Доминирование (генетика) - Dominance (genetics) - Wikipedia
Эта статья нужны дополнительные цитаты для проверка. (Февраль 2018 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |
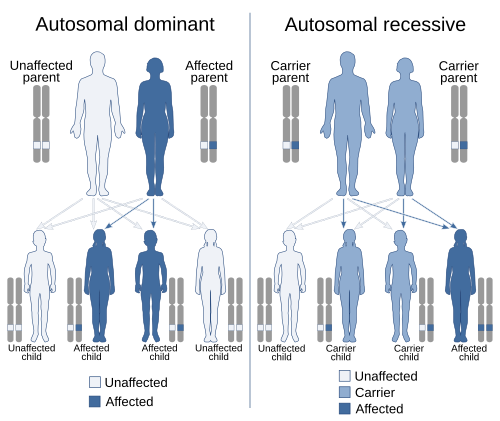
В генетика, господство явление одного варианта (аллель ) из ген на хромосома маскировка или переопределение эффект другого варианта того же гена на другая копия хромосомы.[1][2] Первый вариант называется доминирующий а второй рецессивный. Это состояние обладания два разных варианта одного и того же гена на каждой хромосоме изначально вызвано мутация в одном из генов либо новый (de novo) или же унаследованный. Условия аутосомно-доминантный или же аутосомно-рецессивный используются для описания вариантов генов на неполовых хромосомах (аутосомы ) и связанных с ними черт, а на половые хромосомы (аллосомы) называются Х-сцепленный доминантный, Х-сцепленный рецессивный или же Y-соединенный; у них есть шаблон наследования и представления, который зависит от пола как родителя, так и ребенка (см. Секс связь ). Поскольку существует только одна копия Y-хромосома, Y-сцепленные признаки не могут быть ни доминантными, ни рецессивными. Кроме того, существуют другие формы доминирования, такие как неполное доминирование, при котором вариант гена имеет частичный эффект по сравнению с тем, когда он присутствует на обеих хромосомах, и совместное доминирование, в котором разные варианты на каждой хромосоме демонстрируют связанные с ними черты.
Доминирование не присуще аллелю или его признакам (фенотип ). Это строго относительный эффект между двумя аллелями данного гена любой функции; один аллель может доминировать над вторым аллелем того же гена, рецессивным по отношению к третьему и содоминантный с четвертым. Кроме того, один аллель может быть доминирующим для одного признака, но не для других.
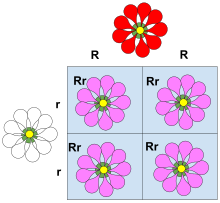
Доминирование - ключевая концепция в Менделирующее наследование и классическая генетика. Письма и Квадраты Пеннета используются для демонстрации принципов доминирования в обучении, а использование прописных букв для доминантных аллелей и строчных букв для рецессивных аллелей является широко применяемым соглашением. Классический пример доминирования - это наследование семя форма в горох. Горох может быть круглым, ассоциированным с аллелем р, или морщинистый, связанный с аллелем р. В этом случае возможны три комбинации аллелей (генотипов): RR, Rr, и rr. В RR (гомозиготный ) у особей круглый горошек, а у rr (гомозиготные) особи имеют морщинистый горох. В Rr (гетерозиготный ) физическим лицам р аллель маскирует присутствие р аллель, поэтому у этих особей также есть круглый горошек. Таким образом, аллель р доминирует над аллелем р, и аллель р рецессивен к аллелю р.
Доминирование отличается от эпистаз, феномен аллеля одного гена, маскирующий действие аллелей разные ген.[3]
Фон

Концепция доминирования была введена Грегор Иоганн Мендель. Хотя Мендель, «отец генетики», впервые использовал этот термин в 1860-х годах, он не был широко известен до начала двадцатого века. Мендель заметил, что для множества признаков садового гороха, связанных с внешним видом семян, семенных коробочек и растений, существует два отдельных фенотипа, таких как круглые или морщинистые семена, желтые или зеленые семена, красные и белые цветы или высокие растения по сравнению с низкорослыми. При раздельном разведении растения всегда давали одни и те же фенотипы, поколение за поколением. Однако когда линии с разными фенотипами скрещивались (скрещивались), у потомства обнаруживался один и только один родительский фенотип (зеленый, круглый, красный или высокий). Однако когда эти гибридный При скрещивании растений-потомков были обнаружены два исходных фенотипа в характерном соотношении 3: 1, причем наиболее распространенным был фенотип родительских гибридных растений. Мендель рассуждал, что каждый родитель в первом скрещивании был гомозиготным по разным аллелям (один родительский AA и другой родительский aa), что каждый вносил один аллель в потомство, в результате чего все эти гибриды были гетерозиготами (Aa) и что один из двух аллелей в гибридном кроссе доминировал над экспрессией другого: A замаскированный a. Окончательное скрещивание двух гетерозигот (Aa X Aa) даст потомство AA, Aa и aa в соотношении генотипов 1: 2: 1, при этом первые два класса демонстрируют фенотип (A), а последний - фенотип (a). , тем самым создавая соотношение фенотипов 3: 1.
