Зеленый флуоресцентный белок - Green fluorescent protein
| Зеленый флуоресцентный белок | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Структура Aequorea victoria зеленый флуоресцентный белок.[1] | |||||||||
| Идентификаторы | |||||||||
| Символ | GFP | ||||||||
| Pfam | PF01353 | ||||||||
| Pfam клан | CL0069 | ||||||||
| ИнтерПро | IPR011584 | ||||||||
| CATH | 1ema | ||||||||
| SCOP2 | 1ema / Объем / СУПФАМ | ||||||||
| |||||||||
| Зеленый флуоресцентный белок | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Организм | |||||||
| Символ | GFP | ||||||
| UniProt | P42212 | ||||||
| |||||||
В зеленый флуоресцентный белок (GFP) это белок состоит из 238 аминокислота остатки (26,9 кДа ) ярко-зеленый флуоресценция при воздействии синего света на ультрафиолетовый ассортимент.[2][3] Подобные белки, которые также флуоресцируют зеленым цветом, встречаются во многих морских организмах, но этикетка GFP традиционно относится к этому конкретному белку, который был впервые выделен из медуза Aequorea victoria и иногда его называют - когда требуется такая точность -avGFP.
GFP от А. Виктория имеет крупный пик возбуждения в длина волны 395 нм и второстепенный на 475 нм. Его пик излучения находится при 509 нм, что находится в нижней зеленой части видимый спектр. Флуоресценция квантовый выход (QY) GFP составляет 0,79. GFP от морские анютины глазки (Renilla reniformis ) имеет один главный пик возбуждения при 498 нм. GFP является отличным инструментом во многих формах биологии благодаря своей способности формировать внутренние хромофор без каких-либо аксессуаров кофакторы, генные продукты или ферменты / субстраты кроме молекулярного кислорода.[4]
В ячейка и молекулярная биология, GFP ген часто используется как репортер выражения.[5] Он был использован в модифицированных формах, чтобы сделать биосенсоры, и было создано много животных, которые экспрессируют GFP, что демонстрирует доказательство концепции что ген может экспрессироваться в данном организме, в выбранных органах или в представляющих интерес клетках. GFP может быть введен животным или другим видам через трансгенные методы, и сохраняются в их геноме и в геноме их потомства. На сегодняшний день GFP экспрессируется у многих видов, включая бактерии, дрожжи, грибы, рыбу и млекопитающих, в том числе в клетках человека. Ученые Роджер Ю. Цзянь, Осаму Шимомура, и Мартин Чалфи были награждены 2008 Нобелевская премия по химии 10 октября 2008 г. за открытие и разработку зеленого флуоресцентного белка.
Задний план


GFP дикого типа (wtGFP)
В 1960-х и 1970-х годах GFP вместе с отдельным люминесцентным белком экуорин (ан фермент что катализирует распад люциферин, испуская свет), впервые был очищен от медуз Aequorea victoria и его свойства, изученные Осаму Шимомура.[6] В А. Виктория, Флуоресценция GFP возникает, когда экворин взаимодействует с Ca2+ ионы, вызывающие голубое свечение. Часть этой люминесцентной энергии передается GFP, смещая общий цвет в сторону зеленого.[7] Однако его полезность в качестве инструмента для молекулярных биологов осознавалась только в 1992 году, когда Дуглас Прашер сообщили о клонировании и нуклеотидной последовательности wtGFP в Ген.[8] Финансирование этого проекта закончилось, поэтому Прашер прислал кДНК образцы в несколько лабораторий. Лаборатория Мартин Чалфи экспрессировали кодирующую последовательность wtGFP с удаленными первыми несколькими аминокислотами в гетерологичных клетках Кишечная палочка и C. elegans, опубликовав результаты в Наука в 1994 г.[9] Лаборатория Фредерика Цуджи независимо сообщила об экспрессии рекомбинантного белка месяц спустя.[10] Примечательно, что молекула GFP сворачивается и флуоресцирует при комнатной температуре без необходимости использования экзогенных кофакторов, специфичных для медуз. Хотя этот близкий к весу GFP был флуоресцентным, он имел несколько недостатков, включая спектры двойного пика возбуждения, чувствительность к pH, чувствительность к хлоридам, низкий квантовый выход флуоресценции, плохую фотостабильность и плохую укладку при 37 ° C.
Первой описанной кристаллической структурой GFP была структура мутанта S65T группой Ремингтона в г. Наука в 1996 г.[11] Через месяц группа Phillips независимо сообщила о структуре GFP дикого типа в Природа Биотехнологии.[12] Эти кристаллические структуры обеспечили жизненно важную основу для хромофор образование и взаимодействия соседних остатков. Исследователи модифицировали эти остатки с помощью направленного и случайного мутагенеза, чтобы получить широкий спектр производных GFP, используемых сегодня. Дальнейшие исследования GFP показали, что он устойчив к детергентам, протеазам, обработке хлоридом гуанидиния (GdmCl) и резким перепадам температуры.[13]
Производные GFP

Из-за возможности широкого использования и растущих потребностей исследователей было создано множество различных мутантов GFP.[14][15] Первым значительным улучшением была единичная точечная мутация (S65T), о которой сообщалось в 1995 г. Природа от Роджер Цзянь.[16] Эта мутация резко улучшила спектральные характеристики GFP, что привело к увеличению флуоресценции, фотостабильности и сдвигу основного пика возбуждения до 488 нм, при этом пик излучения сохранялся на уровне 509 нм. Это соответствовало спектральным характеристикам общедоступных FITC наборы фильтров, повышающие практичность их использования обычным исследователем. Точечный мутант с эффективностью сворачивания при 37 ° C (F64L) по этому каркасу, дающий улучшенный GFP (EGFP), была открыта в 1995 году лабораториями Thastrup[17] и Фалькоу.[18] EGFP позволил практическое использование GFP в клетках млекопитающих. EGFP имеет коэффициент экстинкции (обозначается ε) 55000 M−1см−1.[19] Флуоресценция квантовый выход (QY) EGFP составляет 0,60. Относительная яркость, выраженная как ε • QY, составляет 33 000 M−1см−1.
