Протеасома - Proteasome


Протеасомы находятся белковые комплексы которые ухудшают ненужные или поврежденные белки к протеолиз, а химическая реакция это ломается пептидные связи. Ферменты которые помогают такие реакции называются протеазы.
Протеасомы являются частью основного механизма, с помощью которого клетки регулировать концентрация определенных белков и разлагаются неправильно свернутые белки. Белки маркируются для разложения небольшим белком, называемым убиквитин. Реакция мечения катализируется ферментами, называемыми убиквитинлигазы. Как только белок помечен одной молекулой убиквитина, это сигнал другим лигазам присоединить дополнительные молекулы убиквитина. В результате полиубиквитиновая цепь который связывается протеасомой, позволяя ей расщеплять меченый белок.[1] Процесс деградации дает пептиды примерно от семи до восьми аминокислоты длинные, которые затем можно расщепить на более короткие аминокислотные последовательности и использовать в синтезирующий новые белки.[1]
Протеасомы находятся внутри всех эукариоты и археи, а в некоторых бактерии.У эукариот протеасомы расположены как в ядро и в цитоплазма.[2]
В структура, протеасома представляет собой цилиндрический комплекс, содержащий «ядро» из четырех уложенных друг на друга колец, образующих центральную пору. Каждое кольцо состоит из семи отдельных белков. Два внутренних кольца состоят из семи β субъединицы которые содержат от трех до семи протеаз активные сайты. Эти сайты расположены на внутренней поверхности колец, поэтому целевой белок должен войти в центральную пору, прежде чем он распадется. Два внешних кольца содержат по семь субъединицы α чья функция заключается в поддержании «ворот», через которые белки попадают в ствол. Эти субъединицы α контролируются путем связывания со структурами «кэпа» или регулирующие частицы которые распознают теги полиубиквитина, прикрепленные к белковым субстратам, и инициируют процесс деградации. Общая система убиквитинирования и протеасомной деградации известна как убиквитин-протеасомная система.[3]
Путь протеасомной деградации важен для многих клеточных процессов, включая клеточный цикл, регулирование экспрессия гена, и ответы на окислительный стресс. Важность протеолитической деградации внутри клеток и роль убиквитина в протеолитических путях была признана в награде 2004 г. Нобелевская премия по химии к Аарон Цехановер, Аврам Гершко и Ирвин Роуз.[4]
Открытие
До открытия убиквитин-протеасомной системы считалось, что деградация белка в клетках в основном зависит от лизосомы мембраносвязанный органеллы с кислый и протеаза -заполненные внутренние части, которые могут разрушать, а затем перерабатывать экзогенные белки и состарившиеся или поврежденные органеллы.[1] Однако работы Джозефа Этлингера и Альфред Гольдберг в 1977 г. по АТФ-зависимой деградации белков в ретикулоциты, в которых отсутствуют лизосомы, свидетельствует о наличии второго механизма внутриклеточной деградации.[5] В 1978 году было показано, что он состоит из нескольких отдельных белковых цепей, что в то время было новинкой среди протеаз.[6] Позже работа над модификацией гистоны привело к выявлению неожиданного ковалентный модификация гистонового белка связью между лизин боковая цепь гистона и C-терминал глицин остаток убиквитин, белок, функции которого неизвестны.[7] Затем было обнаружено, что ранее идентифицированный белок, связанный с протеолитической деградацией, известный как АТФ-зависимый фактор протеолиза 1 (APF-1), был тем же белком, что и убиквитин.[8] Протеолитическая активность этой системы была выделена в виде мультибелкового комплекса, первоначально названного Мультикаталитическим протеиназным комплексом Шервином Уилком и Марион Орловски.[9] Позже АТФ -зависимый протеолитический комплекс, ответственный за убиквитин-зависимую деградацию белка, был открыт и назван протеасомой 26S.[10][11]
