Эволюция волка - Evolution of the wolf

В эволюция волка произошло за геологическая шкала времени не менее 300 тысяч лет. Серый волк Canis lupus это высокоадаптивный вид, способный существовать в различных средах и широко распространенный в Голарктический. Исследования современных серых волков выявили отдельные субпопуляции, которые живут в непосредственной близости друг от друга.[2][3] Эти различия в субпопуляциях тесно связаны с различиями в среде обитания - осадками, температурой, растительностью и специализацией добычи - которые влияют на черепно-зубную пластичность.[4][5][6][7]
Археологические и палеонтологические данные свидетельствуют о постоянном присутствии серых волков, по крайней мере, последние 300 000 лет.[8] Это постоянное присутствие контрастирует с геномным анализом, который предполагает, что все современные волки и собаки происходят от общей предковой популяции волков.[9][10][11] Который существовал всего 20 000 лет назад.[9] Эти анализы указывают на узкое место населения с последующим быстрым излучением от предкового населения во время или сразу после Последний ледниковый максимум. Это означает, что первоначальные популяции волков уступили место новому типу волков, который их заменил. Однако географическое происхождение этого излучения неизвестно.
Окаменелости

Летопись окаменелостей древних позвоночных состоит из редко встречающихся фрагментов, из которых зачастую невозможно получить генетический материал. Исследователи ограничены морфологический анализ но трудно оценить внутривидовые и межвидовые вариации и взаимосвязи, которые существовали между особями во времени и месте. Некоторые наблюдения обсуждаются исследователями, которые не всегда согласны с этим, а гипотезы, поддерживаемые одними авторами, оспариваются другими.[12]
Существует общее согласие относительно древнейшей записи, которая показывает, что фелиформ и Caniforms возник в суперсемейке Carnivoramorpha 43 миллиона лет до настоящего (YBP).[13] К собакообразным относился род лисеподобных. Лептоцион различные виды которых существовали от 34 миллионов YBP до того, как 11,9 миллионов YBP разветвлялись на Vulpes (лисы) и Канини (клыки). Размером с шакала Евцион существовали в Северной Америке от 10 миллионов YBP и Ранний плиоцен около 6–5 млн YBP койотоподобный Eucyon davisi[14] вторглись в Евразию. В Северной Америке это дало начало раннему Canis который впервые появился в Миоцен (6 миллионов YBP) на юго-западе США и в Мексике. На 5 млн YBP больше Canis lepophagus появился в том же регионе.[15]:стр58
Псовые, иммигрировавшие из Северной Америки в Евразию - Евцион, Vulpes, и Nyctereutes - были хищниками малого и среднего размера в течение позднего миоцена и раннего плиоцена, но не были главными хищниками. Положение псовых изменится с прибытием Canis стать доминирующим хищником на всем Голарктический. Размером с волка С. chihliensis появились в северном Китае в среднем плиоцене около 4–3 млн. лет назад.[15]:p148 Большой размером с волка Canis появился в середине Плиоцен около 3 миллионов лет назад в бассейне Юшэ, Шаньси Провинция, Китай. 2,5 миллиона лет назад его ареал включал бассейн Нихэван в Уезд Янюань, Хэбэй, Китай и Куруксай, Таджикистан.[16] За этим последовал взрыв Canis эволюция через Евразию в раннем плейстоцене около 1,8 миллиона YBP в том, что обычно называют волк событие. Это связано с формированием мамонтовая степь и континентальное оледенение. Canis распространился в Европу в виде С. arnensis, C. etruscus, и C. falconeri.[15]:p148
Летопись окаменелостей неполна, но вполне вероятно, что волки произошли от популяции небольших ранних псовых.[17]:p241 Морфологические свидетельства[17]:p239[18] и генетические доказательства[19] оба предполагают, что волки эволюционировали во время Плиоцен и рано Плейстоцен эпохи той же линии, которая также привела к появлению койота,[17]:p239 с ископаемыми образцами, указывающими на то, что койот и волк разошлись от общего предка 1,5 миллиона лет назад.[17]:p240[18] Предок шакала и других дошедших до нас представителей рода Canis отделились от линии до этого времени.[17]:p240
После этого отделения от общего предка виды, которые, как считалось, были вовлечены в дальнейшую эволюцию волка и койота, и верования некоторых палеонтологи - разошлись.[17]:p240 Ряд исследователей полагали, что линии C. priscolatrans, C. etruscus, C. rufus и С. волчанка были компоненты, которые каким-то образом привели к появлению современных волков и койотов.[17]:p240[20][21][22][23][24][25]
| Эволюция волка | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Предлагаемая эволюция и ветвление рода Canis к волку.[17]:p240 |
Canis lepophagus
Canis lepophagus жил в раннем плиоцене в Северной Америке.[15] Kurten предложил, чтобы Blancan С. lepophagus[26] полученный из более мелких Миоцен Canis вид в Северной Америке. Затем он получил широкое распространение в Евразии, где был либо идентичен, либо тесно связан с ним. С. arnensis Европы.[17]:p241[27][28]
Джонстон описывает С. lepophagus у них более тонкий череп и скелет, чем у современного койота.[29]:385 Роберт М. Новак обнаружил, что у ранних популяций были маленькие, изящные черепа с узкими пропорциями, которые напоминали маленьких койотов и, по-видимому, были предками C. latrans.[17]:p241 Джонсон отметил, что у некоторых экземпляров, найденных в каньоне Сита, штат Техас, черепа были больше и шире.[29] и вместе с другими фрагментами Новак предположил, что они эволюционировали в волков.[17]:p241[18]
Тедфорд не согласился с предыдущими авторами и обнаружил, что его черепно-дентальная морфология лишена некоторых характеристик, характерных для С. волчанка и C. latrans, и поэтому близких отношений не было, но предполагалось С. lepophagus был предком как волков, так и койотов.[30]:p119
Canis priscolatrans
Canis priscolatrans жил в позднем плиоцене - раннем плейстоцене в Северной Америке.[18] Первый определенный волк появился в конце Blancan /Рано Ирвингтонский,[17]:p240[18][31] и назвал C. priscolatrans это было либо очень близко к[27][28] или синоним для Canis edwardii.[17]:p241[18]:82[32][33] Это напоминало C. rufus по размеру и пропорциям черепа, но с более сложными зубными рядами.[17]:p241 Однако окаменелостей нет. C. rufus до позднего ранчо.[17]:p242
Куртен был неуверен, если C. priscolatrans полученный из С. lepophagus и С. arnensis,[28] но считал, что C. priscolatrans была популяцией крупных койотов, которые были предками Ранчолабрейский и недавний C. latrans. Он отметил, что С. arnensis Европы продемонстрировали поразительное сходство с C. priscolatrans, и они могли представлять то, что когда-то было голарктической популяцией койотов.[27]:стр. 27 Новак не согласился и считал, что C. priscolatrans был аналогом европейского C. etruscus.[18] Куртен позже предположил, что оба C. priscolatrans и C. etruscus были частью группы, которая привела к С. волчанка но не был уверен, развились ли они отдельно от С. lepophagus или возможный общий предок, который произошел от С. lepophagus.[28]
Останки более крупного койота Canis edwardii были найдены в более поздних Плиоцен на юго-западе США вместе с С. lepophagus, что указывает на спуск.[15]:стр60 Тедфорд признал C. edwardii[34] и обнаружили, что черепно-дентальная морфология C. priscolatrans попал внутрь C. edwardii так что название вида C. priscolatrans было сомнительно (номен дубиум).[30]:p131
Canis armbrusteri
Североамериканские волки стали крупнее, образцы зубов указывают на то, что C. priscolatrans разошлись в большом волке C. armbrusteri.[17]:p242[35] в период среднего плейстоцена в Северной Америке.[18] Роберт А. Мартин не согласился и считал, что C. armbrusteri[36] был С. волчанка.[23] Новак не согласился с Мартином и предложил C. armbrusteri не был связан с С. волчанка но C. priscolatrans, что затем привело к C. dirus. Тедфорд предложил, чтобы южноамериканский C. gezi и C. nehringi разделяют сходство зубов и черепа, разработанное для гиперкарнивора, что предполагает C. armbrusteri был общим предком C. gezi, C. nehringi и C. dirus.[30]:148
Canis dirus
В 1908 г. палеонтолог Джон Кэмпбелл Мерриам начал извлекать многочисленные окаменелые фрагменты костей большого волка на ранчо Ла Бреа смоляные ямы. К 1912 году он нашел скелет, достаточно полный, чтобы можно было формально распознать эти и ранее найденные образцы под названием С. Дирус (Лейди 1858).
Canis dirus[37] жили в позднем плейстоцене - начале голоцена в Северной и Южной Америке. [38] и был самым большим из всех Canis виды.[15]:52 В 1987 году новая гипотеза предположила, что популяция млекопитающих могла дать начало более крупной форме, называемой гиперморфом, в те времена, когда пищи было в изобилии, но когда пищи позже стало мало, гиперморф либо приспособился к меньшей форме, либо вымер. Эта гипотеза может объяснить большие размеры тела, обнаруженные у многих позднеплейстоценовых млекопитающих по сравнению с их современными аналогами. И вымирание, и видообразование - новый вид, отделяющийся от более старого - может происходить вместе в периоды экстремальных климатических явлений.[39][40] Глория Д. Гуле согласился с Мартином и предположил, что эта гипотеза может объяснить внезапное появление С. Дирус в Северной Америке, и это из-за схожести форм их черепа, С. волчанка дала начало С. Дирус гиперморфизм из-за обилия игры, стабильной среды и крупных конкурентов.[41] Новак, Куртен и Берта не согласились с Гуле и предложили C. dirus не был получен из С. волчанка.[18][28][42] Три известных палеонтолога Сяомин Ван, Р. Х. Тедфорд и Р. М. Новак предложили, что C. dirus произошел от C. armbrusteri,[15]:p52[30]:181 Новак заявил, что были образцы из пещеры Камберленд, штат Мэриленд, которые указывали на C. armbrusteri расходясь в C. dirus.[17]:p243[43] Эти два таксона имеют ряд общих характеристик (синапоморфия ), что предполагает происхождение C. dirus в конце Ирвингтонский на открытой местности на среднем континенте, а затем расширился на восток и вытеснил своего предка C. armbrusteri.[30]:181
вирус – волчанка гибриды

Мерриам назвал 3 необычных вида на основе образцов, обнаруженных на ранчо. Смоляные ямы Ла Бреа. Новак считал их таксономические синонимы для Canis lupus.[18]
Canis occidentalis furlongi (Мерриам, 1910 г.)[44] описывался как волк, значительно меньший, чем ужасный волк, и более близкий к лесному волку. Западная волчанка Canis lupus occidentalis. Однако его премоляр P4 (верхний хищники ) были массивными, а лицемер из моляр M1 был больше, чем у ужасного волка. Зубы и нёбо одного экземпляра описаны как находящиеся между ужасным волком и Западная волчанка Canis lupus occidentalis.[44] Новак предложил название Canis lupus furlongi так как он считал, что это подвид серого волка.[18] Существует вероятность того, что волки, живущие в маргинальных районах, привели к гибридам ужасных и серых волков.[45]
Canis milleri (Merriam 1912),[46] Волк Миллера был таким же большим, как лесной волк, но с более короткой и тяжелой головой. Его череп и зубной ряд были описаны как промежуточные между Западная волчанка Canis lupus occidentalis и ужасный волк. Его череп отличался от западный из-за его более широкого черепа, особенно во вкусе, и размера его P4 и M1 были намного больше, чем у любого известного деревянного волка, при этом P4 приближался к размеру ужасного волка.[47] Новак считает его таксономическим синонимом Canis lupus furlongi.[18]
Aenocyon milleri (Мерриам, 1918 г.)[48] описывался как волк, отличающийся от лютого волка своим меньшим размером, низким сагиттальным гребнем и менее заметным выступом, но ближе к лютому волку, чем к древесному волку. Был найден только один экземпляр. Новак считает его таксономическим синонимом Canis lupus furlongi.[18]
Canis mosbachensis

Canis mosbachensis, иногда известный как волк Мосбаха, это вымерший маленький волк, который когда-то населял Евразию с Средний к Поздний плейстоцен.[49] Филогенетическое происхождение современного волка С. волчанка от C. etruscus через C. mosbachensis широко распространено.[49][17]:239–245 В 2010 году исследование показало, что разнообразие Canis группа уменьшилась к концу Ранний плейстоцен к Средний плейстоцен и был ограничен в Евразии маленькими волками C. mosbachensis – C. variabilis группы, которые были сопоставимы по размеру с существующими Индийский волк (Canis lupus pallipes), и большой гиперплот Canis (Xenocyon) lycaonoides это было сопоставимо по размеру с существующими северными серыми волками.[8]
Canis variabilis
В 2012 году исследование волкоподобных Canis виды древнего Китая под руководством Сяомин Вана обнаружили, что все они были довольно близки к С. волчанка как в дентальных, так и в посткраниальных размерах, за исключением Canis variabilis, что было «очень странно» по сравнению с другими Canis в Китае, поскольку он имел гораздо меньшие черепно-дентальные размеры, чем более ранние и более поздние виды.[50] Исследование пришло к выводу, что «Весьма вероятно, что этот вид является предком домашней собаки. Собаки фамильярные, гипотеза, которая была предложена предыдущими авторами ".[51][52][53][54][55]
Canis chihliensis
| Эволюция волка - альтернативное предложение | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Предлагаемая эволюция и ответвление от Евцион к волку.[15]:p148[30]:p181 |
Ван и Тедфорд предположил, что род Canis был потомком койотовидных Евцион Дэвиси, а его остатки впервые появились в Миоцен (6 миллионов YBP) на юго-западе США и Мексике. Посредством Плиоцен (5 млн YBP), тем больше Canis lepophagus появились в том же регионе и Ранний плейстоцен (1 миллион YBP) Canis latrans (в койот ) существовало. Они предположили, что прогрессия от Eucyon davisi к C лепофаг для койота была линейная эволюция.[15]:стр58 Дополнительно, C. edwardii, C. latrans и C. aureus образуют небольшую кладу и потому что C. edwardii появился раньше всего в период от середины бланкана (поздний плиоцен) до конца ирвингтона (поздний плейстоцен), он считается предком.[30]:175 180 p.
Новак и Тедфорд также считали, что это возможно С. волчанка происходить от линии псовых миоцена или плиоцена, которая предшествовала и была отдельной от С. lepophagus.[18][31] Основываясь на морфологии из Китая, Плиоцен волк С. chihliensis возможно, был предком для обоих C. armbrusteri и С. волчанка до их миграции в Северную Америку.[15]:p148[30]:p181 С. chihliensis кажется более примитивным и меньшим, чем С. волчанка, а размеры его черепа и зубов аналогичны С. волчанка но посткраниальные элементы его меньше.[50] C. armbrusteri появился в Северной Америке в Средний плейстоцен и волкоподобная форма больше любого Canis в то время.[18] В конце последнего отступления ледников за последние 30 000 лет потепление растопило ледниковые преграды на севере Канады, что позволило арктическим млекопитающим расширить свой ареал до средних широт Северной Америки, включая лосей, карибу, бизонов и серых волков.[15]:стр. 61
В Евразии в среднем плейстоцене C. falconeri дали начало роду гиперплотоядных Ксеноцион, который затем дал начало роду Cuon (волк) и род Ликаон (Африканская охотничья собака).[15]:105 149 стр. Незадолго до появления C. dirus, В Северную Америку вторгся род Ксеноцион это было как C. dirus и более хищные. Летопись окаменелостей показывает, что они редки, и предполагается, что они не могли конкурировать с недавно полученными C. dirus.[15]:стр60 Большой волк C. antonii от позднего плиоцена до раннего плейстоцена Китай оценивался как вариация в пределах С. chihliensis,[30]:p197 и большой волк C. falconeri произошел внезапно в Европе в раннем плейстоцене, возможно, представляя собой расширение на запад C. antonii.[30]:p181
Canis lupus
Раннее Canis lupus Образец был окаменелым зубом, обнаруженным в Old Crow, Юкон, Канада. Экземпляр был найден в отложениях, датированных 1 млн. Лет назад.[30] однако геологическая принадлежность этого осадка подвергается сомнению.[30][56] Немного более молодые экземпляры были обнаружены в Cripple Creek Sump, Фэрбенкс, Аляска, в пластах, датированных 810 000 лет назад. Оба открытия указывают на происхождение этих волков с востока. Берингия вовремя Средний плейстоцен.[30]
Во Франции подвид C. l. lunellensis Бонифай, 1971 г.[57] обнаружен в Люнель-Виль, Эро датированные 400–350 000 лет назад, C. l. santenaisiensis Аргант, 1991[58] от Сантене, Кот-д'Ор датированный 200,000 YBP, и C. lupus maximus Будади-Малинь, 2012 г.[59] из пещеры Яуренс, Nespouls, Коррез от 31000 YBP, показывают прогрессивное увеличение в размере и предлагается хроноподвиды.[60][12] В Италии самые ранние Canis lupus образцы были найдены в Ла-Полледрара-ди-Чеканиббио, в 20 км к северо-западу от Рима, в пластах, датированных 340 000–320 000 лет назад.[61] В 2017 году исследование показало, что размеры верхних и нижних хищных зубов итальянского волка раннего голоцена близки к размерам C. l. Максимус. Колебания размера С. волчанка хищные зубы коррелируют с распространением мегафауны. Итальянский волк претерпел уменьшение размеров тела после потери благородного оленя в Италии в эпоху Возрождения.[12] Предлагаемая линия происхождения:
C. etruscus → C. mosbachensis → C. l. lunellensis → C. l. santenaisiensis → C. l. maximus → К. л. волчанка[12]
В 2020 году появился новый вид волка C. borjgali был обнаружен в Грузия. Предполагается, что этот волк станет следующим эволюционным шагом после C. mosbachensis и предок С. волчанка происхождение.[62]
Canis c.f. фамильяр (Палеолитическая «собака»)
Есть ряд недавно обнаруженных образцов, которые, как предполагается, Палеолитические собаки, однако их таксономия обсуждается. Они были обнаружены либо в Европе, либо в Сибири, и датируются 40 000-17 000 YBP. Они включают Hohle Fels в Германии, Пещеры Гойет в Бельгии, Предмости в Чехии и четыре стоянки в России: пещера Разбойничья, Костёнки-8, Улахан Сулар, Елисеевичи 1. Отпечатки лап из г. Пещера Шове во Франции датированные 26 000 YBP предполагаются как принадлежащие собаке, однако они были оспорены как оставленные волком.[63] Палеолитические собаки были напрямую связаны с человеческими охотничьими лагерями в Европе более 30 000 (YBP), и предполагается, что они были одомашнены. Также предполагается, что они являются протособаками и предками домашней собаки или неизвестным науке типом волков.[64]
Обыкновенная волчанка (домашняя собака)
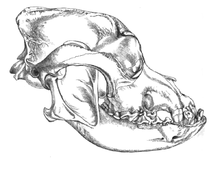

