Рибосома - Ribosome
| Клеточная биология | |
|---|---|
| В животная клетка | |
 Компоненты типичной животной клетки:
|

Рибосомы (/ˈраɪбəˌsoʊм,-бoʊ-/[1]) находятся макромолекулярные машины, найденный во всех живых клетки, которые выполняют биологический синтез белка (трансляция мРНК). Связь рибосом аминокислоты вместе в порядке, установленном кодоны из информационная РНК (мРНК) молекулы для образования полипептид цепи. Рибосомы состоят из двух основных компонентов: малых и больших рибосомных субъединиц. Каждая субъединица состоит из одного или нескольких рибосомная РНК (рРНК) молекулы и многие рибосомальные белки (RP или r-белки).[2][3][4] Рибосомы и связанные с ними молекулы также известны как переводной аппарат.
Обзор
Последовательность ДНК который кодирует последовательность аминокислот в белке, транскрибируется в цепь информационной РНК. Рибосомы связываются с матричной РНК и используют ее последовательность для определения правильной последовательности аминокислот для генерации данного белка. Аминокислоты отбираются и переносятся на рибосому посредством Переносная РНК (тРНК) молекулы, которые входят в рибосому и связываются с цепью информационной РНК через антикодон стеблевая петля. Для каждой кодовой тройки (кодон ) в информационной РНК есть передающая РНК, которая соответствует и несет правильную аминокислоту для включения в растущий полипептид цепь. Как только белок произведен, он может складывать для создания функциональной трехмерной конструкции.
Рибосома состоит из комплексы РНК и белков и поэтому рибонуклеопротеидный комплекс. Каждая рибосома состоит из маленьких (30S) и больших компонентов (50S), называемых субъединицами, которые связаны друг с другом:
- (30S) в основном выполняет функцию декодирования, а также связан с мРНК.
- (50S) выполняет в основном каталитическую функцию; и также связывается с аминоацилированными тРНК.
Синтез белков из их строительных блоков происходит в четыре фазы: инициация, удлинение, завершение и повторное использование. Стартовый кодон во всех молекулах мРНК имеет последовательность AUG. Стоп-кодон может быть одним из UAA, UAG или UGA; нет молекул тРНК, распознающих эти кодоны, поэтому рибосома распознает, что трансляция завершена.[5] Когда рибосома заканчивает считывание молекулы мРНК, две субъединицы разделяются и обычно разбиваются, но могут использоваться повторно. Рибосомы рибозимы, поскольку каталитический пептидилтрансфераза активность, которая связывает аминокислоты вместе, осуществляется рибосомной РНК. Рибосомы часто связаны с внутриклеточными мембранами, составляющими шероховатой эндоплазматической сети.
Рибосомы из бактерии, археи и эукариоты в трехдоменная система похожи друг на друга в значительной степени, что свидетельствует об общем происхождении. Они различаются по размеру, последовательности, структуре и соотношению белка к РНК. Различия в структуре позволяют антибиотики убивать бактерии, подавляя их рибосомы, не затрагивая человеческие рибосомы. У всех видов более одной рибосомы могут перемещаться по одной цепи мРНК одновременно (как полисом ), каждый из которых «читает» определенную последовательность и производит соответствующую молекулу белка.
В митохондриальные рибосомы Эукариотические клетки функционально напоминают многие особенности таковых у бактерий, что отражает вероятное эволюционное происхождение митохондрий.[6][7]
Открытие
Рибосомы были впервые обнаружены в середине 1950-х гг. Румынско-американский клеточный биолог Джордж Эмиль Паладе, используя электронный микроскоп, в виде плотных частиц или гранул.[8] Термин «рибосома» был предложен ученым Ричардом Б. Робертсом в конце 1950-х годов:
В ходе симпозиума стала очевидной смысловая трудность. Для некоторых участников «микросомы» означают частицы рибонуклеопротеина микросомной фракции, загрязненные другим белком и липидным материалом; для других микросомы состоят из белков и липидов, загрязненных частицами. Фраза «микросомальные частицы» не кажется адекватной, а «рибонуклеопротеиновые частицы микросомной фракции» слишком неудобны. Во время встречи было предложено слово «рибосома», имеющее очень удачное название и приятное звучание. Настоящая путаница была бы устранена, если бы термин «рибосома» был принят для обозначения рибонуклеопротеиновых частиц размером от 35 до 100S.
— Альберт, Микросомальные частицы и синтез белка[9]
Альбер Клод, Кристиан де Дюв, и Джордж Эмиль Паладе были совместно награждены Нобелевская премия по физиологии и медицине в 1974 г. за открытие рибосомы.[10] В Нобелевская премия в Химия 2009 был награжден Венкатраман Рамакришнан, Томас А. Стейтц и Ада Э. Йонат для определения подробной структуры и механизма рибосомы.[11]
Структура

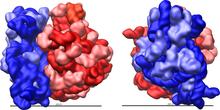
Рибосома - очень сложная клеточная машина. Он в основном состоит из специализированных РНК, известных как рибосомная РНК (рРНК), а также десятки различных белков (точное количество незначительно варьируется между видами). Рибосомные белки и рРНК организованы в две отдельные рибосомные части разного размера, обычно известные как большая и малая субъединица рибосомы. Рибосомы состоят из двух субъединиц, которые подходят друг к другу (рис. 2) и работают как одна, транслируя мРНК в полипептидную цепь во время синтеза белка (рис. 1). Поскольку они состоят из двух субъединиц неравного размера, они немного длиннее по оси, чем по диаметру.
Бактериальные рибосомы
Прокариотических рибосом около 20нм (200 Å ) в диаметре и состоят из 65% рРНК и 35% рибосомальные белки.[12] Рибосомы эукариот составляют от 25 до 30 нм (250–300 Å) в диаметре с отношением рРНК к белку, близким к 1.[13] Кристаллографический работай [14] показал, что рядом с местом реакции синтеза полипептидов рибосомные белки отсутствуют. Это говорит о том, что белковые компоненты рибосом не участвуют напрямую в катализе образования пептидных связей, а скорее, что эти белки действуют как каркас, который может повышать способность рРНК синтезировать белок (см.: Рибозим ).
