Удобрение - Fertilisation

Удобрение или оплодотворение (увидеть орфографические различия ), также известен как генеративное оплодотворение, осеменение, опыление,[1] оплодотворение, сингамия и пропитка,[2] это слияние гаметы дать начало новому индивидуальному организму или потомству и инициировать его развитие. Этот цикл оплодотворения и развития новых особей называется половое размножение. В течение двойное оплодотворение в покрытосеменные то гаплоидный мужская гамета сочетается с двумя гаплоидными полярные ядра сформировать триплоид первичный эндосперм ядро в процессе вегетативного оплодотворения.
История
В древности Аристотель задумал формирование новых индивидуумов путем слияния мужской и женской флюидов, с постепенным появлением формы и функций в режиме, названном им как эпигенетический.[3]
В 1784 г. Спалланцани установили необходимость взаимодействия между яйцеклеткой самки и спермой самца с образованием зиготы у лягушек.[4] В 1827 году фон Бэр заметил терианец яйцо млекопитающих впервые.[3] Оскар Хертвиг (1876) в Германии описали слияние ядер сперматозоидов и яйцеклеток из морской еж.[4]
Эволюция
Эволюция оплодотворения связана с происхождение мейоза, поскольку оба являются частью половое размножение, возникла в эукариоты. Существуют две противоречивые теории о том, как возникла пара мейоз – оплодотворение. Во-первых, он произошел от прокариотического секса (бактериальная рекомбинация ), поскольку эукариоты произошли от прокариот.[нужна цитата ] Другой заключается в том, что митоз возник мейоз.[5]
Удобрение растений

В оплодотворении растений участвуют гаметы: пыльца (мужская) и яйцеклетка (женская). Различные семейства растений имеют разные методы оплодотворения женского гаметофита. В Мохообразный наземные растения, удобрение происходит в архегоний. В цветущие растения во втором случае оплодотворения участвует другая сперматозоид и центральная ячейка которая является второй женской гаметой. У цветковых растений от каждого пыльцевого зерна по два сперматозоида.
В семенные растения, после опыление, а пыльца зерно прорастает, а пыльцевая трубка растет и проникает в яйцеклетка через крошечную пору, называемую микропиле Сперма передается от пыльцы через пыльцевую трубку к семяпочек.
Рост пыльцевой трубки
В отличие от сперматозоидов животных, которые являются подвижными, сперматозоиды растений неподвижны и через пыльцевую трубку переносятся к яйцеклетке, где выделяется сперма.[6] Пыльцевая трубка проникает в клеймо и удлиняется через внеклеточный матрикс стиля, прежде чем достигнет яичника. Затем возле розетки он пробивает яйцеклетка через микропиле (отверстие в стенке семяпочки) пыльцевая трубка «врывается» в зародышевый мешок, высвобождая сперму.[7] Считалось, что рост пыльцевой трубки зависит от химических сигналов от пестика, однако эти механизмы были плохо изучены до 1995 года. табачные растения выявил семью гликопротеины называемые белками TTS, которые усиливают рост пыльцевых трубок.[7] Пыльцевые трубки в среде для прорастания пыльцы без сахара и в среде с очищенными белками TTS росли. Однако в среде TTS пробирки росли в 3 раза быстрее, чем в среде без сахара.[7] Белки TTS также были размещены в различных местах полуфабриката. в вево опыленные пестики и пыльцевые трубки сразу же простирались к белкам. Трансгенные растения, лишенные способности продуцировать белки TTS, демонстрировали более медленный рост пыльцевых трубок и снижение фертильности.[7]
Разрыв пыльцевой трубки
Разрыв пыльцевой трубки для выпуска спермы в Арабидопсис было показано, что он зависит от сигнала от женского гаметофита. Конкретные белки, называемые протеинкиназами FER, присутствующие в яйцеклетке, контролируют производство высокореактивных производных кислорода, называемых активные формы кислорода (ROS). Уровни АФК показаны через GFP быть наивысшим на стадии цветения, когда семяпочка является наиболее восприимчивой к пыльцевым трубкам, и наименьшей во время развития и после оплодотворения.[6] Высокое количество АФК активирует каналы ионов кальция в пыльцевой трубке, заставляя эти каналы поглощать ионы кальция в больших количествах. Это повышенное поглощение кальция вызывает разрыв пыльцевой трубки и высвобождение ее сперматозоидов в яйцеклетку.[6] Анализы питания пестиков, в которых растения получали дифенилйодонийхлорид (DPI), подавляли концентрации ROS в Арабидопсис, что, в свою очередь, предотвратило разрыв пыльцевой трубки.[6]
Мохообразные
Мохообразные - это традиционное название, используемое для обозначения всех эмбриофитов (наземных растений), которые не имеют истинной сосудистой ткани и поэтому называются «несосудистыми растениями». У некоторых мохообразных действительно есть специальные ткани для переноса воды; однако, поскольку они не содержат лигнин, они не считаются настоящей сосудистой тканью.