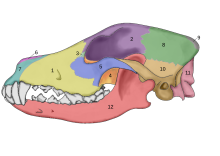
В 2002 году было проведено исследование окаменелых черепов двух крупных псовых, которые были найдены захороненными в нескольких метрах от дверного проема того, что когда-то было хижиной из мамонтовой кости в Елисеевичи-I. Верхний палеолит сайт в Брянская обл. на Русской равнине и с использованием принятого морфологического определения одомашнивания объявили их «собаками ледникового периода». Углеродное датирование дало оценку возраста в календарном году в диапазоне 16 945–13 905 лет назад.[65] В 2013 году в одном из этих черепов было проведено исследование, и его последовательность митохондриальной ДНК была идентифицирована как Обыкновенная волчанка.[66]
В 2015 году зооархеолог заявил, что «с точки зрения фенотипа собаки и волки - принципиально разные животные».[67]
В 1986 году исследование морфологии черепа показало, что домашняя собака морфологически отличается от всех других псовых, кроме волкоподобных. «Разница в размерах и пропорциях между некоторыми породами так же велика, как и между любыми дикими родами, но все собаки явно принадлежат к одному виду».[68] В 2010 году исследование формы черепа собаки по сравнению с дошедшими до нас плотоядные предположил, что «наибольшее расстояние между породами собак явно превосходит максимальное расхождение между видами у хищников. Более того, домашние собаки имеют ряд новых форм за пределами области диких хищников».[69]
Домашняя собака по сравнению с волком показывает самые большие различия в размере и форме черепа (Evans 1979), которые колеблются от 7 до 28 см в длину (McGreevy 2004). Волки долихоцефальный (длинные черепахи), но не такие экстремальные, как у некоторых пород, таких как борзые и русские волкодавы (McGreevy 2004). Собачий брахицефалия (короткомерность) встречается только у домашних собак и связана с педоморфоз (Гудвин 1997). Щенки рождаются с короткой мордой, а более длинный череп долихоцефальных собак появляется в более позднем развитии (Coppinger 1995). Другие различия в форме головы у брахицефальных и долихоцефальных собак включают изменения черепно-лицевого угла (угол между базилярная ось и твердое небо ) (Regodón 1993), морфология височно-нижнечелюстного сустава (Dickie 2001) и рентгенографическая анатомия решетчатая пластина (Шварц 2000).[70]
Новак указал, что угол орбиты глазницы является важной характеристикой, определяющей разницу между собакой и волком, причем волк имеет меньший угол. Новак сравнил орбитальные углы четырех североамериканских клыки (включая индийскую собаку) и дали следующие значения в градусах: койот-42,8, волк-42,8, собака-52,9, ужасный волк-53,1. Орбитальный угол глазницы у собаки был явно больше, чем у койота и волка; почему он был почти таким же, как у лютого волка, не комментировалось.[18]
Многие авторы пришли к выводу, что по сравнению со взрослым существующим волком, у взрослой домашней собаки относительно уменьшенный рострум (передняя часть черепа), повышенный лобная кость, более широкий нёбо, шире череп, и меньшие зубы (Hildebrand1954; Clutton-Brock, Corbet & Hills 1976; Olsen 1985; Wayne 1986; Hemmer 1990; Morey 1990). Другие авторы не согласились и заявили, что эти черты могут перекрываться и варьироваться в пределах двух (Crockford 1999; Harrison 1973). Волчьи детеныши имеют такие же относительные пропорции черепа, как и взрослые собаки, и это было предложено как доказательство того, что домашняя собака является неотенический волк. Было высказано предположение, что это связано либо с человеческим отбором по внешнему виду несовершеннолетних, либо с плейотропный эффект в результате отбора по поведению молоди (Clutton-Brock 1977; Беляев 1979; Wayne 1986; Coppinger and Schneider 1995). Уэйн (1986) пришел к выводу, что образцы его собак не показали значительного относительного укорочения рострума по сравнению с волками, поставив под сомнение эту идентификационную характеристику.[51] Исследование 2004 года, в котором использовались 310 волчьих черепов и более 700 черепов собак, представляющих 100 пород, пришло к выводу, что эволюцию черепов собак, как правило, нельзя описать гетерохронными процессами, такими как неотения, хотя у некоторых педоморфных пород собаки черепа напоминают черепа молодых волков.[71] «Собаки - не педоморфные волки».[72]
По сравнению с волком, зубной ряд собак относительно менее прочен (Olsen 1985; Hemmer 1990), что, как предполагается, связано с ослаблением естественного отбора, когда волки стали комменсальными падальщиками, или с искусственным отбором (Olsen 1985; Clutton-Brock 1995). . Однако Кизер и Греневельд (1992) сравнили нижнечелюстно-дентальные измерения шакалов (C. adustus, C. mesomelas) и песцы накидки (Vulpes chama) собакам эквивалентного размера и обнаружил, что клыки этих других псовых, как правило, были немного меньше, а их вторые коренные зубы были больше по сравнению с собаками, в остальном пропорции были практически одинаковыми у всех видов. Они пришли к выводу, что «... зубы псовых, по-видимому, развивались согласованно друг с другом и относительно независимо от различий в диморфизме, размере или функциональных требованиях». Это ставит под сомнение предположение о том, что собачьи зубы относительно малы из-за недавнего отбора, предполагая, что зубной ряд собаки плезиоморфен от предка, который был меньше волка.[51]
Считается, что уменьшенный размер тела ранней собаки по сравнению с волком связан с выбором ниши (Olsen 1985; Morey 1992; Coppinger & Coppinger 2001). Морей (1992: 199) утверждает, что «Результаты ... согласуются с гипотезой о том, что ранние домашние собаки являются эволюционными педоморфами, продуктами строгого отбора для онтогенетически направленного уменьшения размеров и изменений репродуктивного времени, связанных с новым домашним образом жизни».[51] Однако в эксперименте по приручению одомашненные лисы остались того же размера, что и невыбранные лисы (Trutt 1999: 167).[67]
Уэйн (1986) пришел к выводу, что собака по морфологии черепа ближе к C. latrans, C. aureus, C. adustus, C. mesomelas, Cuon alpinus и Lycaon pictus чем волку. Дар (1942) пришел к выводу, что форма мозга собаки ближе к форме мозга койота, чем к волку. Манвелл и Бейкер (1983) проанализировали работу Дара с добавлением стоматологических данных для собак и пришли к выводу, что предок собаки, вероятно, находился в диапазоне 13,6–20,5 кг, что меньше, чем диапазон 27–54 кг для современных волков (Mech 1970 ) и сравнимо с Динго.[51]
Слуховой булла собаки относительно меньше и более плоской, чем у волка (Harrison 1973; Clutton-Brock, Corbet & Hill 1976; Nowak 1979; Olsen 1985; Wayne 1986), что, как предполагается, связано с ослабленным отбором при одомашнивании, поскольку собака больше не требовалось острого волчьего слуха.Однако было показано, что форма буллы способствует повышенной чувствительности к определенным частотам, но форма и размер могут не коррелировать с остротой зрения (Ewer 1973). Следовательно, наблюдаемая разница может заключаться в том, что собачий булла сохранил свою исконную форму.[51]
Брюшной край собачьей горизонтали. ветвь нижней челюсти имеет выпуклую кривую, которой нет у волка (Olsen 1985; Clutton-Brock 1995), и в литературе нельзя найти обсуждения этого различия. Однако Biknevicius и Van Valkenburgh (1997) заметили, что горизонтальная ветвь хищников, обрабатывающих кости, более толстая в дорсо-вентральном направлении. каудальный к месту обработки кости. Это утолщение могло быть функцией адаптации ниши предком собаки.[51]
Описание морфологии поверхностного мозга шакалов (C. mesomelas, C. aureus), койоты (C. latrans), волки (C. волчанка, C. rufus), а собаки указали, что мозжечок Собаки очень похожи на койота, который тесно связан с шакалами, и что волки демонстрируют многочисленные черты мозга, отличные от других видов (Аткинс и Диллон, 1971). У волков также есть серологические и биохимические признаки, отличные от собак (Leone and Wiens 1956; Lauer, Kuyt & Baker 1969).[51]
Во время последнего ледникового максимума был большой волк. генетическое разнообразие чем сегодня,[9][66] а в популяции серых волков плейстоцена различия между местными средами способствовали появлению ряда экотипов волков, которые были генетически, морфологически и экологически отличны друг от друга.[73] Один автор предположил, что наиболее вероятное объяснение морфологических характеристик собаки по сравнению с волком состоит в том, что предок собаки был приспособлен к другой нише, чем волк.[51]
Генетическая запись
Последовательности ДНК
В митохондрии внутри каждого ячейка содержать много копий небольшого проспекта ДНК геном а у млекопитающих - 16–18 тыс. пар оснований в длину. Клетка содержит сотни или тысячи митохондрий и, следовательно, гены Содержащиеся в этих митохондриях более многочисленны, чем гены, которые встречаются в ядре клетки.[74][75] Обилие митохондриальная ДНК (мДНК) полезна для генетического анализа древних останков, где ДНК деградировала.[75][76]
Митохондриальный Последовательности ДНК иметь более высокий скорость мутации чем частота мутаций ядерных генов, а у млекопитающих она в 5–10 раз выше.[75][77][78] Гены, кодирующие митохондриальные белки, развиваются намного быстрее и являются мощными маркерами для вывода истории эволюции на таких уровнях категорий, как семьи, роды и виды. Однако они эволюционировали более быстрыми темпами, чем другие ДНК-маркеры, и в их молекулярных часах есть временная разница, которую необходимо проверить по другим источникам. Таксономический статус неопределенных видов лучше решить с помощью ядерная ДНК из ядра клетки, что больше подходит для анализа недавней истории.[79] В большинстве случаев мДНК наследуется от материнского предка.[75][80] Следовательно, филогенетический анализ последовательностей мДНК внутри видов обеспечивает историю материнских линий, которая может быть представлена как филогенетическое дерево.[75][81][82]
- От образца к филогенетическому дереву

33500-летний череп «Алтайская собака»

Расположение ядерная ДНК в пределах хромосомы из ядро клетки

Структура части ДНК двойная спираль

Результат Секвенирование ДНК перечисление молекулы ДНК азотистые основания (кодируется как г, C, А или Т )

Молекула ДНК 1 отличается от молекулы ДНК 2 одним базовая пара место, называемое однонуклеотидный полиморфизм (SNP мутация )

Филогенетическое дерево исследуемых волков






| Филогенетическое дерево | |||||||||||||||||||||
| |||||||||||||||||||||
| Филогенетические отношения между четырьмя псовыми.[83][84] |
Мутации, которые отличаются в этих 4 последовательности пронумерованы и выделены жирным шрифтом. Эти мутации затем можно использовать для построения филогенетическое дерево для четырех псовых. В этом примере собака и серый волк отличаются двумя заменами (выделены красным), и каждая из них отличается от койота четырьмя заменами.[75]
1 2 3 4 5 67
Золотой шакал А-ГРАММ-C-T-г-Т-C-GA-Т-TC-CA
Койот А-ГРАММ-C-Т-А-Т-C-GA-А-TC-GA
Волк Т-ГРАММ-C-T-А-Т-г-GA-Т-TC-CT
Собака Т-ГРАММ-г-Т-А-Т-г-GA-Т-TC-CА
Последовательности мДНК собаки и волка различаются только 0–12 заменами в пределах 261 пары оснований, тогда как собаки всегда отличались от койотов и шакалов не менее чем на 20 замен.[75][85] Этот вывод подразумевает, что собака произошла от волка и что имело место повторное обратное скрещивание,[85] или что собака, возможно, произошла от ныне вымершего вида псовых, ближайшим из ныне живущих родственников которых является современный волк.[86]
Проблема с маркером
Различные исследования ДНК могут дать противоречивые результаты из-за выбранных образцов, используемой технологии и предположений, сделанных исследователями.[87] Любой из группы генетические маркеры могут быть выбраны для использования в исследовании. Методы, используемые для извлекать, найти и сравнить генетические последовательности могут быть применены с использованием достижений технологий, что позволяет исследователям наблюдать более длинные длины пар оснований которые предоставляют больше данных для лучшего филогенетический разрешающая способность.[88] Филогенетические деревья составлен с использованием различных генетические маркеры дали противоречивые результаты об отношениях между волком, собакой и койотом. Одно исследование на основе SNP[89] (один мутация ), а другой на основе ядерный ген последовательности[90] (взято из ядро клетки ), показали, что собаки группируются с койотами и отдельно от волков. Другое исследование, основанное на SNPS, показало, что волки объединяются с койотами отдельно от собак.[91] Другие исследования, основанные на ряде маркеров, показывают более общепринятый результат объединения волков с собаками отдельно от койотов.[85][92] Эти результаты демонстрируют необходимость осторожности при интерпретации результатов, полученных с помощью генетических маркеров.[89]
Проблема с синхронизацией
Есть два ключевых предположения, которые сделаны для датировки времени дивергенции видов: время поколения и генетический скорость мутации на поколение. Предполагается, что промежуток времени между поколениями у волков составляет три года, исходя из существующего серого волка, и два года для собаки, исходя из существующей собаки.[83] В одном недавнем крупном исследовании предполагалось, что время генерации у собаки составляло 2 года еще 10000 лет назад, а затем предполагалось, что время генерации составляло 3 года (как у волка) до этого, чтобы рассчитать предлагаемое время расхождения между двумя .[9] В 2017 году ученый-исследователь волков Л. Дэвид Мех спросили, почему биологи-эволюционисты рассчитывали приблизительное время, когда собака отделяется от волка, используя время поколения волков в три года, когда опубликованные работы с использованием больших наборов данных демонстрируют цифру 4,2–4,7 года. Им было рекомендовано соответствующим образом пересчитать даты расхождения.[93]
Исследования ДНК проводятся, но «частота мутаций является основным источником неопределенности».[9] В 2005 году Линдблад-То секвенировал первый черновой вариант генома существующей собаки и рассчитал предполагаемую частоту мутаций 1x10.−8 мутации в поколении.[83] В 2015 году Скоглунд смог секвенировать первый черновой вариант генома 35000 YBP. Таймырский волк и использовал его радиоуглеродную дату для подтверждения предполагаемой скорости генетической мутации 0,4x10−8 мутации в поколении.[10] Разница составляет коэффициент времени 2,5, однако другое исследование показало, что, поскольку до сих пор был секвенирован только один образец плейстоценового волка, к результату следует относиться с осторожностью, а затем в этом исследовании представлены обе оценки для расчета предполагаемого времени расхождения между волк и собака.[11] Однако в 2016 г. частота мутаций 4800 YBP Собака Ньюгрейнджа соответствует таймырскому волку.[94]
Волкоподобные псовые
Волкоподобные псовые (подсемейство псовых Caninae ) представляют собой группу крупных плотоядных животных, которые генетически тесно связаны, поскольку их хромосомы № 78. В группу входит род Canis, Cuon и Ликаон. Члены являются собака (C. lupus familyis), серый волк (С. волчанка), койот (C. latrans), золотой шакал (C. aureus), Эфиопский волк (С. simensis), черноспинный шакал (C. mesomelas), полосатый шакал (C. adustus), волк (Cuon alpinus), и Африканская дикая собака (Lycaon pictus).[95][96][97] К новым предложенным членам относятся красный волк (Canis rufus), восточный волк (Canis Lycaon), и Африканский золотой волк (C. anthus). Поскольку они обладают 78 хромосомами, все представители рода Canis (койоты, волки, шакалы) кариологически неотличимы друг от друга, и от волка, и от африканской охотничьей собаки.[75]:p279[98] Члены Canis потенциально может скрещивание[86] и есть свидетельства того, что эфиопский волк скрещивался с собаками.[99] По мнению зоолога Реджинальд Покок, скрещивание волка с золотым шакалом.[100] Африканская охотничья собака крупная, очень подвижная, она, как известно, рассеивается на большие расстояния и редко встречается на большей части своего географического ареала.[101] затрудняя возможности для гибридизации. Исследование материнской митохондриальная ДНК шакала с черной спиной не смог найти никаких доказательств генотипы от наиболее вероятных партнеров - полосатого шакала или золотого шакала - что указывает на то, что самцы черноспинных шакалов не размножались с ними.[102] Поиск в научной литературе не смог найти свидетельств гибридизации редкого полосатого шакала.
Выравнивание последовательности ДНК волкоподобных псовых дало филогенетическое древо, в котором серый волк и собака были наиболее близкими родственниками, за которыми следовала тесная связь с койотом, золотым шакалом и эфиопским волком, и собака может гибридизоваться в дикой природе с эти три вида. Следующими ближайшими к этой группе являются волк и африканская дикая собака, у которых есть уникальные режущие мясо зубы, что позволяет предположить, что эта адаптация была позже утеряна другими членами.[83] Два африканских шакала показаны как самые базальный члены этой клады, что означает, что это дерево указывает на африканское происхождение клады.[83][103] Дерево иллюстрирует генотип-фенотип различие, где генотип заполнен ли организм наследственный информация и фенотип реально наблюдаемые свойства организма, такие как морфология, развитие, или поведение. По фенотипу волки (род Cuon) и африканская охотничья собака (род Ликаон) не классифицируются как представители рода Canis, но по генотипу они ближе к собакам, волкам и койотам, чем два рода Canis шакалы - полосатый шакал (C. adustus) и черноспинный шакал (C. mesomelas).
В 2015 году исследование последовательностей митохондриального генома и последовательностей ядерного генома африканских и евразийских псовых показало, что современные волкоподобные псовые колонизировали Африку из Евразии по крайней мере 5 раз на протяжении плиоцена и плейстоцена, что согласуется с данными окаменелостей, предполагающими, что большая часть Разнообразие африканских псовых явилось результатом иммиграции евразийских предков, что, вероятно, совпало с климатическими колебаниями плио-плейстоцена между засушливыми и влажными условиями.[84]
Филогенетическое дерево из дошедших до нас волкоподобных псовых,[а] с розовой штриховкой, представляющей вид Canis lupus.
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3,5 млн лет назад |
Смесь с вымершим неизвестным псовым
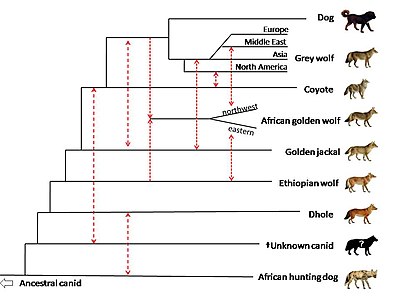
В 2018 г. секвенирование всего генома использовался для сравнения представителей рода Canis, вместе с волком (Cuon alpinus) и африканская охотничья собака (Lycaon pictus). Есть свидетельства потока генов между Африканские золотые волки, золотые шакалы, и серые волки. Исследование предполагает, что африканский золотой волк является потомком генетически смешанный псовый 72% серый волк и 28% Эфиопский волк происхождение, и что эфиопский волк когда-то имел более широкий ареал в Африке. Один африканский золотой волк из Египта Синайский полуостров показали высокую степень смешения с ближневосточными серыми волками и собаками, подчеркнув роль сухопутного моста между африканским и евразийским континентами в эволюции псовых. Есть свидетельства наличия потока генов между золотыми шакалами и ближневосточными волками, в меньшей степени у европейских и азиатских волков и меньше всего у североамериканских волков. Исследование предполагает, что предки золотых шакалов, обнаруженные у североамериканских волков, могли произойти до расхождения серых волков Евразии и Северной Америки. Исследование показывает, что общий предок койот а серый волк генетически смешанный с призрачное население вымершего неопознанного псового. Псовые генетически близки к волк и возник после того, как африканская охотничья собака отделилась от других видов псовых. В базальный Предполагается, что положение койота по сравнению с волком объясняется тем, что койот сохраняет большую часть митохондриального генома этого неизвестного псового.[104]
Геномное исследование китайских волков включало музейные образцы волков из южного Китая, которые были собраны между 1963 и 1988 годами. Волки, участвовавшие в исследовании, образовали три клады: североазиатские волки, в том числе волки из северного Китая и восточной России, Гималайские волки с Тибетского нагорья и уникальное население из южного Китая. Один экземпляр расположен к юго-востоку от г. Цзянси В провинции есть свидетельства того, что тибетские волки смешивались с другими волками в Китае. Один экземпляр из Чжэцзян Провинция в восточном Китае разделяла поток генов с волками из южного Китая, однако ее геном на 12-14 процентов был смешан с псовыми, которые могли быть волками или неизвестными псовыми, которые предшествовали генетической дивергенции волков. Считается, что популяция волков из южного Китая все еще существует в этом регионе.[118]
Две гаплогруппы волков
А гаплотип (гаплоидный генотип ) - это группа гены в организме, которые наследуются вместе от одного родителя.[119][120] А гаплогруппа представляет собой группу похожих гаплотипов, имеющих общего предка с однонуклеотидный полиморфизм (а мутация ).[121][122] Митохондриальная ДНК проходит по материнской линии, которая насчитывает тысячи лет.[121]
В 2010 году исследование сравнивало Последовательности ДНК это было 230 пар оснований в длину от митохондриальная контролирующая область 24 экземпляров древних волков из Западной Европы, датированных между 44,000–1200 YBP, с образцами современных серых волков. Большинство последовательностей можно было бы представить на филогенетическое дерево. Однако гаплотипы Гималайский волк и Индийский серый волк не могли, потому что у них было 8 мутаций, кроме других волков,[111] указывая на четкие линии происхождения, которые ранее были обнаружены в других исследованиях.[111][108][109] Исследование показало, что было 75 разных серых волков. мДНК гаплотипы, которые включают 23 в Европе, 30 в Азии, 18 в Северной Америке, по 3 в Европе и Азии и по 1 в Европе и Северной Америке.[111] Эти гаплотипы можно разделить на две гаплогруппы.[113][111] которые были отделены друг от друга 5 мутации. Гаплогруппа 1 сформировала монофилетический клады (что указывает на то, что все они несли одну и ту же мутацию, унаследованную от одного предка женского пола). Все другие гаплотипы были базальными в дереве, и они образовали 2–3 меньшие клады, которые были отнесены к гаплогруппе 2, которая не была монофилетической.[111][123]
Гаплогруппы 1 и 2 можно было найти разбросанными по Евразии, но только гаплогруппу 1 можно было найти в Северной Америке. Образцы древних волков из Западной Европы отличались от современных волков от 1 до 10 мутаций, и все они принадлежали к гаплогруппе 2, что указывает на преобладание гаплогруппы 2 в этом регионе на протяжении более 40 000 лет до и после Последний ледниковый максимум. Сравнение текущей и прошлой частот показало, что в Европе гаплогруппа 2 уступила по численности гаплогруппа 1 за последние несколько тысяч лет.[113] но в Северной Америке гаплогруппа 2 вымерла и была заменена гаплогруппой 1 после Последний ледниковый максимум.[111][123] Доступ в Северную Америку был доступен между 20 000–11 000 лет назад после Висконсинское оледенение отступили, но до Берингов мост был затоплен морем.[124] Следовательно, гаплогруппа 1 смогла проникнуть в Северную Америку в этот период.
Стабильный изотопный анализ Исследование, проведенное на кости образца, позволяет исследователям делать выводы о диете и, следовательно, об экологии вымерших популяций волков. Этот анализ предполагает, что плейстоценовые волки из гаплогруппы 2, обнаруженные в Берингия а Бельгия охотилась в основном на мегафауну плейстоцена,[64][111][110] который стал редкостью в начале голоцена 12 000 лет назад.[111][125] Одно исследование показало, что берингийский волк базальный всем другим серым волкам, кроме сохранившихся Индийский серый волк и существующие Гималайский волк.[110] Было обнаружено, что плейстоценовые евразийские волки морфологически и генетически сопоставимы с плейстоценовыми восточно-азиатскими волками.Берингийские волки,[126] с некоторыми из древних европейских и берингийских волков, имеющих общий гаплотип (a17),[111][110] что делает вероятным экологическое сходство.[111] Два древних волка с Украины датированы примерно 30 000 юношей и 33 000 юношей. «Алтайская собака» имел ту же последовательность, что и шесть берингийских волков, а еще один из Чешской Республики, датированный 44 000 YBP, имел ту же последовательность, что и два берингийских волка.[110]
Было высказано предположение, что плейстоценовые волки в Северной Евразии и Северной Америке представляли собой непрерывный и почти панмиктический популяция, которая была генетически и, вероятно, также экологически отлична от волков, обитающих в этой области сегодня.[111][127] Специализированные плейстоценовые волки не способствовали генетическому разнообразию современных волков, а современные популяции волков по всей Голарктике, вероятно, являются потомками волков из популяций, пришедших из более южных убежищ.[127] Сохранившиеся волки гаплогруппы 2 можно найти в Италии, на Балканах и в Карпатах, но редко где-либо еще в Европе. В Азии только четыре гаплотипа были идентифицированы как принадлежащие к этой гаплогруппе, и два из них встречаются на Ближнем Востоке.[128] Гаплогруппа 2 не вымерла в Европе, и если до Последний ледниковый максимум гаплогруппа 2 была связана исключительно с волчьим экоморфом, специализирующимся на добыче мегафауны, это означало бы, что в Европе он был способен адаптироваться к изменению добычи.[111]
В 2013 году секвенирование митохондриальной ДНК древних волкоподобных псовых выявило еще одну отдельную линию из 3 гаплотипов (образующих гаплогруппу), которая была обнаружена в 3 образцах позднего плейстоцена из Бельгии; однако у современных волков он не обнаружен.[66][128] Одним из них был "Гойет дог".[66]
Особое мнение