Большая часть ранних работ, приведших к открытию протеасомной системы убиквитина, была проведена в конце 1970-х - начале 1980-х гг. Технион в лаборатории Аврам Гершко, куда Аарон Цехановер работал в аспирантуре. Годовой творческий отпуск Гершко в лаборатории Ирвин Роуз на Онкологический центр Fox Chase предоставил ключевые концептуальные идеи, хотя позже Роуз преуменьшил свою роль в открытии.[12] Трое разделили 2004 год. Нобелевская премия по химии за их работу по открытию этой системы.[4]
Несмотря на то что электронная микроскопия данные, показывающие многослойную кольцевую структуру протеасомы, стали доступны в середине 1980-х годов,[13] первая структура коровой частицы протеасомы не была решена Рентгеновская кристаллография до 1994 г.[14] В 2018 году первые атомные структуры холофермента протеасомы 26S человека в комплексе с полиубиквитилированным белковым субстратом были решены с помощью криогенной электронной микроскопии, выявив механизмы, с помощью которых субстрат распознается, деубиквитилируется, разворачивается и разрушается протеасомой 26S человека.[15]
Структура и организация
Подкомпоненты протеасомы часто называют их Сведберг коэффициент седиментации (обозначается S). Протеасома, наиболее широко используемая у млекопитающих, - это цитозольная протеасома 26S, которая составляет около 2000 килодальтон (кДа) дюйм молекулярная масса содержащие одну субъединицу 20S белка и две субъединицы регуляторной кэпа 19S. Ядро является полым и представляет собой закрытую полость, в которой разрушаются белки; отверстия на двух концах ядра позволяют входить целевому белку. Каждый конец коровой частицы ассоциирует с регуляторной субъединицей 19S, которая содержит несколько АТФаза активные сайты и сайты связывания убиквитина; именно эта структура распознает полиубиквитинированные белки и переносит их в каталитическое ядро.[15] Альтернативная форма регуляторной субъединицы, называемая частица 11S, может ассоциироваться с ядром по существу таким же образом, как частица 19S; 11S может играть роль в деградации чужеродных пептидов, таких как пептиды, полученные после заражения вирус.[16]
20S основная частица
Количество и разнообразие субъединиц, содержащихся в основной частице 20S, зависит от организма; количество отдельных и специализированных субъединиц больше у многоклеточных, чем у одноклеточных организмов, и больше у эукариот, чем у прокариот. Все 20S частицы состоят из четырех уложенных друг на друга гептамерных кольцевых структур, которые сами состоят из двух различных типов субъединиц; субъединицы α имеют структурную природу, тогда как субъединицы β преимущественно каталитический. Α-субъединицы псевдоферменты гомологичен β-субъединицам. Они собраны с их N-концом, смежным с концом субъединиц β.[17] Два внешних кольца в стопке состоят из семи α-субъединиц каждое, которые служат стыковочными доменами для регуляторных частиц и N-концов α-субъединиц (Pfam PF10584 ) образуют заслонку, перекрывающую нерегулируемый доступ подложек во внутреннюю полость.[18] Каждое из двух внутренних колец состоит из семи β-субъединиц и на своих N-концах содержит активные сайты протеазы, которые осуществляют реакции протеолиза.[19] В очищенном комплексе были идентифицированы три различные каталитические активности: химотрипсиноподобный, трипсиноподобный и пептидилглутамилпептидный гидролиз.[20] Размер протеасомы относительно консервативен и составляет около 150 ангстремы (Å) на 115 Å. Внутренняя камера имеет ширину не более 53 Å, хотя вход может иметь ширину до 13 Å, что позволяет предположить, что субстратные белки должны быть по крайней мере частично развернуты, чтобы войти.[21]