Папоротники
Папоротник входит в группу из примерно 12 000 видов сосудистых растений, которые размножаются спорами и не имеют ни семян, ни цветов. Они отличаются от мхов наличием сосудов (т.е. имеют сосуды, проводящие воду). У них есть стебли и листья, как и у других сосудистых растений. У большинства папоротников есть так называемые «головные уборы», которые разрастаются в листья, каждый из которых деликатно делится.
Голосеменные
Голосеменные - это группа семеноводческих растений, в которую входят хвойные, Cycads, Гинкго, и Gnetales. Термин «голосеменные» происходит от греческого сложного слова γυμνόσπερμος (γυμνός gymnos, «голый» и σπέρμα sperma, «семя»), что означает «голые семена», по названию незакрытого состояния их семян (называемых семяпочками в неоплодотворенном состоянии). Их обнаженное состояние контрастирует с семенами и семяпочками цветковых растений (покрытосеменных), которые заключены в завязь. Семена голосеменных развиваются либо на поверхности чешуек или листьев, часто видоизмененных в виде шишек, либо на концах коротких стеблей, как у гинкго.
Цветущие растения
После оплодотворения яичник начинает набухать и превращаться в фрукты.[8] У многосемянных плодов для сингамии с каждой семяпочкой необходимо несколько зерен пыльцы. Рост пыльцевой трубки контролируется вегетативной (или трубочной) цитоплазмой. Гидролитический ферменты выделяются пыльцевой трубкой, переваривающей женские ткани по мере того, как трубка опускается вниз по стигме и стилю; переваренная ткань используется в качестве источника питательных веществ для пыльцевой трубки по мере ее роста. Во время роста пыльцевой трубки по направлению к яичнику генеративное ядро делится с образованием двух отдельных ядер сперматозоидов (гаплоидное число хромосом).[9] - поэтому растущая пыльцевая трубка содержит три отдельных ядра, два сперматозоида и одну трубку.[10] Сперматозоиды взаимосвязаны и диморфны, у ряда растений большой сперматозоид также связан с ядром трубки, а связанные между собой сперматозоид и ядро трубки образуют «мужской зародышевый элемент».[11]
Двойное оплодотворение это процесс в покрытосеменные (цветковые растения) в которых два сперма из каждой пыльцевой трубки удобрять по два клетки в женском гаметофит (иногда называемый эмбриональным мешком), который находится внутри семяпочки. После того, как пыльцевая трубка попадает в гаметофит, ядро пыльцевой трубки распадается, и две сперматозоиды высвобождаются; одна из двух сперматозоидов удобрения яйцеклетка (внизу гаметофита возле микропиле), образующая диплоид (2n) зигота. Это момент, когда действительно происходит оплодотворение; опыление и оплодотворение - это два разных процесса. Ядро другой сперматозоиды сливается с двумя гаплоидными полярными ядрами (содержащимися в центральной клетке) в центре гаметофита. Полученная ячейка триплоид (3н). Эта триплоидная клетка делится на митоз и формирует эндосперм, а питательное вещество -богатые ткань, внутри семя.
Два материнских ядра центральной клетки (полярные ядра), которые вносят вклад в эндосперм, возникают в результате митоза из единственного мейотического продукта, который также дал начало яйцеклетке. Следовательно, материнский вклад в генетическую конституцию триплоидного эндосперма вдвое больше, чем вклад эмбриона.
Один примитивный вид цветкового растения, Nuphar polysepala, имеет диплоидный эндосперм, возникающий в результате слияния сперматозоидов с одним, а не с двумя материнскими ядрами. Считается, что на ранних этапах развития линий покрытосеменных в этом способе размножения имело место дупликация, в результате чего образовывались семиклеточные / восьмиядерные женские гаметофиты и триплоидные эндоспермы с соотношением материнского генома к отцовскому 2: 1.[12]
У многих растений развитие мякоти плода пропорционально проценту оплодотворенных семяпочек. Например, с арбуз необходимо доставить около тысячи зерен пыльцы и равномерно распределить ее по трем лепесткам рыльца, чтобы получить плод нормального размера и формы.
