Мохообразный - Bryophyte

Мохообразные неформальная группа, состоящая из трех подразделения несосудистых наземных растений (эмбриофиты ): печеночники, роголистник и мхи.[1] Обычно они ограничены в размерах и предпочитают влажную среду обитания, хотя могут выжить и в более сухой среде.[2] Мохообразные насчитывают около 20 000 видов растений.[3][4] Мохообразные производят замкнутые репродуктивные структуры (гаметангии и спорангии), но не производят цветы или же семена. Они воспроизводятся через споры.[5] Мохообразные обычно считаются парафилетический группа, а не монофилетический группы, хотя некоторые исследования дали противоположные результаты. Независимо от их статуса, имя удобное и остается неформальным собирательным термином. Термин «мохообразный» происходит от Греческий βρύον, брайон "древесно-моховое, устрично-зеленое" и φυτόν, фитон "растение".
Терминология
Термин «мохообразные» впервые предложил Браун (1964).[нужна цитата ] Г. Смит поместил эту группу между водорослями и Pteridophyta.[6]
Функции
Определяющими чертами мохообразных являются:
- В их жизненных циклах преобладают гаметофит сцена
- Их спорофиты неразветвленные
- У них нет правды сосудистая ткань содержащий лигнин (хотя у некоторых есть специальные салфетки для транспортировки воды)[7]
Среда обитания
Мохообразные существуют в самых разных средах обитания. Их можно найти растущими в диапазоне температур (холодные арктические и жаркие пустыни), возвышенностях (от уровня моря до альпийских) и влажности (от сухих пустынь до влажных тропических лесов).[8]
Мохообразные могут расти там, где васкуляризированные растения не могут, потому что они не зависят от корней в плане поглощения питательных веществ из почвы. Мохообразные могут выжить на камнях и голой почве.[8]
Жизненный цикл
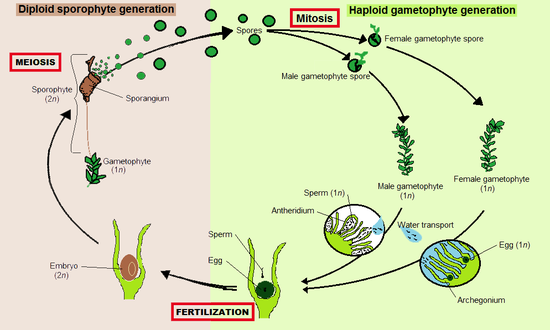
Как и все наземные растения (эмбриофиты), мохообразные имеют жизненные циклы с смена поколений.[9] В каждом цикле гаплоидный гаметофит, каждая из которых содержит фиксированное количество непарных хромосомы, чередуется с диплоид спорофит, клетка которого содержит два набора парных хромосом. Гаметофиты производят гаплоидные сперматозоиды и яйца, которые сливаются, образуя диплоидные зиготы, которые превращаются в спорофиты. Спорофиты продуцируют гаплоидные споры мейоз, которые превращаются в гаметофиты.
Мохообразные являются гаметофитами с преобладанием[10] Это означает, что более заметным и долгоживущим растением является гаплоидный гаметофит. Диплоидные спорофиты появляются только изредка и остаются прикрепленными к гаметофиту и питательно зависимыми от него.[11] У мохообразных спорофиты всегда неразветвленные и образуют один спорангий (капсула, продуцирующая споры), но каждый гаметофит может дать начало нескольким спорофитам одновременно.