Мендель не использовал термины ген, аллель, фенотип, генотип, гомозигота и гетерозигота, все из которых были введены позже. Он ввел обозначения прописных и строчных букв для доминантных и рецессивных аллелей, соответственно, которые используются до сих пор.
В 1928 году британский популяционный генетик Рональд Фишер предположил, что доминирование действует на основе естественного отбора за счет вклада гены-модификаторы. В 1929 году американский генетик Сьюэлл Райт ответил, заявив, что доминирование - это просто физиологическое следствие метаболических путей и относительной необходимости задействованного гена. Объяснение Райта стало установленным фактом в генетике, и дебаты в основном прекратились. Однако на некоторые черты могут влиять эволюционные механизмы.[4][5][6]
Хромосомы, гены и аллели
Большинство животных и некоторые растения спарились хромосомы, и описываются как диплоидные. У них есть две версии каждой хромосомы, одна из которых принадлежит материнской яйцеклетка, а другой по отцовскому сперма, известный как гаметы, описывается как гаплоид и создается посредством мейоз. Эти гаметы затем сливаются во время оплодотворение в течение половое размножение, в новую одиночную ячейку зигота, который делится несколько раз, в результате чего получается новый организм с таким же количеством пар хромосом в каждой (не гаметной) клетке, что и его родители.
Каждая хромосома подходящей (гомологичной) пары структурно похожа на другую и имеет очень похожие ДНК последовательность (места, особый годограф). ДНК в каждой хромосоме функционирует как серия дискретных гены которые влияют на различные черты характера. Таким образом, каждый ген также имеет соответствующий гомолог, который может существовать в разных версиях, называемых аллели. Аллели одного и того же локуса на двух гомологичных хромосомах могут быть идентичными или разными.
В группа крови человека определяется геном, который создает A, B, AB или O группа крови и находится в длинном плече девятой хромосомы. Есть три разных аллеля, которые могут присутствовать в этом локусе, но только два могут присутствовать у любого человека: один унаследован от их матери, а другой - от отца.[7]
Если два аллеля данного гена идентичны, организм называется гомозиготным и считается гомозиготным по отношению к этому гену; если вместо этого два аллеля различны, организм является гетерозиготным и гетерозиготным. Генетический состав организма, находящийся либо в одном локусе, либо по всем его генам в совокупности, называется его генетическим составом. генотип. Генотип организма прямо или косвенно влияет на его молекулярные, физические и другие характеристики, которые по отдельности или вместе называются его фенотип. В гетерозиготных генных локусах эти два аллеля взаимодействуют, создавая фенотип.
Доминирование
Полное господство
При полном доминировании влияние одного аллеля в гетерозиготном генотипе полностью маскирует влияние другого. Аллель, маскирующий другого, называется доминирующий к последнему, и замаскированный аллель называется рецессивный к бывшему.[8] Следовательно, полное доминирование означает, что фенотип гетерозиготы неотличим от фенотипа доминирующей гомозиготы.
Классический пример доминирования - наследование формы семян (формы гороха) у гороха. Горох может быть круглым (ассоциирован с аллелем р) или морщинистый (связанный с аллелем р). В этом случае три комбинации аллелей (генотипы ) возможны: RR и rr гомозиготны и Rr гетерозиготен. В RR у людей есть круглый горошек и rr у отдельных лиц горох сморщенный. В Rr отдельные лица р аллель маскирует присутствие р аллель, поэтому у этих особей также есть круглый горошек. Таким образом, аллель р полностью доминантен для аллеля р, и аллель р рецессивен к аллелю р.
Полу-доминирование
Неполное доминирование (также называемое частичное доминирование, полу-доминирование или же промежуточное наследование) возникает, когда фенотип гетерозиготного генотипа отличается от фенотипов гомозиготных генотипов и часто является промежуточным. Например, львиный зев Цвет цветков гомозиготен по красному или белому цвету. Когда красный гомозиготный цветок сочетается с белым гомозиготным цветком, в результате получается розовый цветок львиный зев. Розовый львиный зев - результат неполного доминирования. Подобный тип неполного доминирования встречается в четырехчасовой завод при этом розовый цвет получается при скрещивании чистокровных родителей белого и красного цветов. В количественная генетика, где фенотипы измеряются и обрабатываются численно, если фенотип гетерозиготы находится точно между (численно) фенотипом двух гомозигот, считается, что фенотип проявляет нет доминирования вообще, т.е. доминирование существует только тогда, когда мера фенотипа гетерозиготы находится ближе к одной гомозиготе, чем к другой.
Когда растения F1 поколения самоопыляемые, фенотипическое и генотипическое соотношение F2 поколение будет 1: 2: 1 (красный: розовый: белый).[9]
Видеть гипотеза частичного доминирования.
Совместное доминирование



Совместное доминирование происходит, когда в фенотипе видны вклады обоих аллелей.