Суперпапка GFP (sfGFP), серия мутаций, позволяющих GFP быстро сворачиваться и созревать даже при слиянии с плохо сворачивающимися пептидами, была описана в 2006 году.[20]
Было сделано много других мутаций, включая цветовые мутанты; особенно, синий флуоресцентный белок (EBFP, EBFP2, Азурит, mKalama1), голубой флуоресцентный белок (ECFP, Cerulean, CyPet, mTurquoise2) и желтый флуоресцентный белок производные (YFP, Citrine, Venus, YPet). Производные BFP (кроме mKalama1) содержат замену Y66H. Они демонстрируют широкую полосу поглощения в ультрафиолетовом диапазоне с центром около 380 нм и максимум излучения при 448 нм. Мутант зеленого флуоресцентного белка (BFPms1) это преимущественно связывает Zn (II) и Cu (II) была разработана. BFPms1 имеет несколько важных мутаций, включая хромофор BFP (Y66H), Y145F для более высокого квантового выхода, H148G для создания дыры в бета-стволе и несколько других мутаций, которые увеличивают растворимость. Связывание Zn (II) увеличивает интенсивность флуоресценции, в то время как связывание Cu (II) гасит флуоресценцию и смещает максимум поглощения с 379 до 444 нм. Следовательно, они могут использоваться как биосенсоры цинка.[21]

Связывание хромофора. Критической мутацией в производных циана является замена Y66W, которая приводит к образованию хромофора с индол а не фенольный компонент. Несколько дополнительных компенсаторных мутаций в окружающем стволе необходимы для восстановления яркости этого модифицированного хромофора из-за увеличенного количества индольной группы. В ECFP и Cerulean N-концевая половина седьмой цепи демонстрирует две конформации. Обе эти конформации обладают сложным набором ван-дер-ваальсовых взаимодействий с хромофором. Мутации Y145A и H148D в Cerulean стабилизируют эти взаимодействия и позволяют хромофору быть более плоским, лучше упакованным и менее склонным к столкновительному тушению.[22]
Дополнительный случайный выбор, ориентированный на сайт мутагенез в сочетании со скринингом на основе времени жизни флуоресценции дополнительно стабилизировал седьмую β-цепь, что привело к яркому варианту, mTurquoise2, с квантовым выходом (QY) 0,93.[23] Смещенная в красную область длина волны производных YFP достигается за счет мутации T203Y и возникает из-за взаимодействий π-стэкинга между замещенным остатком тирозина и хромофором.[3] Эти два класса спектральных вариантов часто используются для Фёрстеровский резонансный перенос энергии (FRET) эксперименты. Генетически кодируемые репортеры FRET, чувствительные к клеточным сигнальным молекулам, таким как кальций или глутамат, состояние фосфорилирования белка, комплементация белка, димеризация рецептора и другие процессы, обеспечивают высокоспецифичные оптические считывания клеточной активности в реальном времени.
Полурациональный мутагенез ряда остатков привел к появлению чувствительных к pH мутантов, известных как pHluorins, а позднее - к суперэклиптическим pHluorins. Используя быстрое изменение pH при слиянии синаптических пузырьков, pHluorins помечены синаптобревин были использованы для визуализации синаптической активности в нейронах.[24]
Редокс-чувствительный GFP (roGFP ) был разработан путем введения цистеинов в структуру бета-ствола. В редокс состояние цистеинов определяет флуоресцентный свойства roGFP.[25]
Номенклатура
Номенклатура модифицированных GFP часто сбивает с толку из-за перекрытия отображения нескольких версий GFP на одно имя. Например, mGFP часто относится к GFP с N-выводом пальмитоилирование что заставляет GFP связываться с клеточные мембраны. Однако этот же термин также используется для обозначения мономерный GFP, который часто достигается с помощью димерного интерфейса, нарушающего мутацию A206K.[26] GFP дикого типа имеет слабую димеризация тенденция при концентрациях выше 5 мг / мл. mGFP также означает «модифицированный GFP», который был оптимизирован за счет замены аминокислот для стабильной экспрессии в растительных клетках.
В природе
Цель обоих (основных) биолюминесценция (от экуорин действие на люциферин) и (вторичный) флуоресценция GFP у медуз неизвестно. GFP совместно экспрессируется с эккорином в небольших гранулах по краю колокольчика медузы. Пик вторичного возбуждения (480 нм) GFP действительно поглощает часть синего излучения экворина, придавая биолюминесценции более зеленый оттенок. Остаток серина 65 GFP хромофор отвечает за спектры возбуждения с двумя пиками GFP дикого типа. Он консервативен во всех трех изоформах GFP, первоначально клонированных Prasher. Почти все мутации этого остатка объединяют спектры возбуждения в один пик при 395 нм или 480 нм. Точный механизм этой чувствительности сложен, но, по-видимому, включает передачу водорода от серина 65 к глутамату 222, который влияет на ионизацию хромофора.[3] Поскольку единичная мутация может значительно усилить пик возбуждения 480 нм, что делает GFP гораздо более эффективным партнером эккорина, А. Виктория похоже, эволюционно предпочитает менее эффективный спектр возбуждения с двумя пиками. Роджер Цзян предположил, что изменение гидростатического давления с глубиной может повлиять на способность серина 65 отдавать водород хромофору и изменить соотношение двух пиков возбуждения. Таким образом, медуза может менять цвет своей биолюминесценции с глубиной. Однако резкое сокращение популяции медуз в Friday Harbour, где GFP был первоначально открыт, препятствует дальнейшему изучению роли GFP в естественной среде обитания медуз.
Другие флуоресцентные белки

Существует множество GFP-подобных белков, которые, несмотря на то, что принадлежат к тому же семейству белков, что и GFP, не происходят напрямую из Aequorea victoria. Они включают dsRed, eqFP611, Dronpa, TagRFP, KFP, EosFP / IrisFP, Dendra и т. д. Эти белки, полученные из белков различных организмов, иногда могут проявлять неантипированный подход к образованию хромофоров. Некоторые из них, такие как KFP, разработаны из естественно не флуоресцентных или слабо флуоресцентных белков, которые могут быть значительно улучшены с помощью мутагенеза.[27] Когда используются GFP-подобные цилиндры с различными спектральными характеристиками, спектры возбуждения одного хромофора могут использоваться для питания другого хромофора (FRET), обеспечивая преобразование между длинами волн света.[28]
FMN-связывающие флуоресцентные белки (FbFP) были разработаны в 2007 году и представляют собой класс небольших (11-16 кДа) кислороднезависимых флуоресцентных белков, которые происходят от рецепторов синего света. Они предназначены, в частности, для использования в анаэробных или гипоксических условиях, поскольку для образования и связывания хромофора Flavin не требуется молекулярный кислород, как в случае синтеза хромофора GFP.[29]

Флуоресцентные белки с другими хромофорами, такие как UnaG с билирубином, могут проявлять уникальные свойства, такие как излучение со смещением в красную область при длине волны более 600 нм или фотопреобразование из состояния излучения зеленого в состояние излучения красного цвета. У них могут быть длины волн возбуждения и излучения, достаточно далеко разнесенные, чтобы обеспечить преобразование между красным и зеленым светом.
