30]
- E-V16 / E-M281 (E1b1b2). Редкий. Встречается у людей в Эфиопии, Йемене и Саудовской Аравии.
- E-V68 (E1b1b1a)
Исключительные случаи мужчин с положительным M215 и отрицательным M35 («E-M215 *») были обнаружены в двух Амхарас Эфиопии и один Йеменцы.[8][31] По крайней мере, некоторые из этих людей, а может быть, и все, известны с начала 2011 года как принадлежащие к редкой родственной группе E-M35, известной как E-V16 или E-M281.[32] Об открытии M281 объявили Semino et al. 2002 г., который нашел его в двух эфиопских Оромо. Trombetta et al. 2011 г. нашли еще 5 эфиопов и аналогичный SNP для M281, V16. Именно в статье 2011 года положение генеалогического дерева (M215 + / M35-) было обнаружено, как описано выше. Производная E-M215, E-M35, определяется SNP M35. 1 туркменский человек из Jawzjan с мутацией, определяющей подклад, обозначается как E-M35 *.[33] По состоянию на июнь 2015 года существует все более сложная древовидная структура, которая делит большинство мужчин в E-M35 на две ветви: E-V68 и E-Z827.
Наиболее часто описываемые субклады - E-M78, часть E-V68, и E-M81, который является веткой E-Z827. Эти два субклада представляют собой самую большую долю современного населения E-M215. E-M78 встречается в большей части ареала, где E-M215 встречается, за исключением Южной Африки. E-M81 встречается в основном в Северной Африке. E-M123 менее распространен, но широко рассредоточен со значительными популяциями в определенных частях Африканского Рога, Леванта, Аравии, Иберии и Анатолии. Новая клада (E-V1515) была определена Trombetta et al. 2015 г., который возник около 12 тыс. Лет назад (95% ДИ 8,6–16,4) в Восточной Африке, где он в настоящее время в основном распространен. Эта клада включает субклады E-V42, E-M293, E-V92 и E-V6, которые были идентифицированы как базальные клады E-M35 в предыдущей филогении.[1]
В E-M35 есть поразительные параллели между двумя гаплогруппами, E-V68 и E-V257. Оба содержат линию, которая часто наблюдалась в Африке (E-M78 и E-M81, соответственно), и группу недифференцированных хромосом, которые в основном встречаются в южной Европе. Расширение авианосцев E-M35, возможно, с Ближнего Востока, как предлагалось другими авторами, и разделение на две ветви, разделенные географическим барьером Средиземного моря, могло бы объяснить эту географическую закономерность. Однако отсутствие E-V68 * и E-V257 * на Ближнем Востоке делает морское распространение между Северной Африкой и Южной Европой более правдоподобной гипотезой.
TMRCA основных узлов в E-M215
| TMRCA (тыс. Шт.) | Тромбетта 2015 | YПолный |
| E-M215 | 39 | 35,4 |
| * E-M35 | 25 | 23,9 |
| ** E-V68 | 20 | 20 |
| *** E-M78 | 15 | 13 |
| ** E-Z827 | ? | 23,6 |
| *** E-V257 / L19 | ? | 13,9 |
| **** E-M81 | ? | 2,7 |
| *** E-Z830 | 20 | 19 |
| **** E-M34 | ? | 15 |
| **** E-V1515 | 19 | ? |
E-V68 (E1b1b1a)
E-V68, преобладает его более известный субклад E-M78. Три человека "E-V68 *", которые находятся в E-V68, но не E-M78, были зарегистрированы в Сардиния, к Trombetta et al. 2011 г., анонсируя открытие V68. Авторы отметили, что, поскольку E-V68 * не был обнаружен в образцах с Ближнего Востока, это, по-видимому, свидетельствует о морской миграции из Африки в юго-западную Европу.E-M78 - часто встречающийся субклад, широко распространенный в Северная Африка, то Африканский рог, Западная Азия, (Ближний Восток и Ближний Восток) «до Южной Азии»,[6] и все Европа.[34] В европейском распределении пик частоты сосредоточен в частях Балканы (в некоторых регионах почти до 50%)[17][35] и Сицилия, и снижение частот очевидно в сторону Западной, Центральной и Северо-Восточной Европы.
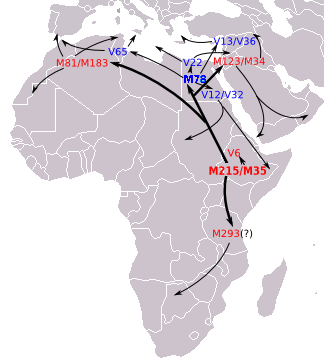
На основе генетических STR данные о дисперсии, Cruciani et al. 2007 г. предполагает, что E-M78 возник в районе Египет и Ливия.[Примечание 4] около 18 600 лет назад (17 300 - 20 000 лет назад).[Примечание 5] Battaglia et al. 2008 г. описывают Египет как «центр распределения различных географически локализованных субкладов, связанных с M78», и, основываясь на археологических данных, они предполагают, что точка происхождения E-M78 (в отличие от более позднего распространения из Египта), возможно, находилась в а рефугиум которые «существовали на рубеже современных Судан и Египет, рядом Озеро Нубия, до начала влажной фазы около 8500 г. до н.э. Сдвигающиеся на север дождевые пояса в этот период могли также стимулировать быструю миграцию Мезолит собирателей на север в Африке, Левант и, в конечном итоге, к Малая Азия и Европа, где каждая из них в конечном итоге разделилась на свои региональные отличительные ветви ". На юг, Hassan et al. 2008 г. также объясните доказательства того, что некоторые субклады E-M78, в частности E-V12 и E-V22, «могли быть занесены в Судан из Северной Африки после прогрессирующего опустынивания Сахары около 6000–8000 лет назад». И аналогично, Cruciani et al. 2007 г. предполагают, что E-M78 в Эфиопии, Сомали и прилегающих районах мигрировал обратно в этот регион со стороны Египта после приобретения мутации E-M78.
Недавно E-M78 был датирован Trombetta et al. 2015. между 20 300 и 14 800 лет назад.[1]
Субклады E-M78
Здесь перечислены основные субклады M78 по состоянию на июнь 2015 года. В субкладе E-M78, Trombetta et al. В 2015 году большая часть бывших хромосом E-M78 * была распределена по трем новым разным ветвям: E-V1083 *, E-V1477 и E-V259. Первый - сестра парагруппы кладов E-V22 и E-V13. Мутация V1477 определяет новую базальную ветвь, которая наблюдалась только в одном образце из Северной Африки. Наконец, родственная клада E-V12, определенная V264, включает E-V65 и V259, новую ветвь, распространенную в Центральной Африке.[1][34]
- E-M78 (E1b1b1a1) Северная Африка, Африканский Рог, Западная Азия, Европа (ранее «E1b1b1a»).
- E-M78 * Встречается в Марокко, южной Португалии, южной Испании и Иране (провинции Тегеран и Семнан).
- E-V1477 Встречается у тунисских евреев.
- E-V1083
- E-V1083 * Встречается только в Эритрее (1,1%) и Сардинии (0,3%).
- E-V13 Это наиболее распространенный субклад E-M215, встречающийся в Европе. Это особенно распространено на Балканах.
- E-V22. Сосредоточен в Северо-Восточной Африке и на Ближнем Востоке. Пики среди сахо.
- E-V1129
- E-V12. Встречается в Египте, Судане и других местах. Имеет важный подклад
- E-V12 * Наиболее распространенная линия среди южных египтян (74,5%).
- E-V32. Очень распространен среди сомалийцев, тигре и оромос.
- E-V264
- E-V259 Найден в Северном Камеруне.
- E-V65 Связан с Северной Африкой, но также встречается на Сицилии, а также в континентальной Италии.
- E-V12. Встречается в Египте, Судане и других местах. Имеет важный подклад
- E-M521 Не упоминается Тромбеттой и др., 2015. Найден у двух человек в Греции Battaglia et al. 2008 г. и у одного человека из Восточно-Альпийского региона Италии Coia et al. (2013)
E-Z827 (E1b1b1b)
В генетике человека E-Z827, это имя крупного гаплогруппа ДНК Y-хромосомы человека в изобилии встречается в Северной Африке, особенно Магриб, и в меньшей степени в Африканский рог, то Ближний Восток и Европа.