Например, в Система групп крови ABO, химические модификации к гликопротеин (H-антиген) на поверхности клеток крови контролируются тремя аллелями, два из которых являются совмещенными друг с другом (яА, яB) и доминирует над рецессивным я на Локус ABO. В яА и яB аллели производят различные модификации. Фермент, кодируемый яА добавляет N-ацетилгалактозамин к мембраносвязанному H-антигену. В яB фермент добавляет галактозу. В я аллель не вызывает модификации. Таким образом яА и яB каждый из аллелей доминирует я (яАяА и яАя у обоих есть кровь типа А, и яBяB и яBя у обоих есть кровь типа B), но яАяB у людей есть обе модификации в своих клетках крови и, следовательно, кровь типа AB, поэтому яА и яB аллели считаются содоминантными.
Другой пример встречается в локусе бета-глобин компонент гемоглобин, где три молекулярных фенотипа HbА/ HbА, HbА/ HbS, и HbS/ HbS все различимы электрофорез белков. (Медицинское состояние, вызванное гетерозиготным генотипом, называется серповидно-клеточная характеристика и это более легкое состояние, отличное от серповидноклеточная анемия, таким образом, аллели показывают неполное доминирование относительно анемии см. выше). Для большинства локусов генов на молекулярном уровне оба аллеля экспрессируются ко-доминантно, поскольку оба являются записано в РНК.
Совместное доминирование, при котором продукты аллелей сосуществуют в фенотипе, отличается от неполного доминирования, когда количественное взаимодействие продуктов аллелей дает промежуточный фенотип. Например, при совместном доминировании красный гомозиготный цветок и белый гомозиготный цветок будут давать потомство с красными и белыми пятнами. Когда растения поколения F1 самоопыляются, фенотипическое и генотипическое соотношение поколения F2 будет 1: 2: 1 (красный: пятнистый: белый). Эти соотношения такие же, как и для неполного доминирования. Опять же, эта классическая терминология неуместна - в действительности нельзя говорить о том, что такие случаи демонстрируют доминирование.
Устранение распространенных заблуждений
Эта секция не цитировать любой источники. (Январь 2020) (Узнайте, как и когда удалить этот шаблон сообщения) |
Хотя часто удобно говорить о рецессивный аллель или доминирующая черта, доминирование не присуще ни аллелю, ни его фенотипу. Доминирование - это связь между двумя аллелями гена и их связанными фенотипами. «Доминантный» аллель является доминантным по отношению к конкретному аллелю того же гена, который может быть выведен из контекста, но он может быть рецессивным по отношению к третьему аллелю и кодоминантным по отношению к четвертому. Точно так же «рецессивный» признак - это признак, связанный с конкретным рецессивным аллелем, подразумеваемым контекстом, но тот же самый признак может встречаться в другом контексте, где он обусловлен каким-то другим геном и доминантным аллелем.
Доминирование не связано с природой самого фенотипа, то есть с тем, считается ли он «нормальным» или «ненормальным», «стандартным» или «нестандартным», «здоровым» или «больным», «более сильным» или «более слабым», "или более или менее экстремально. Доминантный или рецессивный аллель может объяснять любой из этих типов признаков.
Доминирование не определяет, является ли аллель вредоносным, нейтральным или полезным. Тем не мение, отбор должны опосредованно воздействовать на гены через фенотипы, а доминирование влияет на выявление аллелей в фенотипах и, следовательно, на скорость изменения частот аллелей при отборе. Вредные рецессивные аллели могут сохраняться в популяции с низкими частотами, при этом большинство копий переносятся в гетерозиготах без каких-либо затрат для этих людей. Эти редкие рецессивы являются основой многих наследственных заболеваний. генетические нарушения.
Доминирование также не связано с распределением аллелей в популяции. Как доминантные, так и рецессивные аллели могут быть чрезвычайно частыми или крайне редкими.
Номенклатура
В генетике символы начинались как алгебраические заполнители. Когда один аллель является доминантным по отношению к другому, согласно старейшим правилам, доминантный аллель обозначается заглавной буквой. Рецессивному аллелю присваивается такая же буква в нижнем регистре. В примере с горохом, когда известна взаимосвязь доминирования между двумя аллелями, можно обозначить доминантный аллель, который дает круглую форму, заглавным символом. р, и рецессивный аллель, образующий морщинистую форму с помощью символа нижнего регистра р. Затем записываются гомозиготные доминантные, гетерозиготные и гомозиготные рецессивные генотипы. RR, Rr, и rr, соответственно. Также можно было бы обозначить эти два аллеля как W и ш, и три генотипа WW, Ww, и ww, первые два производили круглый горошек, а третье - морщинистый. Выбор "р" или же "W«как символ доминантного аллеля не предопределяет, является ли аллель, вызывающий« круглый »или« морщинистый »фенотип, когда гомозиготный, доминирующим.
У гена может быть несколько аллелей. Каждый аллель обозначен символом локуса, за которым следует уникальный верхний индекс. У многих видов самый распространенный аллель в дикой популяции обозначается как аллель дикого типа. Он обозначается знаком + в качестве надстрочного индекса. Другие аллели являются доминантными или рецессивными по отношению к аллелю дикого типа. Для рецессивных аллелей символ локуса пишется строчными буквами. Для аллелей с любой степенью доминирования по отношению к аллелю дикого типа первая буква символа локуса находится в верхнем регистре. Например, вот некоторые из аллелей на а локус лабораторной мыши, Mus musculus: Ау, преобладающий желтый; а+, дикого типа; и аbt, черно-подпалый. В аbt аллель рецессивен по отношению к аллелю дикого типа, а Ау аллель кодоминантен аллелю дикого типа. В Ау аллель также кодоминантен аbt аллель, но показывая, что родство выходит за рамки правил генетической номенклатуры мышей.