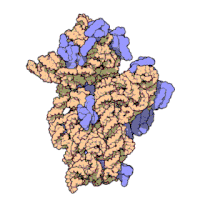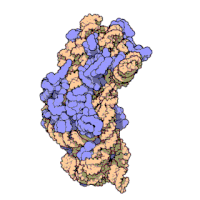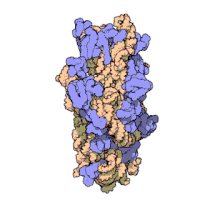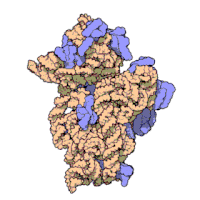
Рибосомные субъединицы бактерии и эукариоты очень похожи.[16]
Единицей измерения, используемой для описания рибосомных субъединиц и фрагментов рРНК, является Сведберг единица, мера скорости осаждение в центрифугирование а не размер. Это объясняет, почему имена фрагментов не складываются: например, бактериальные 70S рибосомы состоят из 50S и 30S субъединиц.
Бактерии 70S рибосомы, каждая из которых состоит из небольшого (30S ) и большой (50S ) субъединица. Кишечная палочка, например, имеет 16S Субъединица РНК (состоящая из 1540 нуклеотидов), связанная с 21 белком. Большая субъединица состоит из 5S Субъединица РНК (120 нуклеотидов), субъединица 23S РНК (2900 нуклеотидов) и 31 белки.[16]
Рибосома Кишечная палочка (бактерия)[17]:962 рибосома подразделение рРНК р-белки 70S 50S 23S (2904 нт ) 31 5S (120 нт) 30S 16S (1542 нт) 21
Аффинная метка для сайтов связывания тРНК на Кишечная палочка рибосома позволила идентифицировать белки сайтов A и P, наиболее вероятно связанные с пептидилтрансферазной активностью; меченые белки - L27, L14, L15, L16, L2; по крайней мере, L27 находится на донорском участке, как показали E. Collatz и A.P. Czernilofsky.[18][19] Дополнительные исследования показали, что белки S1 и S21 в ассоциации с 3'-концом 16S рибосомной РНК участвуют в инициации трансляции.[20]
Эукариотические рибосомы
У эукариот в цитозоле расположены 80S рибосомы, каждая из которых состоит из маленький (40S) и большая (60S) субъединица. Их субъединица 40S имеет 18S РНК (1900 нуклеотидов) и 33 белка.[21][22] Большая субъединица состоит из 5S РНК (120 нуклеотидов), 28S РНК (4700 нуклеотидов), а 5.8S РНК (160 нуклеотидов) субъединиц и 46 белков.[16][21][23]
эукариотические цитозольные рибосомы (Р. norvegicus)[17]:65 рибосома подразделение рРНК р-белки 80-е годы 60S 28S (4718 нт) 49 5.8S (160 нт) 5S (120 нт) 40S 18S (1874 г.) 33
В 1977 году Черниловский опубликовал исследование, в котором маркировка близости для определения сайтов связывания тРНК на рибосомах печени крысы. Некоторые белки, включая L32 / 33, L36, L21, L23, L28 / 29 и L13, были вовлечены как находящиеся на уровне или рядом с ним. пептидилтрансфераза центр.[24]
Пласторибосомы и миторибосомы
У эукариот рибосомы присутствуют в митохондрии (иногда называют миторибосомы ) И в пластиды Такие как хлоропласты (также называемые пласторибосомами). Они также состоят из больших и малых субъединиц, связанных вместе белки в одну частицу 70S.[16] Эти рибосомы похожи на рибосомы бактерий, и считается, что эти органеллы возникли как симбиотический бактерии[16] Из этих двух хлоропластные рибосомы ближе к бактериальным, чем митохрондриальные. Многие части рибосомной РНК в митохрондриях укорачиваются, а в случае 5S рРНК, заменены другими структурами у животных и грибов.[25] Особенно, Лейшмания тарантолы имеет минимизированный набор митохондриальной рРНК.[26]
В криптомонада и хлорарахниофит водоросли могут содержать нуклеоморф что напоминает рудиментарное ядро эукариот.[27] Рибосомы 80S эукариот могут присутствовать в компартменте, содержащем нуклеоморф.[нужна цитата ]
Используя различия
Различия между бактериальными и эукариотическими рибосомами используются химики-фармацевты создавать антибиотики которые могут уничтожить бактериальную инфекцию, не нанося вреда клеткам инфицированного человека. Из-за различий в структуре бактериальные рибосомы 70S уязвимы для этих антибиотиков, в то время как рибосомы 80S эукариот - нет.[28] Хотя митохондрии обладают рибосомами, подобными бактериальным, митохондрии не подвержены действию этих антибиотиков, потому что они окружены двойной мембраной, которая не пропускает эти антибиотики легко. органелла.[29] Однако заслуживающий внимания контрпример включает противоопухолевый антибиотик хлорамфеникол, который успешно ингибирует бактериальные 50S и митохондриальные 50S рибосомы.[30] То же самое о митохондриях нельзя сказать о хлоропластах, где устойчивость рибосомных белков к антибиотикам является признаком, который следует ввести в качестве маркера в генную инженерию.[31]
Общие свойства
Различные рибосомы имеют общую структуру ядра, которая очень похожа, несмотря на большие различия в размере. Большая часть РНК высокоорганизована в различные третичные структурные мотивы, Например псевдоузлы эта выставка коаксиальная укладка. Дополнительный РНК в более крупных рибосомах находится в нескольких длинных непрерывных вставках [32], так что они образуют петли из основной структуры, не нарушая и не изменяя ее.[16] Вся каталитическая активность рибосомы осуществляется РНК; белки находятся на поверхности и, кажется, стабилизируют структуру.[16]
Структура высокого разрешения

Общая молекулярная структура рибосомы известна с начала 1970-х годов. В начале 2000-х годов структура достигла высокого разрешения, порядка нескольких Ангстремс.
Первые статьи, описывающие структуру рибосомы с атомным разрешением, были опубликованы почти одновременно в конце 2000 года. 50S (большая прокариотическая) субъединица была определена из Археон Haloarcula marismortui[33] и бактерия Дейнококк радиодуранс,[34] а структура 30S субъединицы была определена из Термус термофильный.[15] Эти структурные исследования были удостоены Нобелевской премии по химии в 2009 году. В мае 2001 года эти координаты были использованы для реконструкции всего T. thermophilus 70S частица при 5,5Å разрешающая способность.[35]
Две статьи были опубликованы в ноябре 2005 г. структурами кишечная палочка 70S рибосома. Структуры вакантной рибосомы определяли при 3,5Å разрешение с использованием Рентгеновская кристаллография.[36] Затем, две недели спустя, структура на основе криоэлектронная микроскопия был опубликован,[37] что изображает рибосому на уровне 11–15Å разрешение в процессе прохождения вновь синтезированной белковой цепи в белок-проводящий канал.