В трех группах спорофит развивается по-разному. И у мхов, и у роголистников есть меристема зона, где происходит деление клеток. У роголистников меристема начинается у основания, где заканчивается ступня, и деление клеток подталкивает тело спорофита вверх. У мхов меристема расположена между капсулой и верхушкой стебля (щетинки) и продуцирует клетки вниз, удлиняя стебель и поднимая капсулу. У печеночников меристема отсутствует, а удлинение спорофита почти полностью обусловлено размножением клеток.[12]
Печеночники, мхи и роголистники большую часть своей жизни проводят в качестве гаметофитов. Гаметангия (органы, производящие гаметы), архегония и антеридия, образуются на гаметофитах, иногда на концах побегов, в пазухах листьев или скрыты под слоевищами. Некоторые мохообразные, например печеночник Маршанция, создают сложные структуры, несущие гаметангии, которые называются гаметангиофорами. Сперматозоиды жгутируются и должны плавать от антеридий, производящих их, к архегониям, которые могут находиться на другом растении. Членистоногие могут помочь в передаче спермы.[13]
Оплодотворенные яйца становятся зиготами, которые развиваются в зародыши спорофитов внутри архегоний. Зрелые спорофиты остаются прикрепленными к гаметофиту. Они состоят из стебля, называемого щетинкой, и одного спорангия или капсулы. Внутри спорангия гаплоидные споры производятся мейоз. Они распространяются, чаще всего ветром, и, если они приземляются в подходящей среде, могут развиться в новый гаметофит. Таким образом, мохообразные распространяются за счет комбинации плавающих сперматозоидов и спор, аналогично ликофиты, папоротники и другие криптогамы.
Сексуальность
Расположение антеридия и архегония на отдельном мохообразном растении обычно постоянна в пределах одного вида, хотя у некоторых видов она может зависеть от условий окружающей среды. Основное деление происходит между видами, у которых антеридии и архегонии встречаются на одном и том же растении, и видами, у которых они встречаются на разных растениях. Период, термин однообразный может использоваться, если антеридии и архегонии встречаются на одном и том же гаметофит и срок двояковыпуклый где они встречаются на разных гаметофитах.[14]
В семенные растения, "однодомный "используется там, где цветы с пыльники (микроспорангии) и цветы с семяпочки (мегаспорангии) возникают на одном спорофит и "раздельнополый «где они встречаются на разных спорофитах. Эти термины иногда могут использоваться вместо« однодомный »и« двудомный »для описания мохообразных гаметофитов.« Однодомный »и« однодомный »оба произошли от греческого слова« один дом »,« двудомный » и "двудомный" от греческого слова "два дома". Использование терминологии "oicy" относится к гаметофитной сексуальности мохообразных в отличие от спорофитной сексуальности семенных растений.[14]
Одноцветные растения обязательно гермафродиты, то есть одно и то же растение имеет оба пола.[14] Точное расположение антеридий и архегониев у одноядных растений варьируется. Они могут иметь разные побеги (аутогенные или самовлюбленный ), на одном побеге, но не вместе в общей структуре (околоушной или околоушной) или вместе в общем «соцветии» (синоевой или синодезной).[14][15] Двуядные растения однополый, что означает, что одно и то же растение имеет только один пол.[14] Все четыре паттерна (аутоический, околоушный, синойный и двояковыпуклый) встречаются у видов рода мхов. Брюм.[15]
Классификация и филогения


Традиционно все живые наземные растения без сосудистой ткани относили к одной таксономической группе, часто разделение (или тип). В последнее время, филогенетический исследования поставили под вопрос, образуют ли мохообразные монофилетическая группа и, следовательно, должны ли они образовывать единый таксон. Хотя исследование 2005 г. поддержало традиционную точку зрения о том, что мохообразные образуют монофилетическую группу,[16] к 2010 г. среди систематиков сложился широкий консенсус в отношении того, что мохообразные в целом не являются естественной группой (т.е. парафилетический ), хотя каждая из трех существующих (живых) групп является монофилетической.[17][18][19]
Три мохообразных клады (который можно рассматривать как подразделения ) являются Мархантиофита (печеночники), Мохообразные (мхи) и Anthocerotophyta (роголистник).[20] В сосудистые растения или трахеофиты образуют четвертую, безрейтинговую кладу наземных растений, называемую "Полиспорангиофита ". В этом анализе роголистник является сестрой сосудистых растений, а печеночники - сестрой всех других наземных растений, включая роголистник и мхи.[19][21] Филогенетические исследования продолжают давать противоречивые результаты. В частности, те, которые основаны на последовательностях генов, предполагают, что мохообразные являются парафилетическими, тогда как те, которые основаны на трансляции аминокислот тех же генов, предполагают, что они являются монофилетическими. Исследование 2014 года пришло к выводу, что эти различия были вызваны систематическими ошибками и что мохообразные являются монофилетическими.[22] Вопрос остается нерешенным.