E-V257 / E-L19 (E1b1b1b1)
E-V257 * люди в их выборках, которые были E-V257, но не E-M81. Борана из Кении, марракешский бербер, корсиканец, сардинец, южный испанец и кантабриец. Как уже упоминалось выше, Trombetta et al. 2011 г. предполагают, что отсутствие E-V257 * на Ближнем Востоке (Ифулл нашел молодого в Иранский Азербайджан[36] и другой молодой в Армении[37]) делает морское перемещение из северной Африки в южную Европу наиболее правдоподобной гипотезой для объяснения его распространения. Yfull перечисляет 24 человека, все из которых принадлежат к одной ветви, которая на 30% моложе их общего предка с M81.[38]
E-M81
E-M81 является наиболее распространенным субкладом гаплогруппы E-L19 / V257. Он сосредоточен в Магриб, и преобладает его субклад E-M183. Считается, что E-M183 возник в Северо-Западной Африке и имеет предполагаемый возраст 4700 лн.[39]
Эта гаплогруппа достигает средней частоты 61% в Магриб и 51% в Северная Африка, частота снижается примерно с 80% до 100% в популяциях берберов,[40][41] включая сахарцев, примерно до 29% к востоку от этого диапазона в Египте.[42][43][44][45] Из-за своего молодого возраста и распространенности среди этих групп, а также других, таких как Mozabite, Средний Атлас, Kabyle и другие берберский групп, это иногда называют генетическим «берберским маркером». Pereira et al. 2010 г. сообщать о высоких уровнях среди Туареги в двоем Сахара населения - 77,8% около Гором-Гором, в Буркина-Фасо, и 81,8% от Гози в Мали. Была гораздо более низкая частота 11,1% в непосредственной близости от Танут в Нигер. E-M81 также довольно распространен среди Магрибский арабский говорящие группы. Обычно он встречается с частотой около 45% в прибрежных городах Алжир и Тунис (Джиджель, Оран, Тизи Узу, Алжир, Тунис, Сусс ).[42][46]
В этом ключевом районе от Египта до Атлантический океан, Sole-Morata et al. 2017 г. сообщить о паттерне M183-SM001 уменьшения микроспутник вариация гаплотипа (подразумевающая больший возраст родословной в прежних областях) от Племя регибат в Оран и они нашли M183 * (не SM001) в Иберии, Ливии и Марокко. Arredi et al. 2004 г. однако, микросателлитные вариации уменьшаются с востока на запад, что сопровождается значительным увеличением частоты. На восточной окраине этого основного диапазона Куянова и др. 2009 г. обнаружил M81 у 28,6% (10 из 35 мужчин) в Эль-Хайес в Ливийская пустыня в Египте.
Arredi et al. 2004 г. считают, что модель распределения и дисперсии согласуется с гипотезой "демическая диффузия "с Востока. Автохтонного присутствия E-M81 на Ближнем Востоке нет (есть в Ливане.[47]), что указывает на то, что M81, скорее всего, произошел от своей родительской клады M35 либо в Северной Африке, либо, возможно, на юге, до Африканского Рога.[22]
В Европе E-M81 широко распространен, но редко, в Пиренейский полуостров показывает среднюю частоту 4% (45/1140) на Пиренейском полуострове с частотами, достигающими 3,5% в Галиция, 4% в Западной Андалусия и Северо-Запад Кастилия. Однако в это исследование включены 153 человека с островов Майорка, Менорка и Ибица, а также 24 человека из Гаскони, которые не находятся на Пиренейском полуострове. Без этих 177 особей средний показатель для Пиренейского полуострова составляет 4,1% (40/963),[48] он находится на уровне, сопоставимом с E-M78, со средней частотой около 5%. Его частоты выше в западной половине полуострова и достигают 8% в Эстремадура и южной Португалии, от 4% до 5% в Галиция, 5% в западных Андалусия и 4% на северо-западе Кастилия и от 9% до 17% в Кантабрия.[26][49][50][51][52] Самые высокие частоты этой клады, обнаруженные до сих пор в Европе, наблюдались в Valles Pasiegos из Кантабрия от 5,5% (8/45)[52] до 41% (23/56).[8] Средняя частота 8,28% (54/652) также отмечена на испанском языке. Канарские острова с частотами более 10% на трех крупнейших островах Тенерифе (10.68%), Гран-Канария (11,54%) и Фуэртевентура (13.33%).[53]
E-M81 также встречается в Франция,[8] 2,70% (15/555) в целом с частотами выше 5% в Овернь (5/89) и Иль-де-Франс (5/91),[54][55] в Сицилия (примерно 9% в целом, но до 14% в Пьяцца Армерина ),[56] и с гораздо меньшей частотой около Lucera (2,7%), в континентальной Италия,[51] возможно, из-за древних миграций во время Исламский, Римский, и Карфагенский империи. В исследовании 2014 года Стефании Сарно и соавт. с 326 образцами из Козенца, Реджо-ди-Калабрия, Лечче и 5 провинциях Сиклии, E-M81 показывает среднюю частоту 6,5%, но типичный гаплотип ядра Магрибина 13-14-30-24-9-11-13 был обнаружен только у двух из пяти индивидуумов E-M81. Эти результаты, наряду с незначительным вкладом североафриканских популяций, выявленным анализом графиков, подобных примеси, предполагают лишь незначительное влияние транс-средиземноморских потоков генов на текущий генетический пул SSI.[56][57]
E-M81 также был обнаружен в 2013 году на уровне 5,8% в большой выборке из 1204 человек. Сардинцы.[58]
В результате своего распространения в старом мире этот субклад встречается повсюду. Бразилия[59] 5,4% в Бразилия (Рио де Жанейро),[Примечание 6] и среди Латиноамериканец мужчины из Калифорния и Гавайи 2.4%.[60]
В меньшем количестве мужчин E-M81 можно встретить в районах, соприкасающихся с [Северной Африкой, как вокруг Сахары, так и в таких местах, как Судан, и вокруг Средиземного моря в таких местах, как Ливан, индюк, и среди Сефардские евреи.
Есть два признанных субклада E-M81, хотя один гораздо важнее другого.[который? ]
Субклад E-M81 был обнаружен в древних Гуанчи (Bimbapes) окаменелости, раскопанные в Пунта-Асуль, Эль Йерро, Канарские острова, которые датируются X веком (~ 44%).[61]
E-M107
Андерхилл 2000 нашел один образец E-M107 в Мали.
E-M183
E-M183 чрезвычайно доминирует (более 99%[62]) внутри E-M81. Карафет и др. 2008 г. впервые описал его как субклад E-M81. Известные субклады E-M183 включают:
- E-M165 Андерхилл и др. 2000 г. нашел один пример на Ближнем Востоке.
- E-L351 Найдено у двух связанных участников E-M35 Филогенетический проект.
E-Z830 (E1b1b1b2)
Это недавно обнаруженный субклад, который еще не включен в большинство деревьев гаплогрупп, E-Z830 включает подтвержденные субклады E-M123, E-V1515 (E-M293, E-V42, E-V6, E-V92), и E-Z830 * и является родственным классом E-L19. В настоящее время, [постоянная мертвая ссылка ] филогенетический проект E-M35 распознает четыре отдельных кластера носителей Z830 *, два из которых имеют исключительно еврейское происхождение. Остальные два значительно меньше по размеру и включают отдельных особей в Германии, Испании, Латинской Америке, Египте и Эфиопии.[63][64][65][66]
E-M123
E-M123 в основном известен своей основной субкладой E-M34, которая доминирует над этой кладой.[Примечание 7]
E-V1515
Новая клада (E-V1515) была определена Trombetta et al. 2015 г., который возник около 12 тыс. Лет назад (95% ДИ 8,6–16,4) в Восточной Африке, где он в настоящее время в основном распространен. В эту кладу входят все гаплогруппы к югу от Сахары (E-V42, E-M293, E-V92, E-V6), о которых в предыдущей филогении сообщалось как базальные клады E-M35.[1]
E-M293
E-M293 является субкладом E-V1515. Впервые он был идентифицирован ISOGG как второй класс в E-Z830. Он был обнаружен до E-Z830, анонсирован в Хенн 2008, что связывало его с распространением скотоводство из Восточная Африка в Южная Африка. До сих пор высокие уровни были обнаружены у определенных этнических групп в Танзании и Южной Африке. Высшие были Datooga (43%), Кхве (Kxoe) (31%), Burunge (28%), и Народы сандаве (24%). Хенн (2008) в своем исследовании также обнаружил двух кенийских мужчин, говорящих на банту, с мутацией M293.[9]
Другие субклады E-M215 редки в Южной Африке. Авторы заявляют: «Без информации о M293 у масаи, хема и других популяций в Кении, Судане и Эфиопии мы не можем с большей уверенностью определить точный географический источник M293. Однако имеющиеся данные указывают на современную Танзанию как ранний и важный географический локус эволюции M293. ". Они также говорят, что «M293 находится только в Африке к югу от Сахары, что указывает на отдельную филогенетическую историю для образцов M35.1 * (бывших) дальше на север».
E-P72 появляется в Карафете (2008). Trombetta et al. 2011 г. объявил, что это субклад E-M293.
E-V42
Trombetta et al. 2011 г. объявил об открытии E-V42 в двух Бета Израиль человек. Было высказано предположение, что это может быть ограничено регионом вокруг Эфиопии. Однако дальнейшие испытания, проведенные коммерческими компаниями по тестированию ДНК, подтвердили множество положительных результатов для этого субклада в Саудовской Аравии, Кувейте и одного человека в Португалии, имеющего корень из Аравии.[67]
E-V6
Подклад E-V6 для E-V1515 определяется V6. Cruciani et al. (2004) идентифицировали значительное присутствие этих линий в Эфиопии, а также некоторых в соседних Сомалийцы. Среди образцов Эфиопии и Сомали самые высокие показатели были 14,7% среди амхарцев и 16,7% среди Волайта.
На юг, Тишкофф и др. (2007) идентифицировал одного человека V6 + в выборке из 35 датуг из Танзании. И дальше на север, Dugoujon et al. (2009) идентифицировал еще 6 мужчин в выборке из 93 человек из Сива оазис, который является берберским населением
E-V92
Trombetta et al. 2011 г. объявил об обнаружении E-V92 в двух амхарах. Подобно E-V6 и E-V42, возможно, он существует только в районе Эфиопии.