Перекрестное оплодотворение и самооплодотворение представляют собой разные стратегии с разными преимуществами и затратами. По оценкам, 48,7% видов растений являются двудомными или самонесовместимыми облигатными ауткроссами.[13] Также подсчитано, что около 42% цветковых растений в природе имеют смешанную систему спаривания.[14]
В наиболее распространенном виде смешанной системы скрещивания отдельные растения дают один тип цветка, а плоды могут содержать самоопыляющиеся, перекрестные или смешанные типы потомства. Переход от перекрестного оплодотворения к самооплодотворению является наиболее распространенным эволюционным переходом у растений и неоднократно происходил во многих независимых линиях.[15] Около 10-15% цветковых растений являются преимущественно самооплодотворяющимися.[15]
Самоопыление
При обстоятельствах, когда опылители или партнеры редки, самооплодотворение дает преимущество репродуктивная гарантия.[15] Таким образом, самооплодотворение может улучшить способность к колонизации. У некоторых видов самооплодотворение сохранялось на протяжении многих поколений. Capsella краснуха это самооплодотворяющийся вид, который стал самосовместимым от 50 000 до 100 000 лет назад.[16] Arabidopsis thaliana это преимущественно самоопыляющееся растение с пересечение ставка в дикой природе менее 0,3%;[17] исследование показало, что самооплодотворение возникло примерно миллион лет назад или более в A. thaliana.[18] У давно укоренившихся самоопыляющихся растений маскировка вредных мутации и производство генетической изменчивости происходит нечасто и, таким образом, маловероятно, что оно обеспечит достаточное преимущество для многих поколений для поддержания мейотического аппарата. Следовательно, можно было ожидать, что самооплодотворение в природе будет заменено амейотической бесполой формой размножения, которая будет менее затратной. Однако реальное сохранение мейоза и самооплодотворения как формы воспроизводства у давно укоренившихся самоопыляющихся растений может быть связано с непосредственной выгодой от эффективных рекомбинационная репарация повреждений ДНК при формировании половых клеток, обеспечиваемых мейозом в каждом поколении.[нужна цитата ]
Оплодотворение у животных
Механика оплодотворения широко изучена на морских ежах и мышах. Это исследование решает вопрос о том, как сперма и соответствующие яйцеклетки находят друг друга и вопрос, как только один сперматозоид попадает в яйцеклетку и доставляет ее содержимое. Есть три этапа оплодотворения, обеспечивающие видоспецифичность:
- Хемотаксис
- Активация спермы / акросомная реакция
- Прилипание сперматозоидов к яйцам
Внутренние и внешние
Рассмотрение того, использует ли животное (а точнее позвоночное) внутренний или внешнее оплодотворение часто зависит от способа рождения. Яйцекладущие животные откладывают яйца с толстой кальциевой скорлупой, например: куры, или толстая кожистая скорлупа, как правило, воспроизводятся посредством внутреннего оплодотворения, так что сперматозоиды оплодотворяют яйцеклетку без необходимости проходить через толстый защитный третичный слой яйца. Яйцекладущие и живородящий животные также используют внутреннее оплодотворение. Важно отметить, что хотя некоторые организмы размножаются через Amplexus, они все еще могут использовать внутреннее оплодотворение, как некоторые саламандры. Преимущества внутреннего оплодотворения включают: минимальную потерю гамет; больший шанс индивидуального оплодотворения яиц, относительно «более длительный» период защиты яиц и избирательное оплодотворение; многие самки обладают способностью хранить сперму в течение продолжительных периодов времени и могут оплодотворять свои яйцеклетки по собственному желанию.
С другой стороны, яйцекладущие животные, производящие яйца с тонкими третичными оболочками или вообще без оболочек, используют методы внешнего оплодотворения. Преимущества внешнего оплодотворения включают: минимальный контакт и передачу телесных жидкостей; снижение риска передачи болезней и большая генетическая изменчивость (особенно во время широковещательного нереста методами внешнего оплодотворения).
Морские ежи
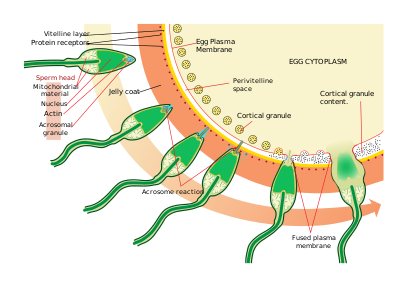
Сперма находит яйца через хемотаксис, тип взаимодействия лиганд / рецептор. Resact - это пептид из 14 аминокислот, очищенный от желеобразной оболочки A. punctulata что привлекает миграцию сперматозоидов.
Найдя яйцеклетку, сперма проникает в желейное пальто через процесс, называемый активацией спермы. При другом взаимодействии лиганд / рецептор олигосахаридный компонент яйца связывает и активирует рецептор на сперматозоиде и вызывает акросомная реакция. Акросомные пузырьки сперматозоидов сливаются с плазматической мембраной и высвобождаются. В этом процессе молекулы, связанные с мембраной акросомных везикул, такие как биндин, обнажаются на поверхности сперматозоидов. Это содержимое переваривает студенистую оболочку и, в конечном итоге, желточную оболочку. Помимо высвобождения акросомальных пузырьков, происходит взрывная полимеризация актин сформировать тонкую иглу на головке сперматозоида, называемую акросомный отросток.