Первые атомные структуры рибосомы в комплексе с тРНК и мРНК молекулы были решены методом рентгеновской кристаллографии двумя группами независимо друг от друга при 2,8Å[38] и 3,7Å.[39] Эти структуры позволяют увидеть детали взаимодействия Термус термофильный рибосома с мРНК и с тРНК связаны на классических рибосомальных участках. Взаимодействие рибосомы с длинными мРНК, содержащими Последовательности Шайна-Далгарно были визуализированы вскоре после этого на 4.5–5.5Å разрешающая способность.[40]
В 2011 году была получена первая полная атомная структура эукариотической 80S рибосомы дрожжей. Saccharomyces cerevisiae был получен кристаллографией.[21] Модель раскрывает архитектуру эукариот-специфичных элементов и их взаимодействие с универсально сохраняемым ядром. В то же время полная модель эукариотической рибосомной структуры 40S в Tetrahymena thermophila была опубликована и описана структура Субъединица 40S, а также многое о взаимодействии субъединицы 40S с eIF1 в течение инициирование перевода.[22] Точно так же эукариотический Субъединица 60S структура была также определена из Tetrahymena thermophila в комплексе с eIF6.[23]
Функция
Рибосомы - это мельчайшие частицы, состоящие из РНК и связанных белков, которые функционируют для синтеза белков. Белки необходимы для многих клеточных функций, таких как восстановление повреждений или управление химическими процессами. Рибосомы можно обнаружить плавающими в цитоплазме или прикрепленными к эндоплазматический ретикулум. По сути, их основная функция - преобразование генетического кода в аминокислотную последовательность и построение белковых полимеров из мономеров аминокислот.
Рибосомы действуют как катализаторы в двух чрезвычайно важных биологических процессах, называемых переносом пептидила и гидролизом пептидила.[41] «Центр PT отвечает за образование белковых связей во время удлинения белка».[41]
Перевод
Рибосомы - это рабочие места биосинтез белка, процесс перевода мРНК в белок. МРНК состоит из ряда кодоны которые расшифровываются рибосомой, чтобы произвести белок. Используя мРНК в качестве матрицы, рибосома пересекает каждый кодон (3 нуклеотиды ) мРНК, спаривая ее с соответствующей аминокислотой, предоставленной аминоацил-тРНК. Аминоацил-тРНК содержит комплементарный антикодон на одном конце и соответствующая аминокислота на другом. Для быстрого и точного распознавания соответствующей тРНК рибосома использует большие конформационные изменения (конформационная корректура ).[42] Маленькая рибосомная субъединица, обычно связанная с аминоацил-тРНК, содержащей первую аминокислоту метионин, связывается с кодоном AUG на мРНК и рекрутирует большую субъединицу рибосомы. Рибосома содержит три сайта связывания РНК, обозначенных A, P и E. Сайт связывает аминоацил-тРНК или факторы терминации высвобождения;[43][44] то P-сайт связывает пептидил-тРНК (тРНК, связанную с полипептидной цепью); и Электронный сайт (выход) связывает свободную тРНК. Синтез белка начинается с стартовый кодон AUG около 5 'конца мРНК. мРНК сначала связывается с сайтом Р рибосомы. Рибосома распознает стартовый кодон с помощью Последовательность Шайна-Далгарно мРНК прокариот и Козак ящик у эукариот.
Хотя катализ пептидная связь включает C2 гидроксил Р-сайта РНК аденозин В механизме протонного челнока другие этапы синтеза белка (например, транслокация) вызываются изменениями конформации белка. Поскольку их каталитическое ядро состоит из РНК, рибосомы классифицируются как «рибозимы,"[45] и считается, что они могут быть остатками Мир РНК.[46]

На рисунке 5 обе рибосомные субъединицы (маленький и большой) собрать в стартовом кодоне (ближе к 5 'концу мРНК ). Рибосома использует тРНК который соответствует текущему кодону (триплету) на мРНК, чтобы добавить аминокислота к полипептидной цепи. Это делается для каждого триплета мРНК, в то время как рибосома перемещается к 3'-концу мРНК. Обычно в бактериальных клетках несколько рибосом работают параллельно над одной мРНК, образуя то, что называется полирибосома или же полисом.
Котрансляционное складывание
Известно, что рибосома активно участвует в сворачивание белка.[47][48] Структуры, полученные таким образом, обычно идентичны структурам, полученным во время химического рефолдинга белка, однако пути, ведущие к конечному продукту, могут быть разными.[49][50] В некоторых случаях рибосома имеет решающее значение для получения функциональной формы белка. Например, один из возможных механизмов складывания глубоко узловатые белки полагается на то, что рибосома проталкивает цепь через прикрепленную петлю.[51]
Добавление трансляционно-независимых аминокислот
Присутствие белка контроля качества рибосом Rqc2 связано с мРНК-независимым удлинением белка.[52][53] Это удлинение является результатом рибосомного добавления (через тРНК, переносимые Rqc2) КОТ хвосты: рибосомы расширяют C-терминал застопорившегося белка со случайными, независимыми от трансляции последовательностями алинии и тхреонины.[54][55]
Расположение рибосом
Рибосомы классифицируются как «свободные» или «мембраносвязанные».
Свободные и связанные с мембраной рибосомы различаются только своим пространственным распределением; они идентичны по структуре. Независимо от того, существует ли рибосома в свободном или связанном с мембраной состоянии, зависит от наличия Сигнальная последовательность ER-нацеливания на синтезируемом белке, поэтому отдельная рибосома может быть связана с мембраной, когда она производит один белок, но свободна в цитозоле, когда она производит другой белок.
Рибосомы иногда называют органеллы, но использование термина органелла часто ограничивается описанием субклеточных компонентов, которые включают фосфолипидную мембрану, чего не делают рибосомы, будучи полностью частицами. По этой причине рибосомы иногда называют «немембранными органеллами».
Свободные рибосомы
Свободные рибосомы могут перемещаться в любом месте цитозоль, но исключены из ядро клетки и другие органеллы. Белки, образованные из свободных рибосом, попадают в цитозоль и используются внутри клетки. Поскольку цитозоль содержит высокие концентрации глутатион и, следовательно, является уменьшение окружающей среды, белки, содержащие дисульфидные связи, которые образуются из окисленных остатков цистеина, не могут образовываться в нем.