Парафилетический вид
| мохообразные |

Если учесть вымершие растения, картина немного изменится. Некоторые вымершие наземные растения, такие как горнеофиты, не являются мохообразными, но также и не сосудистыми растениями, потому что, как и мохообразные, они не имеют настоящей сосудистой ткани. Требуется другое различие. У мохообразных спорофит представляет собой простую неразветвленную структуру с одним спорообразующим органом (спорангий ). У всех других наземных растений полиспорангиофиты спорофит разветвленный и несет множество спорангиев.[23][24] Утверждалось, что этот контраст между мохообразными и другими наземными растениями менее вводит в заблуждение, чем традиционный контраст между несосудистыми и сосудистыми растениями, поскольку многие мхи имеют хорошо развитые водопроводящие сосуды.[25] Контраст показан на немного другой кладограмме:[26]
| наземные растения |
| |||||||||||||||||||||||||||
Термин «мохообразный», таким образом, относится к степени родословных, определяемой в первую очередь тем, чего им не хватает. По сравнению с другими живыми наземными растениями у них отсутствует сосудистая ткань, содержащая лигнин, и разветвленные спорофиты, несущие множественные спорангии. Важность гаметофита в жизненном цикле также является общей чертой трех линий мохообразных (все существующие сосудистые растения являются доминирующими спорофитами).
Другие взгляды
Альтернативная филогения, основанная на аминокислотах, а не на генах, показывает мохообразные как монофилетическую группу:[22]
| эмбриофиты |
| ||||||||||||||||||
Если эта филогения окажется верной, то сложный спорофит живых сосудистых растений мог развиться независимо от более простого неразветвленного спорофита, присутствующего у мохообразных.[22] Другие исследования предполагают монофилетическую группу, состоящую из печеночников и мхов, причем роголистник является сестрой сосудистых растений.[27]
Эволюция
Вероятно, было несколько различных событий земной трансформации, в которых первоначально водные организмы колонизировали землю, как раз в пределах линии происхождения Viridiplantae.[28] Однако между 510-630 миллионами лет назад наземные растения произошли от водных растений, в частности зеленые водоросли. Молекулярно-филогенетические исследования показывают, что мохообразные - самые ранние расходящиеся линии современных наземных растений.[29][1][30][31] Они дают представление о миграции растений из водной среды на сушу. Ряд физических свойств связывает мохообразных как с наземными, так и с водными растениями.