В археи Такие как Термоплазма ацидофильная, все субъединицы α и все β идентичны, тогда как протеасомы эукариот, такие как протеасомы в дрожжи содержат семь различных типов каждой субъединицы. В млекопитающие субъединицы β1, β2 и β5 являются каталитическими; хотя они имеют общий механизм, они имеют три различных специфичности субстрата. химотрипсин -подобно, трипсин -вроде, и пептидил-глутамилпептид-гидролизующий (PHGH).[22] Альтернативные формы β, обозначенные β1i, β2i и β5i, могут быть выражены в кроветворный клетки в ответ на воздействие про-воспалительный сигналы Такие как цитокины, особенно, интерферон гамма. Протеасома, собранная из этих альтернативных субъединиц, известна как иммунопротеасома, субстратная специфичность которого изменена по сравнению с нормальной протеасомой.[21]Недавно в клетках человека была идентифицирована альтернативная протеасома, в которой отсутствует коровая субъединица α3.[23] Эти протеасомы (известные как протеасомы α4-α4) вместо отсутствующей субъединицы α3 образуют 20S ядерные частицы, содержащие дополнительную субъединицу α4. Ранее было известно, что эти альтернативные протеасомы α4-α4 существуют в дрожжах.[24] Хотя точная функция этих изоформ протеасом все еще в значительной степени неизвестна, клетки, экспрессирующие эти протеасомы, демонстрируют повышенную устойчивость к токсичности, вызванной ионами металлов, такими как кадмий.[23][25]
Регуляторная частица 19S
Частица 19S у эукариот состоит из 19 отдельных белков и делится на две части: основание из 9 субъединиц, которое непосредственно связывается с α-кольцом ядерной частицы 20S, и крышку из 10 субъединиц. Шесть из девяти основных белков представляют собой субъединицы АТФазы из семейства AAA, и эволюционный гомолог этих АТФаз существует в архее, названный PAN (нуклеотидаза, активирующая протеасомы).[26] Ассоциация частиц 19S и 20S требует связывания АТФ с субъединицами 19S АТФазы, и гидролиз АТФ необходим для собранного комплекса для разрушения свернутых и убиквитинированных белков. Обратите внимание, что только этап развертывания субстрата требует энергии от гидролиза АТФ, в то время как только связывание АТФ может поддерживать все другие этапы, необходимые для деградации белка (например, сборка комплекса, открытие ворот, транслокация и протеолиз).[27][28] Фактически, связывание АТФ с АТФазами само по себе поддерживает быструю деградацию развернутых белков. Однако, хотя гидролиз АТФ требуется только для разворачивания, еще не ясно, может ли эта энергия использоваться для сочетания некоторых из этих стадий.[28][29]
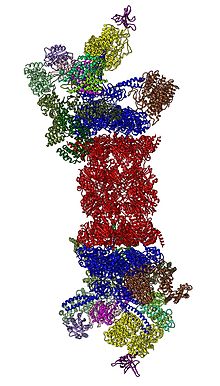
В 2012 году два независимых исследования выяснили молекулярную архитектуру протеасомы 26S с помощью электронная микроскопия одиночных частиц.[31][32] В 2016 году три независимых исследования определили первую структуру близкого к атомному разрешению протеасомы 26S человека в отсутствие субстратов с помощью крио-ЭМ.[33][34][35] В 2018 году была проведена большая работа по выяснению подробных механизмов деубиквитилирования, инициации транслокации и процессивного разворачивания субстратов путем одновременного определения семи атомных структур задействованной в субстрат 26S протеасомы.[15] В основе 19S, непосредственно примыкающих к 20S, находятся AAA-ATPases (Белки AAA ), которые собираются в гетерогексамерное кольцо порядка Rpt1 / Rpt2 / Rpt6 / Rpt3 / Rpt4 / Rpt5. Это кольцо представляет собой тример димеров: Rpt1 / Rpt2, Rpt6 / Rpt3 и Rpt4 / Rpt5 димеризуются через свои N-концевые спиральные спирали. Эти спиральные катушки выступают из гексамерного кольца. Самые большие регуляторные частицы, не являющиеся АТФазами, Rpn1 и Rpn2, связываются с кончиками Rpt1 / 2 и Rpt6 / 3 соответственно. Рецептор убиквитина Rpn13 связывается с Rpn2 и завершает базовый куб-комплекс. Крышка покрывает половину гексамера ААА-АТФазы (Rpt6 / Rpt3 / Rpt4) и, неожиданно, напрямую контактирует с 20S через Rpn6 и в меньшей степени через Rpn5. Субъединицы Rpn9, Rpn5, Rpn6, Rpn7, Rpn3 и Rpn12, которые структурно связаны между собой и с субъединицами Комплекс COP9 и eIF3 (отсюда и называемые субъединицы PCI) собираются в подковообразную структуру, включающую гетеродимер Rpn8 / Rpn11. Rpn11, деубиквитинирующий фермент, помещается в устье гексамера ААА-АТФазы, что идеально подходит для удаления фрагментов убиквитина непосредственно перед транслокацией субстратов в 20S. Второй идентифицированный на сегодняшний день рецептор убиквитина, Rpn10, расположен на периферии крышки, рядом с субъединицами Rpn8 и Rpn9.