Рибосомы, связанные с мембраной
Когда рибосома начинает синтезировать белки, которые необходимы некоторым органеллам, рибосома, производящая этот белок, может стать «связанной с мембраной». В эукариотических клетках это происходит в области эндоплазматического ретикулума (ER), называемой «грубым ER». Вновь полученные полипептидные цепи вставляются непосредственно в ЭР рибосомным предпринимателем. векторный синтез а затем доставляются к месту назначения через секреторный путь. Связанные рибосомы обычно производят белки, которые используются в плазматической мембране или выводятся из клетки через экзоцитоз.[56]
Биогенез
В бактериальных клетках рибосомы синтезируются в цитоплазме через транскрипция гена множественных рибосом опероны. У эукариот этот процесс происходит как в цитоплазме клетки, так и в ядрышко, который является регионом внутри ядро клетки. Процесс сборки включает скоординированную функцию более 200 белков в синтезе и процессинге четырех рРНК, а также сборку этих рРНК с рибосомными белками.
Источник
Рибосома, возможно, впервые возникла в Мир РНК, представляющий собой самовоспроизводящийся комплекс, который только позже развил способность синтезировать белки, когда аминокислоты начали появляться.[57] Исследования показывают, что древние рибосомы, построенные исключительно из рРНК мог развить способность синтезировать пептидные связи.[58][59][60] Кроме того, данные убедительно указывают на древние рибосомы как на самовоспроизводящиеся комплексы, в которых рРНК в рибосомах имела информационные, структурные и каталитические цели, поскольку могла кодировать тРНК и белки, необходимые для саморепликации рибосом.[61] Гипотетические клеточные организмы с самовоспроизводящейся РНК, но без ДНК называются рибоцитами (или рибоцеллами).[62][63]
Поскольку аминокислоты постепенно появлялись в мире РНК в пребиотических условиях,[64][65] их взаимодействия с каталитической РНК увеличивают как диапазон, так и эффективность функции молекул каталитической РНК.[57] Таким образом, движущая сила эволюции рибосомы из древнего самовоспроизводящаяся машина в его нынешнюю форму в качестве трансляционной машины, возможно, оказывалось давление отбора, направленное на включение белков в механизмы саморепликации рибосомы, чтобы увеличить ее способность к саморепликации.[61][66][67]
Гетерогенные рибосомы
Рибосомы неоднородны по составу между видами и даже внутри одной клетки, о чем свидетельствует существование цитоплазматических и митохондриальных рибосом в одних и тех же эукариотических клетках. Некоторые исследователи предположили, что гетерогенность в составе рибосомных белков у млекопитающих важна для регуляции генов, т.е., специализированная гипотеза рибосом.[68][69] Однако эта гипотеза является спорной и тема текущих исследований.[70][71]
Неоднородность в составе рибосом впервые была предложена Винсом Мауро для трансляционного контроля синтеза белка. Джеральд Эдельман.[72] Они предложили гипотезу рибосомного фильтра для объяснения регуляторных функций рибосом. Данные свидетельствуют о том, что специализированные рибосомы, специфичные для разных популяций клеток, могут влиять на то, как транслируются гены.[73] Некоторые рибосомальные белки обмениваются из собранного комплекса с цитозольный копии [74] предполагая, что структура in vivo рибосома может быть изменена без синтеза новой рибосомы.
Некоторые рибосомальные белки абсолютно необходимы для жизни клетки, а другие нет. В бутоньерки, 14/78 рибосомные белки не являются необходимыми для роста, тогда как у людей это зависит от исследуемой клетки.[75] Другие формы гетерогенности включают посттрансляционные модификации рибосомных белков, такие как ацетилирование, метилирование и фосфорилирование.[76] Арабидопсис,[77][78][79][80] Популярный внутренние сайты входа в рибосомы (IRES) может опосредовать переводы разными по составу рибосомами. Например, рибосомные единицы 40S без eS25 в клетках дрожжей и млекопитающих неспособны привлекать CrPV IGR IRES.[81]
Гетерогенность модификаций рибосомной РНК играет важную роль в поддержании структуры и / или функции, и большинство модификаций мРНК обнаруживаются в высококонсервативных областях.[82][83] Наиболее распространенные модификации рРНК: псевдоуридилирование и 2’-O метилирование рибозы.[84]
Смотрите также
- Аминогликозиды
- Биологические машины
- Эукариотический перевод
- Посттрансляционная модификация
- Прокариотический перевод
- Белковая динамика
- Третичная структура РНК
- Перевод (генетика)
- Базовая пара колебаний
- Ада Йонат - Израильский кристаллограф, известный своей новаторской работой по структуре рибосомы, за которую она получила награду Нобелевская премия.
Рекомендации
- ^ Джонс Д., Хартман Дж., Роуч П., Сеттер Дж. (2003) [1917]. Словарь английского произношения. Кембридж: Издательство Кембриджского университета. ISBN 978-3-12-539683-8.
- ^ Konikkat S (февраль 2016 г.). События динамического ремоделирования управляют удалением последовательности спейсера ITS2 во время сборки 60S субъединиц рибосомы у S. cerevisiae (Кандидатская диссертация). Университет Карнеги Меллон. Архивировано из оригинал 3 августа 2017 г.
- ^ Вейлер EW, Новер L (2008). Allgemeine und Molekulare Botanik (на немецком). Штутгарт: Георг Тиме Верлаг. п. 532. ISBN 978-3-13-152791-2.
- ^ де ла Круз Дж., Карбштейн К., Вулфорд Дж. Л. (2015). «Функции рибосомных белков в сборке эукариотических рибосом in vivo». Ежегодный обзор биохимии. 84: 93–129. Дои:10.1146 / annurev-biochem-060614-033917. ЧВК 4772166. PMID 25706898.
- ^ «Scitable по своей природе перевод / перевод РНК».
- ^ Бенне Р., Слоф П. (1987). «Эволюция аппарата синтеза митохондриального белка». Биосистемы. 21 (1): 51–68. Дои:10.1016/0303-2647(87)90006-2. PMID 2446672.
- ^ «Рибосомы». Архивировано из оригинал на 2009-03-20. Получено 2011-04-28.
- ^ Palade GE (январь 1955 г.). «Компонент цитоплазмы в виде мелких частиц». Журнал биофизической и биохимической цитологии. 1 (1): 59–68. Дои:10.1083 / jcb.1.1.59. ЧВК 2223592. PMID 14381428.
- ^ Робертс РБ, изд. (1958). "Вступление". Микросомальные частицы и синтез белка. Нью-Йорк: Pergamon Press, Inc.
- ^ "Нобелевская премия по физиологии и медицине 1974 г.". Nobelprize.org. Нобелевский фонд. В архиве из оригинала 26 января 2013 г.. Получено 10 декабря 2012.