Правила генетической номенклатуры развивались по мере усложнения генетики. Комитеты стандартизировали правила для некоторых видов, но не для всех. Правила для одного вида могут несколько отличаться от правил для другого вида.[10][11]
Связь с другими генетическими концепциями
Множественные аллели
Хотя любой особь диплоидного организма имеет не более двух разных аллелей в одном локусе (за исключением анеуплоидии ), большинство генов существует в большом количестве аллельных версий в популяции в целом. Если аллели по-разному влияют на фенотип, иногда их отношения доминирования можно описать как серию.
Например, на цвет шерсти домашних кошек влияет ряд аллелей TYR ген (кодирующий фермент тирозиназа ). Аллели C, cб, cs, и cа (полноцветный, Бирманский, Сиамский, и альбинос соответственно) производят разные уровни пигмента и, следовательно, разные уровни разбавления цвета. В C аллель (полный цвет) полностью доминирует над последними тремя и cа аллель (альбинос) полностью рецессивен к первым трем.[12][13][14]
Аутосомный против доминирование, связанное с полом
Эта секция не цитировать любой источники. (Январь 2020) (Узнайте, как и когда удалить этот шаблон сообщения) |
У людей и других млекопитающее разновидность, пол определяется двумя половыми хромосомами, называемыми Х хромосома и Y-хромосома. Человеческие женщины обычно XX; мужчины обычно XY. Остальные пары хромосом встречаются у обоих полов и называются аутосомы; генетические признаки, обусловленные локусами на этих хромосомах, описываются как аутосомные и могут быть доминантными или рецессивными. Генетические признаки на Икс и Y хромосомы называются сцепленными с полом, потому что они связаны с половыми хромосомами, а не потому, что они характерны для того или иного пола. На практике этот термин почти всегда относится к Икс-связанные черты и очень многие из них (например, недостаточность красно-зеленого цветового зрения) не зависят от пола. У женщин есть две копии каждого локуса гена, обнаруженного на Х-хромосоме, так же, как и для аутосом, и применяются те же отношения доминирования. Однако самцы имеют только одну копию каждого локуса гена Х-хромосомы и описываются как гемизиготный для этих генов. Y-хромосома намного меньше, чем Икс, и содержит гораздо меньший набор генов, включая, помимо прочего, те, которые влияют на «мужественность», такие как Ген SRY за фактор, определяющий яичко. Правила доминирования локусов сцепленных с полом генов определяются их поведением у самки: поскольку самец имеет только один аллель (за исключением некоторых типов Y-хромосомы анеуплоидия ), этот аллель всегда выражен независимо от того, доминантный он или рецессивный. Птицы имеют противоположные половые хромосомы: у самцов - ZZ, у самок - ZW. Однако в остальном наследование признаков напоминает XY-систему; самцы зебровых зябликов могут нести ген белой окраски в одной из двух Z-хромосом, но у самок белая окраска всегда развивается. У кузнечиков есть XO-система. У женщин XX, а у мужчин только X. Y-хромосомы вообще нет.
Эпистаз
Эпистаз ["эпи + застой = сидеть сверху "] - это взаимодействие между аллелями в двух разные локусы генов, которые влияют на один признак, что иногда может напоминать доминирующее взаимодействие между двумя разные аллели на одно и тоже локус. Эпистаз изменяет характеристику 9:3:3:1 соотношение ожидается для двух неэпистатических генов. Для двух локусов распознается 14 классов эпистатических взаимодействий. В качестве примера рецессивный эпистаз, один локус гена может определять, является ли пигмент цветка желтым (AA или же Аа) или зеленый (аа), а другой локус определяет, производится ли пигмент (BB или же Bb) или нет (bb). В bb растение, цветы будут белыми, независимо от генотипа другого локуса, так как AA, Аа, или же аа. В bb комбинация нет доминирует над А аллель: скорее, B ген показывает рецессивный эпистаз к А ген, потому что B локус, когда гомозиготен по рецессивный аллель (bb) подавляет фенотипическое выражение А локус. В помесь двух AaBb растения, это дает характерный 9:3:4 соотношение, в данном случае желтые: зеленые: белые цветы.
В доминантный эпистаз, один локус гена может определять желтый или зеленый пигмент, как в предыдущем примере: AA и Аа желтые, и аа зеленые. Второй локус определяет, продуцируется ли предшественник пигмента (дд) или нет (DD или же Дд). Здесь, в DD или же Дд растения, цветы будут бесцветными независимо от генотипа на А локус из-за эпистатического эффекта доминантного D аллель. Таким образом, в помеси двух AaDd растений, 3/4 растений будут бесцветными, а фенотипы желтого и зеленого цвета выражаются только в дд растения. Это дает характеристику 12:3:1 соотношение белого: желтого: зеленого растений.