В 2016 году было проведено исследование из-за опасений, что предыдущие исследования мДНК могли быть проведены с недостаточным генетическим разрешением или ограниченным географическим охватом и не включали достаточное количество образцов из России, Китая и Ближнего Востока. В исследовании сравнивали 582 базовая пара последовательность из митохондриальная контролирующая область что дало вдвое более высокое филогенетическое разрешение исследования 2010 года.[111] В исследовании сравнивались последовательности как современных волков, так и древних образцов волков, включая образцы из отдаленных районов Северной Америки, России и Китая. Исследование включало Таймырские волки, то Гойет "собака", то Алтайская "собака", Берингийских волков и др. Древние образцы.[105]
Исследование обнаружило 114 различных гаплотипов волков среди 314 последовательностей, причем новые гаплотипы были обнаружены в Сибири и Китае. Филогенетическое древо разделилось на 19 клад, которые включали как современных, так и древних волков, что показало, что большинство базальных клад включают Индийский серый волк и Гималайский волк, с субкладом волков из Китая и Монголии, попадающим в кладу гималайских волков. Два самых базальных гаплотипа Северной Америки включали Мексиканский волк и Волк острова Ванкувер однако волк с острова Ванкувер показал тот же гаплотип, что и собака, что указывает на примесь,[105] с родословной собак от всех существующих североамериканских подвидов.[11] В Европе два генетически различных гаплотипа образуют Иберийский волк и отдельно Итальянский волк это было расположено рядом с древними волками. В Гренландские волки все принадлежали к одному гаплотипу, который ранее был обнаружен среди североамериканских волков и который указывает на их происхождение из Северной Америки. Восточный волк был подтвержден как гибрид койота и волка. Волки встречаются в регионах Чукотский полуостров, северокорейская граница, Амурская область и Хакасия показали наибольшее генетическое разнообразие и были тесно связаны со всеми другими волками, обитающими в Голарктике. Один древний гаплотип, который был обнаружен на Аляске (Восточная Берингия 28 000 лет назад) и России (Медвежья пещера "Медведь", Печора область, Северная Урал 18000 YBP) был разделен с некоторыми современными волками, найденными в Китае и Монголии.[105]
Предыдущее открытие двух гаплогрупп волков[111] не было четко очерчено в этом исследовании, но оно согласилось с тем, что генетическое разнообразие прошлых волков было утрачено в начале голоцена на Аляске, в Сибири и в Европе с ограниченным совпадением с современными волками. Для древних волков Северной Америки вместо модели вымирания / замещения, предложенной в предыдущем исследовании,[110] это исследование обнаружило существенные доказательства ограниченности популяции в Северной Америке, в которой древнее разнообразие волков было почти утрачено примерно в начале голоцена (без дальнейших разработок в исследовании). В Евразии потерю древних родословных нельзя было просто объяснить и, похоже, происходило медленно с неясными причинами.[105]
В Америку и Японию

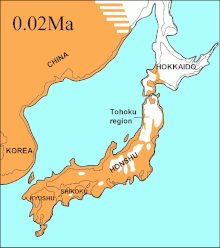
В 2016 году исследование основывалось на результатах другого крупного исследования.[66] и проанализировали последовательности 12 генов, расположенных на тяжелая прядь митохондриального генома вымерших и современных С. волчанка. Из исследования исключены последовательности расходящихся Гималайский волк и Индийский серый волк. Древние образцы были радиоуглеродный датированный и стратаграфически датированы, и вместе с последовательностями сформировали основанное на времени филогенетическое дерево. На основе этого дерева исследователи смогли установить самого недавнего общего предка для всех остальных С. волчанка особей - современных и вымерших - было 80 000 YBP, и эта дата совпадала с данными более раннего исследования.[66][115] Исследование не могло найти никаких доказательств ограниченности популяции волков еще несколько тысяч лет назад.[115]
Филогенетическое древо показало полифилия Что касается американских волков, то мексиканский волк отличался от других североамериканских волков, и эти другие североамериканские волки образовали две тесно связанные клады. Сценарий, соответствующий филогенетическим данным, данным о ледниковом покрове и уровне моря, заключался в том, что во время ледникового периода, когда уровень моря был самым низким, произошла единственная волна колонизации волков в Северной Америке, которая началась с открытия Берингов мост 70,000 YBP и закрытие в течение Поздний ледниковый максимум Юконского коридора, проходящего через разделение Ледяной щит Лаурентиды и Кордильерский ледяной покров 23000 лир. Мексиканские волки были частью единой волны и либо отделились от других волков до того, как попали в Северную Америку, либо однажды попали в Северную Америку из-за изменения окружающей среды.
Поскольку волки были в летописи окаменелостей Северной Америки, но современные волки могли проследить свою родословную только 80 000 лет назад, гаплотипы волков, которые уже были в Северной Америке, были заменены этими захватчиками либо в результате конкурентного вытеснения, либо путем смешения.[115] Замена основной популяции волков в Северной Америке более поздней подтвердила результаты более ранних исследований.[111][123][110][115] Возможно, до закрытия ледяных щитов существовала панмиктическая популяция волков с потоком генов, охватывающим Евразию и Северную Америку.[111][127][115] Когда листы закрылись, южные волки оказались изолированными, а к северу от листов остались только Берингийский волк существовал. Сухопутный мост был затоплен морем на 10 000 YBP, листы отступили на 12 000–6 000 YBP, берингийский волк вымер, а южные волки расширились, чтобы повторно заселить остальную часть Северной Америки. Все североамериканские волки произошли от тех, которые когда-то были изолированы к югу от ледяных щитов. Однако большая часть их разнообразия позже была утрачена в течение двадцатого века.[115]
Исследования с использованием митохондриальная ДНК указали, что волки прибрежных юго-востока Аляски генетически отличаются от внутренних серых волков, что отражает закономерность, наблюдаемую и в других таксонах. Они демонстрируют филогенетические отношения с искорененными волками с юга (Оклахома), указывая на то, что эти волки являются последними останками некогда широко распространенной группы, которая была в значительной степени истреблена в течение прошлого века, и что волки Северной Северной Америки первоначально распространились с южные убежища ниже Висконсинское оледенение после того, как лед растаял в конце Последний ледниковый максимум.[129][130][131] А полногеномный Исследование ДНК показало, что все североамериканские волки были монофилетический и поэтому являются потомками общего предка.[132]
В тот же период Соевый пролив между Хоккайдо и Остров Сахалин просохло 75000 лет, и было высказано предположение, что вымершие Эзо волк (C. l. Хаттай) прибыл на Хоккайдо с Сахалина.[133][115][134] Однако последовательности показали, что он прибыл на Хоккайдо менее чем 10 000 YBP. Волк Эзо был тесно связан с одной из североамериканских клад,[133][115][135] но отличается от более южного Японский волк (C. l. ходофилакс), которая была базальна для современных волков.[133][115] Японский волк населял острова Кюсю, Сикоку и Хонсю.[136][137] но не остров Хоккайдо.[137] Это указывает на то, что его предок мог мигрировать с азиатского континента через Корейский полуостров в Японию.[133][137] Уровни моря в прошлом Корейский пролив вместе с хронометражем японских последовательностей волков указывает на то, что он прибыл на южные острова менее чем 20 000 YBP.[115]
Собака была очень успешным завоевателем Северной Америки и приобрела широкое распространение. экологическая ниша к раннему – среднему голоцену. Между собакой и волком не было никакого совпадения ниши по сравнению с собакой и другими североамериканскими псовыми.К позднему голоцену площадь ниши собаки была меньше, чем ожидали исследователи, что указывает на то, что она была ограничена биотические факторы. Эти регионы включают северо-восток и северо-запад Соединенных Штатов, которые коррелируют с наибольшей плотностью раннего заселения человека, что указывает на то, что собака «перешла» из волчьей ниши в человеческую нишу, и объясняет, почему площадь ниши собаки не была такой большой, как ожидал. Разделение между собакой и волком может отражать быстрые темпы приручения,[138] включая возможность второго случая одомашнивания в Северной Америке.[139][138] Стаи волков и охотники-собиратели охотятся на подобную добычу аналогичным образом в рамках аналогичной социальной структуры группы, которая, возможно, облегчила приручение волков.[140][52]
К середине 20 века волк был истреблен в южной части своего исторического ареала в Северной Америке. Исследование мДНК 34 останков волков из Северной Америки, датированных периодом между 1856 и 1915 годами, показало, что их генетическое разнообразие вдвое больше, чем у современных волков в этих регионах, а две трети идентифицированных гаплотипов были уникальными. Эти результаты показывают, что историческая популяция в несколько сотен тысяч волков когда-то существовала в Мексике и на западе США.[141][113]
Берингия
Серые волки пострадали от узкое место населения (уменьшение) примерно на 25000 YBP во время последнего ледникового максимума. За этим последовала единственная популяция современных волков, выросшая из Берингийский приют заново заселить прежний ареал волков, заменив оставшиеся позднеплейстоценовые популяции волков по всей Евразии и Северной Америке.[114][116][117] Эта исходная популяция, вероятно, не породила собак, а была смешана с собаками, которые позволили им получить гены окраски шерсти, которые также связаны с иммунитетом, и предоставили собакам гены, которые позволили им адаптироваться к высокогорной среде (например, Тибет). Это предполагает, что генетическая дивергенция европейских и восточноазиатских собак может быть основана на смешении с разными субпопуляциями волков.[117]
О древних волках, существовавших до появления узкого места, мало доступной генетической информации. Однако исследования показывают, что одна или несколько из этих древних популяций являются более непосредственными предками собак, чем современные волки, и, возможно, они были более склонны к приручению первыми людьми, вторгшимися в Евразию.[117]
Расхождение с койотом
В 1993 году исследование показало, что волки в Северной Америке обладают чертами черепа, более похожими на койотов, чем волки из Евразии.[41] В 2016 г. полногеномный Исследование ДНК на основе сделанных предположений показало, что все североамериканские волки и койоты произошли от общего предка менее 6000–117000 лет назад. Исследование также показало, что все волки Северной Америки имеют значительное количество предков койотов, а все койоты - некоторую степень предков волков, и что красный волк и восточный волк очень смешанный с разными пропорциями предков серого волка и койота. Одно испытание показало, что время расхождения волков и койотов составляло 51000 лет до настоящего времени, что соответствовало другим исследованиям, показывающим, что дошедший до нас волк появился примерно в это время. Другое испытание показало, что красный волк отделился от койота за 55 000–117 000 лет до настоящего времени, а волк из района Великих озер - за 32 000 лет до настоящего времени. Другие тесты и моделирование показали различные диапазоны дивергенции, и был сделан вывод о диапазоне от 6000 до 117000 лет до настоящего времени.[132][142] Эта находка противоречит летописи окаменелостей, указывающей на наличие похожего на койота экземпляра, датируемого 1 миллионом лет назад.[15]
Современный серый волк распространился из Берингии 25 000 лет назад. Это означает, что если койот и красный волк произошли от этого вторжения, их истории датируются всего десятками тысяч, а не сотнями тысяч лет назад, что согласуется с другими исследованиями.[117]
Домашняя собака
Домашняя собака (Обыкновенная волчанка) - наиболее широко распространенное крупное плотоядное животное.[66][11][143] За последний миллион лет существовало множество волкоподобных форм, но их оборот был высоким, и современные волки не являются прямыми предками собак.[66][9][11][141] Хотя исследования показали, что собаки и волки были генетически очень близкими родственниками,[85][86][95] позже филогенетический анализ решительно поддержал гипотезу о том, что собаки и волки взаимно монофильный таксоны это две сестры клады.[85][9][144] Это говорит о том, что ни одна из современных популяций волков не связана с волками, которые были первыми приручены, и поэтому волк-предок собак считается вымершим.[9][144] Недавние митохондриальная ДНК Анализ древних и современных образцов серого волка подтверждает картину сокращения популяции и обновления.[66][111][110] Альтернативное предложение состоит в том, что во время экологических потрясений позднего плейстоцена все оставшиеся члены сокращающегося рода присоединились к людям.[145][67]
В 2016 году исследование впервые исследовало подразделения популяций, демографию и взаимоотношения серых волков на основе их полногеномные последовательности. Исследование показало, что собака была дивергентным подвидом серого волка и произошла от ныне вымершего призрачное население волков позднего плейстоцена,[66][9][11] и собака и динго не отдельные виды.[11] Общегеномный филогенетическое дерево указал генетическая дивергенция между волками Нового и Старого Света, за которым последовало расхождение между собаками и волками Старого Света 27,000YBP[10][11] - 29000 лыб.[11] Собака образует сестринский таксон с евразийскими серыми волками, но не с североамериканскими волками. После расхождения с волками Старого Света у собаки была значительная предковая родословная, прежде чем она разделилась на отдельные родословные, которые почти так же отличаются друг от друга, как и от волков.[11] Исследование показало, что предыдущие датировки, основанные на расхождении между волками и койотами миллион лет назад с использованием окаменелостей того, что выглядело похожими на койотов, могут не отражать происхождение современных форм.[84][9][10][11]
| Дивергенция серого волка и время | ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| Полный геном филогенетический дерево - сохранившиеся популяции серых волков,[11] с временами расхождения, рассчитанными с использованием предполагаемой скорости мутации Линдблад-То (1x10−8)[83] или [Скоглунд] (0,4x10−8).[10] Собака является генетически дивергентным подвидом серого волка со значительным расхождением между собачьими родословными, столь же отличными друг от друга, как волки от собак, что может отражать большее количество примесей и предков волков, сохраненных в их геноме.[11] |
Исследование показало, что Мексиканский волк также была дивергентной формой серого волка, что позволяет предположить, что это могло быть частью раннего вторжения в Северную Америку.[11][141] Было обнаружено, что тибетский волк является наиболее дивергентным из волков Старого Света, пострадал от исторической проблемы с популяцией и только недавно повторно заселил Тибетское плато. Оледенение могло стать причиной потери среды обитания, генетической изоляции, а затем локальной адаптации.[11]
Исследование показало, что было много генетическая примесь между домашними собаками и волками, причем до 25% генома волков Старого Света демонстрируют признаки собачьей родословной, возможно, в результате поток генов из собак в волков, которые были предками всех современных волков. Были доказательства значительного потока генов между европейскими волками и израильскими волками с басенджи и боксер, что предполагает смешение линий предков этих пород и популяций волков.[9][11] Для волков низинной Азии: волки из Средней и Восточной России и все волки из низинной Китая имели значительный генетический поток от местных собак Китая, Тибетский мастиф и динго. Для высокогорных азиатских волков: тибетские волки не показали значительной примеси с собаками; Однако Цинхай у волков был генетический поток от динго, а у одного из них был геном от китайских собак. У волков Нового Света не было обнаружено какого-либо потока генов у собак-боксеров, динго или коренных китайских собак, но были признаки потока генов между мексиканским волком и африканским басенджи.[11] Все виды в пределах рода Canis, то волкоподобные псовые, филогенетически тесно связаны с 78 хромосомы и потенциально может скрещивание.[86] Было указание на поток генов в золотой шакал от предков популяции до всех волков и собак (11,3–13,6%) и гораздо более низкие показатели (до 2,8%) от существующих популяций волков.[9][11]
Данные показали, что у всех волков были сходные популяционные траектории, за которыми последовало сокращение популяции, которое совпало с распространением современных людей по всему миру и их технологий для отлова крупной дичи.[11][146] Плотоядные животные позднего плейстоцена вели бы социальную жизнь в больших прайдах, кланах и стаях, чтобы охотиться на более крупную дичь, доступную в то время, и эти более крупные группы были бы более заметными целями для преследователей-людей.[146] Крупные собаки, сопровождающие людей, могли ускорить сокращение численности хищников, соревнующихся за дичь.[11][147] поэтому люди расширились по Евразии, встретили волков, одомашнили одних и, возможно, вызвали упадок других.[11]
Исследование пришло к выводу, что добавка помешала сделать выводы о месте приручения собак. Прошлые исследования, основанные на SNP, общегеномном сходстве с китайскими волками и ниже. нарушение равновесия по сцеплению может отражать региональное смешение собак с волками и поток генов между популяциями собак, при этом разные породы собак, возможно, сохраняют больше предков волков в своем геноме. Исследование показало, что анализ древней ДНК может быть лучшим подходом.[11]
В том же году исследование показало, что было всего 11 фиксированных генов, которые показывали различия между волками и собаками. Считается, что эти гены влияют на приручение и способность обрабатывать эмоции.[148] Другое исследование представило список всех гаплотипов мДНК серых волков и собак, объединенных в одно филогенетическое дерево.[149]
В 2018 году в исследовании сравнивали последовательности 61000 Однонуклеотидные полиморфизмы (мутации ) взяты из генома серых волков. Исследование показало, что отдельные волки собачьей / волчьей родословной существуют в большинстве популяций волков Евразии, но в меньшей степени в Северной Америке. Гибридизация происходила в разных временных масштабах и не была недавним событием. Низкоуровневая гибридизация не снизила различимость волков.[150]
Динго