- ^ «Нобелевская премия по химии 2009 г.». Нобелевский фонд. В архиве из оригинала 28 апреля 2012 г.. Получено 10 декабря 2012.
- ^ Курляндская Ц.Г. (1960). «Молекулярная характеристика рибонуклеиновой кислоты из рибосом Escherichia coli». Журнал молекулярной биологии. 2 (2): 83–91. Дои:10.1016 / с0022-2836 (60) 80029-0.
- ^ Уилсон Д. Н., Дудна Кейт Дж. Х. (май 2012 г.). «Структура и функции эукариотической рибосомы». Перспективы Колд-Спринг-Харбор в биологии. 4 (5): a011536. Дои:10.1101 / cshperspect.a011536. ЧВК 3331703. PMID 22550233.
- ^ Ниссен П., Хансен Дж., Бан Н., Мур ПБ, Стейтц Т.А. (август 2000 г.). «Структурные основы активности рибосом в синтезе пептидных связей». Наука. 289 (5481): 920–30. Bibcode:2000Sci ... 289..920N. Дои:10.1126 / science.289.5481.920. PMID 10937990.
- ^ а б Wimberly BT, Brodersen DE, Clemons WM, Morgan-Warren RJ, Carter AP, Vonrhein C, Hartsch T., Ramakrishnan V (сентябрь 2000 г.). «Структура 30S субъединицы рибосомы». Природа. 407 (6802): 327–39. Bibcode:2000Натурал 407..327Вт. Дои:10.1038/35030006. PMID 11014182.
- ^ а б c d е ж грамм Альбертс, Брюс; и другие. (2002). Молекулярная биология клетки (4-е изд.). Наука о гирляндах. п. 342. ISBN 978-0-8153-3218-3.
- ^ а б Гарретт Р., Гришэм К.М. (2009). Биохимия (4-е изд.). Услуги обучения Cengage. ISBN 978-0-495-11464-2.
- ^ Collatz E, Küchler E, Stöffler G, Czernilofsky AP (апрель 1976 г.). «Сайт реакции на рибосомный белок L27 с производной аффинной метки тРНК Met f». Письма FEBS. 63 (2): 283–6. Дои:10.1016/0014-5793(76)80112-3. PMID 770196.
- ^ Черниловский А.П., Коллатц Э., Штеффлер Г, Кюхлер Э. (январь 1974 г.). «Белки в сайтах связывания тРНК рибосом Escherichia coli». Труды Национальной академии наук Соединенных Штатов Америки. 71 (1): 230–4. Bibcode:1974PNAS ... 71..230C. Дои:10.1073 / пнас.71.1.230. ЧВК 387971. PMID 4589893.
- ^ Черниловский А.П., Курляндская Ц.Г., Штеффлер Г. (октябрь 1975 г.). «30S рибосомные белки, связанные с 3'-концом 16S РНК». Письма FEBS. 58 (1): 281–4. Дои:10.1016/0014-5793(75)80279-1. PMID 1225593.
- ^ а б c Бен-Шем А., Гарро де Лубресс Н., Мельников С., Дженнер Л., Юсупова Г., Юсупов М. (декабрь 2011 г.). «Структура эукариотической рибосомы с разрешением 3,0 Å». Наука. 334 (6062): 1524–9. Bibcode:2011Научный ... 334.1524B. Дои:10.1126 / наука.1212642. PMID 22096102.
- ^ а б Рабл Дж., Лейбундгут М., Атаиде С.Ф., Хааг А., Бан Н. (февраль 2011 г.). «Кристаллическая структура эукариотической 40S рибосомной субъединицы в комплексе с фактором инициации 1» (PDF). Наука. 331 (6018): 730–6. Bibcode:2011Научный ... 331..730R. Дои:10.1126 / science.1198308. HDL:20.500.11850/153130. PMID 21205638.
- ^ а б Klinge S, Voigts-Hoffmann F, Leibundgut M, Arpagaus S, Ban N (ноябрь 2011 г.). «Кристаллическая структура эукариотической 60S рибосомной субъединицы в комплексе с фактором инициации 6». Наука. 334 (6058): 941–8. Bibcode:2011Наука ... 334..941K. Дои:10.1126 / наука.1211204. PMID 22052974.
- ^ Фабиянски С., Пеллегрини М. (1977). «Идентификация белков в сайте связывания пептидил-тРНК рибосом печени крысы». Молекулярная и общая генетика. 184 (3): 551–6. Дои:10.1007 / BF00431588. PMID 6950200.
- ^ Агравал РК, Шарма М.Р. (декабрь 2012 г.). «Структурные аспекты митохондриального трансляционного аппарата». Текущее мнение в структурной биологии. 22 (6): 797–803. Дои:10.1016 / j.sbi.2012.08.003. ЧВК 3513651. PMID 22959417.
- ^ Шарма М.Р., Бут TM, Симпсон Л., Маслов Д.А., Агравал РК (июнь 2009 г.). «Структура митохондриальной рибосомы с минимальной РНК». Труды Национальной академии наук Соединенных Штатов Америки. 106 (24): 9637–42. Bibcode:2009ПНАС..106.9637С. Дои:10.1073 / pnas.0901631106. ЧВК 2700991. PMID 19497863.
- ^ Арчибальд Дж. М., Лейн CE (2009). «Идет, идет, не совсем исчезло: нуклеоморфы как пример сокращения ядерного генома». Журнал наследственности. 100 (5): 582–90. Дои:10.1093 / jhered / esp055. PMID 19617523.
- ^ Recht MI, Douthwaite S, Puglisi JD (июнь 1999 г.). «Основы прокариотической специфичности действия аминогликозидных антибиотиков». Журнал EMBO. 18 (11): 3133–8. Дои:10.1093 / emboj / 18.11.3133. ЧВК 1171394. PMID 10357824.
- ^ О'Брайен Т.В. (май 1971 г.). «Общее появление рибосом 55 S в митохондриях печени млекопитающих». Журнал биологической химии. 246 (10): 3409–17. PMID 4930061.
- ^ «Подавление костного мозга, индуцированное хлорамфениколом». JAMA. 213 (7): 1183–1184. 1970-08-17. Дои:10.1001 / jama.1970.03170330063011. ISSN 0098-7484.
- ^ Ньюман С.М., Бойнтон Дж. Э., Гиллхэм Н. В., Рэндольф-Андерсон Б. Л., Джонсон А. М., Харрис Е. Х. (декабрь 1990 г.). «Трансформация генов рибосомной РНК хлоропластов у Chlamydomonas: молекулярная и генетическая характеристика процессов интеграции». Генетика. 126 (4): 875–88. ЧВК 1204285. PMID 1981764.