Дополнительный эпистаз возникает, когда два локуса влияют на один и тот же фенотип. Например, если цвет пигмента производится CC или же Копия но нет cc, и по DD или же Дд но нет дд, то пигмент не образуется ни в одной генотипической комбинации ни с cc или же дд. То есть, обе В локусах должен быть хотя бы один доминантный аллель, чтобы вызвать фенотип. Это дает характеристику 9:7 соотношение пигментированных и непигментированных растений. Дополнительный эпистаз напротив, производит непигментированное растение тогда и только тогда, когда генотип cc и дд, а характеристическое отношение 15:1 между пигментированными и непигментированными растениями.[15]
Классическая генетика рассматривала эпистатические взаимодействия между двумя генами одновременно. В настоящее время из молекулярной генетики очевидно, что все локусы генов вовлечены в сложные взаимодействия со многими другими генами (например, метаболические пути могут включать множество генов), и что это создает эпистатические взаимодействия, которые намного сложнее, чем в классических двухлокусных моделях. .
Принцип Харди – Вайнберга (оценка несущей частоты)
Эта секция не цитировать любой источники. (Январь 2020) (Узнайте, как и когда удалить этот шаблон сообщения) |
Частоту гетерозиготного состояния (которое является состоянием носителя рецессивного признака) можно оценить с помощью Формула Харди – Вайнберга:

Эта формула применяется к гену с ровно двумя аллелями и связывает частоты этих аллелей в большой популяции с частотами их трех генотипов в этой популяции.
Например, если п это частота аллеля А, и q это частота аллеля а тогда условия п2, 2pq, и q2 частоты генотипов AA, Аа и аа соответственно. Поскольку у гена только два аллеля, все аллели должны быть либо А или же а и п + q = 1. Сейчас если А полностью доминирует над а тогда частота генотипа носителя Аа нельзя непосредственно наблюдать (поскольку он имеет те же признаки, что и гомозиготный генотип AA), однако его можно оценить по частоте рецессивного признака в популяции, так как он такой же, как и у гомозиготного генотипа аа. т.е. можно оценить частоту отдельных аллелей: q = √ж(аа), п = 1 − q, и из них можно определить частоту генотипа носителя: ж(Аа) = 2pq.
Эта формула опирается на ряд предположения и точная оценка частоты рецессивного признака. В общем, любая реальная ситуация будет в некоторой степени отклоняться от этих предположений, внося соответствующие неточности в оценку. Если рецессивный признак встречается редко, то будет трудно точно оценить его частоту, так как потребуется очень большой размер выборки.
Доминирующий против выгодного
Эта секция не цитировать любой источники. (Январь 2020) (Узнайте, как и когда удалить этот шаблон сообщения) |
Свойство «доминанта» иногда путают с понятием выгодного, а свойство «рецессивный» иногда путают с понятием «вредный», но явления различны. Доминирование описывает фенотип гетерозигот в отношении фенотипов гомозигот и вне зависимости от того, в какой степени различные фенотипы могут быть полезными или вредными. Поскольку многие аллели генетических заболеваний рецессивны и слово «доминирование» имеет положительный оттенок, часто делается предположение, что доминирующий фенотип превосходит приспособленность. Однако это не гарантировано; как обсуждается ниже, хотя большинство аллелей генетических заболеваний являются вредными и рецессивными, не все генетические заболевания являются рецессивными.
Тем не менее, эта путаница была повсеместной на протяжении всей истории генетики и сохраняется по сей день. Устранение этой путаницы было одной из главных причин публикации Принцип Харди – Вайнберга.
Молекулярные механизмы
Молекулярные основы доминирования были неизвестны Менделю. Теперь понятно, что локус гена включает длинный ряд (от сотен до тысяч) базы или же нуклеотиды из дезоксирибонуклеиновая кислота (ДНК) в определенной точке хромосомы. В центральная догма молекулярной биологии утверждает, что "ДНК делает РНК делает белок ", то есть ДНК записано сделать копию РНК, а РНК переведено сделать белок. В этом процессе разные аллели в локусе могут транскрибироваться или не транскрибироваться, а в случае транскрибирования могут транслироваться в несколько разные версии одного и того же белка (называемого изоформы ). Белки часто функционируют как ферменты которые катализируют химические реакции в клетке, прямо или косвенно производящие фенотипы. В любом диплоидном организме последовательности ДНК двух аллелей, присутствующих в любом локусе гена, могут быть идентичными (гомозиготными) или разными (гетерозиготными). Даже если локус гена гетерозиготен на уровне последовательности ДНК, белки, производимые каждым аллелем, могут быть идентичными. При отсутствии каких-либо различий между белковыми продуктами ни один из аллелей не может считаться доминантным (см. совместное доминирование, над). Даже если два белковых продукта немного отличаются (аллозимы ), вполне вероятно, что они продуцируют один и тот же фенотип в отношении действия ферментов, и снова ни один из аллелей не может считаться доминантным.