Сходства с водорослями и сосудистыми растениями
Зеленые водоросли, мохообразные и сосудистые растения содержат хлорофиллы a и b, и структура хлоропластов схожа.[32] Подобно зеленым водорослям и наземным растениям, мохообразные также производят крахмал, хранящийся в пластидах, и содержат целлюлозу в своих стенках.[32] Отчетливая адаптация, наблюдаемая у мохообразных, позволила растениям колонизировать земную среду Земли. Чтобы предотвратить высыхание тканей растения в земной среде, может присутствовать восковая кутикула, покрывающая мягкие ткани растения, обеспечивающая защиту. У роголистников и мхов устьица обеспечивают газообмен между атмосферой и системой внутреннего межклеточного пространства. Развитие гаметангии обеспечило дополнительную защиту специально для гамет, зиготы и развивающегося спорофита.[33] Мохообразные и сосудистые растения (эмбриофиты ) также имеют эмбриональное развитие, чего не наблюдается у зеленых водорослей.[32] Хотя у мохообразных нет по-настоящему васкуляризированной ткани, у них есть органы, которые специализируются на определенных функциях, аналогичных, например, функциям листьев и стеблей наземных сосудистых растений.[32]
Для размножения и выживания мохообразных необходима вода. Вместе с папоротником и ликофиты, тонкий слой воды необходим на поверхности растения, чтобы обеспечить движение жгутиковых сперматозоидов между гаметофитами и оплодотворением яйцеклетки.[33]
Сравнительная морфология
Резюме морфологических характеристик гаметофиты из трех групп мохообразных:
| Печеночники | Мхи | Роголистники | |
|---|---|---|---|
| Структура | Таллоид или листовой | Foliose | Таллоид |
| Симметрия | Дорсивентральный или же радиальный | Радиальный | Дорсивентральный |
| Ризоиды | Одноклеточный | Многоклеточный | Одноклеточный |
| Хлоропластов / клетка | Много | Много | Один |
| Протонемы | Уменьшенный | Подарок | Отсутствующий |
| Гаметангия (антеридия и архегония ) | Поверхностный | Поверхностный | Погруженный |
Резюме морфологических характеристик спорофиты из трех групп мохообразных:
| Печеночники | Мхи | Роголистники | |
|---|---|---|---|
| Устьица | Отсутствующий | Подарок | Подарок |
| Структура | Маленький, без хлорофилла | Большой, с хлорофиллом | Большой, с хлорофиллом |
| Упорство | Эфемерный | Настойчивый | Настойчивый |
| Рост | Определенный | Определенный | Непрерывный |
| Seta | Подарок | Подарок | Отсутствующий |
| Капсула форма | Простой | Дифференцированный (жаберная крышка, перистом) | Удлиненный |
| Созревание спор | Одновременный | Одновременный | Выпускник |
| Распространение спор | Elaters | Перистомные зубы | Псевдоэлатеры |
| Колумелла | Отсутствующий | Подарок | Подарок |
| Расхождение | Продольные или неровные | Поперечный | Продольный |
Использует
Относящийся к окружающей среде
- Кондиционирование почвы
- Биоиндикаторы
- Моховые сады
- Пестициды
Характеристики мохообразных делают их полезными для окружающей среды. Было показано, что в зависимости от конкретной текстуры растения мохообразные помогают улучшить удержание воды и воздушное пространство в почве.[34] Мохообразные используются в исследованиях загрязнения, чтобы указать на загрязнение почвы (например, присутствие тяжелых металлов), загрязнение воздуха и УФ-В излучение.[34] Сады в Японии созданы из мха, чтобы создать умиротворяющие убежища.[34] Было обнаружено, что некоторые мохообразные производят естественные пестициды. Печеночник, Плагиохила, производит химическое вещество, ядовитое для мышей.[34] Другие мохообразные производят химические вещества, которые являются антифедантами, которые защищают их от поедания слизнями.[34] Когда Фитий сфагнум опрыскивается почвой прорастающих семян, он подавляет рост «гашения грибка», который иначе убил бы молодые всходы.[35]

Коммерческий
- Топливо
- Упаковка
- Перевязочный материал для ран
Торф это топливо, производимое из сушеных мохообразных, обычно Сфагнум. Антибиотические свойства и способность мохообразных удерживать воду делают их полезным упаковочным материалом для овощей, цветов и луковиц.[34] Кроме того, благодаря своим антибиотическим свойствам, Сфагнум использовалась как хирургическая повязка во время Первой мировой войны.[34]
Смотрите также
- Anthocerotophyta (роголистник)
- Мохообразные (мхи)
- Эмбриофит
- Мархантиофита (печеночники)
- Сексуальность растений
- Список британских графств и местных флор мохообразных
Рекомендации
- ^ а б Хеджес, С. Блэр (ноябрь 2002 г.). «Происхождение и эволюция модельных организмов». Природа Обзоры Генетика. 3 (11): 838–849. Дои:10.1038 / nrg929. PMID 12415314. S2CID 10956647.