- ^ Пенев П.И., Фахретаха-Аваль С., Патель В.Дж., Канноне Дж.Дж., Гутелл Р.Р., Петров А.С., Уильямс Л.Д., Гласс Дж.Б. (август 2020 г.). «Увеличенные сегменты экспансии рибосомной РНК в архей Асгарда». Геномная биология и эволюция. Дои:10.1093 / gbe / evaa170. PMID 32785681.
- ^ а б Ban N, Nissen P, Hansen J, Moore PB, Steitz TA (август 2000 г.). «Полная атомная структура большой рибосомной субъединицы при разрешении 2,4 А». Наука. 289 (5481): 905–20. Bibcode:2000Sci ... 289..905B. CiteSeerX 10.1.1.58.2271. Дои:10.1126 / science.289.5481.905. PMID 10937989.
- ^ Schluenzen F, Tocilj A, Zarivach R, Harms J, Gluehmann M, Janell D, Bashan A, Bartels H, Agmon I, Franceschi F, Yonath A (сентябрь 2000 г.). «Структура функционально активированной малой рибосомной субъединицы при разрешении 3,3 ангстрем». Клетка. 102 (5): 615–23. Дои:10.1016 / S0092-8674 (00) 00084-2. PMID 11007480.
- ^ Юсупов М.М., Юсупова Г.З., Бауком А., Либерман К., Эрнест Т.Н., Кейт Дж. Х., Ноллер Х. Ф. (май 2001 г.). «Кристаллическая структура рибосомы при разрешении 5,5 А». Наука. 292 (5518): 883–96. Bibcode:2001Научный ... 292..883Y. Дои:10.1126 / science.1060089. PMID 11283358.
- ^ Шувирт Б.С., Боровинская М.А., Хау К.В., Чжан В., Вила-Санджурджо А., Холтон Дж. М., Кейт Дж. Х. (ноябрь 2005 г.). «Структуры бактериальной рибосомы при разрешении 3,5 А». Наука. 310 (5749): 827–34. Bibcode:2005Наука ... 310..827С. Дои:10.1126 / science.1117230. PMID 16272117.
- ^ Митра К., Шаффицель С., Шейх Т., Тама Ф., Дженни С., Брукс С.Л., Бан Н., Фрэнк Дж. (Ноябрь 2005 г.). «Структура канала, проводящего белок E. coli, связанного с транслирующей рибосомой». Природа. 438 (7066): 318–24. Bibcode:2005Натура.438..318М. Дои:10.1038 / природа04133. ЧВК 1351281. PMID 16292303.
- ^ Селмер М., Данхэм К.М., Мерфи Ф.В., Вейкслбаумер А., Петри С., Келли А.С., Вейр Дж. Р., Рамакришнан В. (сентябрь 2006 г.). «Структура 70S рибосомы в комплексе с мРНК и тРНК». Наука. 313 (5795): 1935–42. Bibcode:2006Научный ... 313.1935S. Дои:10.1126 / science.1131127. PMID 16959973.
- ^ Коростелев А., Траханов С., Лаурберг М., Ноллер Х. Ф. (сентябрь 2006 г.). «Кристаллическая структура комплекса 70S рибосома-тРНК обнаруживает функциональные взаимодействия и перестройки». Клетка. 126 (6): 1065–77. Дои:10.1016 / j.cell.2006.08.032. PMID 16962654.
- ^ Юсупова Г., Дженнер Л., Рис Б., Морас Д., Юсупов М. (ноябрь 2006 г.). «Структурная основа движения матричной РНК по рибосоме». Природа. 444 (7117): 391–4. Bibcode:2006Натура.444..391л. Дои:10.1038 / природа05281. PMID 17051149.
- ^ а б «Специализированные внутренние структуры прокариот | Безграничная микробиология». course.lumenlearning.com. Получено 2018-09-27.
- ^ Савир Ю., Тластий Т. (апрель 2013 г.). «Рибосома как оптимальный декодер: урок молекулярного распознавания». Клетка. 153 (2): 471–9. Дои:10.1016 / j.cell.2013.03.032. PMID 23582332.
- ^ Коркмаз Г., Саньял С. (сентябрь 2017 г.). "Кишечная палочка". Журнал биологической химии. 292 (36): 15134–15142. Дои:10.1074 / jbc.M117.785238. ЧВК 5592688. PMID 28743745.
- ^ Коневега А.Л., Соболева Н.Г., Махно В.И., Семенков Ю.П., Винтермейер В., Роднина М.В., Катунин В.И. (январь 2004 г.). «Пуриновые основания в положении 37 тРНК стабилизируют кодон-антикодонное взаимодействие в рибосомном сайте А посредством стэкинга и Mg2 + -зависимых взаимодействий». РНК. 10 (1): 90–101. Дои:10.1261 / rna.5142404. ЧВК 1370521. PMID 14681588.
- ^ Роднина М.В., Берингер М., Винтермейер В. (январь 2007 г.). «Как рибосомы образуют пептидные связи». Тенденции в биохимических науках. 32 (1): 20–6. Дои:10.1016 / j.tibs.2006.11.007. PMID 17157507.
- ^ Чех TR (август 2000 г.). «Структурная биология. Рибосома - рибозим». Наука. 289 (5481): 878–9. Дои:10.1126 / science.289.5481.878. PMID 10960319.
- ^ Банерджи Д., Саньял С. (октябрь 2014 г.). «Белковая фолдинговая активность рибосомы (PFAR) - мишень для антиприонных соединений». Вирусы. 6 (10): 3907–24. Дои:10.3390 / v6103907. ЧВК 4213570. PMID 25341659.
- ^ Федоров А.Н., Болдуин Т.О. (декабрь 1997 г.). «Котрансляционный фолдинг белка». Журнал биологической химии. 272 (52): 32715–8. Дои:10.1074 / jbc.272.52.32715. PMID 9407040.
- ^ Болдуин Р.Л. (июнь 1975 г.). «Промежуточные продукты в реакциях сворачивания белков и механизм сворачивания белков». Ежегодный обзор биохимии. 44 (1): 453–75. Дои:10.1146 / annurev.bi.44.070175.002321. PMID 1094916.
- ^ Дас Д., Дас А., Саманта Д., Гош Дж., Дасгупта С., Бхаттачарья А., Басу А., Саньял С., Дас Гупта С. (август 2008 г.). «Роль рибосомы в сворачивании белков». Биотехнологический журнал. 3 (8): 999–1009. Дои:10.1002 / биот.200800098. PMID 18702035.