Сперма связывается с яйцеклеткой через реакцию другого лиганда между рецепторами на желточная мембрана. Белок поверхности сперматозоидов биндин связывается с рецептором на желточной мембране, идентифицированным как EBR1.
Слияние плазматических мембран сперматозоидов и яйцеклеток, вероятно, опосредовано биндином. В месте контакта слияние вызывает образование конус удобрения.
Млекопитающие
Млекопитающие внутренне удобрять через совокупление. После мужчины эякулирует, многие сперматозоиды перемещаются в верхнюю часть влагалища (через сокращения из влагалища) через шейка матки и по всей длине матка встретить яйцеклетку. В случаях, когда происходит оплодотворение, самка обычно овулирует в течение периода, который длится от часов до совокупления до нескольких дней после; поэтому у большинства млекопитающих эякуляция чаще предшествует овуляции, чем наоборот.
Когда сперматозоиды откладываются в переднюю часть влагалища, они не способны к оплодотворению (т. Е. Не обладают способностью к оплодотворению) и характеризуются медленной линейной подвижностью. Эта подвижность в сочетании с мышечными сокращениями обеспечивает транспортировку спермы к матке и фаллопиевым трубам.[19] В микросреде женского репродуктивного тракта существует такой градиент pH, что pH около входа во влагалище ниже (примерно 5), чем у маточных труб (примерно 8).[20] Специфический для сперматозоидов pH-чувствительный транспортный белок кальция, называемый CatSper, увеличивает проницаемость сперматозоидов для кальция по мере его продвижения в репродуктивный тракт. Приток внутриклеточного кальция способствует увеличению емкости и гиперактивации сперматозоидов, вызывая более резкую и быструю нелинейную модель подвижности по мере приближения сперматозоидов к ооциту. В уполномоченный сперматозоид и ооцит встречаются и взаимодействуют в ампула из фаллопиевых труб. Реотаксис, термотаксис и хемотаксис - известные механизмы, которые направляют сперматозоиды к яйцеклетке на заключительном этапе миграции сперматозоидов.[21] Сперматозоиды отвечают (см. Термотаксис спермы ) к температурному градиенту ~ 2 ° C между яйцеводом и ампулой,[22] и хемотаксический градиенты прогестерон были подтверждены как сигнал, исходящий от кучевой оофорус клетки, окружающие ооциты кролика и человека.[23] Емкостные и гиперактивированные сперматозоиды реагируют на эти градиенты изменением своего поведения и движением к комплексу кумулюс-ооцит. Другие хемотаксические сигналы, такие как формил-Met-Leu-Phe (fMLF), также могут направлять сперматозоиды.[24]
В zona pellucida, толстый слой внеклеточного матрикса, который окружает яйцо и подобен желточной мембране у морских ежей, связывает сперму. В отличие от морских ежей, сперматозоиды связываются с яйцеклеткой до акросомной реакции. ZP3, гликопротеин в пеллюцидной оболочке, отвечает за адгезию яйцеклеток / сперматозоидов у мышей. Рецептор галактозилтрансфераза (GalT) связывается с остатками N-ацетилглюкозамина на ZP3 и важен для связывания со спермой и активации акросомной реакции. ZP3 достаточно, но не нужен для связывания сперматозоидов / яйцеклеток. Существуют два дополнительных рецептора сперматозоидов: белок 250 кДа, который связывается с секретируемым белком яйцевода, и SED1, который независимо связывается с зоной. Полагают, что после акросомной реакции сперматозоиды остаются связанными с блестящей оболочкой через открытые рецепторы ZP2. Эти рецепторы неизвестны у мышей, но были идентифицированы у морских свинок.
У млекопитающих связывание сперматозоида с GalT инициирует акросомная реакция. Этот процесс освобождает гиалуронидаза который переваривает матрицу гиалуроновая кислота в облачении вокруг ооцита. Кроме того, рядом с ооцитом выделяются гепариноподобные гликозаминогликаны (ГАГ), которые способствуют акросомной реакции.[25] Слияние ооцита плазматические мембраны и сперма следует и позволяет сперме ядро, типичный центриоль, и нетипичный центриоль который прикреплен к жгутик, но не митохондрии, чтобы войти в ооцит.[26] Протеин CD9 вероятно, опосредует это слияние у мышей (связывающий гомолог). Яйцо "активирует "сам по себе при слиянии с одним сперматозоидом и, таким образом, изменяет свою клеточную мембрану, чтобы предотвратить слияние с другими сперматозоидами. Цинк атомы высвобождаются во время этой активации.[нужна цитата ]
Этот процесс в конечном итоге приводит к образованию диплоид ячейка называется зигота. Зигота делится, образуя бластоциста и, войдя в матку, имплантаты в эндометрии, начало беременность. Эмбриональная имплантация не в матка стена приводит к внематочная беременность что может убить мать.