Потеря функции и гаплодостаточность
Доминирование обычно происходит, когда один из двух аллелей нефункционален на молекулярном уровне, то есть не транскрибируется или не производит функциональный белковый продукт. Это может быть результатом мутация который изменяет последовательность ДНК аллеля.[нужна цитата ] Организм, гомозиготный по нефункциональному аллелю, обычно будет демонстрировать отличительный фенотип из-за отсутствия белкового продукта. Например, у людей и других организмов непигментированная кожа альбинос фенотип[16] результаты, когда человек гомозиготен по аллелю, который кодирует нефункциональную версию фермента, необходимого для производства пигмента кожи меланин. Важно понимать, что не отсутствие функции позволяет описывать аллель как рецессивный: это взаимодействие с альтернативным аллелем в гетерозиготе. Возможны три основных типа взаимодействия:
- В типичном случае единственный функциональный аллель производит достаточное количество белка для получения фенотипа, идентичного фенотипу гомозиготы: это называется гаплодостаточность. Например, предположим, что стандартное количество фермента, продуцируемого функциональной гомозиготой, составляет 100%, при этом два функциональных аллеля вносят вклад по 50% каждый. Единственный функциональный аллель в гетерозиготе продуцирует 50% стандартного количества фермента, которого достаточно для получения стандартного фенотипа. Если гетерозигота и гомозигота с функциональным аллелем имеют идентичные фенотипы, функциональный аллель является доминантным по отношению к нефункциональному аллелю. Это происходит в локусе гена альбиноса: гетерозигота вырабатывает достаточно фермента, чтобы преобразовать предшественник пигмента в меланин, и индивидуум имеет стандартную пигментацию.
- Реже наличие единственного функционального аллеля дает фенотип, который не является нормальным, но менее тяжелым, чем фенотип нефункциональной гомозиготы. Это происходит, когда функциональный аллель не является гапло-достаточным. В этих случаях обычно применяются термины гапло-недостаточность и неполное доминирование. Промежуточное взаимодействие происходит там, где гетерозиготный генотип дает фенотип, промежуточный между двумя гомозиготами. В зависимости от того, на какую из двух гомозигот больше всего похожа гетерозигота, считается, что один аллель проявляет неполное доминирование над другим. Например, у людей Hb генный локус отвечает за белок бета-цепи (HBB ), который является одним из двух глобин белки, составляющие пигмент крови гемоглобин.[16] Многие люди гомозиготны по аллелю под названием HbА; некоторые люди несут альтернативный аллель, называемый HbS, либо в виде гомозигот, либо гетерозигот. Молекулы гемоглобина HbS/HbS гомозиготы претерпевают изменение формы, которое искажает морфологию красные кровяные тельца, и вызывает тяжелую, опасную для жизни форму анемия называется серповидноклеточная анемия. Лица гетерозиготные HbА/HbS для этого аллеля есть гораздо менее тяжелая форма анемии, называемая серповидно-клеточная характеристика. Поскольку фенотип заболевания HbА/HbS гетерозиготы больше похожи, но не идентичны HbА/HbА гомозигота, HbА аллель называется неполностью доминирующий к HbS аллель.
- Редко один функциональный аллель в гетерозиготе может продуцировать недостаточный генный продукт для любой функции гена, и фенотип напоминает фенотип гомозиготы для нефункционального аллеля. Это полное гаплонедостаточность очень необычно. В этих случаях нефункциональный аллель будет считаться доминантным по отношению к функциональному аллелю. Эта ситуация может возникнуть, когда нефункциональный аллель продуцирует дефектный белок, который мешает правильной функции белка, продуцируемого стандартным аллелем. Присутствие дефектного белка «доминирует» над стандартным белком, и фенотип заболевания гетерозиготы больше напоминает фенотип гомозиготы по двум дефектным аллелям. Термин «доминантный» часто неправильно применяется к дефектным аллелям, гомозиготный фенотип которых не исследован, но которые вызывают особый фенотип, будучи гетерозиготными с нормальным аллелем. Это явление встречается в ряде тринуклеотидный повтор болезни, одним из примеров является болезнь Хантингтона.[17]
Доминантно-отрицательные мутации
Многие белки обычно активны в форме мультимера, совокупности нескольких копий одного и того же белка, иначе известного как гомомультимерный белок или гомоолигомерный белок. Фактически, большинство из 83000 различных ферментов из 9800 различных организмов в базе данных ферментов BRENDA[18] представляют собой гомоолигомеры.[19] Когда версия белка дикого типа присутствует вместе с мутантной версией, может быть образован смешанный мультимер. Мутация, которая приводит к появлению мутантного белка, который нарушает активность белка дикого типа в мультимере, является доминантно-отрицательной мутацией.