Динго (Canis lupus dingo ) относится к собаке, найденной в Австралия. Динго - это дивергентный подвид серого волка, а не отдельный вид,[11] и считается генетически базальный член клады домашних собак.[11][84][9] Генетические данные показывают, что динго произошел от домашних собак из Восточной Азии и был завезен через архипелаг Юго-Восточной Азии в Австралию.[152][153] с общим происхождением австралийского динго и Поющая собака Новой Гвинеи.[153][154]
Таймырский волк

В мае 2015 года было проведено исследование на частичной реберной кости образца волка (названного "Таймыр-1") найден у реки Большая Балахная в р. Полуостров Таймыр Арктики Северная Азия, что было AMS радиоуглеродный датированный до 34 900YBP. Образец предоставил первый черновик геном из ядро клетки для плейстоцена плотоядное животное, и последовательность была идентифицирована как принадлежащая Canis lupus.[10]
Используя радиоуглеродную дату особи Таймыра-1, последовательность его генома и последовательность генома современного волка, можно сделать прямую оценку частоты мутаций в масштабе всего генома у собак / волков для расчета времени расхождения. Данные показали, что ранее неизвестная линия происхождения Таймыр-1 была популяцией волков, отдельной от современных волков и собак, и указали на то, что генотип Таймыр-1, серые волки и собаки произошли от ныне вымершего общего предка.[10][73][155] перед пиком Последнего ледникового максимума, 27 000–40 000 лет назад. Разделение собаки и волка не обязательно должно совпадать с селекцией людей.[10][156] Такое раннее расхождение согласуется с несколькими палеонтологическими отчетами о собачьих псовых, датируемыми до 36000 лет.YBP, а также свидетельства того, что домашние собаки, скорее всего, сопровождали первых колонизаторов в Америку.[10]
Сравнение с линией серых волков показало, что Таймыр-1 был базальным по отношению к серым волкам с Ближнего Востока, Китая, Европы и Северной Америки, но разделил значительную часть истории с современными серыми волками после их отделения от койота. Это означает, что предки большинства популяций серых волков сегодня происходят от предков, которые жили менее 35000 лет назад, но до затопления Берингов мост с последующей изоляцией евразийских и североамериканских волков.[10]
Сравнение родословной линии Таймыр-1 с родословной собак показало, что некоторые современные породы собак имеют более тесную связь либо с серым волком, либо с Таймыр-1 из-за примесь. В Саарлосская волчья собака показал большую ассоциацию с серым волком, что согласуется с документально подтвержденным историческим скрещиванием с серыми волками в этой породе. Таймыр-1 имеет больше аллелей (экспрессий генов) с теми породами, которые связаны с высокими широтами: Сибирский хаски и Гренландская собака[10][155] которые также связаны с арктическими популяциями людей, и в меньшей степени с Шар-пей и Финский шпиц. График примеси гренландской собаки показывает, что наилучшим образом подходят 3,5% общего материала, хотя соотношение предков в диапазоне от 1,4% до 27,3% согласуется с данными. Это указывает на смешение популяции Таймыра-1 с исконной популяцией собак этих четырех высокоширотных пород. Эти результаты можно объяснить либо очень ранним появлением собак в северной Евразии, либо генетическим наследием Таймыра-1, сохраняющимся в популяциях северных волков до появления собак в высоких широтах. Эта интрогрессия могли предоставить ранним собакам, живущим в высоких широтах, фенотипические вариации, полезные для адаптации к новой и сложной среде. Это также указывает на то, что предки современных пород собак происходят из более чем одного региона.[10]
Попытка изучить примесь Таймыра-1 и серых волков дала ненадежные результаты.[10]
Поскольку таймырский волк внес свой вклад в генетический состав арктических пород, более позднее исследование показало, что потомки таймырского волка дожили до тех пор, пока собаки не были одомашнены в Европе и не прибыли в высокие широты, где они смешались с местными волками. современные арктические породы. Судя по наиболее широко признанным старейшим зооархеологическим останкам собак, домашние собаки, скорее всего, прибыли в высокие широты в течение последних 15000 лет. Скорости мутаций откалиброваны как по таймырскому волку, так и по Собака Ньюгрейнджа геномы предполагают, что современные популяции волков и собак отличались от общего предка между 20 000–60 000YBP. Это указывает на то, что либо собаки были приручены намного раньше, чем их первое появление в археологических записях, либо они прибыли в Арктику рано, либо и то, и другое.[157]
Находка второго экземпляра волка из той же местности («Таймры-2») И датируется 42 000YBP также была секвенирована, но дала только митохондриальную ДНК.[158]
Canis variabilis
В 2015 году в исследовании изучались митохондриальная контролирующая область последовательности 13 останков древних псовых и одного современного волка из пяти памятников арктической северо-восточной Сибири. Четырнадцать псовых показали девять митохондриальных гаплотипы, три из которых были зарегистрированы, а о других не сообщалось ранее. Филогенетическое дерево, полученное из последовательностей, показало, что четыре сибирских псовых датируются 28000 лет назад и один Canis c.f. variabilis от 360 000 YBP сильно расходились. Гаплотип, обозначенный как S805 (28000 YBP) из Река Яна была одна мутация от другого гаплотипа S902 (8000 YBP), который представляет кладу A современных линий волков и домашних собак. Тесно связан с этим гаплотипом тот, который был обнаружен у недавно вымерших Японский волк. Несколько древних гаплотипов были ориентированы на S805, в том числе Canis c.f. variabilis (360 000 YBP), Бельгия (36 000 YBP - «собака Гойет»), Бельгия (30 000 YBP) и Констэки, Россия (22 000 YBP). Учитывая положение гаплотипа S805 на филогенетическом дереве, он потенциально может представлять прямую связь с прародитель (в том числе Canis c.f. variabilis) до домашних собак и современных волков. Серый волк считается предком домашней собаки, однако его родство с С. variabilis, и генетический вклад С. variabilis к собаке, является предметом споров.[159]
В Жохов остров (8700 YBP) и Aachim (1700 YBP) гаплотипы собак попали в кладу домашних собак, кластер с S805, а также разделяют свои гаплотипы с - или находятся на расстоянии одной мутации от - Тибетский волк (C. l. Filchneri) и недавно вымерший японский волк (C. l. ходофилакс). Это может указывать на то, что эти псовые сохранили генетический признак смешения с региональными популяциями волков. Другой гаплотип, обозначенный как S504 (47,000 YBP) от Дуванный Яр появился на филогенетическом древе как не связанный с волками (как древними, так и современными), но являющийся предком собак, и может представлять собой генетический источник местных собак.[159]
Авторы пришли к выводу, что строение современной собаки Генофонд был получен от древних сибирских волков и, возможно, от Canis c.f. variabilis.[159][160]
Восхождение к доминирующему хищнику
В 2015 году было проведено исследование палеоэкологии крупных плотоядных животных в Мамонт степной вовремя Поздний плейстоцен с помощью анализа стабильных изотопов их ископаемых коллаген реконструировать свой рацион. По результатам тестирования в Бельгии, около 40 000 YBP Пещерные гиены охотятся на мамонтов, шерстистых носорогов, лошадей и северных оленей, пещерные львы брать оленей и молодняк пещерные медведи. Волки, похоже, проиграли пещерные гиены и ограничили свой рацион серной, гигантским оленем и благородным оленем. Однако после последнего ледникового максимума около 14000 лет назад волки получили доступ ко всем видам добычи, пещерный лев был ограничен оленями, а пещерная гиена вымерла.[161][162][163] Данные показывают, что исчезновение пещерной гиены позволило волку стать доминирующим хищником, а не пещерным львом, как раз перед исчезновением пещерного льва.[163] Другое исследование показало, что волк процветал по сравнению с пещерной гиеной, когда был больший снежный покров.[164]
Различия в популяциях волков
Серый волк Canis lupus это высокоадаптивный вид, способный существовать в различных средах и широко распространенный в Голарктический. Исследования современных серых волков выявили отдельные субпопуляции, которые живут в непосредственной близости друг от друга.[2][3] Эти различия в субпопуляциях тесно связаны с различиями в среде обитания - осадками, температурой, растительностью и специализацией добычи - которые влияют на черепно-зубную пластичность.[4][5][6][7] Археологические и палеонтологические записи показывают их постоянное присутствие в течение как минимум последних 300 000 лет.[8] Это постоянное присутствие волка контрастирует с геномный исследования, которые показывают, что все современные волки и собаки происходят от общей предковой популяции волков.[9][10][11] Который существовал всего 20 000 лет назад.[9] Эти исследования показывают, что узкое место населения за ним последовало быстрое излучение от наследственного населения во время или сразу после Последний ледниковый максимум. Это означает, что первоначальные популяции волков уступили место новому типу волков, который их заменил.[115] Однако географическое происхождение этого излучения неизвестно.
Помимо приручения, люди причинили вред волку, ограничив его среду обитания путем преследования. Это привело к резкому снижению численности его населения за последние два столетия.[165][166] Сокращение ареалов его обитания, совпадающих с местами обитания близких родственников, таких как собаки и койоты, привело к многочисленным случаям гибридизации.[167][168] Эти события, в дополнение к недавним изменениям (вымиранию и повторному заселению другими генотипами), привели к распаду филогеографический история волка сложная.[105]
Экотипы
An экотип вариант, в котором фенотипический различий слишком мало или слишком тонко, чтобы их можно было причислить к подвидам. Они могут возникать в одном и том же географическом регионе, где различные среды обитания, такие как луг, лес, болото и песчаные дюны, обеспечивают экологические ниши. Если сходные экологические условия встречаются в удаленных друг от друга местах, возможно возникновение аналогичного экотипа. Это отличается от подвида, который может существовать в разных средах обитания. У животных экотипы можно рассматривать как микроподвиды, которые своими различными характеристиками обязаны воздействию очень местной окружающей среды.[169] Экотипы не имеют таксономический ранг.
Серые волки имеют широкое естественное распространение в Голарктический это включает в себя множество различных сред обитания, которые могут варьироваться от арктических до густых лесов, открытых степей и пустынь. Генетические различия между разными популяциями серых волков тесно связаны с типом среды обитания, в которой они живут.[7] Различия в генетических маркерах среди скандинавской популяции волков возникли всего за десять с лишним лет из-за их небольшого размера.[7][170] что указывает на то, что эти различия не зависят от длительного времени, проведенного в изоляции, и что более крупные популяционные структуры могут развиться всего за несколько тысяч лет.[7] Эти различия также могут включать цвет и плотность меха, а также размер тела.[7][171][2] Различия также могут включать поведение, поскольку прибрежные волки едят рыбу.[7][171] и мигрируют тундровые волки.[7][2] Эти различия наблюдались между двумя популяциями волков, живущими в непосредственной близости. Было показано, что горные волки не скрещиваются с соседними прибрежными волками, а французские и швейцарские Альпы были заселены волками из гор соседней Италии.[7][172] и из далеких гор Хорватии[7][173] а не из более близких низменностей, что указывает на то, что расстояние не является движущей силой в различиях между двумя экоморфы.[7]
В 2013 году генетическое исследование показало, что популяция волков в Европе была разделена по оси север-юг и образовала пять основных групп. Были идентифицированы три кластера, занимающие южную и центральную Европу в Италии, Карпатах и Динарско-Балканах. Еще два кластера были идентифицированы, занимая северо-центральную Европу и украинские степи. Итальянский волк состоял из изолированной популяции с низким генетическим разнообразием. Волки из Хорватии, Болгарии и Греции сформировали Динарско-Балканский кластер. Волки из Финляндии, Латвии, Белоруссии, Польши и России сформировали кластер северно-центральной Европы, а волки из Карпат - смесь волков из северо-центрального кластера и Динарско-балканского кластера. Волки из Карпат были больше похожи на волков из украинской степи, чем на волков из северной и центральной Европы. Эти скопления могли быть результатом распространения ледниковых рефугиумов, адаптации к местной среде, фрагментации ландшафта и убийства волков в некоторых районах людьми.[174]
В 2016 году в двух исследованиях сравнивались последовательности 42000 однонуклеотидный полиморфизм у североамериканских серых волков и обнаружил, что они образовали шесть экотипов. Эти шесть экотипов волков были названы Западный лес, Северный лес, Арктика, Высокий Арктический, Баффиновый и Британская Колумбия. Исследования показали, что осадки и диапазон средних суточных температур оказали наибольшее влияние на изменение последовательности.[3][175] Эти результаты согласуются с предыдущими выводами о том, что осадки влияют на морфологию,[176] и эта растительность[4] и тип среды обитания[171][177] повлияли на различия волков. Одно из этих исследований показало, что вариация в 11 ключевых генах влияет на зрение, обоняние, слух, цвет шерсти, метаболизм и иммунитет волков. В ходе исследования было выявлено 1040 генов, которые потенциально подвергаются отбору из-за изменчивости среды обитания, и, следовательно, имеются доказательства локальной адаптации экотипов волков на молекулярном уровне. Наиболее примечательным был положительный отбор генов, влияющих на зрение, цвет шерсти, метаболизм и иммунитет у экотипов Арктики и Высокого Арктика, а также то, что экотип Британской Колумбии обладает уникальным набором адаптаций.[175] Локальная адаптация экотипа волка, скорее всего, отражает предпочтение волка оставаться в той среде обитания, в которой он родился.[3]
Экологические факторы, включая тип среды обитания, климат, специализацию добычи и конкуренцию хищников, будут сильно влиять на серого волка. генетическая структура популяции и кранио-дентальный пластичность.[125][73][7][2][178][179][4][5][6] Во время последнего ледникового максимума был большой волк. генетическое разнообразие чем сегодня,[9][66] а в популяции серых волков плейстоцена различия между местными средами способствовали появлению ряда экотипов волков, которые были генетически, морфологически и экологически отличны друг от друга.[73]
Плейстоценовые волки
Старейший Canis останки, найденные в Европе, были из Франции и датированы 3,1 миллионами лет назад,[180] с последующим Canis cf. этруск (где ср. в переводе с латыни означает совещание, неуверенный) из Италии датируется 2,2 млн юаней.[181] С. волчанка впервые появился в Италии во время Морской изотопный этап 9[182] (337 000 лыб). В Великобритании это был единственный вид псовых, присутствовавший в MIS 7 (243 000 YBP), с самым старым видом из Пещера Понтневидд в северном Уэльсе.[183] Во время ледникового периода Британию отделяла от Европы только Река канал.
Исследование плейстоцена С. волчанка в Великобритании в разные периоды времени обнаружили, что его способность дробить, резать мясо и есть кости подчеркивает его черепно-зубную пластичность. Эти реакции на изменения в питании показали изменения в диете в масштабах всего вида, а не только у местных экоморфов, в ответ на климатические и экологические переменные. Выживание С. волчанка во время плейстоцена во многом можно объяснить его пластической черепно-дентальной морфологией.[6]