Новый класс флуоресцентный белок возникла из цианобактериальный (Trichodesmium erythraeum ) фикобилипротеин, α-аллофикоцианин, и назвал небольшой ультра красный флуоресцентный белок (smURFP ) в 2016 году. smURFP автокаталитически само включает хромофор биливердин без необходимости внешнего белок, известный как лиазе.[30] Медуза - и коралл -производные GFP-подобные белки требуют кислород и произвести стехиометрический количество пероксид водорода на хромофор формирование.[31] smURFP не требует кислород или произвести пероксид водорода и использует хромофор, биливердин. smURFP имеет большой коэффициент экстинкции (180 000 млн−1 см−1) и имеет скромную квантовый выход (0,20), что делает его биофизическую яркость сопоставимой с eGFP и ~ в 2 раза ярче, чем большинство красных или дальних красных флуоресцентные белки полученный из коралл. smURFP спектральные свойства аналогичны органическому красителю Cy5.[30]

Обзоры новых классов флуоресцентных белков и их применения можно найти в цитируемых обзорах.[32][33]
Структура

GFP имеет бета-баррель структура, состоящая из одиннадцати β-нитей с гофрированным расположением листов, с альфа-спиралью, содержащей ковалентно связанные хромофор 4-(п-гидроксибензилиден) имидазолидин-5-он (HBI), проходящий через центр.[3][11][12] Пять более коротких альфа-спиралей образуют шапочки на концах структуры. В бета-баррель структура представляет собой почти идеальный цилиндр длиной 42 Å и диаметром 24 Å (в некоторых исследованиях сообщалось о диаметре 30 Å.[13]),[11] создание того, что называется образованием «β-can», которое является уникальным для семейства GFP.[12] HBI, спонтанно модифицированная форма трипептида Ser65-Tyr66-Gly67, не флуоресцирует в отсутствие правильно свернутого каркаса GFP и существует в основном в неионизированной фенольной форме в wtGFP.[34] Обращенные внутрь боковые цепи цилиндра вызывают специфические реакции циклизации в Ser65-Tyr66-Gly67, которые вызывают ионизацию HBI до фенолятной формы и хромофор формирование. Этот процесс посттрансляционная модификация упоминается как созревание.[35] Сеть водородных связей и взаимодействия электронов с этими боковыми цепями влияют на цвет, интенсивность и фотостабильность GFP и его многочисленных производных.[36] Плотно упакованный характер ствола исключает молекулы растворителя, защищая хромофор флуоресценция от тушения водой. В дополнение к автоциклизации Ser65-Tyr66-Gly67, по остатку Tyr66 происходит реакция 1,2-дегидрирования.[13] Помимо трех остатков, образующих хромофор, все остатки, такие как Gln94, Arg96, His148, Thr203 и Glu222, действуют как стабилизаторы. Остатки Gln94, Arg96 и His148 способны стабилизироваться за счет делокализации заряда хромофора. Arg96 является наиболее важным стабилизирующим остатком из-за того, что он вызывает необходимые структурные перестройки, которые необходимы в кольце HBI. Любая мутация остатка Arg96 приведет к снижению скорости развития хромофора, потому что правильные электростатические и стерические взаимодействия будут потеряны. Tyr66 является реципиентом водородных связей и не ионизируется для создания благоприятной электростатики.[37]

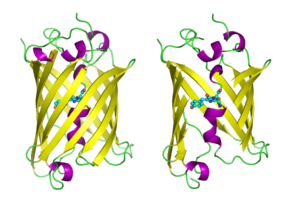 Молекулы GFP, нарисованные в мультяшном стиле, одна целиком и одна сбоку бета-баррель вырезать, чтобы раскрыть хромофор (выделено как клюшка ). От PDB: 1GFL. |

Приложения
Репортерные анализы
Зеленый флуоресцентный белок можно использовать в качестве репортерный ген.[38][39]
Например, GFP можно использовать в качестве репортера уровней токсичности для окружающей среды. Было показано, что этот белок является эффективным способом измерения уровней токсичности различных химических веществ, включая этанол, п-формальдегид, фенол, триклозан и парабен. GFP великолепен как репортерный белок, потому что он не влияет на хозяина при попадании в клеточную среду хозяина. Благодаря этой способности не требуются красители для внешней визуализации, АТФ или кофакторы. Что касается уровней загрязняющих веществ, флуоресценция измерялась, чтобы оценить влияние, которое загрязняющие вещества оказывают на клетку-хозяин. Также измеряли клеточную плотность клетки-хозяина. Результаты исследования, проведенного Сонг, Ким и Сео (2016), показали, что по мере увеличения уровня загрязняющих веществ наблюдается снижение как флуоресценции, так и плотности клеток. Это свидетельствовало о том, что активность клеток снизилась. Дополнительные исследования этого конкретного приложения, чтобы определить механизм, с помощью которого GFP действует как маркер загрязнителя.[40] Подобные результаты наблюдались у рыбок данио, потому что рыбки данио, которым вводили GFP, были примерно в двадцать раз более восприимчивыми к распознаванию клеточных стрессов, чем рыбки данио, которым не вводили GFP.[41]
Преимущества
Самым большим преимуществом GFP является то, что он может передаваться по наследству, в зависимости от того, как он был введен, что позволяет продолжать изучение клеток и тканей, в которых он экспрессируется. Визуализация GFP неинвазивна, требует только освещения синим светом. Сам по себе GFP не влияет на биологические процессы, но при слиянии с представляющими интерес белками требуется тщательный дизайн линкеров для поддержания функции интересующего белка. Более того, при использовании с мономером он может легко диффундировать по клеткам.[42]
Флуоресцентная микроскопия
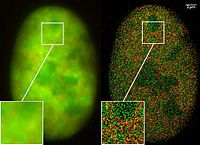
Доступность GFP и его производных полностью изменила определение флуоресцентная микроскопия и способ его использования в клеточной биологии и других биологических дисциплинах.[43] В то время как большинство небольших флуоресцентных молекул, таких как FITC (флюоресцеина изотиоцианат) сильно фототоксичный при использовании в живых клетках флуоресцентные белки, такие как GFP, обычно гораздо менее вредны при освещении в живых клетках. Это послужило толчком к разработке высокоавтоматизированных систем флуоресцентной микроскопии живых клеток, которые можно использовать для наблюдения за клетками с течением времени, экспрессирующими один или несколько белков, меченных флуоресцентными белками. Например, GFP широко использовался для маркировки сперматозоиды различных организмов для целей идентификации, как в Drosophila melanogaster, где выражение GFP может использоваться как маркер для определенной характеристики. GFP также может экспрессироваться в различных структурах, позволяющих различать морфологию. В таких случаях ген для производства GFP включается в геном организма в области ДНК, которая кодирует целевые белки и которая контролируется тем же регуляторная последовательность; то есть регуляторная последовательность гена теперь контролирует продукцию GFP в дополнение к меченному белку (белкам). В клетках, где экспрессируется ген и продуцируются меченые белки, GFP продуцируется одновременно. Таким образом, только те клетки, в которых экспрессируется меченый ген или продуцируются целевые белки, будут флуоресцировать при наблюдении под флуоресцентной микроскопией. Анализ таких интервальных видеороликов переопределил понимание многих биологических процессов, включая сворачивание белка, транспорт белка и динамику РНК, которые в прошлом изучались с использованием фиксированного (т. Е. Мертвого) материала. Полученные данные также используются для калибровки математических моделей внутриклеточных систем и оценки скорости экспрессии генов.[44] Точно так же GFP можно использовать в качестве индикатора экспрессии белка в гетерологичных системах. В этом сценарии слитые белки, содержащие GFP, вводятся косвенно, с использованием РНК конструкции, или напрямую, с самим меченным белком. Этот метод полезен для изучения структурных и функциональных характеристик меченого белка в макромолекулярном или одномолекулярном масштабе с помощью флуоресцентной микроскопии.