Доминантно-отрицательная мутация может возникнуть в соматической клетке человека и обеспечить пролиферативное преимущество мутантной клетки, что приведет к ее клональной экспансии. Например, доминантно-отрицательная мутация в гене, необходимая для нормального процесса запрограммированной гибели клеток (Апоптоз ) в ответ на повреждение ДНК может сделать клетку устойчивой к апоптозу. Это позволит клону размножаться даже при чрезмерном повреждении ДНК. Такие доминантно-отрицательные мутации возникают в гене-супрессоре опухоли. p53.[20][21] Белок P53 дикого типа обычно присутствует в виде четырехбелкового мультимера (олиготетрамера). Доминантно-отрицательный p53 мутации возникают при различных типах рака и предраковых поражениях (например, опухолях головного мозга, раке груди, предраковых поражениях полости рта и раке полости рта).[20]
Доминантно-отрицательные мутации встречаются и в других генах-супрессорах опухолей. Например, две доминантно-отрицательные мутации зародышевой линии были идентифицированы в Атаксия, телеангиэктазия, мутировавшая (ATM) ген, повышающий предрасположенность к раку груди.[22] Доминантные отрицательные мутации фактора транскрипции C / EBPα может вызвать острый миелоидный лейкоз.[23] Унаследованные доминантно-отрицательные мутации также могут повышать риск других заболеваний, помимо рака. Доминантно-отрицательные мутации в Гамма-рецептор, активируемый пролифератором пероксисом (PPARγ) связаны с тяжелой инсулинорезистентностью, сахарным диабетом и гипертонией.[24]
Доминантно-отрицательные мутации были описаны не только у человека, но и у других организмов. Фактически, первое исследование, сообщающее о мутантный белок ингибирование нормальной функции белка дикого типа в смешанном мультимере происходило с белком хвостового волокна бактериофага Т4 GP37.[25] Мутации, которые производят укороченный белок, а не полноразмерный мутантный белок, по-видимому, имеют самый сильный доминантно-негативный эффект в исследованиях P53, ATM, C / EBPα и бактериофага T4 GP37.
Доминантные и рецессивные генетические заболевания человека
У людей многие генетические признаки или заболевания классифицируются просто как «доминантные» или «рецессивные». Особенно с так называемыми рецессивными заболеваниями, которые действительно являются фактором рецессивных генов, но могут чрезмерно упростить лежащую в их основе молекулярную основу и привести к неправильному пониманию природы доминирования. Например, рецессивное генетическое заболевание. фенилкетонурия (ФКУ)[26] является результатом любого из большого количества (> 60) аллелей в локусе гена для фермента фенилаланингидроксилаза (ПАУ).[27] Многие из этих аллелей практически не производят ПАУ, в результате чего подложка фенилаланин (Phe) и его побочные продукты метаболизма накапливаются в Центральная нервная система и может вызвать серьезные Интеллектуальная недееспособность если не лечить.
Чтобы проиллюстрировать эти нюансы, генотипы и фенотипические последствия взаимодействий между тремя гипотетическими аллелями PAH показаны в следующей таблице:[28]
| Генотип | ПАУ Мероприятия | [Phe] конц | ФКУ? |
|---|---|---|---|
| AA | 100% | 60 мкМ | Нет |
| AB | 30% | 120 мкМ | Нет |
| CC | 5% | 200 ~ 300 мкМ | Гиперфенилаланинемия |
| BB | 0.3% | 600 ~ 2400 мкМ | да |
У здоровых людей, гомозиготных по стандартному функциональному аллелю (AA), ПАУ активность стандартная (100%), а концентрация фенилаланина в крови [Phe] составляет около 60 мкМ (= мкмоль / л ). У нелеченных лиц, гомозиготных по одному из аллелей PKU (BB), ПАУ активность близка к нулю, от десяти до сорока раз превышает стандартную норму, и у человека проявляется фенилкетонурия.
в AB гетерозигота, ПАУ активность составляет всего 30% (не 50%) от нормы, кровь [Phe] is elevated two-fold, and the person does not manifest PKU. Таким образом А allele is dominant to the B allele with respect to PKU, but the B allele is incompletely dominant to the А allele with respect to its molecular effect, determination of ПАУ activity level (0.3% < 30% << 100%). Наконец, А allele is an incomplete dominant to B with respect to [Phe], as 60 μM < 120 μM << 600 μM. Note once more that it is irrelevant to the question of dominance that the recessive allele produces a more extreme [Phe] phenotype.
For a third allele C, а CC homozygote produces a very small amount of ПАУ enzyme, which results in a somewhat elevated level of [Phe] in the blood, a condition called гиперфенилаланинемия, which does not result in intellectual disability.
That is, the dominance relationships of any two alleles may vary according to which aspect of the phenotype is under consideration. It is typically more useful to talk about the phenotypic consequences of the allelic interactions involved in any genotype, rather than to try to force them into dominant and recessive categories.
Смотрите также
Рекомендации
- ^ "dominance". Oxford Dictionaries Online. Oxford University Press. Получено 14 мая 2014.
- ^ "express". Oxford Dictionaries Online. Oxford University Press. Получено 14 мая 2014.
- ^ Griffiths AJF; Gelbart WM; Miller JH; и другие. (1999). "Gene Interaction Leads to Modified Dihybrid Ratios". Modern Genetic Analysis. New York: W. H. Freeman & Company. ISBN 978-0-7167-3118-4.
- ^ Mayo, O. and Bürger, R. 1997. The evolution of dominance: A theory whose time has passed? "Biological Reviews", Volume 72, Issue 1, pp. 97–110
- ^ Bourguet, D. 1999. The evolution of dominance Наследственность, Volume 83, Number 1, pp. 1–4
- ^ Bagheri, H.C. 2006 г. Unresolved boundaries of evolutionary theory and the question of how inheritance systems evolve: 75 years of debate on the evolution of dominance "Journal of Experimental Zoology Part B: Molecular and Developmental Evolution", Volume 306B, Issue 4, pp. 329–359
- ^ Ridley, Matt (1999). "Болезнь". Геном: автобиография вида в 23 главах. Харпер Коллинз. pp. 136–146. ISBN 978-0-06-089408-5.