У таких животных, как кролики, половой акт вызывает овуляцию, стимулируя высвобождение гормона гипофиза гонадотропина; этот выпуск значительно увеличивает вероятность беременности.
Люди

Оплодотворение у человека - это союз человека яйцо и сперма, обычно встречающиеся в ампула маточной трубы, производя зигота клетка или оплодотворенная яйцеклетка, инициирующая пренатальное развитие. Ученые обнаружили динамику оплодотворения человека в девятнадцатом веке.
Период, термин зачатие обычно относится к «процессу беременности, включающему оплодотворение или имплантацию, или и то, и другое».[27] Его использование делает его предметом смысловые рассуждения о начале беременности, как правило, в контексте аборт дебаты. гаструляция, который происходит примерно через 16 дней после оплодотворения, имплантированная бластоциста развивает три зародышевых листка, энтодерму, эктодерму и мезодерму, и генетический код отца становится полностью вовлеченным в развитие эмбриона; позднее побратимство невозможно. Кроме того, межвидовые гибриды выживают только до гаструляции и не могут развиваться дальше. Однако в некоторой литературе по биологии развития человека упоминается концептус и такая медицинская литература упоминает «продукты зачатия» как постимплантационный эмбрион и окружающие его оболочки.[28] Термин «концепция» обычно не используется в научной литературе из-за его вариативного определения и коннотации.
Насекомые

Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Февраль 2015 г.) |
Насекомые в разных группах, в том числе Одоната (стрекозы и стрекозы ) и Перепончатокрылые (муравьи, пчелы, и осы ) практикуйте отложенное оплодотворение. У Odonata самки могут спариваться с несколькими самцами и хранить сперму до тех пор, пока не откладываются яйца. Самец может парить над самкой во время откладки яиц, чтобы предотвратить ее спаривание с другими самцами и замену его спермы; в некоторых группах, таких как дротики, самец продолжает хватать самку своими кламмерами во время откладки яиц, и пара летает в тандеме.[29] Среди социальных перепончатокрылых пчела матки спариваются только во время спаривания, в короткие сроки, длившиеся несколько дней; ферзь может спариваться с восемью или более дроны. Затем она хранит сперму до конца своей жизни, возможно, в течение пяти или более лет.[30][31]
Удобрение грибами
Во многих грибы (Кроме хитриды ), как и у некоторых протистов, оплодотворение - это двухэтапный процесс. Во-первых, цитоплазмы двух гаметных клеток сливаются (называемые плазмогамия ), производя дикариотический или гетерокариотический клетка с множеством ядер. Затем эта клетка может делиться с образованием дикариотических или гетерокариотических гифы. Второй этап оплодотворения - это кариогамия, слияние ядер с образованием диплоидной зиготы.
В Читрид У грибов оплодотворение происходит за один этап со слиянием гамет, как у животных и растений.
Оплодотворение у простейших
Оплодотворение простейшими
У простейших есть три типа процессов оплодотворения:[32]
Удобрение водорослями
Оплодотворение водорослей происходит за счет двойное деление. В псевдоподия сначала удаляется, и ядро начинает делиться. Когда цитоплазма делится, цитоплазма также делится на две равные части для каждой дочерней клетки. Две дочерние клетки производятся одной родительской клеткой. Это включает в себя процесс митоз.
Этот раздел пуст. Вы можете помочь добавляя к этому. (Сентябрь 2015 г.) |
Оплодотворение у грибовидных простейших
Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Март 2015 г.) |
Удобрение в грибах. У многих грибов (кроме хитридов), как и у некоторых простейших, оплодотворение представляет собой двухэтапный процесс. ... В Читрид У грибов оплодотворение происходит за один этап со слиянием гамет, как у животных и растений.
Оплодотворение и генетическая рекомбинация
Мейоз приводит к случайной сегрегации генов, вносимых каждым родителем. Каждый родительский организм обычно идентичен, за исключением части их генов; каждый гамета поэтому генетически уникален. При оплодотворении родительский хромосомы объединить. В люди, (2²²) ² = 17,6x1012 хромосомно разные зиготы возможны для неполовых хромосом, даже если хромосомный кроссовер. Если кроссовер происходит один раз, то в среднем (4²²) ² = 309x1024 генетически разные зиготы возможны для каждой пары, не считая, что события кроссовера могут иметь место в большинстве точек вдоль каждой хромосомы. Хромосомы X и Y не подвергаются перекрестным событиям[нужна цитата ] и поэтому исключаются из расчета. В митохондриальная ДНК наследуется только от материнского родителя.