Конформационные изменения 19S
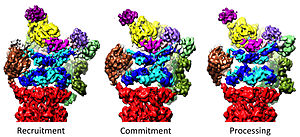
Регуляторная частица 19S в составе голофермента протеасомы 26S на сегодняшний день обнаружена в шести сильно различающихся конформационных состояниях в отсутствие субстратов.[36][37] Отличительной чертой конфигурации ААА-АТФазы в этом преобладающем низкоэнергетическом состоянии является расположение ААА-доменов, напоминающее ступенчатую или стопорную шайбу.[30][31] В присутствии АТФ но при отсутствии субстрата приняты три альтернативные, менее многочисленные конформации 19S, в основном отличающиеся расположением крышки по отношению к модулю ААА-АТФазы.[33][37] В присутствии АТФ-γS или субстрата наблюдается значительно больше конформаций, демонстрирующих драматические структурные изменения модуля ААА-АТФазы.[15][36][38][39] Некоторые из связанных с субстратом конформаций имеют большое сходство с конформациями без субстрата, но они не полностью идентичны, особенно в модуле AAA-ATPase.[15][36] До сборки 26S регуляторная частица 19S в свободной форме также наблюдалась в семи конформационных состояниях.[40] Примечательно, что все эти конформеры несколько отличаются и имеют отличительные особенности. Таким образом, регулирующая частица 19S может отбирать по меньшей мере 20 конформационных состояний в различных физиологических условиях.
Регулирование 20S 19S
Регуляторная частица 19S отвечает за стимуляцию 20S к расщеплению белков. Основная функция 19S регуляторных АТФаз заключается в открытии ворот в 20S, которые блокируют вход субстратов в камеру деградации.[41] Механизм, с помощью которого протеасомная АТФаза открывает эти ворота, был недавно выяснен.[18] Открытие 20S ворот и, следовательно, деградация субстрата требует C-концов протеасомных АТФаз, которые содержат специфический мотив (т.е. мотив HbYX). С-концы АТФазы связываются с карманами в верхней части 20S и привязывают комплекс АТФазы к протеолитическому комплексу 20S, таким образом соединяя оборудование для развертывания субстрата с механизмом деградации 20S. Связывание этих С-концов в эти карманы 20S само по себе стимулирует открытие ворот в 20S во многом так же, как «ключ в замке» открывает дверь.[18] Точный механизм, с помощью которого функционирует этот механизм «ключ-в-замке», был структурно выяснен в контексте протеасомы 26S человека при почти атомном разрешении, предполагая, что вставка пяти С-концов субъединиц АТФазы Rpt1 / 2 / 3/5/6 в поверхностные карманы 20S необходимы для полного открытия затвора 20S.[36][15][33]
Другие регуляторные частицы
Протеасомы 20S могут также связываться со вторым типом регуляторных частиц, регуляторными частицами 11S, гептамерной структурой, которая не содержит никаких АТФаз и может способствовать деградации коротких пептиды но не полных белков. Предполагается, что это связано с тем, что комплекс не может разворачивать более крупные субстраты. Эта структура также известна как PA28, REG или PA26.[17] Механизмы, с помощью которых он связывается с коровой частицей через С-концевые хвосты его субъединиц и индуцирует α-кольцо конформационные изменения открытие ворот 20S предполагает аналогичный механизм для частицы 19S.[42] Экспрессия частицы 11S индуцируется гамма-интерфероном и отвечает, в сочетании с β-субъединицами иммунопротеасомы, за образование пептидов, которые связываются с главный комплекс гистосовместимости.[16]
Еще одним типом нерегулирующих АТФазу частиц является Blm10 (дрожжи) или PA200 /PSME4 (человек). Он открывает только одну α-субъединицу в 20S воротах и сам сворачивается в купол с очень маленькой порой над ним.[17]
сборка
