Кошачья щель - Cat gap
В кошачья щель это период в Окаменелости примерно 25-18,5 миллионов лет назад в котором мало окаменелостей кошки или кошачьи виды нашел в Северная Америка. Причина "кошачьей щели" оспаривается, но могла быть вызвана изменениями климата (глобальное похолодание ), изменения среды обитания и относящийся к окружающей среде экосистема, все больше гиперплотоядный тенденция кошек (особенно нимравиды ), вулканическая активность, эволюционный изменения в стоматологический морфология из Canidae видов, присутствующих в Северной Америке, или периодичность вымирания, называемая циклы Ван дер Хаммена.[1]
Эволюция кошек
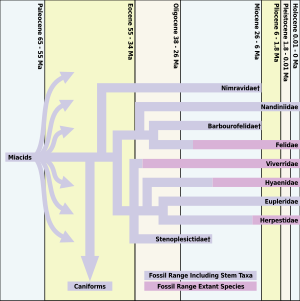
Все современные плотоядные, включая кошек, произошли от миакоиды, который существовал примерно от 66 до 33 миллионов лет назад. Были и другие более ранние кошачьи виды, но Proailurus (что означает «перед кошкой»; также называется «Рассветная кошка Лемана»), появившаяся около 30 миллионов лет назад, обычно считается первой «настоящей кошкой».[2]
После появления рассветной кошки в летописи окаменелостей за 10 миллионов лет мало что говорит о том, что кошки будут процветать. На самом деле, хотя Proailurus сохранялась, по крайней мере, 14 миллионов лет, к концу правления рассветной кошки осталось так мало окаменелостей кошачьих, что палеонтологи называют это «кошачьей пропастью». Переломный момент для кошек наступил с появлением нового рода кошачьих, Псевдэлурус[2]
Увеличение неравенства в раннем миоцене происходит в то время, когда в Северной Америке было обнаружено несколько окаменелостей кошачьих. Гиперплотные кошачьи нимравиды вымерли в Северной Америке после 26 лет. Ма и кошачьи не прибыли в Северную Америку до Средний миоцен с появлением Псевдэлурус. Псевдэлурус перешла в Северную Америку через Берингов мост от выжившего населения в Азия 18,5 миллиона лет назад. Все современные кошки произошли от Псевдэлурус.
Нимравиды и барбурофелиды были саблезубый кот -как животные из семей Nimravidae и Barbourofelidae, соответственно. Хотя не «настоящие кошки» семейства Felidae, они тесно связаны с кошачьими. Nimravidae являются либо базальными кошачьими формами, либо сестринскими группами как кошачьих, так и псовых, в то время как Barbourofelidae являются сестринской группой Felidae. Физически некоторые Nimravidae и Barbourofelidae напоминали саблезубых кошек. Смилодон, который не появится раньше многих миллионов лет спустя. Nimravidae также вымерли в Северной Америке во время «кошачьей пропасти».[3]
Возможные причины
Гиперплотоядность
История плотоядных млекопитающих характеризуется чередой периодов роста и падения разнообразия, в которых наблюдается сокращение клады заменены на филогенетически отчетливые, но функционально похожие клады. За последние 50 миллионов лет последовательные клады мелких и крупных плотоядных млекопитающих разнообразились, а затем пришли к исчезновению. В большинстве случаев причиной упадка были энергетические ограничения и повсеместный отбор на более крупный размер (Правило Копа ), которые приводят к гиперплетение диетическая специализация. Гиперкарниворизм ведет к повышенной уязвимости к исчезновению.
В нимравиды были большими кошачьими животными, которые заняли эту экоморфную нишу в экосистема до 26 Ма. Весьма вероятно, что их гиперкарнивность привела к их исчезновению в Северной Америке. После исчезновения нимравидов не было других кошачьих или кошачий -подобные виды, пока другие кошачьи не прибыли из Евразии после пересечения Берингов мост 18,5 миллиона лет назад. В то время было большое разнообразие среди других хищных млекопитающих в Северной Америке - как гиперплотоядных, так и гиперплотоядных видов - и другие гиперплотоядные виды существовали до, во время и после кошачьей пропасти.