- ^ Леветин, Эстель; МакМахон, Карен (2012). Растения и общество. Нью-Йорк, штат Нью-Йорк: Макгроу-Хилл. п. 139. ISBN 978-0-07-352422-1.
- ^ «Мохообразные (мхи и печеночники) - Список растений». www.theplantlist.org. Получено 2017-04-11.
- ^ «Что такое мохообразные». Университет Южного Иллинойса Карбондейл.
- ^ Вандерпуртен, Ален; Гоффине, Бернар (2009). Введение в мохообразные. Кембридж: Издательство Кембриджского университета. п. 3. ISBN 978-0-511-54013-4.
- ^ Смит, Г. (1955). Криптогамная ботаника. 2 (2-е изд.). Нью-Йорк: Макгроу-Хилл.
- ^ Лукас, Уильям Дж .; Грувер, Эндрю; Лихтенбергер, Рафаэль; Фурута, Каори; Ядав, Шри-Рам; Хелариутта, Юка; Хэ, Синь-Цян; Фукуда, Хироо; Канг, Джули; Brady, Siobhan M .; Патрик, Джон В. (апрель 2013 г.). «Сосудистая система растений: эволюция, развитие и функции F». Журнал интегративной биологии растений. 55 (4): 294–388. Дои:10.1111 / jipb.12041. HDL:10261/76903. PMID 23462277.
- ^ а б «Среда обитания - экология - мохообразные». www.anbg.gov.au. Получено 2017-04-12.
- ^ Лигроне, Роберто; Дакетт, Джеффри Дж .; Ренцалья, Карен С. (апрель 2012 г.). «Основные переходы в эволюции ранних наземных растений: бриологическая перспектива». Анналы ботаники. 109 (5): 851–871. Дои:10.1093 / aob / mcs017. ЧВК 3310499. PMID 22356739.
- ^ CM Шон Кэррингтон (4 ноября 2013 г.). «Мохообразные». Получено 2020-03-05.
- ^ Джонсон, М. Дж .; Шоу, А. Дж. (24 февраля 2016 г.). «Влияние количественной плодовитости на гаплоидной стадии на репродуктивный успех и диплоидную приспособленность водного торфяного мха Sphagnum macrophyllum». Наследственность. 116 (6): 523–530. Дои:10.1038 / hdy.2016.13. ЧВК 4868265. PMID 26905464.
- ^ Развитие и эволюция растений
- ^ Cronberg, N .; Начева, Р .; Хедлунд, К. (2006). «Микроартроподы опосредуют передачу спермы во мхах». Наука. 313 (5791): 1255. Дои:10.1126 / science.1128707. PMID 16946062. S2CID 11555211.
- ^ а б c d е Глим, Дж. М., Бисанг, И. (2014). «Сексуальность: ее определение (гл. 3-1)» (PDF). В Glime, J.M. (ред.). Экология мохообразных. Том 1 Физиологическая экология. Мичиганский технологический университет и Международная ассоциация бриологов. Получено 2014-11-09.
- ^ а б Уотсон, Э. (1981). Британские мхи и печеночники (3-е изд.). Издательство Кембриджского университета. п. 7. (Watson использует термины «oecy», а не «oicy».)
- ^ Горемыкин, В. В., Хеллвиг, Ф. Х. (2005). «Доказательства наиболее базального разделения наземных растений, делящих линии мохообразных и трахеофитных». Систематика и эволюция растений. 254 (1–2): 93–103. Дои:10.1007 / s00606-005-0337-1. S2CID 41403901.
- ^ Конрат, М .; Shaw, A.J .; Ренцалья, К. (2010). "Специальный выпуск Фитотакса посвящено мохообразным: ближайшим живым родственникам ранних наземных растений ». Фитотакса. 9: 5–10. Дои:10.11646 / phytotaxa.9.1.3.