Филогенетика
Филогенетическая история
До 2002 г. в академической литературе существовало по крайней мере семь систем именования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть, прежде всего, своевременным. В приведенной ниже таблице собраны все эти работы на основе знакового Дерева YCC 2002 года. Это позволяет исследователю, просматривающему ранее опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (стенография) | (α) | (β) | (γ) | (δ) | (е) | (ζ) | (η) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E-P29 | 21 | III | 3А | 13 | Eu3 | H2 | B | E * | E | E | E | E | E | E | E | E | E | E |
| E-M33 | 21 | III | 3А | 13 | Eu3 | H2 | B | E1 * | E1 | E1a | E1a | E1 | E1 | E1a | E1a | E1a | E1a | E1a |
| E-M44 | 21 | III | 3А | 13 | Eu3 | H2 | B | E1a | E1a | E1a1 | E1a1 | E1a | E1a | E1a1 | E1a1 | E1a1 | E1a1 | E1a1 |
| E-M75 | 21 | III | 3А | 13 | Eu3 | H2 | B | E2a | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 |
| E-M54 | 21 | III | 3А | 13 | Eu3 | H2 | B | E2b | E2b | E2b | E2b1 | - | - | - | - | - | - | - |
| E-P2 | 25 | III | 4 | 14 | Eu3 | H2 | B | E3 * | E3 | E1b | E1b1 | E3 | E3 | E1b1 | E1b1 | E1b1 | E1b1 | E1b1 |
| E-M2 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a * | E3a | E1b1 | E1b1a | E3a | E3a | E1b1a | E1b1a | E1b1a | E1b1a1 | E1b1a1 |
| E-M58 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a1 | E3a1 | E1b1a1 | E1b1a1 | E3a1 | E3a1 | E1b1a1 | E1b1a1 | E1b1a1 | E1b1a1a1a | E1b1a1a1a |
| E-M116.2 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a2 | E3a2 | E1b1a2 | E1b1a2 | E3a2 | E3a2 | E1b1a2 | E1b1a2 | E1ba12 | удаленный | удаленный |
| E-M149 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a3 | E3a3 | E1b1a3 | E1b1a3 | E3a3 | E3a3 | E1b1a3 | E1b1a3 | E1b1a3 | E1b1a1a1c | E1b1a1a1c |
| E-M154 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a4 | E3a4 | E1b1a4 | E1b1a4 | E3a4 | E3a4 | E1b1a4 | E1b1a4 | E1b1a4 | E1b1a1a1g1c | E1b1a1a1g1c |
| E-M155 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a5 | E3a5 | E1b1a5 | E1b1a5 | E3a5 | E3a5 | E1b1a5 | E1b1a5 | E1b1a5 | E1b1a1a1d | E1b1a1a1d |
| E-M10 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a6 | E3a6 | E1b1a6 | E1b1a6 | E3a6 | E3a6 | E1b1a6 | E1b1a6 | E1b1a6 | E1b1a1a1e | E1b1a1a1e |
| E-M35 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b * | E3b | E1b1b1 | E1b1b1 | E3b1 | E3b1 | E1b1b1 | E1b1b1 | E1b1b1 | удаленный | удаленный |
| E-M78 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b1 * | E3b1 | E1b1b1a | E1b1b1a1 | E3b1a | E3b1a | E1b1b1a | E1b1b1a | E1b1b1a | E1b1b1a1 | E1b1b1a1 |
| E-M148 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b1a | E3b1a | E1b1b1a3a | E1b1b1a1c1 | E3b1a3a | E3b1a3a | E1b1b1a3a | E1b1b1a3a | E1b1b1a3a | E1b1b1a1c1 | E1b1b1a1c1 |
| E-M81 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b2 * | E3b2 | E1b1b1b | E1b1b1b1 | E3b1b | E3b1b | E1b1b1b | E1b1b1b | E1b1b1b | E1b1b1b1 | E1b1b1b1a |
| E-M107 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b2a | E3b2a | E1b1b1b1 | E1b1b1b1a | E3b1b1 | E3b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1a | E1b1b1b1a1 |
| E-M165 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b2b | E3b2b | E1b1b1b2 | E1b1b1b1b1 | E3b1b2 | E3b1b2 | E1b1b1b2a | E1b1b1b2a | E1b1b1b2a | E1b1b1b2a | E1b1b1b1a2a |
| E-M123 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b3 * | E3b3 | E1b1b1c | E1b1b1c | E3b1c | E3b1c | E1b1b1c | E1b1b1c | E1b1b1c | E1b1b1c | E1b1b1b2a |
| E-M34 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b3a * | E3b3a | E1b1b1c1 | E1b1b1c1 | E3b1c1 | E3b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1b2a1 |
| E-M136 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3ba1 | E3b3a1 | E1b1b1c1a | E1b1b1c1a1 | E3b1c1a | E3b1c1a | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1b2a1a1 |
Научные публикации
Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
Обсуждение
E-M215 и E1b1b1 являются принятыми в настоящее время именами, найденными в предложениях Консорциум Y-хромосомы (YCC) для клад, определяемых мутациями M215 и M35 соответственно, которые также могут обозначаться как E-M215 и E-M35.[4] Номенклатура E3b (E-M215) и E3b1 (E-M35) соответственно были определенные YCC имена, используемые для обозначения тех же гаплогруппы в более ранней литературе с разветвлением E-M35 как отдельным субклад E-M215 в 2004 году.[8] До 2002 г. эти гаплогруппы не были обозначены единообразно, и их связь с другими родственными клады в гаплогруппа E и гаплогруппа DE. Но в нестандартной или старой терминологии E-M215, например, примерно то же самое, что и «гаплотип V», все еще используемый в таких публикациях, как Жерар и др. (2006).[5]
Смотрите также
Генетика
- Африканская примесь в Европе
- генетическая генеалогия
- Гаплогруппа D
- Гаплогруппа DE
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- Молекулярная филогенетика
- Парагруппа
- Субклад
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК по этнической группе
- Гаплогруппы Y-ДНК в популяциях стран Африки к югу от Сахары
Y-ДНК E Субклады
- Гаплогруппа E-L485
- Гаплогруппа E-M123
- Гаплогруппа E-M180
- Гаплогруппа E-M215
- Гаплогруппа E-M33
- Гаплогруппа E-M521
- Гаплогруппа E-M75
- Гаплогруппа E-M96
- Гаплогруппа E-P147
- Гаплогруппа E-P177
- Гаплогруппа E-P2
- Гаплогруппа E-V12
- Гаплогруппа E-V13
- Гаплогруппа E-V22
- Гаплогруппа E-M2
- Гаплогруппа E-V65
- Гаплогруппа E-V68
- Гаплогруппа E-Z820
- Гаплогруппа E-Z827
Основное дерево Y-ДНК
| Филогенетическое древо человека Гаплогруппы ДНК Y-хромосомы [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-хромосомный Адам " | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| грамм | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | ЧАС | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| я | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | Т | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | п [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| НЕТ | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | О | Q | р | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Примечания
- ^ Cruciani et al. (2004): «Несколько наблюдений указывают на Восточную Африку как на родину гаплогруппы E3b, то есть в ней было (1) наибольшее количество различных кладов E3b (таблица 1), (2) высокая частота этой гаплогруппы и высокое микросателлитное разнообразие, и, наконец, (3) исключительное присутствие недифференцированного E3b * парагруппа. »Как упоминалось выше,« E3b »- старое название E-M215. Semino et al. (2004): "Этот вывод дополнительно подтверждается наличием дополнительной линейной диверсификации Hg E и самой высокой частотой встречаемости E-P2 * и E-M35 * в том же регионе. Распространение E-P2 *, по-видимому, ограничивается восточноафриканскими народами. Линия E-M35 * показывает самую высокую частоту (19,2%) в эфиопском оромо, но с более широким диапазоном распространения, чем E-P2 * ». Для E-M215 Cruciani et al. (2007) снизили свою оценку до 22 400 с 25 600 в Cruciani et al. (2004), повторная калибровка тех же данных.
- ^ «Данные по Y-хромосоме показывают сигнал для отдельной миграции позднего плейстоцена из Африки в Европу через Синай, о чем свидетельствует распределение клонов гаплогруппы E3b, которое не проявляется в распределении гаплогрупп мтДНК».Андерхилл и Кивисилд (2007:547)
- ^ «Парагруппа E-M35 * и гаплогруппа J-12f2a * соответствуют критериям основных линий происхождения AJ, потому что они широко распространены как в популяциях AJ, так и в ближневосточных популяциях, и встречаются гораздо реже в европейских нееврейских популяциях». Behar et al. (2004)
- ^ Cruciani et al. 2007 г. используйте термин «Северо-Восточная Африка» для обозначения Египта и Ливии, как показано в таблице 1 исследования. До Cruciani et al. 2007 г., Semino et al. 2004 г. Восточная Африка как возможное место происхождения E-M78, на основании испытаний в Эфиопии. Это произошло из-за высокой частоты и разнообразия линий E-M78 в регионе Эфиопия. Тем не мение, Cruciani et al. 2007 г. смогли изучить больше данных, включая население из Северной Африки, которое не было представлено в Semino et al. 2004 г. исследования и нашли доказательства того, что линии E-M78, которые составляют значительную часть некоторых популяций в этом регионе, были относительно молодыми ветвями (см. E-V32 ниже). Таким образом, они пришли к выводу, что «Северо-Восточная Африка» была вероятным местом происхождения E-M78, основываясь на «периферийном географическом распределении наиболее производных подгаплогрупп по отношению к северо-востоку Африки, а также на результатах количественного анализа UEP и микросателлитного разнообразия». . Итак, согласно Cruciani et al. 2007 г. E-M35, родительский класс E-M78, возник в Восточной Африке, впоследствии распространился на Северо-Восточную Африку, а затем произошла «обратная миграция» хромосом E-M215, которые приобрели мутацию E-M78. Cruciani et al. 2007 г. поэтому отметьте это как свидетельство «коридора для двунаправленных миграций» между Северо-Восточной Африкой (Египет и Ливия по их данным) с одной стороны и Восточной Африкой с другой. Авторы считают, что было «как минимум 2 эпизода между 23,9–17,3 тыс. Лет назад и 18,0–5,9 тыс. Лет назад».