- ^ King, RC; и другие. (2006). Словарь по генетике (7-е изд.). Издательство Оксфордского университета. п. 129. ISBN 978-0-19-530761-0.
Dominance [refers] to alleles that fully manifest their phenotype when present in the гетерозиготный ... state.
- ^ Pennington, Sandra (1999). 11th Hour: Introduction to Genetics. Вайли. п. 43. ISBN 978-0-632-04438-2.
- ^ [1], Online 'Guidelines for nomenclature of genes, genetic markers, alleles, and mutations in mouse and rat'
- ^ [2], Online 'A standard for maize genetic nomenclature'
- ^ "Cat Coat Color". Veterinary Genetics Laboratory, University of California. Получено 2011-11-02.
- ^ Imes, D. L.; Geary, L. A.; Grahn, R. A.; Lyons, L. A. (April 2006). "Albinism in the domestic cat (Felis catus) ассоциируется с тирозиназа (TYR) mutation". Генетика животных. 37 (2): 175–8. Дои:10.1111/j.1365-2052.2005.01409.x. ЧВК 1464423. PMID 16573534.
- ^ Schmidt-Küntzel, A.; Eizirik, E .; O'Brien, S. J.; Menotti-Raymond, M. (April 2005). "Tyrosinase and tyrosinase related protein 1 alleles specify domestic cat coat color phenotypes of the albino and brown loci". Журнал наследственности. 96 (4): 289–301. Дои:10.1093/jhered/esi066. PMID 15858157.
- ^ Carr, Steven M. "Extensions to Mendelian Analysis". Мемориальный университет Ньюфаундленда.
- ^ а б Онлайн-менделевское наследование в человеке (OMIM): Albinism, oculocutaneous, type IA - 203100
- ^ Онлайн-менделевское наследование в человеке (OMIM): Huntington disease - 143100
- ^ Schomburg I; Chang A; Ebeling C; и другие. (Январь 2004 г.). "BRENDA, the enzyme database: updates and major new developments". Нуклеиновые кислоты Res. 32 (Database issue): D431–3. Дои:10.1093/nar/gkh081. ЧВК 308815. PMID 14681450.
- ^ Hashimoto K; Nishi H; Bryant S; Panchenko AR (June 2011). "Caught in self-interaction: evolutionary and functional mechanisms of protein homooligomerization". Phys Biol.. 8 (3): 035007. Bibcode:2011PhBio...8c5007H. Дои:10.1088/1478-3975/8/3/035007. ЧВК 3148176. PMID 21572178.
- ^ а б Marutani M; Tonoki H; Tada M; и другие. (Октябрь 1999 г.). "Dominant-negative mutations of the tumor suppressor p53 relating to early onset of glioblastoma multiforme". Рак Res. 59 (19): 4765–9. PMID 10519380.
- ^ Goh AM; Coffill CR; Lane DP (January 2011). "The role of mutant p53 in human cancer". Дж. Патол. 223 (2): 116–26. Дои:10.1002/path.2784. PMID 21125670. S2CID 23998813.
- ^ Chenevix-Trench G; Spurdle AB; Gatei M; и другие. (Февраль 2002 г.). "Dominant negative ATM mutations in breast cancer families". J. Natl. Институт рака. 94 (3): 205–15. Дои:10.1093/jnci/94.3.205. PMID 11830610.
- ^ Pabst T; Mueller BU; Zhang P; и другие. (Март 2001 г.). "Dominant-negative mutations of CEBPA, encoding CCAAT/enhancer binding protein-alpha (C/EBPalpha), in acute myeloid leukemia". Nat. Genet. 27 (3): 263–70. Дои:10.1038/85820. PMID 11242107. S2CID 33788907.
- ^ Barroso I; Gurnell M; Crowley VE; и другие. (1999). "Dominant negative mutations in human PPARgamma associated with severe insulin resistance, diabetes mellitus and hypertension". Природа. 402 (6764): 880–3. Bibcode:1999Natur.402..880B. Дои:10.1038/47254. PMID 10622252. S2CID 4423555.
- ^ Bernstein H; Fisher KM (March 1968). "Dominance in bacteriophage T4D". Генетика. 58 (3): 307–18. ЧВК 1211863. PMID 5662621.
- ^ Онлайн-менделевское наследование в человеке (OMIM): Hyperphenylalaninemia, non-PKU mild - 261600
- ^ Онлайн-менделевское наследование в человеке (OMIM): Phenylalanine Hydroxylase; PAH - 612349
- ^ Carr, Steven M. "One Gene, One Enzyme". Мемориальный университет Ньюфаундленда.
- "On-line notes for Biology 2250 – Principles of Genetics". Мемориальный университет Ньюфаундленда.
- Онлайн-менделевское наследование в человеке (OMIM): Hemoglobin—Beta Locus; HBB - 141900 — Sickle-Cell Anemia
- Онлайн-менделевское наследование в человеке (OMIM): ABO Glycosyltransferase - 110300 — ABO blood groups