В Вертико СМИ Микроскоп, использующий технологию SPDM Phymod, использует так называемый эффект «обратимого фотообесцвечивания» флуоресцентных красителей, таких как GFP и его производные, для локализации их в виде отдельных молекул с оптическим разрешением 10 нм. Это также может быть выполнено как совместная локализация двух производных GFP (2CLM).[45]
Еще одно мощное применение GFP - экспрессия белка в небольших наборах определенных клеток. Это позволяет исследователям оптически обнаруживать определенные типы клеток. in vitro (в блюде) или даже in vivo (в живом организме).[46] Генетическое объединение нескольких спектральных вариантов GFP - полезный трюк для анализа схем мозга (Brainbow ).[47] Другие интересные применения флуоресцентных белков в литературе включают использование FP в качестве сенсоров нейрон мембранный потенциал,[48] отслеживание AMPA рецепторы на клеточных мембранах,[49] вирусное проникновение и заражение отдельных грипп вирусы и лентивирусные вирусы,[50][51] и т.п.
Также было обнаружено, что новые линии трансгенных крыс GFP могут иметь значение для генной терапии, а также регенеративной медицины.[52] При использовании «высокоэкспрессирующего» GFP трансгенные крысы демонстрируют высокую экспрессию в большинстве тканей и во многих клетках, которые не были охарактеризованы или были плохо охарактеризованы у ранее трансгенных крыс с GFP.
Показано, что GFP полезен в криобиология как анализ жизнеспособности. Корреляция жизнеспособности, измеренная трипановый синий анализы были 0,97.[53] Другое применение - использование котрансфекции GFP в качестве внутреннего контроля эффективности трансфекции в клетках млекопитающих.[54]
Новое возможное использование GFP включает использование его в качестве чувствительного монитора внутриклеточных процессов с помощью лазерной системы eGFP, созданной из линии клеток эмбриональной почки человека. Первый инженерный живой лазер состоит из клетки, экспрессирующей eGFP внутри отражающей оптической полости и воздействующей на нее импульсами синего света. При определенном пороге импульса оптический выход eGFP становится более ярким и полностью однородным по цвету чистого зеленого цвета с длиной волны 516 нм. Прежде чем испускаться в виде лазерного излучения, свет отражается взад и вперед в полости резонатора и многократно проходит через ячейку. Изучая изменения оптической активности, исследователи могут лучше понять клеточные процессы.[55][56]
GFP широко используется в исследованиях рака для маркировки и отслеживания раковых клеток. Раковые клетки, меченные GFP, использовались для моделирования метастазирования - процесса распространения раковых клеток в отдаленные органы.[57]
Разделить GFP
GFP можно использовать для анализа совместной локализации белков. Это достигается «расщеплением» белка на два фрагмента, способных к самосборке, а затем слиянием каждого из них с двумя интересующими белками. По отдельности эти неполные фрагменты GFP не могут флуоресцировать. Однако, если два интересующих белка колокализуются, то два фрагмента GFP собираются вместе с образованием GFP-подобной структуры, способной флуоресцировать. Следовательно, измеряя уровень флуоресценции, можно определить, колокализуются ли два интересующих белка.[58]
Макро-фотография
Макромасштабные биологические процессы, такие как распространение вирусных инфекций, можно отслеживать с помощью маркировки GFP.[59] В прошлом мутагенный ультрафиолетовый свет (УФ) использовался для освещения живых организмов (например, см.[60]), чтобы обнаружить и сфотографировать выражение GFP. В последнее время появилась техника, использующая немутагенные светодиодные лампы.[61] были разработаны для макросъемки.[62] В технике используется насадка для эпифлуоресцентной камеры.[63] основанный на том же принципе, что и при построении эпифлуоресцентные микроскопы.
Трансгенные животные

Альба зеленый флуоресцентный кролик, был создан французской лабораторией по заказу Эдуардо Кац использование GFP для художественных и социальных комментариев.[64] Американская компания Yorktown Technologies поставляет в аквариумные магазины зеленые флуоресцентные лампы. данио (GloFish ), которые изначально были разработаны для обнаружения загрязнения водных путей. Американская компания NeonPets продала зеленых флуоресцентных мышей индустрии домашних животных под названием NeonMice.[65] Зеленые флуоресцентные свиньи, известные как Ноэлс, были выведены группой исследователей под руководством Ву Шинн-Чи из Департамента зоотехники и технологий в Национальный Тайваньский университет.[66] Японско-американская команда создала зеленый флуоресцентный кошки как доказательство концепции их потенциального использования в качестве модельных организмов для болезней, особенно ВИЧ.[67] В 2009 году южнокорейская команда из Сеульского национального университета вывели первых трансгенных животных. гончие с клетками фибробластов морских анемонов. Собаки излучают красный флуоресцентный свет, и они предназначены для того, чтобы ученые могли изучать гены, вызывающие такие заболевания человека, как нарколепсия и слепота.[68]
Искусство
Джулиан Восс-Андреэ, художник немецкого происхождения, специализирующийся на «белковых скульптурах»,[69] создал скульптуры на основе структуры GFP, в том числе "Зеленый флуоресцентный белок" высотой 1,70 м (5 футов 6 дюймов) (2004 г.)[70] и "Стальная медуза" высотой 1,40 м (4'7 ") (2006 г.). Последняя скульптура находится на месте открытия GFP Шимомура в 1962 г. Вашингтонский университет с Friday Harbour Laboratories.[71]

Смотрите также
использованная литература
- ^ Ормо М., Кубитт А.Б., Каллио К., Гросс Л.А., Цзянь Р.Ю., Ремингтон С.Дж. (сентябрь 1996 г.). «Кристаллическая структура зеленого флуоресцентного белка Aequorea victoria». Наука. 273 (5280): 1392–5. Bibcode:1996 Наука ... 273.1392O. Дои:10.1126 / science.273.5280.1392. PMID 8703075. S2CID 43030290.