- ^ Cruciani et al. 2007 г. используйте два метода расчета для оценки возраста E-M78, которые дают очень разные результаты. В течение 18 600 лет назад Метод ASD используется, а на секунду "ρ метод ", используемый в качестве проверки, дает 13,7кья с стандартное отклонение 2,3 тыс. лет назад, но разница между двумя методами велика только для оценки возраста E-M78, а не его субкладов. Авторы заявляют, что большая разница «объясняется соответствующим отклонением от звездообразной структуры из-за повторяющихся эффекты основателя "
- ^ (6 из 112): «Присутствие хромосом североафриканского происхождения (E3b1b-M81; Cruciani et al., 2004) также можно объяснить притоком португальцев, но в первую очередь итальянским, поскольку эта гаплогруппа достигает частоты 4,6. % в Португалия и 4,8% Италия, очень похожая на частоту, найденную в Рио де Жанейро (4,4%) среди европейских вкладчиков ». Silva et al. 2006 г.
- ^ Например, с 11 ноября 2008 г. филогенетический проект E-M35[постоянная мертвая ссылка ] имел записи о четырех тестах E-M123 * по сравнению с 93 результатами тестов с E-M34.
Рекомендации
- ^ а б c d е ж грамм час Trombetta et al. 2015, Филогеографические уточнения и крупномасштабное генотипирование гаплогруппы Е Y-хромосомы человека позволяют по-новому взглянуть на расселение ранних скотоводов на африканском континенте.
- ^ а б Хабер М., Джонс А.Л., Коннел Б.А., Асан, Арчиеро Э., Хуанмин И., Томас М.Г., Сюэ Ю., Тайлер-Смит К. (июнь 2019 г.). «Редкая африканская гаплогруппа Y-хромосомы с глубокими корнями D0 и ее значение для распространения современного человека за пределы Африки». Генетика. 212 (4): 1421–1428. Дои:10.1534 / генетика.119.302368. ЧВК 6707464. PMID 31196864.
- ^ ISOGG (2011)
- ^ а б c Карафет и др. (2008)
- ^ а б c Консорциум Y-хромосомы "YCC" (2002)
- ^ а б c Cruciani et al. (2007)
- ^ Недавнее крупномасштабное расширение отцовства в Европе, показанное повторным упорядочением населения, Кьяра Батини и др., nature.com, 2015
- ^ а б c d е ж грамм Cruciani et al. (2004)
- ^ а б Henn et al. (2008)
- ^ Hassan et al. (2008)
- ^ Lacan et al. (2011)
- ^ «Молекулярно-генетическое исследование истории неолитического населения Западного Карпатского бассейна» (PDF). Архивировано из оригинал (PDF) на 21.07.2015.
- ^ Schuenemann, Verena J .; и другие. (2017). «Геномы древних египетских мумий предполагают увеличение числа африканских предков к югу от Сахары в постримские периоды». Nature Communications. 8: 15694. Bibcode:2017НатКо ... 815694S. Дои:10.1038 / ncomms15694. ЧВК 5459999. PMID 28556824.
- ^ Фрегель; и другие. (2018). «Древние геномы из Северной Африки свидетельствуют о доисторических миграциях в Магриб как из Леванта, так и из Европы». bioRxiv 10.1101/191569.
- ^ Родригес-Варела; и другие. (2017). «Геномный анализ человеческих останков с Канарских островов до европейского завоевания выявил близкое родство с современными североафриканцами». Текущая биология. 27 (1–7): 3396–3402.e5. Дои:10.1016 / j.cub.2017.09.059. PMID 29107554. Получено 27 октября 2017.
- ^ Loosdrecht et al. (2018), Плейстоценовые геномы Северной Африки связывают человеческие популяции Ближнего Востока и Африки к югу от Сахары, Наука, 15 марта 2018 г .: eaar8380 DOI: 10.1126 / science.aar8380
- ^ а б c d Semino et al. (2004)
- ^ Россер и др. (2000)
- ^ Firasat et al. (2006)
- ^ Ehret et al. (2004)
- ^ Кейта и Бойс (2005)
- ^ а б Кейта 2008
- ^ Behar et al. (2003)
- ^ Behar et al. (2004)
- ^ Шен и др. (2004)
- ^ а б Адамс и др. (2008)
- ^ Nebel et al. (2001)
- ^ а б c Moran, Colin N .; и другие. (2004). «Гаплогруппы Y-хромосомы элитных бегунов на выносливость из Эфиопии». Генетика человека. 115 (6): 492–7. Дои:10.1007 / s00439-004-1202-у. PMID 15503146. S2CID 13960753. Получено 6 февраля 2017.
- ^ ISOGG (2008)
- ^ а б c ISOGG 2015
- ^ Cadenas et al. 2007 г.
- ^ Trombetta et al. 2011 г.
- ^ (Рисунок S.7) Дж. Д. Кристофаро и др., 2013, «Афганский Гиндукуш: где сходятся потоки генов на евразийском субконтиненте», http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0076748
- ^ а б Cruciani et al. 2006 г.
- ^ Peričic et al. 2005 г.
- ^ https://yfull.com/tree/E-Y133414/
- ^ https://yfull.com/tree/E-FGC18960*/
- ^ https://yfull.com/tree/E-L19/
- ^ https://www.yfull.com/tree/E-M81/
- ^ Кручиани, Фульвио; Ла Фратта, Роберта; Сантоламацца, Пьеро; Селлитто, Даниэле; Пасконе, Роберто; Мораль, Педро; Уотсон, Элизабет; Гуида, Валентина; Коломб, Элиан Беро (май 2004 г.). «Филогеографический анализ хромосом гаплогруппы E3b (E-M215) Y выявил множественные миграционные события в Африке и за ее пределами». Американский журнал генетики человека. 74 (5): 1014–1022. Дои:10.1086/386294. ISSN 0002-9297. ЧВК 1181964. PMID 15042509.
- ^ Фадхлауи-Зид, Карима; Мартинес-Крус, Бегонья; Ходжет-эль-Хил, Хуссейн; Мендизабал, Изабель; Бенаммар-Эльгаайед, Амель; Комас, Дэвид (октябрь 2011 г.). «Генетическая структура тунисских этносов по отцовской линии». Американский журнал физической антропологии. 146 (2): 271–280. Дои:10.1002 / ajpa.21581. ISSN 1096-8644. PMID 21915847.
- ^ а б Arredi et al. 2004 г.
- ^ Альварес и др. 2009 г.
- ^ Bosch et al. 2001 г.
- ^ Куянова, Мартина; Перейра, Луиза; Фернандес, Вероника; Перейра, Жоана Б.; Черны, Виктор (октябрь 2009 г.). «Ближневосточные неолитические генетические данные в небольшом оазисе египетской Западной пустыни». Американский журнал физической антропологии. 140 (2): 336–346. Дои:10.1002 / ajpa.21078. ISSN 1096-8644. PMID 19425100.
- ^ Робино и др. 2008 г.
- ^ https://yfull.com/tree/E-A5604/
- ^ см. таблицу В архиве 2013-07-30 в Wayback Machine.
- ^ Флорес и др. 2005 г.
- ^ Beleza et al. 2006 г.
- ^ а б Capelli et al. 2009 г.
- ^ а б MacaMaca-Meyer et al. 2003 г.
- ^ Fregel et al. 2009 г., см. таблицу
- ^ Рамос-Луиза и др. 2009 г.
- ^ Были проанализированы только мужчины с французской фамилией, чтобы попытаться исключить более недавних иммигрантов.
- ^ а б Ди Гаэтано и др. 2009 г.
- ^ Сарно, S; Боаттини, А; Carta, M; Ферри, G; Alù, M; Yao, DY; Ciani, G; Петтенер, Д; Луизелли, Д. (2014). «Древний средиземноморский плавильный котел: исследование однородной генетической структуры и истории населения Сицилии и Южной Италии». PLOS ONE. 9 (4): e96074. Bibcode:2014PLoSO ... 996074S. Дои:10.1371 / journal.pone.0096074. ЧВК 4005757. PMID 24788788.
 Эта статья содержит цитаты из этого источника, который доступен под Лицензия Creative Commons Attribution 4.0 International (CC BY 4.0).
Эта статья содержит цитаты из этого источника, который доступен под Лицензия Creative Commons Attribution 4.0 International (CC BY 4.0). - ^ Francalacci et al. (2013), Низкочастотное секвенирование ДНК 1200 сардинцев реконструирует европейскую филогению Y-хромосомы
- ^ (8 из 132), Mendizabal et al. 2008 г.
- ^ (7 из 295), Paracchini et al. 2003 г.
- ^ Ordóñez, A.C .; Fregel, R .; Трухильо-Медерос, А .; Hervella, M .; де-ла-Руа, С .; Арне-де-ла-Роса, М. (2017). «Генетические исследования доиспанского населения, захороненного в пещере Пунта Асуль (Эль Йерро, Канарские острова)». Журнал археологической науки. 78: 20–28. Дои:10.1016 / j.jas.2016.11.004.