Изменения климата и среды обитания

Другое возможное объяснение исчезновения кошачьих в Северной Америке: изменения в экологии континента. Свидетельства из геологическая запись температуры показывает, что Земля переживала период глобальное похолодание, вызывая леса уступить место саванны.[2] Климатические изменения в засушливых условиях, приглушенные колебания около 25,8 млн лет назад, совпадают с первым появлением свиноподобный креодонты и из карманные суслики, и это тоже начало "кошачьей пропасти" и "энтелодонт пробел », период около 7 миллионов лет, когда в Северной Америке не было нимравидов, кошачьих или энтелодонтов. Фаунал переворот на 25,8 млн. лет является основанием для разделения Арикарийский период времени (30,5–19 млн лет назад) и Arikareen NALMA (Североамериканский возраст наземных млекопитающих) в Монрукреческий период (29,5–25,8 млн лет), а затем Харрисониан период (25,8–23,5 млн лет).[4]
Почему эти кошачьи существа вымерли в Северной Америке (при выживании в Евразии) без замены настоящими кошками? Их судьба может быть обязана тем же факторам, которые создали разнообразие травоядных млекопитающих, поскольку большинству кошек нужен лес или укрытие, из которого они могут охотиться. Во все более открытой Америке нимравиды могли оказаться без экологического окуня для охоты, особенно если конкуренция с собаками помешала им колонизировать саванны.[5]
Другой
Вулканическая активность также рассматривается как возможная причина пропасти кошек, а также других вымираний в течение этого периода времени. В Ла Гарита Кальдера это большой вулканический кальдера расположен в Горы Сан-Хуан на юго-западе Колорадо, США, и является одной из многих кальдер, образовавшихся во время массивного игнимбрит вспышки в Колорадо, Юте и Неваде во время Олигоцен Эпоха. Кальдера Ла Гарита была местом Извержение Рыбного каньона, огромное извержение около 27 миллионов лет назад. Масштабы извержения Рыбного каньона были намного больше, чем что-либо известное в истории человечества (извержение более 10 000 км3 или 2400 куб. миль за VEI 8+) и, возможно, было самым энергичным событием на Земле со времен Chicxulub удар, который, по мнению многих палеонтологов, вызвал вымирание из динозавры в Меловое – палеогеновое вымирание. Возникший в результате взрывной вулканизм, вероятно, выбросил большое количество пыли и мусора в стратосфера вызывая сильное охлаждение (см. вулканическая зима ). Климатические эффекты также могли быть вызваны сера выброшен в стратосфера, который быстро преобразуется в серная кислота, аэрозоль который охлаждает тропосфера путем блокировки входящих солнечная радиация.
Другой возможной причиной кошачьей щели могла быть Поздний кайнозойский ледниковый период это началось 33,9 миллиона лет назад. Этот ледниковый период вызвал оледенение в Антарктида что в конечном итоге распространилось на Арктический регионы южных Аляска, Гренландия, и Исландия. Ледники на североамериканском континенте, а также тенденция к похолоданию, могли сделать экосистему непригодной для проживания кошачьих видов, хотя и пригодных для жизни в холодную погоду. Caniformia такие виды, как псовые (собачьи породы), куньих (ласкоподобные виды) и урсиды (медвежьи виды).
Есть также свидетельства того, что в миоцене подоконник, окружающий Северный Ледовитый океан, известный как Гренландско-Шотландский хребет, просел, позволив большему количеству холодной полярной воды уйти в Северную Атлантику. По мере роста солености Северной Атлантики и увеличения оттока холодной полярной воды термохалинная циркуляция увеличилась в силе, обеспечивая мягкие зимние температуры и большое количество влаги в Северной Атлантике, что является предпосылкой для образования крупных континентальных ледяных шапок на прилегающих холодных континентах.[6]
Эволюция псовых во время разрыва

Некоторые предполагали, что в результате кошачьей щели Caniforms (собачьи виды, включая псовых, медведей, ласки и другие родственные таксоны) эволюционировали, чтобы заполнить более плотоядных и гиперплотоядный экологические ниши которые иначе были бы заполнены кошками.[7] Этот вывод, однако, оспаривается.[8]