- ^ Прендергаст Ф.Г., Манн К.Г. (август 1978 г.).«Химические и физические свойства экворина и зеленого флуоресцентного белка, выделенного из Aequorea forskålea». Биохимия. 17 (17): 3448–53. Дои:10.1021 / bi00610a004. PMID 28749.
- ^ а б c d Цзянь Р.Ю. (1998). «Зеленый флуоресцентный белок» (PDF). Ежегодный обзор биохимии. 67: 509–44. Дои:10.1146 / annurev.biochem.67.1.509. PMID 9759496.
- ^ Степаненко О.В., Верхуша В.В., Кузнецова И.М., Уверский В.Н., Туроверов К.К. (август 2008 г.). «Флуоресцентные белки как биомаркеры и биосенсоры: бросая цветные огни на молекулярные и клеточные процессы». Современная наука о белках и пептидах. 9 (4): 338–69. Дои:10.2174/138920308785132668. ЧВК 2904242. PMID 18691124.
- ^ Филлипс GJ (октябрь 2001 г.). «Зеленый флуоресцентный белок - яркая идея для изучения локализации бактериального белка». Письма о микробиологии FEMS. 204 (1): 9–18. Дои:10.1016 / S0378-1097 (01) 00358-5. PMID 11682170.
- ^ Шимомура О., Джонсон Ф. Х., Сайга Й. (июнь 1962 г.). «Извлечение, очистка и свойства экворина, биолюминесцентного белка из светящегося гидромедузана, Aequorea». Журнал клеточной и сравнительной физиологии. 59 (3): 223–39. Дои:10.1002 / jcp.1030590302. PMID 13911999.
- ^ Мориз Х., Шимомура О., Джонсон Ф.Х., Винант Дж. (Июнь 1974 г.). «Межмолекулярный перенос энергии в биолюминесцентной системе Aequorea». Биохимия. 13 (12): 2656–62. Дои:10.1021 / bi00709a028. PMID 4151620.
- ^ Prasher DC, Eckenrode VK, Ward WW, Prendergast FG, Cormier MJ (февраль 1992 г.). «Первичная структура зеленого флуоресцентного белка Aequorea victoria». Ген. 111 (2): 229–33. Дои:10.1016 / 0378-1119 (92) 90691-Н. PMID 1347277.
- ^ Chalfie M, Tu Y, Euskirchen G, Ward WW, Prasher DC (февраль 1994 г.). «Зеленый флуоресцентный белок как маркер экспрессии генов». Наука. 263 (5148): 802–5. Bibcode:1994Sci ... 263..802C. Дои:10.1126 / наука.8303295. PMID 8303295. S2CID 9043327.
- ^ Inouye S, Tsuji FI (март 1994 г.). «Зеленый флуоресцентный белок Aequorea. Экспрессия гена и характеристики флуоресценции рекомбинантного белка». Письма FEBS. 341 (2–3): 277–80. Дои:10.1016/0014-5793(94)80472-9. PMID 8137953.
- ^ а б c Ормо М., Кубитт А.Б., Каллио К., Гросс Л.А., Цзянь Р.Ю., Ремингтон С.Дж. (сентябрь 1996 г.). «Кристаллическая структура зеленого флуоресцентного белка Aequorea victoria». Наука. 273 (5280): 1392–5. Bibcode:1996 Наука ... 273.1392O. Дои:10.1126 / science.273.5280.1392. PMID 8703075. S2CID 43030290.
- ^ а б c Ян Ф., Мосс Л.Г., Филлипс Г.Н. (октябрь 1996 г.). «Молекулярная структура зеленого флуоресцентного белка» (PDF). Природа Биотехнологии. 14 (10): 1246–51. Дои:10.1038 / nbt1096-1246. HDL:1911/19233. PMID 9631087. S2CID 34713931.
- ^ а б c Brejc, K .; Sixma, T. K .; Kitts, P.A .; Kain, S. R .; Tsien, R. Y .; Ормё, М .; Ремингтон, С. Дж. Структурные основы двойного возбуждения и фотоизомеризации Aequorea victoria зеленый флуоресцентный белок. Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ.. 1997, 94 (6), 2306-2311.
- ^ Shaner NC, Steinbach PA, Tsien RY (декабрь 2005 г.). «Руководство по выбору флуоресцентных белков» (PDF). Методы природы. 2 (12): 905–9. Дои:10.1038 / nmeth819. PMID 16299475. S2CID 10024284.
- ^ Вильгельмссон М, Тор Y (2016). Флуоресцентные аналоги биомолекулярных строительных блоков: конструкция и применение. Нью-Джерси: Уайли. ISBN 978-1-118-17586-6.
- ^ Heim R, Cubitt AB, Tsien RY (февраль 1995 г.). «Улучшенная зеленая флуоресценция» (PDF). Природа. 373 (6516): 663–4. Bibcode:1995Натура.373..663H. Дои:10.1038 / 373663b0. PMID 7854443. S2CID 40179694.
- ^ Патент США 6172188, Thastrup O, Tullin S, Kongsbak Poulsen L, Bjørn S, "Fluorescent Proteins", опубликовано 9 января 2001 г.
- ^ Кормак Б.П., Вальдивия Р.Х., Фалькоу С. (1996). «Оптимизированные для FACS мутанты зеленого флуоресцентного белка (GFP)». Ген. 173 (1 Спецификация): 33–38. Дои:10.1016/0378-1119(95)00685-0. PMID 8707053.