- ^ https://yfull.com/tree/E-M81/
- ^ «Архивная копия». Архивировано из оригинал на 2015-09-24. Получено 2012-04-01.CS1 maint: заархивированная копия как заголовок (связь)
- ^ «Архивная копия». Архивировано из оригинал на 2015-09-24. Получено 2012-04-01.CS1 maint: заархивированная копия как заголовок (связь)
- ^ «Архивная копия». Архивировано из оригинал на 2015-09-24. Получено 2012-04-01.CS1 maint: заархивированная копия как заголовок (связь)
- ^ «Архивная копия». Архивировано из оригинал на 2015-09-24. Получено 2012-04-01.CS1 maint: заархивированная копия как заголовок (связь)
- ^ «Архивная копия». Архивировано из оригинал на 2015-09-24. Получено 2013-01-14.CS1 maint: заархивированная копия как заголовок (связь)
Дополнительные источники
- Адамс, Сьюзан М; Босх, Елена; Balaresque, Patricia L .; Ballereau, Stéphane J .; Ли, Эндрю С .; Арройо, Эдуардо; Лопес-Парра, Ана М .; Aler, Mercedes; и другие. (2008), «Генетическое наследие религиозного разнообразия и нетерпимости: отцовское происхождение христиан, евреев и мусульман на Пиренейском полуострове», Американский журнал генетики человека, 83 (6): 725–36, Дои:10.1016 / j.ajhg.2008.11.007, ЧВК 2668061, PMID 19061982
- Альварес; Сантос, Кристина; Монтиэль, Рафаэль; Каейро, Бласкес; Баали, Абделлатиф; Дюгужон, Жан-Мишель; Алуха, Мария Пилар (2009), «Вариации Y-хромосомы в Южной Иберии: взгляд на вклад Северной Африки», Американский журнал биологии человека, 21 (3): 407–409, Дои:10.1002 / ajhb.20888, PMID 19213004, S2CID 7041905
- Арреди, Б; Poloni, E; Параккини, S; Зерджал, Т; Фатхаллах, D; Макрелуф, М; Паскали, V; Новеллетто, А; Тайлерсмит, К. (2004), «Преимущественно неолитическое происхождение вариации Y-хромосомы ДНК в Северной Африке», Американский журнал генетики человека, 75 (2): 338–345, Дои:10.1086/423147, ЧВК 1216069, PMID 15202071
- Бадро, Даниэль А .; Дуайхи, Бушра; Габер, Марк; Юханна, Соня С .; Саллум, Анжелик; Гассиб-Саббаг, Мишелла; Джонсруд, Брайан; Хазен, Жорж; Матисоо-Смит, Элизабет; Сориа-Эрнанц, Дэвид Ф .; Уэллс, Р. Спенсер; Тайлер-Смит, Крис; Platt, Daniel E .; Заллуа, Пьер А. (2013), «Генетика Y-хромосомы и мтДНК выявляет значительные различия в сходстве современных народов Ближнего Востока с европейскими и африканскими популяциями», PLOS ONE, 8 (1: e54616): e54616, Bibcode:2013PLoSO ... 854616B, Дои:10.1371 / journal.pone.0054616, ЧВК 3559847, PMID 23382925
- Батталья, Винченца; Форнарино, Симона; Аль-Захери, Надия; Оливьери, Анна; Пала, Мария; Майрес, Натали М; Кинг, Рой Дж; Рутси, Сиири; и другие. (2008), «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе», Европейский журнал генетики человека, 17 (6): 820–830, Дои:10.1038 / ejhg.2008.249, ЧВК 2947100, PMID 19107149
- Бехар, Дорон М .; Thomas, Mark G .; Скорецкий, Карл; Хаммер, Майкл Ф .; Булыгина, Екатерина; Розенгартен, Дрор; Джонс, Эбигейл Л .; Хелд, Карен; и другие. (Октябрь 2003 г.), "Множественное происхождение левитов ашкенази: свидетельства Y-хромосомы как для ближневосточных, так и для европейских предков", Являюсь. J. Hum. Genet., 73 (4), стр. 768–779, Дои:10.1086/378506, ЧВК 1180600, PMID 13680527. Также на http://www.ucl.ac.uk/tcga/tcgapdf/Behar-AJHG-03.pdf и https://web.archive.org/web/20090304100321/http://www.familytreedna.com/pdf/400971.pdf
- Бехар; Гарриган; Каплан; Мобашер; Розенгартен (ноябрь 2004 г.), «Контрастные модели вариации Y-хромосомы у евреев-ашкенази и нееврейского европейского населения» (PDF), Гм. Genet., 114 (4), стр. 354–365, Дои:10.1007 / s00439-003-1073-7, PMID 14740294, S2CID 10310338, заархивировано из оригинал (PDF) на 2011-11-10, получено 2009-09-11
- Белеза, Сандра; Гусмао, Леонор; Лопес, Александра; Алвес, Синтия; Гомеш, Ива; Джузели, Мария; Калафель, Франсеск; Карраседо, Ангел; Аморим, Антонио (2006), "Микрофилогеографическая и демографическая история португальских мужских линий", Анналы генетики человека, 70 (2): 181–194, Дои:10.1111 / j.1529-8817.2005.00221.x, PMID 16626329, S2CID 4652154[мертвая ссылка ]
- Птица, Стивен (2007), «Гаплогруппа E3b1a2 как возможный индикатор заселения римской Британии солдатами балканского происхождения», Журнал генетической генеалогии, 3 (2)
- Бортолини; Thomas, Mark G .; Чихи, Лурдес; Агилар, Хуан А .; Кастро-де-Герра, Динора; Salzano, Francisco M .; Руис-Линарес, Андрес (2004), «Типология Рибейро, геномы и испанский колониализм с точки зрения Гран-Канарии и Колумбии» (PDF), Генетика и молекулярная биология, 27 (1): 1–8, Дои:10.1590 / S1415-47572004000100001
- Босх, Елена; Калафель, Франсеск; Комас, Дэвид; Oefner, Питер Дж .; Андерхилл, Питер А .; Бертранпетит, Жауме (2001), «Анализ вариаций Y-хромосомы человека с высоким разрешением показывает резкий разрыв и ограниченный поток генов между северо-западной Африкой и Пиренейским полуостровом», Am J Hum Genet, 68 (4): 1019–1029, Дои:10.1086/319521, ЧВК 1275654, PMID 11254456
- Bosch, E .; Calafell, F .; Gonzalez-Neira, A .; Flaiz, C .; Mateu, E .; Scheil, H.-G .; Huckenbeck, W .; Ефремовская, Л .; и другие. (2006), «Отцовские и материнские линии на Балканах демонстрируют однородный ландшафт через языковые барьеры, за исключением изолированных аромунов», Анналы генетики человека, 70 (4): 459–487, Дои:10.1111 / j.1469-1809.2005.00251.x, PMID 16759179, S2CID 23156886, заархивировано из оригинал на 2012-12-10
- Кадены; Животовский, Лев А; Кавалли-Сфорца, Лука Л.; Андерхилл, Питер А; Эррера, Рене Дж. (2007), «Разнообразие Y-хромосомы характеризует Оманский залив», Европейский журнал генетики человека, 16 (3): 1–13, Дои:10.1038 / sj.ejhg.5201934, PMID 17928816
- Капелли, Кристиан; Рыжая, Никола; Абернети, Джулия К .; Гратрикс, Фиона; Уилсон, Джеймс Ф .; Моэн, Торольф; Хервиг, Тор; Ричардс, Мартин; и другие. (2003), «Перепись хромосомы Y на Британских островах», Текущая биология, 13 (11): 979–84, Дои:10.1016 / S0960-9822 (03) 00373-7, PMID 12781138 также в [1]
- Каратти; Джино, С .; Torre, C .; Робино, К. (2009), «Подтипирование Y-хромосомной гаплогруппы E-M78 (E1b1b1a) с помощью анализа SNP и его применения в судебной медицине», Международный журнал судебной медицины, 123 (4): 357–360, Дои:10.1007 / s00414-009-0350-у, PMID 19430804, S2CID 5657112
- Капелли, Кристиан; Онофри, Валерио; Бризигелли, Франческа; Боски, Илария; Скарниччи, Франческа; Масулло, Мара; Ферри, Джанмарко; Тофанелли, Серджио; и другие. (2009), «Мавры и сарацины в Европе: оценка средневекового наследия мужчин Северной Африки в южной Европе», Европейский журнал генетики человека, 17 (6): 848–852, Дои:10.1038 / ejhg.2008.258, ЧВК 2947089, PMID 19156170
- Чинниоглу, Дженгиз; Кинг, Рой; Кивисилд, Тоомас; Калфоглу, Эрси; Атасой, Севиль; Cavalleri, Gianpiero L .; Лилли, Анита С .; Roseman, Charles C .; и другие. (2004), «Раскопки гаплотипов Y-хромосомы в Анатолии», Hum Genet, 114 (2): 127–48, Дои:10.1007 / s00439-003-1031-4, PMID 14586639, S2CID 10763736
- Конту, Даниэла; Морелли, Даниэла; Сантони, Федерико; Фостер, Джейми У .; Франкалаччи, Паоло; Кукка, Франческо (2008), «Доказательства на основе Y-хромосомы для донеолитического происхождения генетически однородного, но разнообразного населения Сардинии: выводы для ассоциативного сканирования» (PDF), PLOS ONE, 3 (1): e1430, Bibcode:2008PLoSO ... 3.1430C, Дои:10.1371 / journal.pone.0001430, ЧВК 2174525, PMID 18183308, заархивировано из оригинал (PDF) на 2016-03-04, получено 2012-01-14
- Кручиани, Фульвио; Сантоламацца, Пьеро; Шен, Пейдун; Маколей, Винсент; Мораль, Педро; Олькерс, Антонель; Модиано, Дэвид; Холмс, Сьюзан (2002), "Обратная миграция из Азии в Африку к югу от Сахары подтверждается анализом гаплотипов Y-хромосомы человека с высоким разрешением", Американский журнал генетики человека, 70 (5): 1197–1214, Дои:10.1086/340257, ЧВК 447595, PMID 11910562
- Cruciani; Ла Фратта; Сантоламацца; Селлитто (май 2004 г.), «Филогеографический анализ хромосом гаплогруппы E3b (E-M215) Y выявил множественные миграционные события в Африке и за ее пределами» (PDF), Американский журнал генетики человека, 74 (5): 1014–1022, Дои:10.1086/386294, ЧВК 1181964, PMID 15042509, заархивировано из оригинал (PDF) на 2008-06-26, получено 2008-05-17
- Cruciani; Ла Фратта; Торрони; Андерхилл; Скоццари (2006), «Молекулярное расслоение гаплогруппы Y-хромосомы E-M78 (E3b1a): апостериорная оценка подхода на основе микросателлитной сети с помощью шести новых двуаллельных маркеров», Человеческая мутация, 27 (8): 831–2, Дои:10.1002 / humu.9445, PMID 16835895, S2CID 26886757
- Cruciani, F .; La Fratta, R .; Trombetta, B .; Santolamazza, P .; Sellitto, D .; Colomb, E. B .; Dugoujon, J.-M .; Crivellaro, F .; и другие. (2007), «Отслеживание прошлых движений мужчин в Северной / Восточной Африке и Западной Евразии: новые ключи к разгадке на основе гаплогрупп Y-хромосомы E-M78 и J-M12», Молекулярная биология и эволюция, 24 (6): 1300–1311, Дои:10.1093 / молбев / msm049, PMID 17351267 Также см Дополнительные данные.