| Время YBP | Переменные |
|---|---|
| 243,000 MIS 7 | Палеосреда представляла собой открытые луга с летними температурами от 16 ° C до 23 ° C и зимними температурами от -7 ° C до -6 ° C, где преобладали степные мамонты и лошади. Среди конкурентов были лев, бурый медведь и, реже, пятнистая гиена. Волки из MIS 7 были немного меньше по размеру тела, чем волки из MIS 5 и те, что встречаются сегодня в Швеции. Эти волки уступили более крупным конкурентам, что привело к более всеядной диете с повышенной способностью к раздавливанию в открытой среде, которая поддерживала больше видов добычи и больше немясной пищи, чем в период MIS 5. У них были более мелкие и узкие челюсти, чем у волков MIS 5 и тех, которые встречаются сегодня в Швеции, что указывало на то, что они могли ловить только мелкую и среднюю добычу. У них более низкий процент поломки зубов, чем у волков MIS-3. Однако у них был самый высокий процент умеренно изношенных зубов.[6] |
| 82000 MIS 5A | Палеосреда была холодной, открытой тундрой с летними температурами от 7 ° C до 11 ° C и зимними температурами от -10 ° C до -30 ° C, где преобладали олени и бизоны. Крупный бурый медведь был главным хищником, в то время гиены не было. Волки из MIS 5 были больше по размеру тела, чем те, что встречаются сегодня в Швеции. Эти волки страдали от сурового климата, низкой доступности добычи и диетического стресса, что привело к более плотоядной диете с увеличением количества замороженных туш и потребления костей. У них были сильные челюсти и наивысшая способность резать плоть по сравнению с другими волками, с более мелкими челюстями, чем у современных волков, но более широкими и глубокими, чем у волков MIS 7 и MIS 5. У них была самая длинная и самая узкая верхняя часть P4, что предполагает улучшенную режущую способность, и самую длинную верхнюю часть M1 и M2, но с меньшей шириной и, следовательно, уменьшенной способностью к раздавливанию, что указывает на гиперплотоядность. У них был более высокий процент поломки зубов и сильно изношенных зубов по сравнению с другими волками, и, возможно, они использовали верхний P4 и нижний m1 для дробления костей, а не коренные зубы, что приводило к более высокой частоте повреждений.[6] |
| 57000 MIS 3 | Палеосреда открытых лугов с летней температурой около 12 ° C и зимой около -20 ° C, где преобладают шерстистый мамонт, шерстистый носорог, лошадь и гигантский олень. Среди лучших хищников были лев, бурый медведь и пятнистая гиена. Волки из MIS 3 были меньше по размеру тела, чем волки из MIS 5 и те, что встречаются сегодня в Швеции. Эти волки уступили место льву и гиене, что привело к более всеядной диете с повышенной способностью к раздавливанию в открытой среде, которая поддерживала больше видов добычи и больше немясной пищи, чем в период MIS 5. У них были более мелкие и узкие челюсти, чем у волков MIS 5 и тех, что встречаются сегодня в Швеции, что указывало на то, что они могли ловить только мелкую и среднюю добычу. У них был более низкий процент поломки зубов, чем у волков MIS-7 с умеренным износом зубов.[6] |
| Сегодня (Швеция) | Волки были истреблены в Великобритании, но не в Швеции, где температуры аналогичны температурам в Британии в период MIS 7. Окружающая среда бореальный лес с летними температурами от 14 ° C до 18 ° C и зимними температурами от 1 ° C до −10 ° C. Хищные виды включают лосей, северных оленей, косуль, кабанов, зайцев, кроликов и бобров. Среди конкурентов - бурый медведь и рысь, но волк - главное хищник. Волки, встречающиеся сегодня в Швеции, меньше по размеру тела, чем волки MIS 5, но больше, чем у волков MIS 7 и MIS 3. Верхняя длина M1 и M2 длиннее, чем у волков MIS 7 и MIS 3, а челюсти глубже и шире, что указывает на способность охотиться и покорять крупную добычу. Однако большие коренные зубы сохранили способность измельчать и обрабатывать немясные продукты. Эти волки живут в бореальных лесах, где мелкую и среднюю дичь трудно обнаружить и усмирить трудоемко, что приводит к адаптации к охоте на крупную дичь с более высокой наградой. Это гиперплотоядные животные, похожие на волков из MIS 5, но не обладающие такой же способностью резать.[6] |
В течение Последний ледниковый максимум 20000 YBP, Плейстоценовая степь простирался через северную и центральную Евразию и через Берингия в Северную Америку. Плейстоцен волки Берингии и, возможно, степные, приспособились к этой среде обитания. Морфология их зубов и черепа указывает на то, что они специализировались на добыче ныне вымерших Мегафауна плейстоцена Износ зубов указывает на то, что их поведение отличалось от поведения современных волков.[110][7][184][185] Это подчеркивает успех С. волчанка как вид приспосабливается к различным условиям окружающей среды.[6] Этот экоморф серого волка вымер в конце оледенения вместе с лошадьми и другими видами, от которых он зависел, и был заменен волками из южной части Северной Америки. Это указывает на то, что специализированные волчьи экоморфы могут исчезнуть при изменении среды их обитания, даже если среда обитания все еще может поддерживать других волков.[7] Волки прошли через узкое место населения 20000 YBP, что совпадает с Последним ледниковым максимумом,[9][128][73][7] что указывает на то, что многие популяции волков, возможно, вымерли одновременно с берингийскими волками.[7]
Есть небольшое количество Canis останки, которые были найдены в пещере Гойе, Бельгия (36 500 YBP)[64] Пещера Разбойничья, Россия (33,500 лыб)[186] Костенки 8, Россия (33,500–26,500 YBP)[187] Предмости, Чехия (31,000 YBP)[188] и Елисеевичи 1, Россия (17000 юаней).[65] На основании морфометрического исследования черепа характеристик, которые, как считается, связаны с процессом одомашнивания, эти собаки были предложены как ранние палеолитические собаки.[187] Эти характеристики укороченного рострума, скученности зубов и отсутствия или вращения премоляров были зарегистрированы как у древних, так и у современных волков.[110][73][6][189][190][191] Вместо того, чтобы представлять первых собак, эти образцы могут представлять «морфологически отличную местную, ныне вымершую популяцию волков».[73][192]
Заметки
- ^ Филогенетические отношения и время генетическая дивергенция для сохранившихся волкоподобные псовые на основе ДНК всего ядро клетки.[104] Исключение 1: полосатый шакал и шакал с черной спиной добавлены на основе сегментов ДНК, взятых из ядро клетки[83][84] с предполагаемым временем расхождения в миллионах лет.[84] Исключение 2: положение гималайского волка основано на сегментах митохондриальная ДНК[84][105][106][107] и сегменты из ядра клетки.[84][106][107] Исключение 3: Положение индийского равнинного волка основано на мДНК.[108][109][110][111][112][113] и его время 120 000 лет назад.[114] Исключение 4: Поздний плейстоцен волк включен только для целей временной шкалы - самый последний общий предок для всех остальных Canis lupus образцы - современные и вымершие, за исключением равнинных индейских волков и гималайских волков - были за 80 000 лет до настоящего времени.[66][115] Исключение 5: Серый волк и собака отсчитывают время 25 000 лет назад.[114][116][117]
использованная литература
- ^ Докинз, Уильям Бойд, сэр; Сэнфорд, У. Эйшфорд; Рейнольдс, Сидней Х. (1912). "Британские плейстоцены Hyænid, Ursid, Canid и Mustelid". Монография британских плейстоценовых млекопитающих. 2. Лондон: Палеонтографическое общество.
- ^ а б c d е Мусиани, Марко; Леонард, Дженнифер А .; Клафф, Х. Дин; Гейтс, К. Кормак; Мариани, Стефано; Paquet, Paul C .; Вила, Карлес; Уэйн, Роберт К. (2007). «Дифференциация волков тундры / тайги и бореальных хвойных лесов: генетика, цвет шерсти и ассоциация с мигрирующим карибу». Молекулярная экология. 16 (19): 4149–70. Дои:10.1111 / j.1365-294X.2007.03458.x. PMID 17725575.
- ^ а б c d Schweizer, Rena M .; Vonholdt, Bridgett M .; Харриган, Райан; Ноулз, Джеймс С.; Мусиани, Марко; Колтман, Дэвид; Новембре, Джон; Уэйн, Роберт К. (2016). «Генетическое подразделение и гены-кандидаты при отборе у североамериканских серых волков». Молекулярная экология. 25 (1): 380–402. Дои:10.1111 / mec.13364. PMID 26333947.
- ^ а б c d Геффен, ELI; Андерсон, Марти Дж .; Уэйн, Роберт К. (2004). «Климатические и средовые барьеры для распространения высокомобильного серого волка». Молекулярная экология. 13 (8): 2481–90. Дои:10.1111 / j.1365-294X.2004.02244.x. PMID 15245420.
- ^ а б c Пилот Малгожата; Енджеевский, Влодзимеж; Браницки, Войцех; Сидорович, Вадим Е .; Енжеевска, Богумила; Стахура, Кристина; Функ, Стефан М. (2006). «Экологические факторы влияют на популяционно-генетическую структуру европейских серых волков». Молекулярная экология. 15 (14): 4533–53. Дои:10.1111 / j.1365-294X.2006.03110.x. PMID 17107481.
- ^ а б c d е ж г час я j Цветок, Люси О.Х .; Шреве, Даниэль С. (2014). "Исследование палеодиетической изменчивости европейских псовых плейстоцена". Четвертичные научные обзоры. 96: 188–203. Bibcode:2014QSRv ... 96..188F. Дои:10.1016 / j.quascirev.2014.04.015.
- ^ а б c d е ж г час я j k л м п о п Леонард, Дженнифер (2014). «Экология движет эволюцией серых волков» (PDF). Исследования эволюции экологии. 16: 461–473.
- ^ а б c Сотникова, М (2010). «Распространение Canini (Mammalia, Canidae: Caninae) по Евразии в период от позднего миоцена до раннего плейстоцена». Четвертичный международный. 212 (2): 86–97. Bibcode:2010QuInt.212 ... 86S. Дои:10.1016 / j.quaint.2009.06.008.
- ^ а б c d е ж г час я j k л м п о п q Freedman, Adam H .; Гронау, Илан; Schweizer, Rena M .; Ортега-дель-Векчё, Диего; Хан, Ынджунг; Silva, Pedro M .; Галаверни, Марко; Фань, Чжэньсинь; Маркс, Петр; Лоренте-Галдос, Белен; Бил, Холли; Рамирес, Оскар; Хормоздиари, Фархад; Алкан, банка; Вила, Карлес; Сквайр, Кевин; Геффен, Эли; Кусак, Иосип; Бойко, Адам Р .; Паркер, Хайди Дж .; Ли, Кларенс; Тадиготла, Васишт; Siepel, Адам; Bustamante, Carlos D .; Харкинс, Тимоти Т .; Нельсон, Стэнли Ф .; Острандер, Элейн А .; Маркес-Боне, Томас; Уэйн, Роберт К .; и другие. (2014). «Секвенирование генома подчеркивает динамическую раннюю историю собак». PLOS Genetics. 10 (1): e1004016. Дои:10.1371 / journal.pgen.1004016. ЧВК 3894170. PMID 24453982.
- ^ а б c d е ж г час я j k л м п Скоглунд, Понт; Эрсмарк, Эрик; Палкопулу, Элефтерия; Дален, Любовь (2015). «Геном древнего волка свидетельствует о раннем расхождении предков домашних собак и их смешении с высокоширотными породами». Текущая биология. 25 (11): 1515–1519. Дои:10.1016 / j.cub.2015.04.019. PMID 26004765.
- ^ а б c d е ж г час я j k л м п о п q р s т ты v ш Икс у Фань, Чжэньсинь; Сильва, Педро; Гронау, Илан; Ван, Шуогуо; Армеро, Айтор Серрес; Schweizer, Rena M .; Рамирес, Оскар; Поллингер, Джон; Галаверни, Марко; Ортега Дель-Векчё, Диего; Ду, Ляньминь; Чжан, Вэньпин; Чжан, Чжихэ; Син, Цзиньчуань; Вила, Карлес; Маркес-Боне, Томас; Годиньо, Ракель; Юэ, Бисон; Уэйн, Роберт К. (2016). «Мировые закономерности геномной изменчивости и смешения серых волков». Геномные исследования. 26 (2): 163–73. Дои:10.1101 / гр.197517.115. ЧВК 4728369. PMID 26680994.
- ^ а б c d Сарделла, Рафаэле; Берте, Давиде; Юрино, Давид Адам; Черин, Марко; Тальякоццо, Антонио (2014). «Волк из грота Романелли (Апулия, Италия) и его значение в эволюционной истории Canis lupus в позднем плейстоцене Южной Италии». Четвертичный международный. 328–329: 179–195. Bibcode:2014QuInt.328..179S. Дои:10.1016 / j.quaint.2013.11.016.
- ^ Флинн, Джон Дж .; Уэсли-Хант, Джина Д. (2005). «Филогения плотоядных: основные отношения между хищными морфами и оценка положения« Miacoidea »по отношению к хищникам». Журнал систематической палеонтологии. 3: 1–28. Дои:10.1017 / с1477201904001518. S2CID 86755875.
- ^ "Eucyon davisi". Окаменелости. Получено 11 июля 2016.
- ^ а б c d е ж г час я j k л м п Ван, Сяомин; Тедфорд, Ричард Х .; Собаки: их ископаемые родственники и история эволюции. Нью-Йорк: Издательство Колумбийского университета, 2008.
- ^ Вислобокова И., Сотникова М., Додонов А., 2003 - Биологические события и разнообразие позднемиоцен-плиоценовой фауны млекопитающих России и сопредельных территорий - в: Reumer, J.W.F. & Wessels, W. (ред.) - РАСПРОСТРАНЕНИЕ И МИГРАЦИЯ ТРЕТИЧНЫХ МЛЕКОПИТАЮЩИХ В ЕВРАЗИИ. ТОМ В ПОЧЕТУ ГАНСА ДЕ БРЮЙНА - DEINSEA 10: 563–574 [ISSN 0923-9308] Опубликовано 1 декабря 2003 г.
- ^ а б c d е ж г час я j k л м п о п q р Р.М. Новак (2003). «Глава 9 - Эволюция и систематика волков». В Mech, Л. Дэвид; Бойтани, Луиджи (ред.). Волки: поведение, экология и сохранение. Издательство Чикагского университета. С. 239–258. ISBN 978-0-226-51696-7.
- ^ а б c d е ж г час я j k л м п о п Новак, Р. М. (1979). Североамериканский четвертичный период Canis. 6. Монография Музея естественной истории Канзасского университета. С. 1–154. ISBN 978-0-89338-007-6.
- ^ Уэйн, Р. К. (1995). «Красный волк: сберегать или не сберегать» (PDF). В Macdonald, D .; Handoca, L. (ред.). Canid News. 3. Информационный бюллетень группы специалистов МСОП / SSC. С. 7–12.
- ^ Хоффмайстер Д. Ф. и У. У. Гудпастер. 1954. Млекопитающие гор Уачука, юго-восток Аризоны. Биологические монографии Иллинойса 24: 1–152.
- ^ Лоуренс, Б.; Боссерт, В. Х. (1967). "Многосимвольный анализ Canis lupus, латранс и Нигер". Американский зоолог. 7 (2): 223–232. Дои:10.1093 / icb / 7.2.223. JSTOR 3881428.
- ^ Лоуренс, Б.; Боссерт, В. Х. (1975). "Отношения североамериканских Canis показано многократным анализом отдельных популяций ". В Fox, M. W. (ed.). Дикие собаки: их систематика, поведенческая экология и эволюция. Нью-Йорк: Ван Ностранд-Рейнхольд. ISBN 978-0-442-22430-1.
- ^ а б Martin, R.A .; Уэбб, С. Д. (1974). «Позднеплейстоценовые млекопитающие из фауны Логова Дьявола, графство Леви». В Уэббе, С. Д. (ред.). Плейстоценовые млекопитающие Флориды. Гейнсвилл: Университетские прессы Флориды. стр.114–145. ISBN 978-0-8130-0361-0.
- ^ Уэбб, С. Д. (1974). «Хронология плейстоценовых млекопитающих Флориды». Плейстоценовые млекопитающие Флориды. Гейнсвилл: Университетские прессы Флориды. стр.5–31. ISBN 978-0-8130-0361-0.
- ^ Уэйн, Р. К .; Дженкс, С. М. (1991). «Анализ митохондриальной ДНК поддерживает обширную гибридизацию красного волка, находящегося под угрозой исчезновения (Canis rufus)». Природа. 351 (6327): 565–68. Bibcode:1991 Натур.351..565Вт. Дои:10.1038 / 351565a0. S2CID 4364642.
- ^ "Canis lepophagus". Окаменелости. Получено 11 июля 2016.
- ^ а б c Куртен Б. (1974). «История койотоподобных собак (Canidae, Mamalia)». Acta Zoologica Fennica (140): 1–38.
- ^ а б c d е Kurten, B .; Андерсон, Э. (1980). Плейстоценовые млекопитающие Северной Америки. Нью-Йорк: издательство Колумбийского университета. С. 1–442. ISBN 978-0-231-03733-4.
- ^ а б Джонстон, С. С. (1938). «Предварительный отчет о местонахождении позвоночных в каньоне Сита и описание койота-предка». Американский журнал науки. 5. 35 (209): 383–390. Bibcode:1938AmJS ... 35..383J. Дои:10.2475 / ajs.s5-35.209.383.
- ^ а б c d е ж г час я j k л м Тедфорд, Ричард Х .; Ван, Сяомин; Тейлор, Берил Э. (2009). "Филогенетическая систематика североамериканских ископаемых собак (Carnivora: Canidae)" (PDF). Бюллетень Американского музея естественной истории. 325: 1–218. Дои:10.1206/574.1. HDL:2246/5999. S2CID 83594819.