Партеногенез
Организмы, которые обычно размножаются половым путем, также могут воспроизводиться через партеногенез, при этом неоплодотворенная женская гамета дает жизнеспособное потомство. Эти потомки могут быть клонами матери или в некоторых случаях генетически отличаться от нее, но унаследовать только часть ее ДНК. Партеногенез происходит у многих растений и животных и может быть вызван у других посредством химического или электрического раздражения яйцеклетки. В 2004 г. японские исследователи под руководством Томохиро Коно удалось после 457 попыток объединить яйцеклетка двух мышей, блокируя определенные белки, которые обычно предотвращают такую возможность; в результате эмбрион обычно развивался в мышь.[35]
Аллогамия и автогамия
Аллогамия, которое также известно как перекрестное оплодотворение, относится к оплодотворению яйцеклетки одного человека мужской гаметой другого.
Автогамия, также известная как самооплодотворение, встречается у таких гермафродитных организмов, как растения и плоские черви; в нем две гаметы от одного индивидуального предохранителя.
Другие варианты бисексуального размножения
Некоторые относительно необычные формы размножения:[36][37]
Гиногенез: Сперма стимулирует развитие яйцеклетки без оплодотворения или сингамии. Сперма может попасть в яйцеклетку.
Гибридогенез: Один геном удаляется, чтобы произвести гаплоидные яйца.
Canina мейоз: (иногда называемая «перманентная нечетная полиплоидия») один геном передается по менделевской манере, другие передаются клонально.
Преимущества перекрестного оплодотворения
Обычно считается, что основным преимуществом перекрестного оплодотворения является предотвращение инбридинговая депрессия. Чарльз Дарвин в своей книге 1876 г. Эффекты перекрестного оплодотворения и самооплодотворения в растительном мире (страницы 466-467) резюмировал свои выводы следующим образом.[38]
«В настоящем сборнике показано, что потомки от союза двух разных особей, особенно если их предки находились в очень разных условиях, имеют огромное преимущество в росте, весе, конституциональной силе и плодовитости над самооплодотворяющимися. потомок от одних и тех же родителей. И этого факта вполне достаточно, чтобы объяснить развитие половых элементов, то есть возникновение двух полов ».
Кроме того, некоторые считают,[39] что долгосрочным преимуществом ауткроссинга в природе является повышенная генетическая изменчивость, которая способствует адаптации или предотвращению исчезновения (см. Генетическая изменчивость ).
Смотрите также
- Слияние клеток
- Шапка зачатия
- Устройство зачатия
- Женская сперма
- Развитие плода
- Экстракорпоральное оплодотворение
- Кагуя (мышь)
- Партеногенез, тип воспроизводства, не предполагающий оплодотворения
- Опыление
- Proembryo
- Пронуклеус
- Сверхоплодотворение
- Суперфетация
- Нарушение симметрии и кортикальное вращение
использованная литература
- ^ «Удобрение». Мерриам-Вебстер. Получено 10 июля, 2018.
- ^ http://www.oxfordlearners Commandaries.com/definition/english/impregnation[требуется полная цитата ][постоянная мертвая ссылка ]
- ^ а б Майеншайн Дж. 2017. Первое столетие теории клетки: от структурных единиц до сложных живых систем. В: Штадлер Ф. (ред.), Комплексная история и философия науки. Ежегодник Венского кружка. Институт Венского кружка, Венский университет, Общество Венского кружка, Общество по продвижению концепций научного мира, том 20. Springer, Cham. ссылка на сайт.
- ^ а б Биркхед, Т. И Монтгомери Р. (2009). Три века исследований спермы, стр. 1-42 в: Birkhead, T. R., Hosken, D. J. & Pitnick, S. Биология спермы: эволюционная перспектива. Elsevier / Academic Press, Амстердам. 642 стр., [1].
- ^ Уилкинс А.С., Холлидей Р. (январь 2009 г.). «Эволюция мейоза из митоза». Генетика. 181 (1): 3–12. Дои:10.1534 / генетика.108.099762. ЧВК 2621177. PMID 19139151.
- ^ а б c d Дуань, Цяохун; Кита, Даниэль; Джонсон, Эрик А; Аггарвал, Мини; Гейтс, Лаура; Ву, Хен-Мин; Чунг, Алиса Y (2014). «Активные формы кислорода опосредуют разрыв пыльцевых трубок и высвобождение сперматозоидов для оплодотворения у Arabidopsis». Nature Communications. 5: 3129. Bibcode:2014 НатКо ... 5.3129D. Дои:10.1038 / ncomms4129. PMID 24451849.
- ^ а б c d Cheung, Алиса Y; Ван, Хун; Ву, Хен-Мин (1995). «Тканевый гликопротеин, передающий цветы, привлекает пыльцевые трубки и стимулирует их рост». Ячейка. 82 (3): 383–93. Дои:10.1016/0092-8674(95)90427-1. PMID 7634328.
