Кариогамы - Karyogamy

Кариогамы это последний шаг в процессе объединения двух гаплоидный эукариотический клеток, и относится конкретно к слиянию двух ядра. До кариогамии каждая гаплоидная клетка имела одну полную копию организма. геном. Для того, чтобы произошла кариогамия, необходимо клеточная мембрана и цитоплазма каждой ячейки должны сливаться с другой в процессе, известном как плазмогамия. Оказавшись внутри соединенной клеточной мембраны, ядра называются пронуклеусы. Как только клеточные мембраны, цитоплазма и пронуклеусы сливаются вместе, в результате получается единичная клетка. диплоид, содержащий две копии генома. Эта диплоидная клетка, называемая зигота или же зигоспора затем можно войти мейоз (процесс дупликации, рекомбинации и деления хромосом с образованием четырех новых гаплоидных клеток) или продолжить деление на митоз. При оплодотворении млекопитающих используется аналогичный процесс для объединения гаплоидных сперматозоидов и яйцеклеток (гаметы ) для создания диплоидного оплодотворенного яйца.
Термин кариогамия происходит от Греческий карио- (от κάρυον карион), что означает «орех» и γάμος гамос, что означает «брак».[1]
Важность гаплоидных организмов
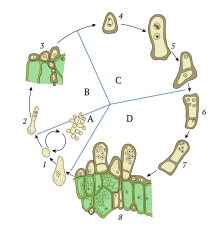
Гаплоидные организмы, такие как грибы, дрожжи, и водоросли может иметь сложный клеточные циклы, в котором выбор между сексуальный или же бесполое размножение жидкий и часто подверженный влиянию окружающей среды. Некоторые организмы, в дополнение к их обычному гаплоидному состоянию, могут также существовать как диплоидные в течение короткого времени, что позволяет генетическая рекомбинация происходить. Кариогамия может происходить в любом режиме воспроизводства: во время полового цикла или в соматический (не репродуктивные) клетки.[2]
Таким образом, кариогамия является ключевым шагом в объединении двух наборов различного генетического материала, который может рекомбинировать во время мейоза. У гаплоидных организмов, у которых отсутствуют половые циклы, кариогамия также может быть важным источником генетическая вариация в процессе формирования соматического диплоид клетки. Образование соматических диплоидов обходит процесс гамета образуются во время цикла полового размножения и вместо этого создают вариации в соматических клетках уже развитого организма, таких как грибок.[2]
Роль в половом размножении

Роль кариогамии в половом размножении проще всего продемонстрировать одноклеточные гаплоидный организмы, такие как водоросли рода Хламидомонада или дрожжи Saccharomyces cerevisiae. Такие организмы обычно существуют в гаплоидном состоянии, содержащем только один набор хромосом на клетку. Однако механизм остается в основном одинаковым для всех гаплоидных эукариот.[3]
При воздействии окружающей среды, например, при азотном голодании в случае Хламидомонада, клетки вынуждены образовывать гаметы.[4] Формирование гамет у одноклеточных гаплоидных организмов, таких как дрожжи, называется спороношение, в результате чего происходит множество клеточных изменений, повышающих устойчивость к стрессу. Образование гамет у многоклеточных грибов происходит в гаметангия, орган, специализирующийся на таком процессе, обычно за счет мейоза.[5] Когда напротив типы вязки встречаются, они вынуждены покинуть вегетативный цикл и войти в цикл спаривания. У дрожжей существует два типа спаривания, а и а.[6] У грибов может быть два, четыре или даже до 10 000 типов спаривания, в зависимости от вида.[7][8] Распознавание партнера у простейших эукариот достигается за счет феромон передача сигналов, которая индуцирует образование shmoo (проекция клетки) и запускает процесс организации и миграции микротрубочек. Феромоны, используемые для распознавания типа спаривания, часто представляют собой пептиды, но иногда и триспоровую кислоту или другие молекулы, распознаваемые клеточными рецепторами противоположной клетки. Примечательно, что передача сигналов феромона отсутствует у высших грибов, таких как грибы.[3]
Клеточные мембраны и цитоплазма этих гаплоидных клеток затем сливаются вместе в процессе, известном как плазмогамия. В результате получается одна клетка с двумя ядрами, известная как пронуклеусы. Затем пронуклеусы сливаются вместе в хорошо регулируемом процессе, известном как кариогамия. Это создает диплоидную клетку, известную как зигота, или зигоспора,[4] который затем может войти мейоз, процесс дупликации, рекомбинации и деления хромосом для создания четырех новых гаплоидных клеток гамет. Одним из возможных преимуществ полового размножения является то, что оно приводит к большей генетической изменчивости, предоставляя возможность адаптации посредством естественного отбора. Еще одно преимущество - эффективная рекомбинационная репарация повреждений ДНК во время мейоза. Таким образом, кариогамия является ключевым шагом в объединении разнообразного генетического материала, чтобы гарантировать рекомбинацию в мейозе.[3]
В Амебозоа представляет собой большую группу в основном одноклеточных видов, которые, как недавно было установлено, обладают механизмами кариогамии и мейоз.[9] Поскольку Amoeboza рано отделилась от генеалогического древа эукариот, это открытие предполагает, что кариогамия и мейоз присутствовали на ранней стадии развития. эукариотический эволюция.