- ^ а б Тедфорд, Р.Х. и Цю, З.-Х., 1996 - Новый род псовых из плиоцена Юшэ, провинция Шаньси - Vertebrata PalAsiatica 34 (1): 27-40
- ^ Андерсон, Э. (1996). «Предварительный отчет о хищниках из пещеры Дикобраза, округ Парк, Колорадо». In Churcher, C. S. R .; Стюарт, К. М .; Сеймур, К. Л. (ред.). Палеоэкология и палеообстановка позднекайнозойских млекопитающих. Торонто: Университет Торонто Press. С. 259–282. ISBN 978-0-8020-0728-5.
- ^ Олбрайт, III, Л. Б. (2000). Биостратиграфия и палеонтология позвоночных бесплодных земель Сан-Тимотео, Южная Калифорния. Публикации Калифорнийского университета в области геологических наук. 144. Калифорнийский университет Press. С. 1–121. ISBN 978-0-520-91598-5.
- ^ "Canis edwardii". Окаменелости. Получено 11 июля 2016.
- ^ Берта, А. (1995). «Ископаемые плотоядные животные из карьеров Leisey Shell Pits, округ Хиллсборо, Флорида». В Hulbert, Jr., R.C .; Морган, Г. С .; Уэбб, С. Д. (ред.). Палеонтология и геология карьера Leisey Shell Pits, ранний плейстоцен Флориды. 37. Бюллетень Флоридского музея естественной истории. С. 463–499.
- ^ "Canis ambrusteri". Окаменелости. Получено 11 июля 2016.
- ^ "Canis dirus". Окаменелости. Получено 11 июля 2016.
- ^ Куртен Б. 1984: Географическая дифференциация лютого волка ранчолабрейных (Canis dirus Leidy) в Северной Америке. В Genoways, Х. Х. и Доусон, М. Р. (ред.): Вклад в палеонтологию четвертичных позвоночных: Том в память Джона Э. Гилдея, 218–227. Специальная публикация 8 Музея естественной истории Карнеги
- ^ Гейст, Валериус (1998). Олени мира: их эволюция, поведение и экология (1-е изд.). Stackpole Books, Механиксберг, Пенсильвания. п. 10. ISBN 0811704963.
- ^ Гейст, Валериус (1987). «О видообразовании млекопитающих ледникового периода, с особым вниманием к шейкам и капридам». Канадский журнал зоологии. 65 (5): 1067–1084. Дои:10.1139 / z87-171.
- ^ а б Гуле, Г. Д. (1993). Сравнение временных и географических изменений черепа у серых волков неарктического, современного голоцена и позднего плейстоцена (Canis lupus) и выбран Canis (Степень магистра). Виннипег: Университет Манитобы. С. 1–116.
- ^ Берта, А. (1988). Четвертичная эволюция и биогеография крупных южноамериканских Canidae (Mammalia: Carnivora). 132. Публикации Калифорнийского университета в области геологических наук. С. 1–49. ISBN 978-0-520-09960-9.
- ^ Новак, Р. М .; Федерофф, Н. Э. (2002). «Систематический статус итальянского волка. Canis lupus". Acta Theriol. 47 (3): 333–338. Дои:10.1007 / BF03194151. S2CID 366077.
- ^ а б Canis occidentalis furlongi Мерриам (1910), Univ. California Publ. Бык. Отдел геол., 5: 393
- ^ Стивенсон, Марк (1978). «9 - Систематика и поведение лютого волка». In Hall, Roberta L .; Шарп, Генри С. (ред.). Волк и человек: эволюция параллельно. Академическая пресса. С. 183–4. ISBN 0123192501.
- ^ Canis milleri Мерриам (1912) Mem. Univ. Калифорния, 1: 247
- ^ Мерриам, Дж. К. (1912). «Фауна Ранчо Ла Бреа. Часть II. Псовые». Мемуары Калифорнийского университета. 1: 217–273.
- ^ Aenocyon milleri Merriam (1918) Univ. California Publ. Бык. Геол., 10: 533.
- ^ а б Куртен, Б. (1968). Плейстоценовые млекопитающие Европы. Лондон: Вайденфельд и Николсон. п. 317. ISBN 978-0-202-30953-8.
- ^ а б Тонг; Hu, N .; Ван, X. (2012). «Новые останки Canis chihliensis (Mammalia, Carnivora) из Шаньшенмяозуй, стоянки нижнего плейстоцена в Янъюане, Хэбэй». Позвоночные PalAsiatica. 50 (4): 335–360.
- ^ а б c d е ж г час я Колер-Мацник, Дженис (2002). "Снова о происхождении собаки" (PDF). Anthrozoos: мультидисциплинарный журнал о взаимодействии людей и животных. 15 (2): 98–118. Дои:10.2752/089279302786992595. S2CID 14850835.
- ^ а б Олсен, С. Дж. (1985). Происхождение домашней собаки: летопись окаменелостей. Univ. Аризона Пресс, Тусон, США. С. 88–89.
- ^ Олсен С. Дж., Олсен Дж. В., Ци Г. К., 1982. Положение Canis lupus variabilis из Чжоукоудиана в родословной домашней собаки Canis familis. Vert PalAsia, 20 (3): 264–267 (на китайском языке с аннотацией последней страницы на английском языке)
- ^ Лоуренс Б. 1967. Ранние домашние собаки. Zeitschrift für Säugetierkunde 32: 44–59.
- ^ Пей, W.C. (1934). Плотоядное животное из населенного пункта 1 Чукутьен. Palaeontologia Sinica, серия C, т. 8, пучок 1. Геологическая служба Китая, Пекин. С. 1–45.
- ^ Вестгейт, Джон А; Пирс, Дж. Уильям; Прис, Шари Дж; Schweger, Charles E; Морлан, Ричард Э; Пирс, Николас Дж. Г.; Перкинс, Т. Уильям (2017). «Тефрохронология, магнитостратиграфия и фауны млекопитающих отложений среднего и раннего плейстоцена на двух участках реки Олд Кроу, северная территория Юкон, Канада». Четвертичное исследование. 79: 75–85. Дои:10.1016 / j.yqres.2012.09.003.
- ^ Бонифай, М.Ф., 1971. Четвертичные плотоядные животные Южного Востока Франции. Национальный музей естественной истории, Париж, 334 стр. (Mémoires du Muséum national d'Histoire naturelle, Sér. C - Sciences de la Terre (1950–1992); 21 (2)). [Четвертичные хищники юго-востока Франции]
- ^ Аргант А., 1991 - Четвероядные хищники Бургонь. Documents du Laboratoire de Géologie de Lyon, № 115, 301 стр.
- ^ Будади-Малинь, Мириам (2012). "Une nouvelle sous-espèce de loup (Canis lupus maximus nov. Subsp.) Dans le Pléistocène supérieur d'Europe occidentale [Новый подвид волка (Canis lupus maximus nov. Subsp.) Из верхнего плейстоцена Западной Европы]". Comptes Rendus Palevol. 11 (7): 475. Дои:10.1016 / j.crpv.2012.04.003.
- ^ Brugal, J. P .; Будади-Малинь, М. (2011). «Четвертичные от малых до крупных псовых в Европе: таксономический статус и биохронологический вклад». Четвертичный международный. 243: 171–182. Дои:10.1016 / j.quaint.2011.01.046.
- ^ Анзидей, Анна Паола; Булгарелли, Грация Мария; Каталано, Паола; Керилли, Эухенио; Галлотти, Розалия; Леморини, Кристина; Милли, Сальваторе; Паломбо, Мария Рита; Пантано, Уолтер; Сантуччи, Эрнесто (2012). «Текущие исследования на территории позднего среднего плейстоцена Ла-Полледрара-ди-Чеканиббио (центральная Италия) с упором на взаимоотношения человека и слона». Четвертичный международный. 255: 171–187. Дои:10.1016 / j.quaint.2011.06.005.
- ^ Лученти, Саверио Бартолини; Бухсианидзе, Майя; Мартинес-Наварро, Бьенвенидо; Лордкипанидзе, Давид (2020). «Волк из Дманиси и дополненная реальность: обзор, последствия и возможности». Границы науки о Земле. 8: 131. Bibcode:2020FrEaS ... 8..131B. Дои:10.3389 / feart.2020.00131. S2CID 216337804.
- ^ Тельман, Олаф; Перри, Анджела Р. (2018). "Палеогеномные выводы об одомашнивании собак". In Lindqvist, C .; Раджора, О. (ред.). Палеогеномика. Популяционная геномика. Спрингер, Чам. С. 273–306. Дои:10.1007/13836_2018_27. ISBN 978-3-030-04752-8.
- ^ а б c Жермонпре, Митье; Саблин, Михаил В .; Стивенс, Рианнон Э .; Hedges, Роберт E.M .; Хофрайтер, Майкл; Стиллер, Матиас; Депре, Вивиан Р. (2009). «Ископаемые собаки и волки из палеолитических памятников Бельгии, Украины и России: остеометрия, древняя ДНК и стабильные изотопы». Журнал археологической науки. 36 (2): 473–490. Дои:10.1016 / j.jas.2008.09.033.
- ^ а б Саблин, Михаил В .; Хлопачев, Геннадий А. (2002). "Древнейшие собаки ледникового периода: свидетельства из Елисеевичей I" (PDF). Фонд Веннер-Грена антропологических исследований. Получено 10 января 2015. Цитировать журнал требует
| журнал =(Помогите) - ^ а б c d е ж г час я j k л Thalmann, O .; Шапиро, Б .; Cui, P .; Schuenemann, V.J .; Сойер, С. К .; Гринфилд, Д. Л .; Germonpre, M. B .; Саблин, М. В .; Lopez-Giraldez, F .; Доминго-Роура, X .; Napierala, H .; Uerpmann, H.-P .; Loponte, D.M .; Acosta, A. A .; Giemsch, L .; Schmitz, R.W .; Worthington, B .; Buikstra, J. E .; Дружкова, А .; Графодацкий, А. С .; Оводов, Н. Д .; Wahlberg, N .; Freedman, A. H .; Schweizer, R.M .; Koepfli, K.-P .; Леонард, Дж. А .; Мейер, М .; Krause, J .; Paabo, S .; и другие. (2013). «Полные митохондриальные геномы древних псовых предполагают европейское происхождение домашних собак». Наука. 342 (6160): 871–4. Bibcode:2013Наука ... 342..871Т. Дои:10.1126 / science.1243650. HDL:10261/88173. PMID 24233726. S2CID 1526260.
- ^ а б c Мори, Дарси Ф .; Jeger, Rujana (2015). «Палеолитические собаки: зачем тогда продолжалось приручение?». Журнал археологической науки: отчеты. 3: 420–428. Дои:10.1016 / j.jasrep.2015.06.031.
- ^ Уэйн, Роберт К. (1986). "Морфология черепа домашних и диких собак: влияние развития на морфологические изменения". Эволюция. 40 (2): 243–261. Дои:10.2307/2408805. JSTOR 2408805. PMID 28556057.
- ^ Дрейк, Эбби Грейс; Клингенберг, Кристиан Питер (2010). «Крупномасштабная диверсификация формы черепа у домашних собак: несоответствие и модульность». Американский натуралист. 175 (3): 289–301. Дои:10.1086/650372. PMID 20095825.
- ^ Робертс, Тарин; МакГриви, Пол; Валенсуэла, Майкл (2010). «Индуцированное человеком вращение и реорганизация мозга домашних собак». PLOS ONE. 5 (7): e11946. Дои:10.1371 / journal.pone.0011946. ЧВК 2909913. PMID 20668685.
- ^ Дрейк, Эбби Грейс, «Эволюция и развитие морфологии черепа собак: исследование морфологической интеграции и гетерохронии» (1 января 2004 г.). Докторские диссертации доступны через Proquest. Бумага AAI3136721. ссылка на сайт
- ^ Дрейк, Эбби Грейс (2011). "Рассеивание догмы собак: исследование гетерохронии у собак с использованием трехмерного геометрического морфометрического анализа формы черепа". Эволюция и развитие. 13 (2): 204–213. Дои:10.1111 / j.1525-142X.2011.00470.x. PMID 21410876.
- ^ а б c d е ж г Перри, Анджела (2016). «Волк в собачьей шкуре: начальное приручение собаки и вариация плейстоценового волка». Журнал археологической науки. 68: 1–4. Дои:10.1016 / j.jas.2016.02.003.
- ^ Рэнд, Дэвид М. (2001). «Единицы отбора на митохондриальной ДНК». Ежегодный обзор экологии и систематики. 32: 415–448. Дои:10.1146 / annurev.ecolsys.32.081501.114109.
- ^ а б c d е ж г час Роберт К. Уэйн, Дженнифер А. Леонард, Карлес Вила (2006). «Глава 19: Генетический анализ одомашнивания собак». В Мелинде А. Зедер (ред.). Документирование приручения: новые генетические и археологические парадигмы. Калифорнийский университет Press. С. 279–295. ISBN 978-0-520-24638-6.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ Хофрайтер, Майкл; Серр, Дэвид; Пойнар, Хендрик Н .; Куч, Мелани; Паабо, Сванте (2001). «Древняя ДНК». Природа Обзоры Генетика. 2 (5): 353–9. Дои:10.1038/35072071. PMID 11331901. S2CID 205016024.
- ^ Ли, Вэнь-Сюн (1997). Молекулярная эволюция. Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-463-8.[страница нужна ]
- ^ Песоле, G; Gissi, C; Де Кирико, А; Сакконе, С. (1999). «Скорость нуклеотидных замен митохондриальных геномов млекопитающих». Журнал молекулярной эволюции. 48 (4): 427–34. Дои:10.1007 / pl00006487. PMID 10079281. S2CID 1665621.
- ^ Ван, Цю-Хун; Ву, Хуа; Фуджихара, Цутому; Фанг, Шэн-Го (2003). «Какой генетический маркер для какой генетической проблемы сохранения?» (PDF). Электрофорез. 25 (14): 2165–2176. Дои:10.1002 / elps.200305922. PMID 15274000.
- ^ Бирки, К. Уильям (2001). «Наследование генов в митохондриях и хлоропластах: законы, механизмы и модели». Ежегодный обзор генетики. 35: 125–48. Дои:10.1146 / annurev.genet.35.102401.090231. PMID 11700280.
- ^ Ависе, Дж. К. (1994). Молекулярные маркеры, естественная история и эволюция. Нью-Йорк: Чепмен и Холл. ISBN 978-0-412-03781-8.
- ^ Ависе, Дж. К. (2000). Филогеография: история и формирование видов. Кембридж, Массачусетс: Издательство Гарвардского университета. ISBN 978-0-674-66638-2.
- ^ а б c d е ж г Линдблад-То, Керстин; Уэйд, Клэр М .; Mikkelsen, Tarjei S .; Карлссон, Элинор К .; Джефф, Дэвид Б .; Камаль, Майкл; Зажим, Микеле; Чанг, Жан Л .; Кульбокас, Эдвард Дж .; Зоди, Майкл С .; Мусели, Эван; Се, Сяохуэй; Брин, Мэтью; Уэйн, Роберт К .; Острандер, Элейн А .; Понтинг, Крис П .; Галиберт, Фрэнсис; Smith, Douglas R .; Деджонг, Питер Дж .; Киркнесс, Юэн; Альварес, Пабло; Бьяджи, Тара; Брокман, Уильям; Батлер, Джонатан; Чин, Чи-Уай; Повар, апрель; Кафф, Джеймс; Дейли, Марк Дж .; Декаприо, Дэвид; и другие. (2005). «Последовательность генома, сравнительный анализ и структура гаплотипов домашней собаки». Природа. 438 (7069): 803–819. Bibcode:2005Натура.438..803л. Дои:10.1038 / природа04338. PMID 16341006.
- ^ а б c d е ж г час Koepfli, K.-P .; Pollinger, J .; Godinho, R .; Робинсон, Дж .; Lea, A .; Хендрикс, С .; Schweizer, R.M .; Thalmann, O .; Silva, P .; Fan, Z .; Юрченко, А. А .; Добрынин, П .; Макунин, А .; Cahill, J. A .; Шапиро, Б .; Álvares, F .; Brito, J.C .; Geffen, E .; Леонард, Дж. А .; Helgen, K. M .; Johnson, W. E .; О’Брайен, С. Дж .; Van Valkenburgh, B .; Уэйн, Р. К. (17 августа 2015 г.). «Данные по всему геному показывают, что африканские и евразийские золотые шакалы являются разными видами». Текущая биология. 25 (16): 2158–65. Дои:10.1016 / j.cub.2015.06.060. PMID 26234211.
- ^ а б c d е Вила, К. (1997). «Множественное и древнее происхождение домашней собаки». Наука. 276 (5319): 1687–9. Дои:10.1126 / science.276.5319.1687. PMID 9180076.
- ^ а б c d Уэйн, Р. и Острандер, Элейн А. (1999). «Происхождение, генетическое разнообразие и структура генома домашней собаки». BioEssays. 21 (3): 247–57. Дои:10.1002 / (SICI) 1521-1878 (199903) 21: 3 <247 :: AID-BIES9> 3.0.CO; 2-Z. PMID 10333734.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ Бойко, А. (2009). «Сложная структура популяции африканских деревенских собак и ее значение для вывода истории одомашнивания собак». Труды Национальной академии наук. 106 (33): 13903–13908. Bibcode:2009PNAS..10613903B. Дои:10.1073 / pnas.0902129106. ЧВК 2728993. PMID 19666600.
- ^ Панг, Дж. (2009). «Данные мтДНК указывают на единственное происхождение собак к югу от реки Янцзы, менее 16 300 лет назад, от многочисленных волков». Молекулярная биология и эволюция. 26 (12): 2849–64. Дои:10.1093 / молбев / msp195. ЧВК 2775109. PMID 19723671.
- ^ а б Cronin, M. A .; Canovas, A .; Bannasch, D. L .; Обербауэр, А. М .; Медрано, Дж. Ф. (2014). «Вариация полиморфизма одиночных нуклеотидов (SNP) у волков (Canis lupus) на юго-востоке Аляски и сравнение с волками, собаками и койотами в Северной Америке». Журнал наследственности. 106 (1): 26–36. Дои:10.1093 / jhered / esu075. PMID 25429025.
- ^ Барделебен, Кэролайн; Мур, Рэйчел Л .; Уэйн, Роберт К. (2005). «Молекулярная филогения Canidae на основе шести ядерных локусов». Молекулярная филогенетика и эволюция. 37 (3): 815–31. Дои:10.1016 / j.ympev.2005.07.019. PMID 16213754.
- ^ Грей, Мелисса М; Саттер, Натан Б; Острандер, Элейн А; Уэйн, Роберт К (2010). «Гаплотип маленькой собаки IGF1 происходит от серых волков Ближнего Востока». BMC Биология. 8: 16. Дои:10.1186/1741-7007-8-16. ЧВК 2837629. PMID 20181231.
- ^ Уэйн, Роберт К .; Вонхольдт, Бриджит М. (2012). «Эволюционная геномика одомашнивания собак». Геном млекопитающих. 23 (1–2): 3–18. Дои:10.1007 / s00335-011-9386-7. PMID 22270221. S2CID 16003335.
- ^ Мех, Л. Дэвид; Барбер-Мейер, Шеннон (2017). «Использование ошибочного времени поколения волков в оценке эволюции домашних собак и человека» (PDF). Наука. 352: 1228–31. Bibcode:2016Научный ... 352.1228F. Дои:10.1126 / science.aaf3161. PMID 27257259. S2CID 206647686.