- ^ Джонстон, Адам (2001). Биология: факты и практика для уровня A. Издательство Оксфордского университета. п.95. ISBN 0-19-914766-3.
- ^ Справочник по растениеводству. Чичестер, Западный Сассекс, Англия: Джон Вили. 2007. с. 466. ISBN 978-0-470-05723-0.
- ^ Кирк, Дэвид; Старр, Сеси (1975). Биология сегодня. Дель Мар, Калифорния: CRM. п.93. ISBN 978-0-394-31093-0.
- ^ Рагхаван, Валаямгхат (2006). Двойное оплодотворение: развитие эмбриона и эндосперма у цветкового растения. Берлин: Springer-Verlag. п.12. ISBN 978-3-540-27791-0.
- ^ Фридман, Уильям Э; Уильямс, Джозеф H (2003). "Модульность женского гаметофита покрытосеменных и ее влияние на раннюю эволюцию эндосперма цветковых растений". Эволюция. 57 (2): 216–30. Дои:10.1111 / j.0014-3820.2003.tb00257.x. PMID 12683519.
- ^ Igic B, Кон-младший (2006). «Распределение систем спаривания растений: систематическая ошибка исследования против обязательных ауткроссинговых видов». Эволюция. 60 (5): 1098–103. Дои:10.1554/05-383.1. PMID 16817548.
- ^ Гудвилли С., Калиш С., Эккерт К. Г. (2005). «Эволюционная загадка смешанных систем спаривания у растений: возникновение, теоретические объяснения и эмпирические данные». Анну. Rev. Ecol. Evol. Syst. 36: 47–79. Дои:10.1146 / annurev.ecolsys.36.091704.175539. S2CID 3755371.
- ^ а б c Райт, С. I; Калиш, S; Слотт, Т (2013). «Эволюционные последствия самоопыления растений». Труды Королевского общества B: биологические науки. 280 (1760): 20130133. Дои:10.1098 / rspb.2013.0133. ЧВК 3652455. PMID 23595268.
- ^ Брандвайн, Янив; Слотте, Таня; Hazzouri, Khaled M; Райт, Стивен I; Куп, Грэм (2013). «Геномная идентификация основополагающих гаплотипов раскрывает историю самоопытывающихся видов Capsella rubella». PLOS Genetics. 9 (9): e1003754. arXiv:1307.4118. Bibcode:2013arXiv1307.4118B. Дои:10.1371 / journal.pgen.1003754. ЧВК 3772084. PMID 24068948.
- ^ Abbott, RJ; Гомес, М.Ф. (1989). «Генетическая структура популяции и скорость ауткроссинга Arabidopsis thaliana (L.) Heynh». Наследственность. 62 (3): 411–418. Дои:10.1038 / хди.1989.56.
- ^ Тан С., Тоомаджан С., Шерман-Бройлс С., Планьол В., Го Ю.Л., Ху Т.Т., Кларк Р.М., Насралла Дж. Б., Вейгель Д., Нордборг М. (2007). «Эволюция самоопыления у Arabidopsis thaliana». Наука. 317 (5841): 1070–2. Bibcode:2007Научный ... 317.1070Т. Дои:10.1126 / science.1143153. PMID 17656687.
- ^ Suarez, S.S .; Пейси, А. А. (2006). «Транспорт спермы в женских половых путях». Обновление репродукции человека. 12 (1): 23–37. Дои:10.1093 / humupd / dmi047. PMID 16272225.
- ^ Чеонг, Инь; МакКлон, Ник; Морган, Хиуел; Мингельс, Роэль; Нг, Ка Ин Бонни (январь 2018 г.). «In vivo динамика кислорода, температуры и pH в женских половых путях и их значение для контрацепции человека: системный обзор». Обновление репродукции человека. 24 (1): 15–34. Дои:10.1093 / humupd / dmx028. PMID 29077897.
- ^ Ли, Шуай; Winuthayanon, Wipawee (2016). «Яйцевод: роль в оплодотворении и раннем развитии эмбриона». Журнал эндокринологии. 232 (1): R1 – R26. Дои:10.1530 / JOE-16-0302. PMID 27875265.
- ^ Бахат, Анат; Айзенбах, Майкл (2006). «Термотаксис спермы». Молекулярная и клеточная эндокринология. 252 (1–2): 115–9. Дои:10.1016 / j.mce.2006.03.027. PMID 16672171.
- ^ Тевес, Мария Э; Гвидобальди, Гектор А; Uñates, Diego R; Санчес, Рауль; Миска, Вернер; Publicover, Стивен Дж; Моралес Гарсия, Адуэн А; Гиохалас, Лаура C (2009). «Молекулярный механизм хемотаксиса спермы человека, опосредованный прогестероном». PLOS ONE. 4 (12): e8211. Bibcode:2009PLoSO ... 4.8211T. Дои:10.1371 / journal.pone.0008211. ЧВК 2782141. PMID 19997608.