Клеточные механизмы
Пронуклеарная миграция
Конечная цель кариогамии - слияние двух гаплоидных ядер. Первым шагом в этом процессе является движение двух пронуклеусов друг к другу, которое происходит сразу после плазмогамии. Каждый пронуклеус имеет корпус полюса шпинделя который встроен в ядерная оболочка и служит точкой крепления для микротрубочки. Микротрубочки, важный волокноподобный компонент цитоскелет, выходят из корпуса полюса шпинделя. Точка крепления к корпусу полюса шпинделя отмечает отрицательный конец, а положительный конец проходит в цитоплазма. Плюс имеет нормальные роли в митотическое деление, но во время ядерной конгресса положительные стороны перенаправляются. Плюс концы микротрубочек прикрепляются к противоположному пронуклеусу, в результате чего два пронуклеуса притягиваются друг к другу.[10]
Движение микротрубочек опосредуется семейством моторные белки известный как кинезины, например, Kar3 в дрожжах. Дополнительные белки, такие как Spc72 в дрожжах, действуют как клей, соединяющий моторный белок, корпус полюса шпинделя и микротрубочка в структуре, известной как полумост. Другие белки, такие как Kar9 и Bim1 у дрожжей, прикрепляются к плюсовому концу микротрубочек. Они активируются сигналами феромона, которые прикрепляются к кончику шму. Шму - это проекция клеточной мембраны, которая является местом первоначального слияния клеток в плазмогамии. После плазмогамии плюс концы микротрубочек продолжают расти в направлении противоположного пронуклеуса. Считается, что растущий положительный конец микротрубочки прикрепляется непосредственно к моторному белку противоположного пронуклеуса, запуская реорганизацию белков на полумосту. Сила, необходимая для миграции, возникает непосредственно в ответ на это взаимодействие.[11]
Были предложены две модели ядерной конгресса: скользящая перемычка и модель с положительным концом. В модели скользящего поперечного моста микротрубочки проходят антипараллельный друг к другу на всем расстоянии между двумя пронуклеусами, образуя перекрестные ссылки друг к другу, и каждое присоединяется к противоположному ядру на плюсовом конце. Это излюбленная модель. Альтернативная модель предполагает, что положительные концы контактируют друг с другом на полпути между двумя пронуклеусами и лишь слегка перекрываются. В любой модели считается, что укорочение микротрубочек происходит на плюсовом конце и требует наличия Kar3p (у дрожжей), члена семейства кинезин -подобные белки.[10]
Микротрубочка организация в цитоскелет было показано, что он необходим для правильного ядерного конгресса во время кариогамии. Дефектная организация микротрубочек вызывает полное нарушение кариогамии, но не прерывает полностью мейоз и производство спор у дрожжей. Нарушение происходит потому, что процесс ядерной конгресса не может происходить без функциональных микротрубочек. Таким образом, пронуклеусы не подходят достаточно близко друг к другу, чтобы слиться вместе, и их генетический материал остается разделенным.[12]
Пронуклеарное слияние (кариогамия)
Слияние ядерные оболочки Пи происходит в три этапа: слияние внешней мембраны, слияние внутренней мембраны и слияние тел полюсов веретена. В дрожжах несколько членов семейства белков Kar, а также протамин, необходимы для слияния ядерных мембран. Протамин Prm3 расположен на внешней поверхности каждой ядерной мембраны и необходим для слияния внешней мембраны. Точный механизм неизвестен. Kar5, кинезин-подобный белок, необходим для увеличения расстояния между внешней и внутренней мембранами в явлении, известном как расширение мостика. Считается, что Kar8 и Kar2 необходимы для слияния внутренних мембран.[13]Как описано выше, реорганизация вспомогательных и моторных белков во время миграции пронуклеусов также служит для ориентации тел полюсов веретена в правильном направлении для эффективной ядерной конгресса. Ядерный конгресс все еще может происходить без этой предварительной ориентации полюсных тел веретена, но он происходит медленнее. В конечном итоге два пронуклеуса объединяют содержимое своих нуклеоплазмы и сформируйте вокруг результата единый конверт.[11]