- ^ Frantz, L.A. F .; Муллин, В. Э .; Pionnier-Capitan, M .; Lebrasseur, O .; Ollivier, M .; Perri, A .; Linderholm, A .; Mattiangeli, V .; Teasdale, M.D .; Dimopoulos, E.A .; Tresset, A .; Duffraisse, M .; McCormick, F .; Bartosiewicz, L .; Gal, E .; Nyerges, E. A .; Саблин, М. В .; Brehard, S .; Машкур, М .; b l Escu, A .; Gillet, B .; Hughes, S .; Chassaing, O .; Hitte, C .; Vigne, J.-D .; Добней, К .; Hanni, C .; Bradley, D.G .; Ларсон, Г. (2016). «Геномные и археологические данные свидетельствуют о двойном происхождении домашних собак». Наука. 352 (6290): 1228–31. Bibcode:2016Научный ... 352.1228F. Дои:10.1126 / science.aaf3161. PMID 27257259. S2CID 206647686.
- ^ а б Уэйн, Р. (1993). «Молекулярная эволюция семейства собак». Тенденции в генетике. 9 (6): 218–24. Дои:10.1016 / 0168-9525 (93) 90122-Х. PMID 8337763.
- ^ Уэйн, Р. К .; Benvenista, R.E .; Janczewski, D. N .; О'Брайен, С. Дж. (1989). «17-Молекулярная и биомеханическая эволюция хищников». В Гиттлман, Дж. Л. (ред.). Поведение, экология и эволюция хищников. Springer. С. 477–478. ISBN 9780412343605.
- ^ Vila, C .; Леонард, Дж. (2012). "Филогения 1-псовых и происхождение домашней собаки". In Ostrander, E. A .; Рувинский, А. (ред.). Генетика собаки (2-е изд.). КАБИ. п. 3. ISBN 9781845939403.
- ^ Wurster-Hill, D.H .; Центерволл, В. Р. (1982). «Взаимосвязь образцов полос хромосом у псовых, куньих, гиен и кошачьих». Цитогенетика и клеточная генетика. 34 (1–2): 178–192. Дои:10.1159/000131806. PMID 7151489.
- ^ Gottelli, D .; Sillero-Zubiri, C .; Applebaum, G.D .; Рой, М. С .; Гирман, Д. Дж .; Garcia-Moreno, J .; Острандер, Э. А .; Уэйн, Р. К. (1994). «Молекулярная генетика собак, находящихся под наибольшей угрозой исчезновения: эфиопский волк Canis simensis». Молекулярная экология. 3 (4): 301–12. Дои:10.1111 / j.1365-294X.1994.tb00070.x. PMID 7921357.
- ^ Покок, Р.И. (1941). Фауна Британской Индии: Млекопитающие. 2. Тейлор и Фрэнсис. С. 146–163.
- ^ Гирман, Д. Дж .; Vilà, C .; Geffen, E .; Creel, S .; Миллс, М. Г. Л .; McNutt, J. W .; Ginsberg, J .; Kat, P.W .; Mamiya, K. H .; Уэйн, Р. К. (2001). "Паттерны деления популяции, потока генов и генетической изменчивости африканской дикой собаки (Lycaon pictus)". Молекулярная экология. 10 (7): 1703–23. Дои:10.1046 / j.0962-1083.2001.01302.x. PMID 11472538.
- ^ Уэйн, Р.К .; Мейер, А .; Lehman, N .; van Valkenburgh, B .; Kat, P.W .; Фуллер, Т.К .; Гирман, Д .; О'Брайен, С.Дж. (1990). «Большое расхождение в последовательностях генотипов митохондриальной ДНК в популяциях восточноафриканских черноспинных шакалов» (PDF). Труды Национальной академии наук Соединенных Штатов Америки. 87 (5): 1772–1776. Bibcode:1990PNAS ... 87.1772W. Дои:10.1073 / pnas.87.5.1772. ЧВК 53565. PMID 1968637. Получено 21 декабря 2011.
- ^ Джулиана Камински и Сара Маршалл-Пескини (2014). «Глава 1 - Социальная собака: история и эволюция». Социальная собака: поведение и познание. Эльзевир. п. 4. ISBN 978-0-12407-931-1.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ а б c Гопалакришнан, Шьям; Sinding, Mikkel-Holger S .; Рамос-Мадригал, Джасмин; Ниманн, Йонас; Саманьего Каструита, Хосе А .; Vieira, Filipe G .; Карё, Кристиан; Монтеро, Марк де Мануэль; Кудерна, Лукас; Серрес, Айтор; Гонсалес-Базаллот, Виктор Мануэль; Лю, Ян-Ху; Ван, Го-Донг; Маркес-Боне, Томас; Мирараб, Сиаваш; Фернандес, Карлос; Гобер, Филипп; Кёпфли, Клаус-Петер; Бадд, Джейн; Руенесс, Эли Книспель; Хайде-Йоргенсен, Мэдс Петер; Петерсен, Бент; Зихериц-Понтен, Томас; Бахманн, Лутц; Уиг, Эйстейн; Hansen, Anders J .; Гилберт, М. Томас П. (2018). «Межвидовой поток генов повлиял на эволюцию рода Canis». Текущая биология. 28 (21): 3441–3449.e5. Дои:10.1016 / j.cub.2018.08.041. ЧВК 6224481. PMID 30344120.
- ^ а б c d е ж Эрсмарк, Эрик; Klütsch, Cornelya F.C .; Chan, Yvonne L .; Sinding, Mikkel-Holger S .; Файн, Стивен Р .; Илларионова Наталья А .; Оскарссон, Маттиас; Улен, Матиас; Чжан, Я-Пин; Дален, Любовь; Саволайнен, Питер (2016). «От прошлого к настоящему: филогеография и демографическая история Вольфа на основе контрольной области митохондрий». Границы экологии и эволюции. 4. Дои:10.3389 / fevo.2016.00134.
- ^ а б Верхан, Джеральдин; Сенн, Хелен; Каден, Дженнифер; Джоши, Джоти; Бхаттараи, Сусмита; Куси, Нареш; Силлеро-Зубири, Клаудио; Макдональд, Дэвид В. (2017). «Филогенетическое свидетельство древнего гималайского волка: к уточнению его таксономического статуса на основе генетической выборки из западного Непала». Королевское общество открытой науки. 4 (6): 170186. Bibcode:2017RSOS .... 470186W. Дои:10.1098 / rsos.170186. ЧВК 5493914. PMID 28680672.
- ^ а б Верхан, Джеральдин; Сенн, Хелен; Газали, Мухаммед; Кармачарья, Дибеш; Шерхан, Адарш Ман; Джоши, Джоти; Куси, Нареш; Лопес-Бао, Хосе Винсенте; Розен, Таня; Качел, Шеннон; Силлеро-Зубири, Клаудио; Макдональд, Дэвид В. (2018). «Уникальная генетическая адаптация гималайского волка к высокогорью и последствия для сохранения». Глобальная экология и сохранение. 16: e00455. Дои:10.1016 / j.gecco.2018.e00455.
- ^ а б Aggarwal, R.K .; Кивисилд, Т .; Ramadevi, J .; Сингх, Л. (2007). «Последовательности кодирующей области митохондриальной ДНК подтверждают филогенетическое различие двух видов индийских волков». Журнал зоологической систематики и эволюционных исследований. 45 (2): 163–172. Дои:10.1111 / j.1439-0469.2006.00400.x.
- ^ а б Шарма, Д. К .; Мальдонадо, Дж. Э .; Jhala, Y. V .; Флейшер, Р. К. (2004). «Древние родословные волков в Индии». Труды Королевского общества B: биологические науки. 271 (Приложение 3): S1 – S4. Дои:10.1098 / рсбл.2003.0071. ЧВК 1809981. PMID 15101402.
- ^ а б c d е ж г час я j Леонард, Дж. А .; Vilà, C; Фокс-Доббс, К; Koch, P. L .; Уэйн, Р. К .; Ван Валкенбург, Б. (2007). «Мегафауна вымирания и исчезновение специализированного волка-экоморфа» (PDF). Текущая биология. 17 (13): 1146–50. Дои:10.1016 / j.cub.2007.05.072. PMID 17583509. S2CID 14039133.
- ^ а б c d е ж г час я j k л м п о п q р Пилот, М .; и другие. (2010). «Филогеографическая история серых волков в Европе». BMC Эволюционная биология. 10: 104. Дои:10.1186/1471-2148-10-104. ЧВК 2873414. PMID 20409299.
- ^ Руенесс, Эли Книспель; Асмир, Мария Гулбрандсен; Силлеро-Зубири, Клаудио; Макдональд, Дэвид В; Бекеле, Афворк; Атикем, Анагау; Стенсет, Нильс Хр (2011). «Загадочный африканский волк: Canis aureus lupaster не золотой шакал и не является эндемиком Египта». PLOS ONE. 6 (1): e16385. Bibcode:2011PLoSO ... 616385R. Дои:10.1371 / journal.pone.0016385. ЧВК 3027653. PMID 21298107.
- ^ а б c d Миклоси, Адам (2015). Поведение, эволюция и познание собаки. Оксфордская биология (2-е изд.). Издательство Оксфордского университета. С. 106–107. ISBN 978-0199545667.
- ^ а б c Лоог, Лийза; Тельман, Олаф; Sinding, Mikkel ‐ Holger S .; Schuenemann, Verena J .; Перри, Анджела; Жермонпре, Митье; Бохеренс, Эрве; Witt, Kelsey E .; Саманьего Каструита, Хосе А .; Velasco, Marcela S .; Lundstrøm, Inge K.C .; Уэльс, Натан; Сонет, Гонтран; Франц, Лоран; Шредер, Ханнес; Бадд, Джейн; Хименес, Элоди-Лор; Федоров, Сергей; Гаспарян, Борис; Кандел, Эндрю В .; Лазничкова ‐ Галетова, Мартина; Напьерала, Ханнес; Уерпманн, Ханс-Петер; Никольский, Павел А .; Павлова, Елена Ю .; Питулько, Владимир В .; Герциг, Карл-Хайнц; Malhi, Ripan S .; Виллерслев, Эске; и другие. (2019). «Древняя ДНК предполагает, что современные волки ведут свое происхождение от позднего плейстоцена из Берингии». Молекулярная экология. 29 (9): 1596–1610. Дои:10.1111 / mec.15329. ЧВК 7317801. PMID 31840921.
- ^ а б c d е ж г час я j k л Кобльмюллер, Стефан; Вила, Карлес; Лоренте-Галдос, Белен; Дабад, Марк; Рамирес, Оскар; Маркес-Боне, Томас; Уэйн, Роберт К .; Леонард, Дженнифер А. (2016). «Целые митохондриальные геномы освещают древние межконтинентальные расселения серых волков (Canis lupus)». Журнал биогеографии. 43 (9): 1728–1738. Дои:10.1111 / jbi.12765. HDL:10261/153364.
- ^ а б Верхан, Джеральдин; Сенн, Хелен; Газали, Мухаммед; Кармачарья, Дибеш; Шерхан, Адарш Ман; Джоши, Джоти; Куси, Нареш; Лопес-Бао, Хосе Винсенте; Розен, Таня; Качел, Шеннон; Силлеро-Зубири, Клаудио; Макдональд, Дэвид В. (2018). «Уникальная генетическая адаптация гималайского волка к высокогорью и последствия для сохранения». Глобальная экология и сохранение. 16: e00455. Дои:10.1016 / j.gecco.2018.e00455.
- ^ а б c d е Schweizer, Rena M .; Уэйн, Роберт К. (2020). «Раскрывая тайны истории волков». Молекулярная экология. 29 (9): 1589–1591. Дои:10.1111 / MEC.15438. PMID 32286714.
- ^ Ван, Го-Донг; Чжан, Мин; Ван, Сюань; Ян, Мелинда А .; Цао, Пэн; Лю, Фэн; Лу, Хэн; Фэн, Сяотянь; Скоглунд, Понт; Ван, Лу; Фу, Цяомэй; Чжан, Я-Пин (2019). «Геномные подходы выявляют эндемичную субпопуляцию серых волков в Южном Китае». iScience. 20: 110–118. Дои:10.1016 / j.isci.2019.09.008. ЧВК 6817678. PMID 31563851.
- ^ Cox, C. B .; Мур, Питер Д .; Ковш, Ричард (2016). Биогеография: экологический и эволюционный подход. Вили-Блэквелл. п. 106. ISBN 978-1-118-96858-1.
- ^ Редакционная коллегия (2012 г.). Краткий научный словарь. Нью-Дели: Издательство V&S. п. 137. ISBN 978-93-81588-64-2.
- ^ а б Арора, Девендер; Сингх, Аджит; Шарма, Викрант; Бхадурия, Харвендра Сингх; Патель, Рам Бахадур (2015). "Hgs Db: База данных гаплогрупп для понимания миграции и оценки молекулярных рисков ". Биоинформация. 11 (6): 272–5. Дои:10.6026/97320630011272. ЧВК 4512000. PMID 26229286.
- ^ «Глоссарий по генетике». Международное общество генетической генеалогии. 2015 г.. Получено 22 июля 2016.
- ^ а б c Рэнди, Этторе (2011). «Генетика и сохранение волков Canis lupus в Европе». Обзор млекопитающих. 41 (2): 99–111. Дои:10.1111 / j.1365-2907.2010.00176.x.
- ^ Тамм, Э .; Кивисилд, Т .; Reidla, M .; Мецпалу, М .; Smith, D.G .; Mulligan, C.J .; Bravi, C.M .; Rickards, O .; Martinez-Labarga, C .; Хуснутдинова, Э.К .; Федорова, С. А .; Голубенко, М. В .; Степанов, В. А .; Губина, М. А .; Жаданов, С. И .; Осипова, Л. П .; Damba, L .; Воевода, М. И .; Dipierri, J. E .; Villems, R .; Малхи, Р. С. (2007). Картер, Ди (ред.). "Берингийский застой и распространение основателей коренных американцев". PLOS ONE. 2 (9): e829. Bibcode:2007PLoSO ... 2..829T. Дои:10.1371 / journal.pone.0000829. ЧВК 1952074. PMID 17786201.
- ^ а б Хофрайтер, Майкл; Барнс, Ян (2010). «Разнообразие утрачено: все ли виды крупных голарктических млекопитающих - это просто реликтовые популяции?». BMC Биология. 8: 46. Дои:10.1186/1741-7007-8-46. ЧВК 2858106. PMID 20409351.
- ^ Жермонпре, Митье; Саблин, Михаил В .; Деспрес, Вивиан; Хофрайтер, Майкл; Лазничкова-Галетова, Мартина; Стивенс, Рианнон Э .; Стиллер, Матиас (2013). «Палеолитические собаки и раннее приручение волка: ответ на комментарии Крокфорда и Кузьмина (2012)». Журнал археологической науки. 40: 786–792. Дои:10.1016 / j.jas.2012.06.016.
- ^ а б c Хофрейтер, Майкл (2007). «Плейстоценовые вымирания: преследующие выживших». Текущая биология. 17 (15): R609–11. Дои:10.1016 / j.cub.2007.06.031. PMID 17686436.
- ^ а б c Пилот, М; Греко, C; Vonholdt, B.M .; Jędrzejewska, B; Randi, E; Jędrzejewski, W; Сидорович, В Э; Острандер, Э А; Уэйн, Р. К. (2013). «Полногеномные признаки узких мест в популяции и диверсификация селекции европейских волков». Наследственность. 112 (4): 428–42. Дои:10.1038 / hdy.2013.122. ЧВК 3966127. PMID 24346500.
- ^ Weckworth, Byron V .; Талбот, Сандра; Сейдж, Джордж К .; Человек, Дэвид К .; Кук, Джозеф (2005). «Сигнал для независимых прибрежных и континентальных историй среди североамериканских волков». Молекулярная экология. 14 (4): 917–31. Дои:10.1111 / j.1365-294X.2005.02461.x. PMID 15773925.
- ^ Weckworth, Byron V .; Talbot, Sandra L .; Кук, Джозеф А. (2010). "Филогеография волков (Canis lupus) Тихоокеанского Северо-Запада". Журнал маммологии. 91 (2): 363–375. Дои:10.1644 / 09-МАММ-А-036.1.
- ^ Weckworth, Byron V .; Доусон, Натали Дж .; Talbot, Sandra L .; Flamme, Мелани Дж .; Кук, Джозеф А. (2011). «Побережье: общая история эволюции прибрежных районов Британской Колумбии и волков Юго-Восточной Аляски (Canis lupus)». PLOS ONE. 6 (5): e19582. Bibcode:2011PLoSO ... 619582W. Дои:10.1371 / journal.pone.0019582. ЧВК 3087762. PMID 21573241.
- ^ а б Vonholdt, B.M .; Cahill, J. A .; Fan, Z .; Gronau, I .; Робинсон, Дж .; Pollinger, J. P .; Шапиро, Б .; Wall, J .; Уэйн, Р. К. (2016). «Анализ последовательности всего генома показывает, что два эндемичных вида североамериканского волка представляют собой смесь койота и серого волка». Достижения науки. 2 (7): e1501714. Bibcode:2016SciA .... 2E1714V. Дои:10.1126 / sciadv.1501714. ЧВК 5919777. PMID 29713682.
- ^ а б c d Мацумура, Шуичи; Иношима, Ясуо; Исигуро, Наотака (2014). «Реконструкция истории колонизации потерянных линий волков с помощью анализа митохондриального генома». Молекулярная филогенетика и эволюция. 80: 105–12. Дои:10.1016 / j.ympev.2014.08.004. PMID 25132126.
- ^ Абэ, Х. (1999). «Разнообразие и сохранение млекопитающих Японии». In Yokohata, Y .; Накамура, С. (ред.). Последние достижения в биологии японских насекомоядных. Шубара: Общество естествознания Хиба. С. 89–104.
- ^ Исигуро, Наотака; Иношима, Ясуо; Шигехара, Нобуо; Итикава, Хидео; Като, Масару (2010). «Остеологический и генетический анализ вымершего волка эзо (Canis Lupus Hattai) с острова Хоккайдо, Япония». Зоологическая наука. 27 (4): 320–4. Дои:10.2108 / zsj.27.320. PMID 20377350. S2CID 11569628.
- ^ Уокер, Бретт (2008). Заблудшие волки Японии. Вашингтонский университет Press. п. 42.
- ^ а б c Исигуро, Наотака; Иношима, Ясуо; Шигехара, Нобуо (2009). «Анализ митохондриальной ДНК японского волка (Canis lupus hodophilax Temminck, 1839) и сравнение с репрезентативными гаплотипами волков и домашних собак ». Зоологическая наука. 26 (11): 765–70. Дои:10.2108 / zsj.26.765. PMID 19877836. S2CID 27005517.
- ^ а б Парди, Мелисса I .; Смит, Фелиса А. (2016). «Биотические реакции псовых на окончательное вымирание мегафауны плейстоцена». Экография. 39 (2): 141–151. Дои:10.1111 / ecog.01596.
- ^ Witt, Kelsey E .; Джадд, Кэтлин; Кухня, Андрей; Гриер, Колин; Колер, Тимоти А .; Ортман, Скотт Дж .; Кемп, Брайан М .; Малхи, Рипан С. (2015). «Анализ ДНК древних собак Америки: Определение возможных основополагающих гаплотипов и реконструкция истории популяции». Журнал эволюции человека. 79: 105–18. Дои:10.1016 / j.jhevol.2014.10.012. PMID 25532803.
- ^ Волк и человек. Эволюция параллельно. Р.Л. Холл, Х.С. Sharp. Academic Press, Нью-Йорк, 1978.
- ^ а б c Леонард, Дженнифер А .; Вила, Карлес; Уэйн, Роберт К. (2004). "Fast Track: Legacy lost: Генетическая изменчивость и размер популяций искорененных серых волков США (Canis lupus)". Молекулярная экология. 14 (1): 9–17. Дои:10.1111 / j.1365-294X.2004.02389.x. PMID 15643947.
- ^ Морелл, Вирджиния (2016). «Как спасти волка, который на самом деле не волк?». Наука. 353 (6300). Дои:10.1126 / science.aag0699.
- ^ Vila, C .; Amorim, I.R .; Леонард, Дж. А .; Posada, D .; Castroviejo, J .; Petrucci-Fonseca, F .; Crandall, K. A .; Ellegren, H .; Уэйн, Р. К. (1999). «Филогеография митохондриальной ДНК и популяционная история серого волка Canis lupus» (PDF). Молекулярная экология. 8 (12): 2089–103. Дои:10.1046 / j.1365-294x.1999.00825.x. HDL:10261/58565. PMID 10632860.