- ^ Троицкий, А. В .; Игнатов, М. С .; Боброва, В. К .; Милютина И. А. (декабрь 2007 г.). «Вклад геносистематики в современные концепции филогении и классификации мохообразных». Биохимия (Москва). 72 (12): 1368–1376. Дои:10.1134 / с0006297907120115. PMID 18205621. S2CID 13509400.
- ^ а б Кноп, Волкер (31 декабря 2010 г.). «Ищем смысл в чепухе: краткий обзор некодирующей органеллярной ДНК, проливающий свет на филогению мохообразных». Разнообразие и эволюция мохообразных. 31 (1): 51–60. Дои:10.11646 / bde.31.1.10.
- ^ «ГЛОССАРИЙ B». Архивировано из оригинал на 2009-04-02. Получено 2009-03-26.
- ^ Цю, Инь-Лун; Ли, Либо; Ван, Бин; Чен, Чжидуань; Knoop, Volker; Грот-Малонек, Милена; Домбровская, Елена; Ли, Чонхо; Кент, Ливия; Отдыхай, Джошуа; Истабрук, Джордж Ф .; Хендри, Тори А .; Тейлор, Дэвид В .; Testa, Christopher M .; Амброс, Мэтью; Крэндалл-Стотлер, Барбара; Дафф, Р. Джоэл; Стеч, Майкл; Фрей, Вольфганг; Квандт, Дитмар; Дэвис, Чарльз К. (17 октября 2006 г.). «Самые глубокие расхождения в наземных растениях, выведенные из филогеномных данных». Труды Национальной академии наук Соединенных Штатов Америки. 103 (42): 15511–15516. Bibcode:2006PNAS..10315511Q. Дои:10.1073 / pnas.0603335103. ЧВК 1622854. PMID 17030812.
- ^ а б c Кокс, Саймон Дж .; Ли, Блэз; Фостер, Питер Дж .; Эмбли, Т. Мартин; Civá, Питер (2014). «Конфликтные филогении ранних наземных растений вызваны систематическими ошибками среди синонимичных замен». Систематическая биология. 63 (2): 272–279. Дои:10.1093 / sysbio / syt109. ЧВК 3926305. PMID 24399481.
- ^ Кенрик, Пол и Крейн, Питер Р. (1997a). Происхождение и ранняя диверсификация наземных растений: кладистическое исследование. Вашингтон, округ Колумбия: Smithsonian Institution Press. ISBN 978-1-56098-730-7.
- ^ Кенрик П. и Крейн П. Р. (1997b). «Происхождение и ранняя эволюция растений на суше». Природа. 389 (6646): 33–39. Bibcode:1997 Натур.389 ... 33К. Дои:10.1038/37918. S2CID 3866183.
- ^ Белл, Н. Э. и Хивёнен, Дж. (2010). «Филогения мхов класса Polytrichopsida (BRYOPHYTA): структура общего уровня и неконгруэнтные генные деревья». Молекулярная филогенетика и эволюция. 55 (2): 381–398. Дои:10.1016 / j.ympev.2010.02.004. PMID 20152915.
- ^ Крейн, Питер Р .; Херендин, Патрик; Фриис, Эльза Мари (октябрь 2004 г.). «Ископаемые и филогения растений». Американский журнал ботаники. 91 (10): 1683–1699. Дои:10.3732 / ajb.91.10.1683. PMID 21652317.
- ^ Karol, Kenneth G .; Арумуганатан, Катираветпиллай; Boore, Джеффри Л .; Даффи, Аарон М .; Эверетт, Карин DE; Холл, Джон Д .; Hansen, S.K .; Kuehl, Jennifer V .; Мандоли, Дина Ф .; Mishler, Brent D .; Olmstead, Ричард Дж .; Ренцалья, Карен С. и Вольф, Пол Г. (2010). «Полные пластомные последовательности Equisetum arvense и Isoetes flaccida: значение для филогении и эволюции пластидного генома ранних линий наземных растений». BMC Эволюционная биология. 10 (1): 321. Дои:10.1186/1471-2148-10-321. ЧВК 3087542. PMID 20969798.