- ^ McRae SR, Brown CL, Bushell GR (май 2005 г.). «Быстрая очистка EGFP, EYFP и ECFP с высоким выходом и чистотой». Экспрессия и очистка белков. 41 (1): 121–127. Дои:10.1016 / j.pep.2004.12.030. PMID 15802229.
- ^ Педелак Дж. Д., Кабантус С., Тран Т., Тервиллигер Т.С., Уолдо Г.С. (январь 2006 г.). «Разработка и характеристика зеленого флуоресцентного белка superfolder». Природа Биотехнологии. 24 (1): 79–88. Дои:10.1038 / nbt1172. PMID 16369541. S2CID 2966399.
- ^ Барондо Д.П., Кассманн С.Дж., Тайнер Дж.А., Getzoff ED (Апрель 2002 г.). «Структурная химия зеленого флуоресцентного белкового биосенсора Zn». Журнал Американского химического общества. 124 (14): 3522–3524. Дои:10.1021 / ja0176954. PMID 11929238.
- ^ Lelimousin M, Noirclerc-Savoye M, Lazareno-Saez C, Paetzold B, Le Vot S, Chazal R, Macheboeuf P, Field MJ, Bourgeois D, Royant A (октябрь 2009 г.). «Внутренняя динамика в квантовом выходе флуоресценции ECFP и Cerulean». Биохимия. 48 (42): 10038–10046. Дои:10.1021 / bi901093w. PMID 19754158.
- ^ Goedhart J, von Stetten D, Noirclerc-Savoye M, Lelimousin M, Joosen L, Hink MA, van Weeren L, Gadella TW, Royant A (2012). «Управляемая структурой эволюция голубых флуоресцентных белков в сторону квантового выхода 93%». Nature Communications. 3: 751. Bibcode:2012NatCo ... 3..751G. Дои:10.1038 / ncomms1738. ЧВК 3316892. PMID 22434194.
- ^ Miesenböck G, De Angelis DA, Rothman JE (июль 1998 г.). «Визуализация секреции и синаптической передачи с помощью pH-чувствительных зеленых флуоресцентных белков». Природа. 394 (6689): 192–5. Bibcode:1998Натура.394..192М. Дои:10.1038/28190. PMID 9671304. S2CID 4320849.
- ^ Hanson GT, Aggeler R, Oglesbee D, Cannon M, Capaldi RA, Tsien RY, Remington SJ (март 2004 г.). «Исследование окислительно-восстановительного потенциала митохондрий с помощью индикаторов окислительно-восстановительного потенциала зеленых флуоресцентных белков». Журнал биологической химии. 279 (13): 13044–53. Дои:10.1074 / jbc.M312846200. PMID 14722062.
- ^ Захариас Д.А., Скрипка Д.Д., Ньютон А.С., Цзянь Р.Й. (май 2002 г.). «Разделение липид-модифицированных мономерных GFP на мембранные микродомены живых клеток». Наука. 296 (5569): 913–16. Bibcode:2002Наука ... 296..913Z. Дои:10.1126 / science.1068539. PMID 11988576. S2CID 14957077.
- ^ Чудаков Д.М., Белоусов В.В., Зарайский А.Г., Новоселов В.В., Староверов Д.Б., Зоров Д.Б., Лукьянов С., Лукьянов К.А. (февраль 2003 г.). «Kindling флуоресцентные белки для точного фотомечения in vivo». Природа Биотехнологии. 21 (2): 191–4. Дои:10.1038 / nbt778. PMID 12524551. S2CID 52887792.
- ^ Wiens MD, Shen Y, Li X, Salem MA, Smisdom N, Zhang W., Brown A, Campbell RE (декабрь 2016 г.). «Тандемный зелено-красный гетеродимерный флуоресцентный белок с высокой эффективностью FRET». ChemBioChem. 17 (24): 2361–2367. Дои:10.1002 / cbic.201600492. PMID 27781394.
- ^ Дреппер, Т., Эггерт, Т., Цирколоне, Ф., Хек, А., Краусс, У., Гутерл, Дж. К., Вендорф, М., Лози, А., Гертнер, В., Йегер, К. Э. (2007) . «Репортерные белки для флуоресценции in vivo без кислорода». Nat Biotechnol. 25 (4): 443–445. Дои:10.1038 / nbt1293. PMID 17351616. S2CID 7335755.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ а б Родригес Е.А., Тран Г.Н., Гросс Л.А., Крисп Дж.Л., Шу Х, Лин Дж.Й., Цзянь Р.Й. (сентябрь 2016 г.). «Дальний красный флуоресцентный белок произошел из цианобактериального фикобилипротеина». Методы природы. 13 (9): 763–9. Дои:10.1038 / nmeth.3935. ЧВК 5007177. PMID 27479328.
- ^ Цзянь Р.Ю. (01.01.1998). «Зеленый флуоресцентный белок». Ежегодный обзор биохимии. 67 (1): 509–44. Дои:10.1146 / annurev.biochem.67.1.509. PMID 9759496. S2CID 8138960.
- ^ Родригес Е.А., Кэмпбелл Р.Э., Лин Дж.Й., Лин М.З., Мияваки А., Палмер А.Е., Шу Х, Чжан Дж., Цзянь Р.Й. (февраль 2017 г.). "Набор инструментов для выращивания и свечения флуоресцентных и фотоактивных белков". Тенденции в биохимических науках. 42 (2): 111–129. Дои:10.1016 / j.tibs.2016.09.010. ЧВК 5272834. PMID 27814948.
- ^ Монтесинос-Франьола Ф, Лин Дж.Й., Родригес Э.А. (2020-11-16). «Флуоресцентные белки для визуализации in vivo, где биливердин?». Сделки Биохимического Общества: BST20200444. Дои:10.1042 / BST20200444.
- ^ Бокман Ш., Вард WW (1982). «Обратимая денатурация зеленого флуоресцентного белка Aequorea: физическое разделение и характеристика ренатурированного белка». Биохимия. 21 (19): 4535–4540. Дои:10.1021 / bi00262a003. PMID 6128025.
- ^ Pouwels LJ, Zhang L, Chan NH, Dorrestein PC, Wachter RM (сентябрь 2008 г.). «Исследования кинетического изотопного эффекта на скорость de novo образования хромофора в быстро- и медленно созревающих вариантах GFP». Биохимия. 47 (38): 10111–22. Дои:10.1021 / bi8007164. ЧВК 2643082. PMID 18759496.