- ^ Dabrowski-Tumanski P, Piejko M, Niewieczerzal S, Stasiak A, Sulkowska JI (декабрь 2018 г.). «Узелок белка за счет активного переплетения растущей полипептидной цепи, выходящей из канала выхода рибосомы». Журнал физической химии B. 122 (49): 11616–11625. Дои:10.1021 / acs.jpcb.8b07634. PMID 30198720.
- ^ Брэндман О., Стюарт-Орнштейн Дж., Вонг Д., Ларсон А., Уильямс С.К., Ли Г.В., Чжоу С., Кинг Д., Шен П.С., Вайбезан Дж., Данн Дж. Г., Рускин С., Инада Т., Фрост А., Вайсман Дж. С. (ноябрь 2012 г.) . «Связанный с рибосомами комплекс контроля качества запускает деградацию растущих пептидов и сигнализирует о стрессе трансляции». Клетка. 151 (5): 1042–54. Дои:10.1016 / j.cell.2012.10.044. ЧВК 3534965. PMID 23178123.
- ^ Defenouillère Q, Yao Y, Mouaikel J, Namane A, Galopier A, Decourty L, Doyen A, Malabat C, Saveanu C, Jacquier A, Fromont-Racine M (март 2013 г.). «Cdc48-ассоциированный комплекс, связанный с частицами 60S, необходим для удаления аберрантных продуктов трансляции». Труды Национальной академии наук Соединенных Штатов Америки. 110 (13): 5046–51. Bibcode:2013PNAS..110.5046D. Дои:10.1073 / пнас.1221724110. ЧВК 3612664. PMID 23479637.
- ^ Шен П.С., Пак Дж., Цинь И, Ли Х, Парсавар К., Ларсон М. Х., Кокс Дж, Ченг И, Ламбовиц А. М., Вайсман Дж. С., Брандман О., Фрост А. (январь 2015 г.). «Синтез белка. Rqc2p и 60S рибосомные субъединицы опосредуют мРНК-независимое удлинение растущих цепей». Наука. 347 (6217): 75–8. Bibcode:2015Научный ... 347 ... 75S. Дои:10.1126 / science.1259724. ЧВК 4451101. PMID 25554787.
- ^ Кили Дж., Гутникофф Р. (02.01.2015). «Исследования рибосом открывают новый механизм синтеза белка» (Пресс-релиз). Медицинский институт Говарда Хьюза. В архиве из оригинала от 12.01.2015. Получено 2015-01-16.
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Рибосомы, связанные с мембраной, определяют грубую ER». Молекулярная биология клетки (4-е изд.). Нью-Йорк: Наука Гарланд. ISBN 978-0-8153-4072-0.
- ^ а б Ноллер HF (апрель 2012 г.). «Эволюция синтеза белка из мира РНК». Перспективы Колд-Спринг-Харбор в биологии. 4 (4): a003681. Дои:10.1101 / cshperspect.a003681. ЧВК 3312679. PMID 20610545.
- ^ Даббс ER (1986). Мутантные исследования прокариотической рибосомы. Нью-Йорк: Springer-Verlag.
- ^ Ноллер Х.Ф., Хоффарт В., Зимняк Л. (июнь 1992 г.). «Необычная устойчивость пептидилтрансферазы к процедурам экстракции белков». Наука. 256 (5062): 1416–9. Bibcode:1992Научный ... 256.1416N. Дои:10.1126 / science.1604315. PMID 1604315.
- ^ Номура М., Мидзусима С., Одзаки М., Трауб П., Лоури К.В. (1969). «Структура и функции рибосом и их молекулярных компонентов». Симпозиумы Колд-Спринг-Харбор по количественной биологии. 34: 49–61. Дои:10.1101 / sqb.1969.034.01.009. PMID 4909519.
- ^ а б Рут-Бернштейн М, Рут-Бернштейн Р (февраль 2015 г.). «Рибосома как недостающее звено в эволюции жизни». Журнал теоретической биологии. 367: 130–158. Дои:10.1016 / j.jtbi.2014.11.025. PMID 25500179.
- ^ Ярус М (2002). «Изначальная генетика: фенотип рибоцита». Ежегодный обзор генетики. 36: 125–51. Дои:10.1146 / annurev.genet.36.031902.105056. PMID 12429689.
- ^ Фортер П, Крупович М (2012). «Происхождение вирионов и вироэлементов: пересмотр гипотезы побега». Вирусы: важные агенты жизни. С. 43–60. Дои:10.1007/978-94-007-4899-6_3. ISBN 978-94-007-4898-9.
- ^ Каэтано-Аноллес Г, Сеуфферхельд MJ (2013). «Коэволюционные корни биохимии и клеточной организации бросают вызов парадигме мира РНК». Журнал молекулярной микробиологии и биотехнологии. 23 (1–2): 152–77. Дои:10.1159/000346551. PMID 23615203.
- ^ Саладино Р., Ботта Дж., Пино С., Костанцо Дж., Ди Мауро Е. (август 2012 г.). «Генетика превыше всего или метаболизм превыше всего? Ключ к разгадке формамида». Обзоры химического общества. 41 (16): 5526–65. Дои:10.1039 / c2cs35066a. PMID 22684046.
- ^ Fox GE (сентябрь 2010 г.). «Происхождение и эволюция рибосомы». Холодная весна Харб Perspect Biol. 2 (9). Дои:10.1101 / cshperspect.a003483. PMID 20534711.
- ^ Fox GE (2016). «Происхождение и ранняя эволюция рибосомы». В Hernández G, Jagus R (ред.). Эволюция оборудования для синтеза белка и его регулирование. Швейцария: Шпрингер, Чам. С. 31–60. ISBN 978-3-319-39468-8.
- ^ Ши, Чжэнь; Фудзи, Котаро; Ковари, Кайл М .; Genuth, Наоми Р .; Röst, Hannes L .; Теруэль, Мэри Н .; Барна, Мария (2017). «Гетерогенные рибосомы предпочтительно транслируют отдельные субпулы мРНК в масштабе всего генома». Молекулярная клетка. Elsevier BV. 67 (1): 71–83.e7. Дои:10.1016 / j.molcel.2017.05.021. ISSN 1097-2765.
- ^ Сюэ, Шифэн; Барна, Мария (23.05.2012). «Специализированные рибосомы: новый рубеж в регуляции генов и биологии организма». Обзоры природы Молекулярная клеточная биология. ООО "Спрингер Сайенс энд Бизнес Медиа". 13 (6): 355–369. Дои:10.1038 / nrm3359. ISSN 1471-0072. ЧВК 4039366.