- ^ Жерриенн, Филипп; Серве, Томас; Веколи, Марко (апрель 2016 г.). «Эволюция растений и террестриализация в палеозойские времена - филогенетический контекст». Обзор палеоботаники и палинологии. 227: 4–18. Дои:10.1016 / j.revpalbo.2016.01.004.
- ^ Конрат, М .; Shaw, A.J .; Ренцалья, К. (2010). "Специальный выпуск Фитотакса посвящено мохообразным: ближайшим живым родственникам ранних наземных растений ». Фитотакса. 9: 5–10. Дои:10.11646 / phytotaxa.9.1.3.
- ^ Karol, Kenneth G .; Арумуганатан, Катираветпиллай; Boore, Джеффри Л .; Даффи, Аарон М .; Эверетт, Карин DE; Холл, Джон Д .; Hansen, S.K .; Kuehl, Jennifer V .; Мандоли, Дина Ф .; Mishler, Brent D .; Olmstead, Ричард Дж .; Ренцалья, Карен С. и Вольф, Пол Г. (2010). «Полные пластомные последовательности Equisetum arvense и Isoetes flaccida: значение для филогении и эволюции пластидного генома ранних линий наземных растений». BMC Эволюционная биология. 10 (1): 321. Дои:10.1186/1471-2148-10-321. ЧВК 3087542. PMID 20969798.
- ^ Шоу, А. Джонатан; Сёвеньи, Петер; Шоу, Бланка (март 2011). «Разнообразие и эволюция мохообразных: окна в раннюю эволюцию наземных растений». Американский журнал ботаники. 98 (3): 352–369. Дои:10.3732 / ajb.1000316. PMID 21613131.
- ^ а б c d Эверет, Рэй; Эйххорн, Сьюзен (2013). Биология растений. W.H. Freeman and Company Publishers.
- ^ а б Перселл, Адам. «Мохообразные». Основы биологии.
- ^ а б c d е ж грамм Glime, Дженис. «Экономическое и этническое использование мохообразных» (PDF). harvard.edu.
- ^ Вольфехель, Х. (апрель 1988 г.). «Подавляющее действие сфагнового торфа на Pythium spp». Acta Horticulturae (221): 217–222. Дои:10.17660 / actahortic.1988.221.22.
Библиография
- Lesica, P .; McCune, B .; Купер, С. В .; Хонг, В. С. (1991). «Различия в сообществах лишайников и мохообразных между старовозрастными и управляемыми второстепенными лесами в Лебединой долине, штат Монтана». Канадский журнал ботаники. 69 (8): 1745–1755. Дои:10.1139 / b91-222.
внешняя ссылка
- Участок Андреевского мха Фото мохообразных
- 27-мая-2013 Возродились вековые замороженные растения, 400-летние экземпляры мохообразных, оставленные отступающие ледники в Канаде возвращаются к жизни в лаборатории.
- Фарж, Катрин Ла; Уильямс, Криста Х .; Англия, Джон Х. (22 мая 2013 г.). «Регенерация мохообразных малой ледниковой эпохи, выходящих из полярного ледника с последствиями тотипотентности в экстремальных условиях». Труды Национальной академии наук. 110 (24): 9839–9844. Bibcode:2013ПНАС..110.9839Л. Дои:10.1073 / pnas.1304199110. ЧВК 3683725. PMID 23716658.
- Мэджилл, Р. Э., изд. (1990). Glossarium polyglottum bryologiae. Многоязычный глоссарий по бриологии. Монографии по систематической ботанике из ботанического сада Миссури, т. 33, 297 стр. Онлайн-версия: Интернет-архив.
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Растение группы | |||||||||||
| |||||||||||
| |||||||||||
| Рост и привычка растений | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||
| |||||||||||
| Авторитетный контроль |
|---|