- Ди Гаэтано; Черутти, Франческа; Кробу, Карло; Робино (2009), «Дифференциальная миграция греков и северных африканцев на Сицилию подтверждается генетическими данными по Y-хромосоме», Европейский журнал генетики человека, 17 (1): 91–99, Дои:10.1038 / ejhg.2008.120, ЧВК 2985948, PMID 18685561
- Дугужон; Coudray; Торрони; Cruciani; Скоццари; Мораль; Луали; Косманн (2009), д'Эррико; Гомберт (ред.), «Берберы и берберы: генетическое и лингвистическое разнообразие» (PDF), Становление красноречивым успехом в развитии языка, человеческого познания и современных культур: 123–146, Дои:10.1075 / з.152.05ч4, ISBN 978-90-272-3269-4
- Ehret, C .; Кейта, SO; Ньюман, П. (2004), «Истоки афроазиатского», Наука, 306 (5702): 1680, Дои:10.1126 / science.306.5702.1680c, PMID 15576591, S2CID 8057990
- Эль-Сибай, Мирват; Platt, Daniel E .; Габер, Марк; Сюэ, Яли; Юханна, Соня С .; Уэллс, Р. Спенсер; Изаабель, Хасан; Sanyoura, May F .; и другие. (2009), «Географическая структура Y-хромосомного генетического ландшафта Леванта: контраст между прибрежными и внутренними районами», Анналы генетики человека, 73 (6): 568–581, Дои:10.1111 / j.1469-1809.2009.00538.x, ЧВК 3312577, PMID 19686289, заархивировано из оригинал на 2013-01-05
- Firasat; Халик, Шагуфта; Мохьюддин, Аиша; Папайоанну, Мирто; Тайлер-Смит, Крис; Андерхилл, Питер А; Аюб, Касим (2006), «Доказательства Y-хромосомы ограниченного вклада Греции в патанское население Пакистана», Европейский журнал генетики человека, 15 (1): 121–126, Дои:10.1038 / sj.ejhg.5201726, ЧВК 2588664, PMID 17047675
- Флорес, Карлос; Мака-Мейер, Николь; Гонсалес, Ана М; Эфнер, Питер Дж; Шен, Пейдун; Перес, Хосе А; Рохас, Антонио; Ларруга, Хосе М; Андерхилл, Питер А (2004), «Уменьшение генетической структуры Пиренейского полуострова, выявленное анализом Y-хромосомы: последствия для демографии населения» (PDF), Европейский журнал генетики человека, 12 (10): 855–863, Дои:10.1038 / sj.ejhg.5201225, PMID 15280900, S2CID 16765118, заархивировано из оригинал (PDF) на 2008-04-06
- Флорес; Мака-Мейер, Николь; Ларруга, Хосе М .; Cabrera, Vicente M .; Карадшех, Наиф; Гонсалес, Ана М. (2005), «Изоляты в коридоре миграций: анализ вариации Y-хромосомы в Иордании с высоким разрешением», J Hum Genet, 50 (9): 435–441, Дои:10.1007 / s10038-005-0274-4, PMID 16142507
- Francalacci, P .; Morelli, L .; Андерхилл, П.А.; Lillie, A.S .; Пассарино, G .; Усели, А .; Madeddu, R .; Paoli, G .; и другие. (2003), «Население трех средиземноморских островов (Корсика, Сардиния и Сицилия), обусловленное двуаллельной изменчивостью Y-хромосомы», Американский журнал физической антропологии, 121 (3): 270–279, Дои:10.1002 / ajpa.10265, PMID 12772214
- Фрегель, Роза; Гомеш, Вероника; Гужман, Леонор; Гонсалес, Ана М; Кабрера, Висенте М; Аморим Антониу; Ларруга, Хосе М. (2009), "Демографическая история мужского генофонда Канарских островов: замена местных линий европейскими", BMC Эволюционная биология, 9: 181, Дои:10.1186/1471-2148-9-181, ЧВК 2728732, PMID 19650893
- Жерар; Berriche, S; Aouizérate, A; Diéterlen, F; Люкотт, Джи (2006), «Североафриканские берберы и арабские влияния в западном Средиземноморье выявлены гаплотипами ДНК Y-хромосомы», Человеческая биология, 78 (3): 307–316, Дои:10.1353 / ступица.2006.0045, PMID 17216803, S2CID 13347549
- Гонсалвес, Р. Фрейтас, А; Бранко, М; Роза, А; Fernandes, AT; Животовский, Л.А.; Андерхилл, Пенсильвания; Кивисилд, Т; Брем, А (2005), «Происхождение Y-хромосомы из Португалии, Мадейры и Акорес, записывающих элементы сефардского и берберского происхождения», Анналы генетики человека, 69 (Пт 4): 443–454, Дои:10.1111 / j.1529-8817.2005.00161.x, PMID 15996172, S2CID 3229760[мертвая ссылка ]
- Хаммер (2003), "Структура популяции человека и ее влияние на выборку вариации последовательности Y-хромосомы", Генетика, 164 (4): 1495–1509, ЧВК 1462677, PMID 12930755
- Hassan, Hisham Y .; Андерхилл, Питер А .; Cavalli-Sforza, Luca L .; Ибрагим, Мунтасер Э. (2008), «Вариации Y-хромосомы среди суданцев: ограниченный поток генов, соответствие языку, географии и истории» (PDF), Американский журнал физической антропологии, 137 (3): 316–23, Дои:10.1002 / ajpa.20876, PMID 18618658, заархивировано из оригинал (PDF) на 2009-03-04
- Henn, B.M .; Gignoux, C .; Лин, Алиса А; Oefner, Питер Дж .; Shen, P .; Scozzari, R .; Cruciani, F .; Тишкофф, С. А .; Mountain, J. L .; Андерхилл, П. А. (2008), "Y-хромосомные свидетельства миграции скотоводов через Танзанию в южную Африку", PNAS, 105 (31): 10693–8, Bibcode:2008PNAS..10510693H, Дои:10.1073 / pnas.0801184105, ЧВК 2504844, PMID 18678889. Видеть прокомментировать блог Dienekes, комментарий в блоге Spitoon и публичный выпуск.
- ISOGG (2011), Y-ДНК гаплогруппа E и ее субклады - 2011, Международное общество генетиков-генеологов "ISOGG"
- Jobling, M.A .; Тайлер-Смит, К. (2000), «Новые способы использования новых гаплотипов Y-хромосомы человека, болезнь и отбор», Тенденции Genet., 16 (8): 356–362, Дои:10.1016 / S0168-9525 (00) 02057-6, PMID 10904265
- Карафет, Т. М .; Mendez, F. L .; Meilerman, M. B .; Андерхилл, П. А .; Zegura, S.L .; Хаммер, М. Ф. (май 2008 г.), «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека», Геномные исследования, 18 (5): 830–8, Дои:10.1101 / gr.7172008, ЧВК 2336805, PMID 18385274. Опубликовано в Интернете 2 апреля 2008 г. См. Также Дополнительный материал.