- ^ Ферретти, Макс Б.; Карбштейн, Катрин (07.02.2019). "Действительно ли существует функциональная специализация рибосом?". РНК. Лаборатория Колд-Спринг-Харбор. 25 (5): 521–538. Дои:10.1261 / rna.069823.118. ISSN 1355-8382.
- ^ Фарли-Барнс, Кэтрин I .; Огава, Лиза М .; Базерга, Сьюзан Дж. (2019). «Рибосомопатии: старые концепции, новые противоречия». Тенденции в генетике. Elsevier BV. 35 (10): 754–767. Дои:10.1016 / j.tig.2019.07.004. ISSN 0168-9525.
- ^ Мауро В.П., Эдельман Г.М. (сентябрь 2002 г.). «Гипотеза рибосомного фильтра». Труды Национальной академии наук Соединенных Штатов Америки. 99 (19): 12031–6. Bibcode:2002ПНАС ... 9912031М. Дои:10.1073 / пнас.192442499. ЧВК 129393. PMID 12221294.
- ^ Сюэ С., Барна М. (май 2012 г.). «Специализированные рибосомы: новый рубеж в регуляции генов и биологии организма». Обзоры природы. Молекулярная клеточная биология. 13 (6): 355–69. Дои:10.1038 / nrm3359. ЧВК 4039366. PMID 22617470.
- ^ Mathis AD, Naylor BC, Carson RH, Evans E, Harwell J, Knecht J, Hexem E, Peelor FF, Miller BF, Hamilton KL, Transtrum MK, Bikman BT, Price JC (февраль 2017 г.). «Механизмы изменения содержания рибосом in vivo в ответ на сигналы питательных веществ». Молекулярная и клеточная протеомика. 16 (2): 243–254. Дои:10.1074 / mcp.M116.063255. ЧВК 5294211. PMID 27932527.
- ^ Steffen, Kristan K .; Маккормик, Марк А .; Pham, Kim M .; MacKay, Vivian L .; Делани, Джо Р .; Мураками, Кристофер Дж .; Kaeberlein, Matt; Кеннеди, Брайан К. (29 февраля 2012 г.). «Дефицит рибосом защищает от стресса ER у Saccharomyces cerevisiae». Генетика. Американское общество генетиков. 191 (1): 107–118. Дои:10.1534 / генетика.111.136549. ISSN 0016-6731. ЧВК 3338253.
- ^ Ли С.В., Бергер С.Дж., Мартинович С., Паша-Толич Л., Андерсон Г.А., Шен И., Чжао Р., Смит Р.Д. (апрель 2002 г.). «Прямой масс-спектрометрический анализ интактных белков большой рибосомной субъединицы дрожжей с использованием капиллярной ЖХ / FTICR». Труды Национальной академии наук Соединенных Штатов Америки. 99 (9): 5942–7. Bibcode:2002PNAS ... 99.5942L. Дои:10.1073 / pnas.082119899. ЧВК 122881. PMID 11983894.
- ^ Кэрролл А.Дж., Хизлвуд Д.Л., Ито Дж., Миллар А.Х. (февраль 2008 г.). «Анализ протеома цитозольной рибосомы Arabidopsis дает подробное представление о его компонентах и их посттрансляционных модификациях». Молекулярная и клеточная протеомика. 7 (2): 347–69. Дои:10.1074 / mcp.m700052-mcp200. PMID 17934214.
- ^ Одинцова Т.И., Мюллер Э.С., Иванов А.В., Егоров Т.А., Бинерт Р., Владимиров С.Н., Костка С., Отто А., Виттманн-Либольд Б., Карпова Г.Г. (апрель 2003 г.). «Характеристика и анализ посттрансляционных модификаций белков большой цитоплазматической рибосомной субъединицы человека с помощью масс-спектрометрии и секвенирования по Эдману». Журнал химии белков. 22 (3): 249–58. Дои:10.1023 / а: 1025068419698. PMID 12962325.
- ^ Ю Й, Джи Х, Дудна Дж. А., Лири Дж. А. (июнь 2005 г.). «Масс-спектрометрический анализ субъединицы рибосомы 40S человека: нативные и связанные с IRES комплексы HCV». Белковая наука. 14 (6): 1438–46. Дои:10.1110 / пс. 041293005. ЧВК 2253395. PMID 15883184.
- ^ Зейдан К., Ван З., Де Майо А., Харт Г. В. (июнь 2010 г.). «Циклические ферменты O-GlcNAc связываются с механизмом трансляции и модифицируют основные рибосомные белки». Молекулярная биология клетки. 21 (12): 1922–36. Дои:10.1091 / mbc.e09-11-0941. ЧВК 2883937. PMID 20410138.
- ^ Ландри Д.М., Герц М.И., Томпсон С.Р. (декабрь 2009 г.). «RPS25 необходим для инициации трансляции IRES вирусов Dicistroviridae и гепатита C». Гены и развитие. 23 (23): 2753–64. Дои:10.1101 / gad.1832209. ЧВК 2788332. PMID 19952110.
- ^ Decatur WA, Fournier MJ (июль 2002 г.). «Модификации рРНК и функция рибосом». Тенденции в биохимических науках. 27 (7): 344–51. Дои:10.1016 / s0968-0004 (02) 02109-6. PMID 12114023.
- ^ Натчар С.К., Мясников А.Г., Кратзат Х., Хаземанн И., Клахольц Б.П. (ноябрь 2017 г.). «Визуализация химических модификаций в структуре рибосомы 80S человека». Природа. 551 (7681): 472–477. Bibcode:2017Натура.551..472Н. Дои:10.1038 / природа24482. PMID 29143818.
- ^ Гуо Х (август 2018). «Специализированные рибосомы и контроль трансляции». Сделки Биохимического Общества. 46 (4): 855–869. Дои:10.1042 / BST20160426. PMID 29986937.
внешняя ссылка
- Лабораторный компьютер имитирует движение рибосомы
- Роль рибосомы, Гвен В. Чайлдс, копия здесь
- Рибосома в Протеопедия - Бесплатная совместная трехмерная энциклопедия белков и других молекул.
- Семейства рибосомных белков в ExPASy
- Молекула месяца © RCSB Protein Data Bank:
- Структура рибосом с помощью трехмерной электронной микроскопии в банке данных EM (EMDB)
 Эта статья включаетматериалы общественного достояния от NCBI документ: «Учебник по науке».
Эта статья включаетматериалы общественного достояния от NCBI документ: «Учебник по науке».