- ^ Чудаков Д.М., Мац М.В., Лукьянов С., Лукьянов К.А. (июль 2010 г.). «Флуоресцентные белки и их применение в визуализации живых клеток и тканей». Физиологические обзоры. 90 (3): 1103–63. Дои:10.1152 / физрев.00038.2009. PMID 20664080. S2CID 10767597.
- ^ Степанеко, О.В .; Верхуша, В. В .; Шавловский, М. М .; Кузнецова, И. М .; Уверский, В. Н .; Туроверов, К. К. Понимание роли Arg96 в структуре и стабильности зеленого флуоресцентного белка. Белки: Struct., Funct., Bioinf. 1999, 73 (3), 539-551.
- ^ Джагдер Б. Э., Уэлч Дж., Брейди Н., Маркиз С. П. (2016-07-26). «Создание и использование слияния растворимого промотора гидрогеназы (PSH) Cupriavidus necator H16 с gfp (зеленый флуоресцентный белок)». PeerJ. 4: e2269. Дои:10.7717 / peerj.2269. ЧВК 4974937. PMID 27547572.
- ^ Арун К.Х., Каул С.Л., Рамарао П. (2005). «Зеленые флуоресцентные белки в исследовании рецепторов: новый инструмент для открытия лекарств». Журнал фармакологических и токсикологических методов. 51 (1): 1–23. Дои:10.1016 / j.vascn.2004.07.006. PMID 15596111.
- ^ Сон Й.Х., Ким С.С., Со Дж.Х. (апрель 2016 г.). «Неинвазивный мониторинг токсичности окружающей среды с помощью зеленого флуоресцентного белка, экспрессирующего Escherichia coli». Корейский журнал химической инженерии. 33 (4): 1331–6. Дои:10.1007 / s11814-015-0253-1. S2CID 62828580.
- ^ Пан Y, Leifert A, Graf M, Schiefer F, Thoröe-Boveleth S, Broda J, Halloran MC, Hollert H, Laaf D, Simon U, Jahnen-Dechent W (март 2013 г.). «Высокочувствительный анализ в реальном времени токсичности наночастиц у зеленых флюоресцентных белок-экспрессирующих рыбок данио». Маленький. Weinheim an Der Bergstrasse, Германия. 9 (6): 863–9. Дои:10.1002 / smll.201201173. PMID 23143852.
- ^ Chalfie M (июнь 2009 г.). "GFP: освещая жизнь". Труды Национальной академии наук Соединенных Штатов Америки. 106 (25): 10073–10080. Bibcode:2009PNAS..10610073C. Дои:10.1073 / pnas.0904061106. ЧВК 2700921. PMID 19553219.
- ^ Юсте Р. (декабрь 2005 г.). «Флуоресцентная микроскопия сегодня». Методы природы. 2 (12): 902–4. Дои:10.1038 / nmeth1205-902. PMID 16299474. S2CID 205418407.
- ^ Коморовски М., Финкенштедт Б., Рэнд Д. (июнь 2010 г.). «Использование одного флуоресцентного репортерного гена для определения периода полужизни внешнего шума и других параметров экспрессии гена». Биофизический журнал. 98 (12): 2759–2769. Bibcode:2010BpJ .... 98.2759K. Дои:10.1016 / j.bpj.2010.03.032. ЧВК 2884236. PMID 20550887.
- ^ Гункель М., Эрдель Ф., Риппе К., Леммер П., Кауфманн Р., Хёрманн С., Амбергер Р., Кремер С. (июнь 2009 г.). «Двухцветная локализационная микроскопия клеточных наноструктур». Биотехнологический журнал. 4 (6): 927–38. Дои:10.1002 / biot.200900005. PMID 19548231.
- ^ Чудаков Д.М., Лукьянов С., Лукьянов К.А. (декабрь 2005 г.). «Флуоресцентные белки как инструментарий для визуализации in vivo». Тенденции в биотехнологии. 23 (12): 605–13. Дои:10.1016 / j.tibtech.2005.10.005. PMID 16269193.
- ^ Livet J, Weissman TA, Kang H, Draft RW, Lu J, Bennis RA, Sanes JR, Lichtman JW (ноябрь 2007 г.). «Трансгенные стратегии комбинаторной экспрессии флуоресцентных белков в нервной системе». Природа. 450 (7166): 56–62. Bibcode:2007Натура 450 ... 56л. Дои:10.1038 / природа06293. PMID 17972876. S2CID 4402093.
- ^ Бейкер Б.Дж., Муто Х., Димитров Д., Акеманн В., Перрон А., Ивамото Ю., Джин Л., Коэн Л. Б., Исакофф Е. Ю., Пиерибоне В. А., Хьюз Т., Кнопфель Т. (август 2008 г.). «Генетически кодируемые флуоресцентные сенсоры мембранного потенциала». Клеточная биология мозга. 36 (1–4): 53–67. Дои:10.1007 / s11068-008-9026-7. ЧВК 2775812. PMID 18679801.
- ^ Адесник Х., Николл Р.А., Премьер-министр Англии (декабрь 2005 г.). «Фотоинактивация нативных рецепторов AMPA показывает их трафик в реальном времени». Нейрон. 48 (6): 977–85. Дои:10.1016 / j.neuron.2005.11.030. PMID 16364901.
- ^ Lakadamyali M, Rust MJ, Babcock HP, Zhuang X (август 2003 г.). «Визуализация заражения отдельными вирусами гриппа». Труды Национальной академии наук Соединенных Штатов Америки. 100 (16): 9280–5. Bibcode:2003ПНАС..100.9280Л. Дои:10.1073 / pnas.0832269100. ЧВК 170909. PMID 12883000.
- ^ Джу Ки, Ван П (октябрь 2008 г.). «Визуализация направленной трансдукции с помощью сконструированных лентивирусных векторов». Генная терапия. 15 (20): 1384–96. Дои:10.1038 / gt.2008.87. ЧВК 2575058. PMID 18480844.
- ^ Реми С., Тессон Л., Усал С., Менорет С., Боннамейн В., Нерриер-Дагин В., Россиньоль Дж., Бойер С., Нгуен Т.Х., Навейхан П., Лескодрон Л., Анегон I (октябрь 2010 г.). «Новые линии трансгенных крыс GFP, актуальные для регенеративной медицины и генной терапии». Трансгенные исследования. 19 (5): 745–63. Дои:10.1007 / s11248-009-9352-2. PMID 20094912. S2CID 42499768.
- ^ Эллиотт Дж., Макграт Дж., Крокетт-Тораби Э. (июнь 2000 г.). «Зеленый флуоресцентный белок: новый анализ жизнеспособности для криобиологических приложений». Криобиология. 40 (4): 360–369. Дои:10.1006 / крио.2000.2258. PMID 10924267.