Роль в соматических диплоидах
Хотя грибы обычно гаплоидны, диплоидные клетки могут возникать по двум механизмам. Первый - это отказ митотического веретена во время регулярного деления клеток, и он не связан с кариогамией. Полученная клетка может быть только генетически гомозиготный так как он производится из одной гаплоидной клетки. Второй механизм, связанный с кариогамией соматических клеток, может производить гетерозиготный диплоиды, если два ядра различаются по генетической информации. Образование соматических диплоидов, как правило, встречается редко и, как полагают, происходит из-за мутации в гене репрессора кариогамии (KR).[2]
Однако есть несколько грибов, которые существуют в основном в диплоидном состоянии. Одним из примеров является грибковые микроорганизмы албиканс, грибок, который обитает в желудочно-кишечном тракте многих теплокровных животных, включая человека. Хотя обычно безобидно, C. albicans может стать патогенным и представляет особую проблему у пациентов с ослабленным иммунитетом. В отличие от большинства других грибов, диплоидные клетки разных типов спаривания сливаются, образуя тетраплоидные клетки, которые впоследствии возвращаются в диплоидное состояние, теряя хромосомы.[14]
Сходства и отличия от оплодотворения млекопитающих
Млекопитающие, включая человека, также объединяют генетический материал из двух источников - отца и матери. оплодотворение. Этот процесс похож на кариогамию. Как и в случае кариогамии, микротрубочки играют важную роль в оплодотворении и необходимы для соединения сперматозоидов и яйцеклетки (ооцит ) ДНК.[15] Такие препараты как гризеофульвин которые мешают микротрубочкам, препятствуют слиянию пронуклеусов сперматозоидов и яйцеклеток. Ген KAR2, который играет большую роль в кариогамии, имеет аналог у млекопитающих под названием Bib / GRP78.[16] В обоих случаях генетический материал объединяется для создания диплоидной клетки, которая имеет большее генетическое разнообразие, чем любой исходный источник.[17] Вместо того, чтобы слиться, как это делают низшие эукариоты при кариогамии, ядро сперматозоидов образует пузырьки, а его ДНК деконденсируется. Центриоль сперматозоидов действует как центр организации микротрубочек и образует звездочку, которая распространяется по всему яйцеклетке, пока не соприкоснется с ядром яйцеклетки. Два пронуклеуса перемещаются друг к другу, а затем сливаются, образуя диплоидную клетку.[18]
Смотрите также
Рекомендации
- ^ «кариогамия». Бесплатный словарь.
- ^ а б c Коул, Гарри Т. (2012-12-02). Биология конидиальных грибов. Эльзевир. ISBN 9780323143547.
- ^ а б c Ni, мин; Марианна Ферецаки; Шэн Сунь; Сюин Ван; Джозеф Хейтман (декабрь 2011 г.). "Секс в грибах". Ежегодный обзор генетики. 45: 405–430. Дои:10.1146 / annurev-genet-110410-132536. ЧВК 3310392. PMID 21942368.
- ^ а б Рэйвен, Питер Х .; Рэй Франклин Эверт; Сьюзан Э. Эйххорн (январь 2005 г.). Биология растений. Макмиллан. ISBN 9780716710073.
- ^ Ünal, E .; А. Амон (01.01.2011). «Образование гамет сбрасывает часы старения дрожжей». Симпозиумы Колд-Спринг-Харбор по количественной биологии. 76: 73–80. Дои:10.1101 / sqb.2011.76.011379. ISSN 0091-7451. ЧВК 3912942. PMID 21890640.