- ^ Гнесси Л., Фаббри А., Сильвестрони Л., Моретти С., Фрайоли Ф., Перт CB, Исидори А. (1986). «Доказательства наличия специфических рецепторов для N-формилхемотаксических пептидов на сперматозоидах человека». J Clin Endocrinol Metab. 63 (4): 841–846. Дои:10.1210 / jcem-63-4-841. PMID 3018025.
- ^ Axe, R. L .; Во-первых, Н. Л .; Bushmeyer, S.M .; Clayton, M. K .; Ли, К. Н. (1986-09-01). «Гликозаминогликаны в репродуктивных трактах овец и их влияние на акросомные реакции в сперматозоидах крупного рогатого скота in vitro». Журнал зоотехники. 63 (3): 861–867. Дои:10.2527 / jas1986.633861x. ISSN 0021-8812. PMID 3759713. S2CID 853558.
- ^ Фишман, Эмили Л; Джо, Кён; Nguyen, Quynh P.H; Конг, Донг; Ройфман, Рэйчел; Cekic, Anthony R; Ханал, Сушил; Миллер, Энн Л; Симерли, Кальвин; Шаттен, Джеральд; Лончарек, Ядранка; Меннелла, Вито; Авидор-Рейсс, Томер (2018). «Новая атипичная центриоль сперматозоидов функционирует во время оплодотворения человека». Nature Communications. 9 (1): 2210. Bibcode:2018НатКо ... 9.2210F. Дои:10.1038 / s41467-018-04678-8. ЧВК 5992222. PMID 29880810.
- ^ «Зачатие». Мерриам-Вебстер. Получено 27 января, 2019.
- ^ Moore, K. L .; Т. В. М. Персо (2003). Развивающийся человек: клинически ориентированная эмбриология. Компания W. B. Saunders. ISBN 0-7216-6974-3.
- ^ Дейкстра, Клаас-Доуе Б. (2006). Полевой справочник по стрекозам Британии и Европы. Британское издательство дикой природы. С. 8–9. ISBN 0-9531399-4-8.
- ^ Вальдбауэр, Гилберт (1998). Книга ошибок птицелова. Издательство Гарвардского университета.
- ^ Сельское хозяйство и защита прав потребителей. «Пчеловодство в Африке: жизнь колонии и социальная организация». ФАО.
- ^ Оплодотворение простейших и многоклеточных животных: клеточные и молекулярные аспекты. Хуан Х. Тарин; Антонио Кано (ред.). Springer-Verlag, 2000.
- ^ Репродукция # Автогамия
- ^ «Автогамия | биология».
- ^ Коно, Томохиро; Обата, Яёи; Ву, Куионг; Нива, Кацутоши; Оно, Юкико; Ямамото, Юдзи; Пак, Ын Сон; Со, Чон Сон; Огава, Хидехико (2004). «Рождение партеногенетических мышей, которые могут развиваться до взрослой жизни». Природа. 428 (6985): 860–4. Bibcode:2004Натура.428..860K. Дои:10.1038 / природа02402. PMID 15103378. Сложить резюме – Национальная география (2004-04-21).
- ^ Стенберг, П; Саура, А (2013). «Мейоз и его отклонения у полиплоидных животных». Цитогенетические и геномные исследования. 140 (2–4): 185–203. Дои:10.1159/000351731. PMID 23796636.
- ^ Шток, М; Устинова, Дж; Бетто-Коллиард, К; Schartl, M; Moritz, C; Перрин, Н. (2011). «Одновременная передача менделевского и клонального генома у репродуктивного триплоидного позвоночного животного». Труды Королевского общества B: биологические науки. 279 (1732): 1293–1299. Дои:10.1098 / rspb.2011.1738. ЧВК 3282369. PMID 21993502.
- ^ Дарвин CR (1876 г.). Эффекты перекрестного и самооплодотворения в растительном мире. Лондон: Джон Мюррей. http://darwin-online.org.uk/converted/published/1881-Worms-CrossandSelfFertilisation-F1249/1876-F1249.html[постоянная мертвая ссылка ] см. стр. 466-467
- ^ Отто, С.П .; Герштейн, A.C (2006). «Зачем заниматься сексом? Популяционная генетика пола и рекомбинации». Сделки Биохимического Общества. 34 (4): 519–22. Дои:10.1042 / BST0340519. PMID 16856849. S2CID 14542950.
внешние ссылки
- Оплодотворение (зачатие) видео
| Режимы |
|  |
|---|---|---|
| Удобрение | ||
| Родительская забота | ||
| похожие темы | ||
| Биологические термины | |
|---|---|
| Сексуальный воспроизведение | |
| Сексуальность | |
| |
| Авторитетный контроль |
|---|