- ^ а б Ларсон Г., Брэдли Д. Г. (2014). «Сколько это в собачьих годах? Появление геномики собачьей популяции». PLOS Genetics. 10 (1): e1004093. Дои:10.1371 / journal.pgen.1004093. ЧВК 3894154. PMID 24453989.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ Мори, Дарси Ф .; Jeger, Rujana (2016). «От волка к собаке: экологическая динамика позднего плейстоцена, измененные трофические стратегии и смещение человеческого восприятия». Историческая биология. 29 (7): 1–9. Дои:10.1080/08912963.2016.1262854. S2CID 90588969.
- ^ а б Ван Валкенбург, Блэр; Хейворд, Мэтью В .; Ripple, Уильям Дж .; Мелоро, Карло; Рот, В. Луиза (2016). «Воздействие крупных наземных хищников на экосистемы плейстоцена». Труды Национальной академии наук. 113 (4): 862–867. Дои:10.1073 / pnas.1502554112. ЧВК 4743832. PMID 26504224.
- ^ Шипман, П. (2015). Захватчики: как люди и их собаки довели неандертальцев до исчезновения. Издательство Гарвардского университета. ISBN 978-0-674-73676-4.
- ^ Каган, Алекс; Бласс, Торстен (2016). «Идентификация геномных вариантов, предположительно нацеленных на отбор во время одомашнивания собак». BMC Эволюционная биология. 16: 10. Дои:10.1186 / s12862-015-0579-7. ЧВК 4710014. PMID 26754411.
- ^ Сун, Цзяо-Цзяо; Ван, Вэнь-Чжи; Otecko, Newton O .; Пэн, Мин-Шэн; Чжан, Я-Пин (2016). «Урегулирование конфликтов между деревьями гаплогруппы митохондриальной ДНК Canis lupus». Международная судебная экспертиза: генетика. 23: 83–85. Дои:10.1016 / j.fsigen.2016.03.008. PMID 27042801.
- ^ Пилот Малгожата; Греко, Клаудиа; Vonholdt, Bridgett M; Рэнди, Этторе; Енджеевский, Влодзимеж; Сидорович, Вадим Е; Конопинский, Мацей К; Острандер, Элейн А; Уэйн, Роберт К (2018). «Широко распространенное, долгосрочное смешение серых волков и домашних собак по всей Евразии и его последствия для сохранения статуса гибридов». Эволюционные приложения. 11 (5): 662–680. Дои:10.1111 / eva.12595. ЧВК 5978975. PMID 29875809.
- ^ Грейг, Карен; Букок, Джеймс; Прост, Стефан; Хорсбург, К. Энн; Джакомб, Крис; Уолтер, Ричард; Матисоо-Смит, Элизабет (2015). «Полные митохондриальные геномы первых собак Новой Зеландии». PLOS ONE. 10 (10): e0138536. Bibcode:2015PLoSO..1038536G. Дои:10.1371 / journal.pone.0138536. ЧВК 4596854. PMID 26444283.
- ^ Savolainen, P .; Leitner, T .; Wilton, A.N .; Matisoo-Smith, E .; Лундеберг, Дж. (2004). «Подробная картина происхождения австралийского динго, полученная в результате изучения митохондриальной ДНК». Труды Национальной академии наук. 101 (33): 12387–12390. Bibcode:2004ПНАС..10112387С. Дои:10.1073 / pnas.0401814101. ЧВК 514485. PMID 15299143.
- ^ а б Оскарссон, М. С. Р .; Klutsch, C.F.C .; Boonyaprakob, U .; Wilton, A .; Tanabe, Y .; Саволайнен, П. (2011). «Данные митохондриальной ДНК указывают на интродукцию австралийских динго и полинезийских домашних собак через материковую часть Юго-Восточной Азии». Труды Королевского общества B: биологические науки. 279 (1730): 967–974. Дои:10.1098 / rspb.2011.1395. ЧВК 3259930. PMID 21900326.
- ^ Ардалан, Арман; Оскарссон, Маттиас; Натанаэльссон, Кристиан; Уилтон, Алан Н .; Ахмадиан, Афшин; Саволайнен, Питер (2012). «Узкая генетическая основа австралийского динго подтверждена анализом отцовского происхождения». Genetica. 140 (1–3): 65–73. Дои:10.1007 / s10709-012-9658-5. ЧВК 3386486. PMID 22618967.
- ^ а б Гросс, Майкл (2015). "Собаки такие же, как мы?". Текущая биология. 25 (17): R733–6. Дои:10.1016 / j.cub.2015.08.018. PMID 26561653.
- ^ «Древний геном волка отодвигает собачий рассвет». Природа. 2015.
- ^ Machugh, David E .; Ларсон, Грегер; Орландо, Людовик (2016). «Укрощение прошлого: древняя ДНК и исследование приручения животных». Ежегодный обзор биологических наук о животных. 5: 329–351. Дои:10.1146 / annurev-animal-022516-022747. PMID 27813680.
- ^ Эрсмарк, Э (2016). «Обновление популяции крупных хищников и экологические изменения в позднем четвертичном периоде». Стокгольмский университет. Кандидатская диссертация под руководством Дж. Леонарда и Л. Далена
- ^ а б c Ли, Э. (2015). «Анализ древней ДНК старейших видов псовых из сибирской Арктики и генетический вклад в развитие домашних собак». PLOS ONE. 10 (5): e0125759. Bibcode:2015PLoSO..1025759L. Дои:10.1371 / journal.pone.0125759. ЧВК 4446326. PMID 26018528.
- ^ Irizarry, Kristopher J. L .; Васконселос, Элтон Дж. Р. (2018). «Популяционная геномика одомашнивания и развития пород у собак в контексте когнитивных, социальных, поведенческих и болезненных черт». В Раджоре, О. (ред.). Популяционная геномика. С. 755–806. Дои:10.1007/13836_2018_43. ISBN 978-3-030-04587-6.
- ^ Бохеренс, Эрве; Drucker, Dorothée G .; Бонжан, Доминик; Бридо, Энн; Конард, Николас Дж .; Купиллард, Кристоф; Жермонпре, Митье; Хёнейзен, Маркус; Münzel, Susanne C .; Напьерала, Ханнес; Пату-Матис, Мэрилен; Стефан, Элизабет; Уерпманн, Ханс-Петер; Циглер, Рейнхард (2011). «Изотопные данные по экологии питания пещерного льва (Panthera spelaea) в Северо-Западной Европе: выбор добычи, конкуренция и последствия для исчезновения». Четвертичный международный. 245 (2): 249–261. Bibcode:2011QuInt.245..249B. Дои:10.1016 / j.quaint.2011.02.023.
- ^ Yeakel, J.D .; Guimaraes, P. R .; Bocherens, H .; Кох, П. Л. (2013). «Влияние изменения климата на структуру трофических сетей плейстоцена в мамонтовой степи». Труды Королевского общества B: биологические науки. 280 (1762): 20130239. Дои:10.1098 / rspb.2013.0239. ЧВК 3673045. PMID 23658198.
- ^ а б Бохеренс, Эрве (2015). «Изотопное отслеживание палеоэкологии крупных хищников в мамонтовой степи». Четвертичные научные обзоры. 117: 42–71. Дои:10.1016 / j.quascirev.2015.03.018.
- ^ Dinnis, R .; Pate, A .; Рейнольдс, Н. (2016). "Средне-поздние морские изотопы фауны млекопитающих Британии: новый взгляд". Труды ассоциации геологов. 127 (4): 435–444. Дои:10.1016 / j.pgeola.2016.05.002.
- ^ Бойтани, Л. (2003). «Сохранение и восстановление волков» в книге «Волки: экология поведения и сохранение», ред. Л.Д. Мех и Л. Бойтани (Чикаго, Иллинойс: издательство Chicago University Press), 317–340. Дои:10.7208 / Чикаго / 9780226516981.001.0001
- ^ Леонард, Дженнифер А .; Вила, Карлес; Уэйн, Роберт К. (2004).«FAST TRACK: Наследие утрачено: генетическая изменчивость и размер популяций искорененных серых волков США (Canis lupus)». Молекулярная экология. 14 (1): 9–17. Дои:10.1111 / j.1365-294X.2004.02389.x. PMID 15643947.
- ^ Vonholdt, Bridgett M .; Pollinger, John P .; Earl, Dent A .; Ноулз, Джеймс С.; Бойко, Адам Р .; Паркер, Хайди; Геффен, Эли; Пилот Малгожата; Енджеевский, Влодзимеж; Енжеевска, Богумила; Сидорович, Вадим; Греко, Клаудиа; Рэнди, Этторе; Мусиани, Марко; Кейс, Роланд; Bustamante, Carlos D .; Острандер, Элейн А .; Новембре, Джон; Уэйн, Роберт К. (2011). «Полногеномный взгляд на эволюционную историю загадочных волкоподобных псовых». Геномные исследования. 21 (8): 1294–305. Дои:10.1101 / гр.116301.110. ЧВК 3149496. PMID 21566151.
- ^ Vonholdt, Bridgett M .; Pollinger, John P .; Earl, Dent A .; Паркер, Хайди Дж .; Острандер, Элейн А .; Уэйн, Роберт К. (2012). «Идентификация недавней гибридизации серых волков и домашних собак с помощью генотипирования SNP». Геном млекопитающих. 24 (1–2): 80–8. Дои:10.1007 / s00335-012-9432-0. PMID 23064780. S2CID 1161310.
- ^ Эрнст Майр (1999). "VIII-Негеографическое видообразование". Систематика и происхождение видов с точки зрения зоолога. Издательство Гарвардского университета. С. 194–195. ISBN 9780674862500.
- ^ Vila, C .; Sundqvist, A.-K .; Flagstad, O .; Seddon, J .; Rnerfeldt, S. B .; Kojola, I .; Casulli, A .; Sand, H .; Wabakken, P .; Эллегрен, Х. (2003). «Спасение популяции волка (Canis lupus) с серьезным ограничением возможностей одним иммигрантом». Труды Королевского общества B: биологические науки. 270 (1510): 91–97. Дои:10.1098 / rspb.2002.2184. ЧВК 1691214. PMID 12590776.
- ^ а б c Муньос-Фуэнтес, Виолетта; Даримонт, Крис Т .; Уэйн, Роберт К .; Paquet, Paul C .; Леонард, Дженнифер А. (2009). «Экологические факторы приводят к дифференциации волков из Британской Колумбии». Журнал биогеографии. 36 (8): 1516–1531. Дои:10.1111 / j.1365-2699.2008.02067.x. HDL:10261/61670.
- ^ Вальер, Натаниэль; Фумагалли, Лука; Гилли, Людовик; Микель, Кристиан; Лекетт, Бенуа; Пуль, Мари-Лазарин; Вебер, Жан-Марк; Арлеттаз, Рафаэль; Таберле, Пьер (2003). «Дальнейшая реколонизация волков во Франции и Швейцарии по результатам неинвазивной генетической выборки в течение 10 лет». Сохранение животных. 6: 83–92. Дои:10.1017 / S1367943003003111.
- ^ Fabbri, E .; Caniglia, R .; Kusak, J .; Галов, А .; Gomerčić, T .; Arbanasić, H .; Huber, D .; Рэнди, Э. (2014). «Генетическая структура растущих популяций волков (Canis lupus) в Италии и Хорватии и первые шаги реколонизации Восточных Альп». Биология млекопитающих - Zeitschrift für Säugetierkunde. 79 (2): 138–148. Дои:10.1016 / j.mambio.2013.10.002.
- ^ Стронен, Астрид V .; Jä ™ Drzejewska, Bogumiå'a; Pertoldi, Cino; Демонтис, Дитте; Рэнди, Этторе; Недзяа'Ковска, Магдалена; Пилот, Ма'Горзата; Сидорович, Вадим Е .; Дикий, Игорь; Кусак, Иосип; Цингарская, Елена; Коджола, Илпо; Karamanlidis, Alexandros A .; Орниканс, Айварс; Лобков, Владимир А .; Думенко, Виталий; Чарномская, Сильвия Д. (2013). «Дифференциация между Севером и Югом и область высокого разнообразия европейских волков (Canis lupus)». PLOS ONE. 8 (10): e76454. Дои:10.1371 / journal.pone.0076454. ЧВК 3795770. PMID 24146871.
- ^ а б Schweizer, Rena M .; Робинсон, Жаклин; Харриган, Райан; Сильва, Педро; Галверни, Марко; Мусиани, Марко; Грин, Ричард Э .; Новембре, Джон; Уэйн, Роберт К. (2016). «Целенаправленный захват и повторное секвенирование 1040 генов выявляют функциональные вариации у серых волков, обусловленные окружающей средой». Молекулярная экология. 25 (1): 357–79. Дои:10.1111 / mec.13467. PMID 26562361.
- ^ О'Киф, Ф. Робин; Мичен, Джули; Fet, Elizabeth V .; Бранник, Александрия (2013). «Экологические детерминанты клинальной морфологической изменчивости черепа североамериканского серого волка». Журнал маммологии. 94 (6): 1223–1236. Дои:10.1644 / 13-MAMM-A-069.
- ^ Кармайкл, Л. Э .; Krizan, J .; Nagy, J. A .; Fuglei, E .; Dumond, M .; Johnson, D .; Veitch, A .; Berteaux, D .; Стробек, К. (2007). «Исторические и экологические детерминанты генетической структуры арктических псовых». Молекулярная экология. 16 (16): 3466–83. Дои:10.1111 / j.1365-294X.2007.03381.x. PMID 17688546.
- ^ Кармайкл, Л. Э .; Nagy, J. A .; Larter, N.C .; Стробек, К. (2001). «Специализация добычи может влиять на структуру потока генов у волков Северо-Запада Канады». Молекулярная экология. 10 (12): 2787–98. Дои:10.1046 / j.0962-1083.2001.01408.x. PMID 11903892.
- ^ Кармайкл, Л. Э. (2006). Экологическая генетика северных волков и песцов. Кандидат наук. Диссертация (PDF). Университет Альберты.
- ^ Lacombat, F .; Abbazzi, L .; Ferretti, M.P .; Martinez-Navarro, B .; Moulle, P.-E .; Palombo, M.-R .; Ладья, Л .; Тернер, А .; Валли, А.М.-Ф. (2008). «Новые данные о ранней виллафранкской фауне из Виалетт (Верхняя Луара, Франция) на основе коллекции Музея Крозатье (Ле-Пюи-ан-Веле, Верхняя Луара, Франция)». Четвертичный международный. 179 (1): 64–71. Дои:10.1016 / j.quaint.2007.09.005.
- ^ Ладья, Л .; Торре, Д. (1996). «Волк-событие в Западной Европе и начало позднего Виллафранка». Neues Jahrbuch für Geologie und Paläontologie - Monatshefte. 1996 (8): 495–501. Дои:10.1127 / njgpm / 1996/1996/495.
- ^ Gliozzi, E .; Abbazzi, L .; Argenti, A .; Azzaroli, A .; Caloi, L .; Capasso Barbato, L .; ди Стефано, G .; Esu, D .; Ficcarelli, G .; Girotti, O .; Kotsakis, T .; Masini, F .; Mazza, P .; Mezzabotta, C .; Palombo, M. R .; Petronio, C .; Ладья, Л .; Сала, Б .; Sardella, R .; Zanalda, E .; Торре, Д. (1997). «Биохронология отдельных млекопитающих, моллюсков и остракод от среднего плиоцена до позднего плейстоцена в Италии. Современное состояние» (PDF). Итальянская Ривиста ди Палеонтология и Стратиграфия. 103 (3): 369–388. Дои:10.13130/2039-4942/5299.
- ^ Смородина, А. П. (1984). «Остатки млекопитающих». В Грин, Х.С. (ред.). Пещера Понтневидд: местонахождение гоминидов нижнего палеолита в Уэльсе: первое сообщение. Журнал археологической науки. 12. Amgueddfa Genedlaethol Cymru. С. 177–181. Дои:10.1016 / 0305-4403 (85) 90038-Х. ISBN 978-0-7200-0282-9.
- ^ Фокс-Доббс, Кена; Леонард, Дженнифер А .; Кох, Пол Л. (2008). «Плейстоценовая мегафауна восточной Берингии: палеоэкологические и палеоэкологические интерпретации стабильных изотопов углерода и азота и радиоуглеродных записей». Палеогеография, палеоклиматология, палеоэкология. 261 (1–2): 30–46. Дои:10.1016 / j.palaeo.2007.12.011.
- ^ Барышников, Геннадий Ф .; Мол, Дик; Тихонов, Алексей Н (2009). «Обнаружение позднеплейстоценовых хищников на Таймыре (Россия, Сибирь) с палеоэкологическим контекстом» (PDF). Российский журнал териологии. 8 (2): 107–113. Дои:10.15298 / rusjtheriol.08.2.04. Получено 23 декабря, 2014.
- ^ Оводов, Н. (2011). «Зарождающаяся собака возрастом 33 000 лет из Горного Алтая в Сибири: свидетельство самого раннего приручения, нарушенного Последним ледниковым максимумом». PLOS ONE. 6 (7): e22821. Bibcode:2011PLoSO ... 622821O. Дои:10.1371 / journal.pone.0022821. ЧВК 3145761. PMID 21829526.
- ^ а б Жермонпре, Митье; Лазничкова-Галетова, Мартина; Лоузи, Роберт Дж .; Райкконен, Янникке; Саблин, Михаил В. (2015). «Крупные псовые на стоянке Gravettian Předmostí, Чехия: нижняя челюсть». Четвертичный международный. 359–360: 261–279. Дои:10.1016 / j.quaint.2014.07.012.
- ^ Жермонпре, Митье; Лазничкова-Галетова, Мартина; Саблин, Михаил В. (2012). "Палеолитические черепа собак на могиле Граветтианских Предмостей, Чехия". Журнал археологической науки. 39: 184–202. Дои:10.1016 / j.jas.2011.09.022.
- ^ Будади-Малинь, М. (2010). "Les Canis Pleistocenes du Sud de la France: подход к биосистематике, эволюции и биохронологии. Докторская диссертация" (PDF). Archéologie et Préhistoire. Университет Бордо (4126).
- ^ Dimitrijević, V .; Вукович, С. (2015). «Была ли собака локально одомашнена в Дунайских ущельях? Морфометрическое исследование черепных останков собак из четырех археологических памятников эпохи мезолита и раннего неолита по сравнению с современными волками». Международный журнал остеоархеологии. 25: 1–30. Дои:10.1002 / oa.2260.
- ^ Жермонпреа, Митье; Саблинб, Михаил В .; Stevensc, Rhiannon E .; Hedgesd, Роберт Э. М .; Хофрайтере, Майкл; Стиллере, Матиас; Деспрез, Вивиан Р. (2008). «Ископаемые собаки и волки из палеолитических памятников Бельгии, Украины и России: остеометрия, древняя ДНК и стабильные изотопы». Журнал археологической науки. 36 (2): 473–490. Дои:10.1016 / j.jas.2008.09.033.
- ^ Ларсон Дж. (2012). "Переосмысление приручения собак путем интеграции генетики, археологии и биогеографии". PNAS. 109 (23): 8878–8883. Bibcode:2012PNAS..109.8878L. Дои:10.1073 / пнас.1203005109. ЧВК 3384140. PMID 22615366.
| Соответствующие поля | ||
|---|---|---|
| Базовые концепты | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||
| ||
| Ключевые компоненты | |
|---|---|
| Поля | |
| Археогенетика из | |
| похожие темы | |
| Списки | |
| |