- Кейта, Шомарка (2008), «География, отдельные афро-азиатские семьи и вариация линии Y-хромосомы», В горячих поисках языка в предыстории: очерки в четырех областях антропологии: в честь Гарольда Крейна Флеминга, ISBN 978-90-272-3252-6
- Keita, S.O.Y .; Бойс, А. Дж. (Энтони Дж.) (2005), «Генетика, Египет и история: интерпретация географических закономерностей вариации Y-хромосомы», История в Африке, 32: 221–246, Дои:10.1353 / hia.2005.0013, S2CID 163020672
- King, R.J .; Özcan, S. S .; Картер, Т .; Kalfolu, E .; Атасой, С .; Triantaphyllidis, C .; Kouvatsi, A .; Lin, A. A .; и другие. (2008), «Дифференциальные анатолийские влияния Y-хромосомы на греческий и критский неолит» (PDF), Анналы генетики человека, 72 (2): 205–214, Дои:10.1111 / j.1469-1809.2007.00414.x, PMID 18269686, S2CID 22406638, заархивировано из оригинал (PDF) на 2009-03-05
- Король; Андерхилл (2002), «Конгруэнтное распределение неолитической расписной керамики и керамических фигурок с линиями Y-хромосомы», Античность, 76 (293): 707–14, Дои:10.1017 / S0003598X00091158
- Куянова; Перейра; Фернандес; Перейра; Cerný (2009), "Ближневосточные неолитические генетические данные в небольшом оазисе египетской Западной пустыни", Американский журнал физической антропологии, 140 (2): 336–346, Дои:10.1002 / ajpa.21078, PMID 19425100
- Лакан, Мари; Кейзер, Кристина; Рико, Франсуа-Ксавье; Брукато, Николас; Таррус, Хосеп; Босх, Ангел; Guilaine, Жан; Крабези, Эрик; Лудес, Бертран (2011), «Древняя ДНК предполагает ведущую роль, которую играли люди в неолитическом распространении», PNAS, 108 (45): 18255–9, Bibcode:2011PNAS..10818255L, Дои:10.1073 / pnas.1113061108, ЧВК 3215063, PMID 22042855
- Ланкастер, Эндрю (2009), «Гаплогруппы Y, археологические культуры и языковые семьи: обзор междисциплинарных сравнений на примере E-M35» (PDF), Журнал генетической генеалогии, 5 (1)
- Луис, Дж; Роулд, D; Регейро, М; Caeiro, B; Cinnioglu, C; Roseman, C; Андерхилл, П; Cavallisforza, L; Эррера, Р. (2004), "Левант против Африканского Рога: свидетельства двунаправленных коридоров миграции людей" (PDF), Американский журнал генетики человека, 74 (3): 532–544, Дои:10.1086/382286, ЧВК 1182266, PMID 14973781, заархивировано из оригинал (PDF) на 2012-02-16. (Также см Опечатки )
- Мака-Мейер Н., Санчес-Веласко П., Флорес С., Ларруга Дж. М., Гонсалес А. М., Отерино А., Лейва-Кобиан Ф. и др. (2003), «Характеристика Y-хромосомы и митохондриальной ДНК Pasiegos, человеческого изолята из Кантабрии (Испания)», Анналы генетики человека, 67 (Pt 4): 329–339, CiteSeerX 10.1.1.584.4253, Дои:10.1046 / j.1469-1809.2003.00045.x, PMID 12914567, S2CID 40355653.
- Мартинес, Лайзель; Андерхилл, Питер А; Животовский, Лев А; Гайден, Тензин; Мошонас, Николай К; Чоу, Шерил-Эмилиан Т; Конти, Саймон; Мамолини, Элизабетта; Кавалли-Сфорца, Л. Лука; Эррера, Рене (1 апреля 2007 г.), «Палеолитическое наследие Y-гаплогруппы преобладает на критском высокогорном плато», Европейский журнал генетики человека, 15 (4): 485–493, Дои:10.1038 / sj.ejhg.5201769, ISSN 1018-4813, PMID 17264870
- Мендизабал, Изабель; Сандовал, Карла; Берниелл-Ли, Джемма; Калафель, Франсеск; Салас, Антонио; Мартинес-Фуэнтес, Антонио; Комас, Дэвид (2008), «Генетическое происхождение, примеси и асимметрия в материнских и отцовских линиях человеческого происхождения на Кубе», BMC Evol. Биол., 8: 213, Дои:10.1186/1471-2148-8-213, ЧВК 2492877, PMID 18644108
- Небель; Филон, Д; Бринкманн, Б; Majumder, P; Фаерман, М; Оппенгейм, А. (2001), «Пул Y-хромосомы евреев как часть генетического ландшафта Ближнего Востока», Американский журнал генетики человека, 69 (5): 1095–1112, Дои:10.1086/324070, ЧВК 1274378, PMID 11573163
- Онофри, Валерио; Алессандрини, Федерика; Турчи, Кьяра; Песареси, Мауро; Бушеми, Лоредана; Тальабраччи, Адриано (2006), «Разработка мультиплексных ПЦР для эволюционного и судебно-медицинского применения 37 SNP Y-хромосомы человека» (PDF), Международная криминалистическая экспертиза, 157 (1): 23–35, Дои:10.1016 / j.forsciint.2005.03.014, PMID 15896936[постоянная мертвая ссылка ]
- Параккини; Пирс, CL; Колонель, LN; Альтшулер, Д; Хендерсон, BE; Тайлер-Смит, К. (2003), «Влияние Y-хромосомы на риск рака простаты: многоэтническое когортное исследование», J Med Genet, 40 (11): 815–819, Дои:10.1136 / jmg.40.11.815, ЧВК 1735314, PMID 14627670
- Пелотти; Чеккарди, S; Lugaresi, F; Trane, R; Falconi, M; Бини, С; Willuweit, S; Roewer, L (2007), "Микрогеографические генетические вариации Y-хромосомы в выборке населения области Равенны в регионе Эмилия-Романья (север Италии)", Forensic Science International: Серия дополнений по генетике, 1 (1): 242–243, Дои:10.1016 / j.fsigss.2007.10.025
- Перейра, Луиза; Черный Виктор; Сересо, Мария; Silva, Nuno M; Гайек, Мартин; Вашикова, Альжбета; Куянова, Мартина; Брдичка, Радим; Салас, Антонио (2010), «Объединение генофондов к югу от Сахары и Западной Евразии: материнское и отцовское наследие кочевников туарегов из африканского Сахеля» (PDF), Европейский журнал генетики человека, 18 (8): 915–923, Дои:10.1038 / ejhg.2010.21, ЧВК 2987384, PMID 20234393, заархивировано из оригинал (PDF) на 2013-05-28
- Перичич, М .; Lauc, LB; Кларич, ИМ; Рутси, S; Яничевич, B; Рудан, я; Terzić, R; Чолак, I; и другие. (2005), «Филогенетический анализ с высоким разрешением в юго-восточной Европе отслеживает основные эпизоды отцовского потока генов среди славянского населения», Мол. Биол. Evol., 22 (10), стр. 1964–75, Дои:10.1093 / molbev / msi185, PMID 15944443.
- Понтикос Д. "Филогеографическое уточнение гаплогруппы E" http://dienekes.blogspot.ru/2015/07/phylogeographic-refinement-of.html
- Рамос-Луиза, Э .; Blanco-Verea, A .; Brión, M .; Van Huffel, V .; Карраседо, А .; Санчес-Диз, П. (2009), "Филогеография французских мужских линий (неопубликованные данные 23-го Международного Конгресса ISFG)", Международная криминалистическая экспертиза, 2: 439–441, Дои:10.1016 / j.fsigss.2009.09.026[мертвая ссылка ]
- Regueiro, M .; Кадены, AM; Гайден, Т .; Андерхилл, П.А.; Эррера, Р.Дж. (2006), "Иран: триконтинентальная связь для миграции, управляемой Y-хромосомой", Hum Hered, 61 (3): 132–143, Дои:10.1159/000093774, PMID 16770078, S2CID 7017701
- Робино, С .; Crobu, F .; Gaetano, C .; Bekada, A .; Benhamamouch, S .; Cerutti, N .; Piazza, A .; Inturri, S .; Торре, К. (2008), "Анализ гаплогрупп SNP Y-хромосомы и гаплотипов STR в выборке алжирского населения", Журнал Международный журнал судебной медицины, 122 (3): 251–5, Дои:10.1007 / s00414-007-0203-5, PMID 17909833, S2CID 11556974
- Роза, Александра; Орнелас, Каролина; Джоблинг, Марка А; Брем, Антониу; Виллемс, Ричард (2007), «Разнообразие Y-хромосомы у населения Гвинеи-Бисау: мультиэтническая перспектива» (PDF), BMC Эволюционная биология, 7: 124, Дои:10.1186/1471-2148-7-124, ЧВК 1976131, PMID 17662131
- Россер, З; Зерджал, Т; Hurles, M; Адохаан, М; Алавантик, Д; Аморим, А; Амос, Вт; Арментерос, М; и другие. (2000), «Разнообразие Y-хромосомы в Европе является клинальным и определяется в первую очередь географией, а не языком», Американский журнал генетики человека, 67 (6): 1526–1543, Дои:10.1086/316890, ЧВК 1287948, PMID 11078479, заархивировано из оригинал на 2008-05-06
- Санчес, Хуан Дж; Халленберг, Шарлотта; Борстинг, Клаус; Эрнандес, Алексис; Горлин, Р.Дж. (2005), "Высокие частоты клонов Y-хромосомы, характеризующиеся E3b1, DYS19-11, DYS392-12 у сомалийских мужчин", Европейский журнал генетики человека, 13 (7): 856–866, Дои:10.1038 / sj.ejhg.5201390, PMID 15756297. Опубликовано онлайн 9 марта 2005 г.
- Скоццари, Розария; Cruciani, F; Панграцио, А; Santolamazza, P; Вона, G; Мораль, P; Латини, V; Варези, L; и другие. (2001), "Изменения Y-хромосомы человека в районе Западного Средиземноморья: последствия для населения региона" (PDF), Иммунология человека, 62 (9): 871–884, CiteSeerX 10.1.1.408.4857, Дои:10.1016 / S0198-8859 (01) 00286-5, PMID 11543889
- Семино, О .; Пассарино, G; Oefner, PJ; Линь А.А.; Арбузова, С; Бекман, LE; Де Бенедиктис, G; Francalacci, P; и другие. (2000), «Генетическое наследие палеолита. Homo sapiens sapiens в современных европейцах: перспектива хромосомы Y " (PDF), Наука, 290 (5494), стр. 1155–59, Bibcode:2000Sci ... 290.1155S, Дои:10.1126 / наука.290.5494.1155, PMID 11073453, заархивировано из оригинал (PDF) на 2003-11-25.