Во время или незадолго до этого «кошачьего промежутка» у многих видов псовых развиваются кошачьи черты, указывающие на гиперпарнивность, такие как уменьшенная морда, несколько увеличенные клыки и довольно резкое уменьшение их раздавливания коренные зубы. В Северной Америке первой группой собак среднего размера, которая двинулась в сторону гиперкарнавии, были эндемичный геспероционин псовые, с тремя родами (Паренгидроцион, Энгидроцион, и Мезоцион ), размером от шакалов до маленьких койотов, появившихся в начале Арикарийский (около 28 млн лет назад). Примечательно, что эти три эволюционировали вместе с последним гиенодонт а остальные три нимравида, два из которых были пума -размер. К маленьким гиперплотоядным псовым вскоре присоединились и в конечном итоге на смену им пришли многочисленные виды из других семейств, у которых также развились более специализированные зубы и черепа для мясоедения. Сюда входили, по крайней мере, три более крупных рода одинаково адаптированных амфициониды, один эндемик (Дафенодон ) и два из Старого Света (Temnocyon и Маммоцон ), размером с леопарда куньих (Megalictis ), а также двух гиперплотных медведей, гемиционины Цефалогала и Фобероцион.[7]
Однако другие палеонтологи не согласны с этим выводом:
Было высказано предположение, что у псовых эволюционировала морфология гиперплотоядных, потому что в этот период отсутствовали кошачьи формы («кошачья щель», 26–16 млн лет назад). Представленные здесь данные не подтверждают эту гипотезу. В расчетных морфопространство... Псовые никогда не занимают область морфопространства, в которой кошачьи, нимравиды и гиперплотные креодонты найдены. Однако более актуальным для рассматриваемого вопроса является то, что большинство этих гиперплотоядных собак присутствовало до исчезновения нимравид, и все вымерли до появления кошачьих ... «кошачья щель». 28–20 млн лет характеризуются интенсивностью вымирания выше средней и интенсивностью образования ниже средней. 20 млн. Лет ознаменовались увеличением интенсивности зарождения, а 18 млн. Лет показали снижение интенсивности вымирания и значительное увеличение интенсивности зарождения. Тем не менее, несмотря на увеличение интенсивности возникновения и снижение интенсивности вымирания ближе к концу «кошачьей пропасти» (20–16 млн лет назад), до иммиграции кошачьих в Северную Америку существенного вторжения в морфопространство гиперплотоядных не было ».[8]
Рекомендации
- ^ Т. Дж. Михан; Л. Д. Мартин (2003). «Исчезновение и повторная эволюция подобных адаптивных типов (экоморфов) у кайнозойских североамериканских копытных и хищников отражает циклы Ван-дер-Хамменса» (PDF). Naturwissenschaften. 90 (3): 131–135. Bibcode:2003NW ..... 90..131M. Дои:10.1007 / s00114-002-0392-1. PMID 12649755. Получено 2008-11-28.
- ^ а б c Хантер, Люк; Хайнде, Джеральд (2006). Кошки Африки: поведение, экология и сохранение. Кейптаун: Издательство Университета Джона Хопкинса. С. 40–42. ISBN 978-0-8018-8482-5.
- ^ Р. М. Джокель; Стефан Пейнеб; Роберт М. Хант; Роберт И. Сколник (2002). «Слуховая область и носовая полость олигоценовых Nimravidae». Журнал палеонтологии позвоночных. 22 (4): 131–135. Дои:10.1671 / 0272-4634 (2002) 022 [0830: TARANC] 2.0.CO; 2. Получено 2008-11-28.
- ^ Грегори Дж. Реталлак (2004). «Палеопочвы позднего олигоцена и дерново-луговые палеопочвы раннего миоцена в центральном Орегоне, США» (PDF). Палеогеография, палеоклиматология, палеоэкология. 207 (3–4): 203–237. Bibcode:2004ППП ... 207..203Р. Дои:10.1016 / j.palaeo.2003.09.027. Архивировано из оригинал (PDF) на 2008-08-30. Получено 2008-11-28.
- ^ Фланнери, Тим (2002). Вечные рубежи: экологическая история Северной Америки и ее народов. Нью-Йорк: Grove Press. С. 113–114. ISBN 0-8021-3888-8.
- ^ Хаггарт, Б.А. (2000). "Теории ледникового периода". Оксфордский компаньон Земли. Нью-Йорк: Издательство Оксфордского университета.
- ^ а б c Ван Валкенбург, Блэр (1999). «Основные закономерности в истории хищных млекопитающих». Ежегодный обзор наук о Земле и планетах. 27 (1): 463–493. Bibcode:1999AREPS..27..463V. Дои:10.1146 / annurev.earth.27.1.463.
- ^ а б Джина Д. Уэсли-Хант (2005). «Морфологическая диверсификация хищников Северной Америки». Палеобиология. 31 (1): 35–55. Дои:10.1666 / 0094-8373 (2005) 031 <0035: TMDOCI> 2.0.CO; 2. Получено 2008-11-28.