- ^ Fakhrudin N, Ladurner A, Atanasov AG, Heiss EH, Baumgartner L, Markt P, Schuster D, Ellmerer EP, Wolber G, Rollinger JM, Stuppner H, Dirsch VM (апрель 2010 г.). «Компьютерное открытие, проверка и механистическая характеристика новых неолигнановых активаторов гамма-рецепторов, активируемых пролифератором пероксисом». Молекулярная фармакология. 77 (4): 559–66. Дои:10.1124 / моль 109.062141. ЧВК 3523390. PMID 20064974.
- ^ Собери MC, Юн Ш (2011). «Биологические одноклеточные лазеры». Природа Фотоника. 5 (7): 406–410. Bibcode:2011НаФо ... 5..406Г. Дои:10.1038 / nphoton.2011.99. S2CID 54971962.
- ^ Матсон Дж (2011). «Зеленый флуоресцентный протеин для живых лазеров». Scientific American. Получено 2011-06-13.
- ^ Курос-Мехр Х., Бечис С.К., Слорах Е.М., Литтлпейдж Л.Е., Эгеблад М., Эвальд А.Дж., Пай С.Ю., Хо IC, Верб З. (февраль 2008 г.). «GATA-3 связывает дифференцировку и распространение опухоли в модели рака молочной железы в просвете». Раковая клетка. 13 (2): 141–52. Дои:10.1016 / j.ccr.2008.01.011. ЧВК 2262951. PMID 18242514.
- ^ Cabantous S, Terwilliger TC, Waldo GS (январь 2005 г.). «Маркировка и обнаружение белков с помощью сконструированных самособирающихся фрагментов зеленого флуоресцентного белка». Природа Биотехнологии. 23 (1): 102–7. Дои:10.1038 / nbt1044. PMID 15580262. S2CID 25833063.
- ^ Родман М.К., Ядав Н.С., Артус Н.Н. (01.09.2002). «Развитие сайленсинга трансгена, индуцированного геминивирусом, связано с метилированием трансгена». Новый Фитолог. 155 (3): 461–468. Дои:10.1046 / j.1469-8137.2002.00467.x.
- ^ Чжу Й.Дж., Агбаяни Р., Мур PH (апрель 2004 г.). «Зеленый флуоресцентный белок как визуальный маркер отбора для трансформации папайи (Carica papaya L.)». Отчеты о растительных клетках. 22 (9): 660–7. Дои:10.1007 / s00299-004-0755-5. PMID 14749892. S2CID 23198182.
- ^ Нива Ю., Хирано Т., Ёсимото К., Симидзу М., Кобаяши Н. (1999). «Неинвазивное количественное обнаружение и применение нетоксичного зеленого флуоресцентного белка S65T-типа в живых растениях». Журнал растений. 18 (4): 455–63. Дои:10.1046 / j.1365-313X.1999.00464.x. PMID 10406127. S2CID 292648.
- ^ Бейкер С.С., Видикан С.Б., Кэмерон Д.С., Грейб Х.Г., Яроцки С.К., Сетапутри А.В., Спикуцца С.Х., Берр А.А., Вакас М.А., Толберт Д.А. «Эпифлуоресцентная насадка улучшает цифровую фотографию всего растения Arabidopsis thaliana, экспрессирующего зеленый флуоресцентный белок с красным смещением». ЗАВОДЫ AoB. 2012: pls003. Дои:10.1093 / aobpla / pls003. ЧВК 3296078. PMID 22479674.
- ^ «PlantEdDL - Использование цифровых камер SRL в количественных исследованиях растений, экспрессирующих зеленый флуоресцентный белок (GFP)». plantted.botany.org. Получено 2016-03-23.
- ^ Эдуардо Кац. "Зайчик GFP".
- ^ "Светящиеся в темноте неоновые мыши". Архивировано из оригинал 14 февраля 2009 г.. Получено 30 августа, 2016.
- ^ Тайваньские ученые разводят флуоресцентных зеленых свиней
- ^ Wongsrikeao P, Saenz D, Rinkoski T, Otoi T, Poeschla E (2011). «Трансгенез антивирусного фактора рестрикции у домашней кошки». Природные методы. 8 (10): 853–9. Дои:10.1038 / nmeth.1703. ЧВК 4006694. PMID 21909101.
- ^ «Флуоресцентный щенок - первая в мире трансгенная собака».
- ^ Восс-Андреэ Дж (2005). "Белковые скульптуры: строительные блоки жизни вдохновляют искусство". Леонардо. 38: 41–45. Дои:10.1162 / leon.2005.38.1.41. S2CID 57558522.
- ^ Павляк А (2005). «Протеин Inspirierende». Физический журнал. 4: 12.
- ^ "Скульптура Джулиана Восс-Андреэ". Получено 2007-06-14.
дальнейшее чтение
- Пьерибон В., Грубер Д. (2006). Сияние в темноте: революционная наука биофлуоресценции. Кембридж: Белкнап Пресс. ISBN 978-0-674-01921-8. OCLC 60321612. Научно-популярная книга, описывающая историю и открытие GFP
- Циммер М (2005). Светящиеся гены: революция в биотехнологии. Буффало, Нью-Йорк: Книги Прометея. ISBN 978-1-59102-253-4. OCLC 56614624.
внешние ссылки
| Библиотечные ресурсы около Зеленый флуоресцентный белок |
- Подробная статья о флуоресцентных белках в Scholarpedia
- Краткое изложение важных документов GFP
- Интерактивный Java-апплет, демонстрирующий химию, лежащую в основе образование хромофора GFP
- Видео с лекции Роджера Цзяня о флуоресцентных белках, получившей Нобелевскую премию
- Спектры возбуждения и испускания различных флуоресцентных белков
- Тематический выпуск Green Fluorescent Protein Chem Soc Rev посвящена лауреатам Нобелевской премии по химии 2008 г., профессорам Осаму Шимомура, Мартин Чалфи и Роджер Ю. Цзянь
- Молекула месяца, июнь 2003 г.: иллюстрированный обзор GFP Дэвида Гудселла.
- Молекула месяца, июнь 2014 г.: иллюстрированный обзор GFP-подобных вариантов Дэвида Гудселла.
- Зеленый флуоресцентный белок on FPbase, база данных флуоресцентных белков
- Обзор всей структурной информации, доступной в PDB для UniProt: P42212 (Зеленый флуоресцентный белок) на PDBe-KB.