- Семино; Сантачиара-Бенеречетти, А. Сильвана; Фаласки, Франческо; Кавалли-Сфорца, Л. Лука; Андерхилл, Питер А. (2002), «Эфиопы и койсаны разделяют самые глубокие клады филогении Y-хромосомы человека» (PDF), Am J Hum Genet, 70 (1), стр. 265–268, Дои:10.1086/338306, ЧВК 384897, PMID 11719903, заархивировано из оригинал (PDF) на 2006-03-15
- Семино, Орнелла; Магри, Кьяра; Бенуцци, Джорджия; Лин, Алиса А .; Аль-Захери, Надия; Батталья, Винченца; Макчони, Лилиана; Triantaphyllidis, Costas; и другие. (2004), "Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморском регионе", Американский журнал генетики человека, 74 (5), стр. 1023–1034, Дои:10.1086/386295, ЧВК 1181965, PMID 15069642
- Шен, Пейдун; Лави, Тал; Кивисилд, Тоомас; Чоу, Вивиан; Сенгун, Дениз; Гефель, Дов; Шпирер, Иссак; Вульф, Эйлон; и другие. (2004), «Реконструкция отцовства и материнства самаритян и других израильских популяций на основе Y-хромосомы и вариации последовательности митохондриальной ДНК» (PDF), Человеческая мутация, 24 (3), стр. 248–60, Дои:10.1002 / humu.20077, PMID 15300852, S2CID 1571356, заархивировано из оригинал (PDF) на 2009-03-05, получено 2008-11-07
- Сильва; Карвалью, Элизеу; Коста, Гильерме; Таварес, Лигия; Аморим Антониу; Гужман, Леонор (2006), «Генетическая изменчивость Y-хромосомы в популяции Рио-де-Жанейро», Американский журнал биологии человека, 18 (6): 829–837, Дои:10.1002 / ajhb.20567, PMID 17039481, S2CID 23778828, заархивировано из оригинал на 2012-10-18
- Шлуш; Бехар, Дорон М .; Юдковский, Геннадий; Темплтон, Алан; Хадид, Ярин; Базис, Фуад; Хаммер, Майкл; Ицковиц, Шалев; Скорецкий, Карл (2008), Геммелл, Нил Джон (редактор), «Друзы: популяционный генетический рефугиум Ближнего Востока», PLOS ONE, 3 (5): e2105, Bibcode:2008PLoSO ... 3.2105S, Дои:10.1371 / journal.pone.0002105, ЧВК 2324201, PMID 18461126
- Сайкс, Брайан (2006), Кровь островов: исследование генетических корней нашей племенной истории, Бантам, ISBN 978-0-593-05652-3
- Тишкофф; Гондер; Хенн; Мортенсен; Рыцарь (2007), "История кликабельных популяций Африки на основе генетической изменчивости мтДНК и Y-хромосомы", Мол Биол Эвол, 24 (10): 2180–2195, Дои:10.1093 / молбев / мсм155, PMID 17656633
- Томас; Штумпф, М. П. Х .; Харке, Х. (2006), «Доказательства социальной структуры, подобной апартеиду, в ранней англосаксонской Англии» (PDF), Труды Королевского общества B, 273 (1601): 2651–2657, Дои:10.1098 / rspb.2006.3627, ЧВК 1635457, PMID 17002951, заархивировано из оригинал (PDF) на 2009-03-05
- Тромбетта, Бениамино; Кручиани, Фульвио; Селлитто, Даниэле; Скоццари, Розария (2011), Маколей, Винсент (редактор), «Новая топология гаплогруппы E1b1 (E-P2) Y-хромосомы человека, выявленная благодаря использованию недавно охарактеризованных бинарных полиморфизмов», PLOS ONE, 6 (1): e16073, Bibcode:2011PLoSO ... 616073T, Дои:10.1371 / journal.pone.0016073, ЧВК 3017091, PMID 21253605
- Тромбетта Б. «Филогеографическое уточнение и крупномасштабное генотипирование Y-хромосомы гаплогруппы E человека дает новый взгляд на рассредоточение ранних скотоводов на африканском континенте» http://gbe.oxfordjournals.org/content/7/7/1940.long
- Андерхилл, Питер А .; Шен, Пейдун; Лин, Алиса А .; Джин, Ли; Пассарино, Джузеппе; Ян, Вэй Х .; Кауфман, Эрин; Бонне-Тамир, Батшева; и другие. (2000), "Вариации последовательности Y-хромосомы и история человеческих популяций", Нат Жене, 26 (3), стр. 358–361, Дои:10.1038/81685, PMID 11062480, S2CID 12893406
- Андерхилл; Пассарино, G .; Lin, A. A .; Shen, P .; Mirazon Lahr, M .; Foley, R.A .; Oefner, P.J .; Кавалли-Сфорца, Л. Л. (2001), «Филогеография бинарных гаплотипов Y-хромосомы и происхождение современных человеческих популяций» (PDF), Энн Хам Жене, 65 (Pt 1), pp. 43–62, Дои:10.1046 / j.1469-1809.2001.6510043.x, PMID 11415522, S2CID 9441236, заархивировано из оригинал (PDF) на 2011-07-21
- Андерхилл (2002), Беллвуд и Ренфрю (ред.), Заключение неолитической истории населения с использованием гаплотипов Y-хромосомы, Кембридж: Институт археологических исследований Макдональда, ISBN 978-1-902937-20-5
- Андерхилл, Питер А .; Кивисилд, Тоомас (2007), «Использование структуры популяции Y-хромосомы и митохондриальной ДНК в отслеживании миграций человека», Анну. Преподобный Жене., 41: 539–64, Дои:10.1146 / annurev.genet.41.110306.130407, PMID 18076332
- Weale, M.E .; Weiss, D.A .; Jager, R. F .; Bradman, N .; Томас, М. Г. (2002), "Свидетельства Y-хромосомы об англосаксонской массовой миграции" (PDF), Мол. Биол. Evol., 19 (7), стр. 1008–1021, Дои:10.1093 / oxfordjournals.molbev.a004160, PMID 12082121.
- Уил; Шах, Т; Джонс, Алабама; Greenhalgh, J; Wilson, JF; Nymadawa, P; Zeitlin, D; Коннелл, BA; и другие. (1 сентября 2003 г.), "Редкие глубоко укоренившиеся линии Y-хромосомы у людей: уроки филогеографии", Генетика, 165 (1), стр. 229–234, ЧВК 1462739, PMID 14504230
- Дерево; Стовер; Эрет; Дестро-Бисол (2005 г.), «Контрастные модели вариаций Y-хромосомы и мтДНК в Африке: свидетельства предвзятых по полу демографических процессов», Европейский журнал генетики человека, 13 (7): 867–876, Дои:10.1038 / sj.ejhg.5201408, PMID 15856073
- Консорциум Y-хромосомы "YCC" (2002), "Система номенклатуры древа бинарных гаплогрупп Y-хромосомы человека", Геномные исследования, 12 (2), стр. 339–348, Дои:10.1101 / гр.217602, ЧВК 155271, PMID 11827954
- Zalloua, Pierre A .; Сюэ, Яли; Халифе, Джейд; Махоул, Надин; Debiane, Labib; Platt, Daniel E .; Ройюру, Аджай К .; Эррера, Рене Дж .; и другие. (2008), «Разнообразие Y-хромосомы в Ливане структурировано недавними историческими событиями», Американский журнал генетики человека, 82 (4): 873–882, Дои:10.1016 / j.ajhg.2008.01.020, ЧВК 2427286, PMID 18374297
- Zalloua, Pierre A .; Platt, Daniel E .; Эль-Сибай, Мирват; Халифе, Джейд; Махоул, Надин; Габер, Марк; Сюэ, Яли; Изаабель, Хасан; и другие. (2008), «Идентификация генетических следов исторической экспансии: финикийские следы в Средиземном море», Американский журнал генетики человека, 83 (5): 633–642, Дои:10.1016 / j.ajhg.2008.10.012, ЧВК 2668035, PMID 18976729
- Зерджал (1999), Использование вариации ДНК Y-хромосомы для изучения истории популяции; в Papiha SS, Deka R, Chakraborty R (eds): Геномное разнообразие: применение в популяционной генетике человека, Kluwer Academic / Plenum Publishers, стр. 91–101.
- Чжао; Хан, Фейсал; Боркар, Минал; Эррера, Рене; Агравал, Суракша (2009), «Наличие трех разных отцовских линий среди северных индейцев: исследование 560 Y-хромосом», Анналы биологии человека, 36 (1): 1–14, Дои:10.1080/03014460802558522, ЧВК 2755252, PMID 19058044
внешняя ссылка
- E-M35 Филогенетический проект вики
- E-M35 Phylogeny Project (сайт всех лабораторий)
- Общественный форум проекта филогенеза E-M35
- Еврейский проект E3b в FTDNA
- Карта распространения E-M35 в Европе
- Гаплогруппа E3B1 (E1b1b1) Группа в Facebook (сообщество M35)
- E1b1b1b * -A Проект
- E1b1.org - Международный проект Y-ДНК гаплогруппы E1b1 и ее субкладов
{kind=link}