- ^ Гатри, Кристина; Джеральд Р. Финк (2004). Руководство по генетике дрожжей, молекулярной и клеточной биологии. Gulf Professional Publishing. ISBN 9780121827786.
- ^ Волк, Том. "Грибок месяца Тома Волка за февраль 2000 года". Грибки Тома Волка. Университет Висконсин-Ла-Кросс. Получено 14 декабря 2013.
- ^ Ходж, Кэти. «Грибок заходит в бар для одиноких». Блог Cornell Mushroom. Корнелл Университет. Получено 23 ноября 2013.
- ^ Hofstatter PG, Brown MW, Lahr DJG (ноябрь 2018 г.). «Сравнительная геномика поддерживает секс и мейоз у различных амебозоа». Геном Биол Эвол. 10 (11): 3118–3128. Дои:10.1093 / gbe / evy241. ЧВК 6263441. PMID 30380054.
- ^ а б Молк, Джеффри Н .; E. D. Лосось; Керри Блум (02.01.2006). «Ядерная конгрессия обусловлена взаимодействиями цитоплазматических микротрубочек и концов у S. cerevisiae». Журнал клеточной биологии. 172 (1): 27–39. Дои:10.1083 / jcb.200510032. ISSN 0021-9525. JSTOR 4134115. ЧВК 2063526. PMID 16380440.
- ^ а б Жибо, Ромен; Антонио З. Полити; Франсуа Неделек; Клод Антоний; Майкл Кноп (01.02.2013). «Заякоренный в теле полюса веретена Kar3 движет ядро по микротрубочкам из другого ядра в процессе подготовки к слиянию ядер во время кариогамии дрожжей». Гены и развитие. 27 (3): 335–349. Дои:10.1101 / gad.206318.112. ISSN 0890-9369. ЧВК 3576518. PMID 23388829.
- ^ Ямасита, Акира; Ёсихиро Фудзита; Масаюки Ямамото (05.06.2013). «Правильная структура микротрубочек жизненно важна для своевременного прохождения мейоза у делящихся дрожжей». PLoS ONE. 8 (6): e65082. Дои:10.1371 / journal.pone.0065082. ЧВК 3673945. PMID 23755176.
- ^ Меллой, Патриция; Шу Шэнь; Эрин Уайт; Марк Д. Роуз (1 сентября 2009 г.). «Отчетливые роли ключевых белков кариогамии во время слияния ядер дрожжей». Молекулярная биология клетки. 20 (17): 3773–3782. Дои:10.1091 / mbc.E09-02-0163. ISSN 1059-1524. ЧВК 2735476. PMID 19570912.
- ^ Благородный, Сюзанна М .; Джонсон, Александр Д. (декабрь 2007 г.). "Генетика Candida albicans, диплоидного грибкового патогена человека". Ежегодный обзор генетики. 41 (1): 193–211. Дои:10.1146 / annurev.genet.41.042007.170146. PMID 17614788.
- ^ Schatten, G; Симерли, К; Schatten, H (июнь 1985 г.). «Конфигурации микротрубочек во время оплодотворения, митоза и раннего развития у мышей и потребность в опосредованной микротрубочками яйцеклетки подвижности во время оплодотворения млекопитающих». Труды Национальной академии наук Соединенных Штатов Америки. 82 (12): 4152–6. Дои:10.1073 / пнас.82.12.4152. ЧВК 397953. PMID 3889922.
- ^ Роза, Марк Д .; Misra, Leanne M .; Фогель, Джозеф П. (1 июня 1989 г.). «KAR2, ген кариогамии, является дрожжевым гомологом гена BiP / GRP78 млекопитающих». Клетка. 57 (7): 1211–1221. Дои:10.1016/0092-8674(89)90058-5. PMID 2661018.
- ^ Вассарман, премьер-министр; Джовин, L; Личер, ES (февраль 2001 г.). «Профиль оплодотворения у млекопитающих». Природа клеточной биологии. 3 (2): E59–64. Дои:10.1038/35055178. PMID 11175768.
- ^ Wassarman, Paul M .; Джовин, Лука; Личер, Эвелин С. (1 февраля 2001 г.). «Профиль оплодотворения у млекопитающих». Природа клеточной биологии. 3 (2): E59 – E64. Дои:10.1038/35055178. PMID 11175768.