Сборка протеасомы - сложный процесс из-за количества субъединиц, которые должны связываться, чтобы сформировать активный комплекс. Субъединицы β синтезируются с N-концевой "пропептиды", которые посттрансляционно модифицированный во время сборки 20S частицы, чтобы обнажить протеолитический активный центр. Частица 20S собирается из двух полупротеасом, каждая из которых состоит из семичленного про-β-кольца, присоединенного к семичленному α-кольцу. Ассоциация β-колец двух полупротеасом запускает треонин -зависимый автолиз пропептидов, чтобы открыть активный сайт. Эти β-взаимодействия в основном опосредуются соляные мосты и гидрофобный взаимодействия между сохраненными альфа спирали чье нарушение мутация нарушает способность протеасомы к сборке.[43] Сборка полупротеасом, в свою очередь, инициируется сборкой субъединиц α в их гептамерное кольцо, формируя матрицу для ассоциации соответствующего про-β кольца. Сборка субъединиц α не охарактеризована.[44]
Только недавно процесс сборки 19S регуляторной частицы был в значительной степени выяснен. Регуляторная частица 19S состоит из двух отдельных субкомпонентов: основания и крышки. Сборку базового комплекса облегчают четыре сборки шапероны, Hsm3 / S5b, Nas2 / p27, Rpn14 / PAAF1 и Nas6 /ганкирин (названия дрожжей / млекопитающих).[45] Эти сборочные шапероны связываются с AAA-АТФаза субъединиц и их основная функция, по-видимому, заключается в обеспечении правильной сборки гетерогексамерных AAA-АТФаза звенеть. На сегодняшний день все еще ведутся дискуссии о том, собирается ли базовый комплекс по отдельности, является ли сборка шаблоном ядерной частицы 20S или существуют альтернативные пути сборки. Помимо четырех шаперонов сборки, деубиквитинирующий фермент Ubp6 /УСП14 также способствует сборке основания, но это не существенно.[46] Крышка собирается отдельно в определенном порядке и не требует сборочных шаперонов.[47]
Процесс деградации белка

Убиквитинирование и таргетинг
Белки нацелены на деградацию протеасомой с ковалентной модификацией остатка лизина, которая требует скоординированных реакций трех ферменты. На первом этапе убиквитин-активирующий фермент (известный как E1) гидролизует АТФ и аденилилирует убиквитин молекула. Затем он передается на активный сайт E1. цистеин остаток вместе с аденилилированием второго убиквитина.[48] Этот аденилилированный убиквитин затем переносится в цистеин второго фермента, убиквитин-конъюгированный фермент (E2). На последнем этапе член очень разнообразного класса ферментов, известных как убиквитинлигазы (E3) распознает конкретный белок, который должен быть убиквитинирован, и катализирует перенос убиквитина от E2 к этому белку-мишени. Целевой белок должен быть помечен как минимум четырьмя мономерами убиквитина (в форме полиубиквитиновой цепи), прежде чем он будет распознан крышкой протеасомы.[49] Следовательно, именно E3 дает субстрат специфика этой системы.[50] Количество экспрессируемых белков E1, E2 и E3 зависит от организма и типа клеток, но у человека присутствует много различных ферментов E3, что указывает на огромное количество мишеней для протеасомной системы убиквитина.
Механизм, с помощью которого полиубиквитинированный белок нацелен на протеасому, полностью не изучен. Несколько снимков с высоким разрешением протеасомы, связанной с полиубиквитинированным белком, предполагают, что рецепторы убиквитина могут быть скоординированы с деубиквитиназой Rpn11 для начального нацеливания на субстрат и взаимодействия.[15] Белки-рецепторы убиквитина обладают N-концевой убиквитиноподобный (UBL) и один или несколько убиквитин-ассоциированных (UBA) доменов. Домены UBL распознаются крышками протеасомы 19S, а домены UBA связывают убиквитин через трехспиральные пучки. Эти рецепторные белки могут сопровождать полиубиквитинированные белки в протеасому, хотя специфика этого взаимодействия и его регуляция неясны.[51]
В убиквитин сам белок - 76 аминокислоты долго и был назван из-за его повсеместного распространения, так как он консервированный последовательность и встречается во всех известных эукариотических организмах.[52] Гены, кодирующие убиквитин в эукариоты расположены в тандем повторяет, возможно, из-за тяжелого транскрипция требует от этих генов производить достаточное количество убиквитина для клетки. Было высказано предположение, что убиквитин является самым медленнымразвивающийся белок, идентифицированный на сегодняшний день.[53] Убиквитин содержит семь остатков лизина, с которыми может быть лигирован другой убиквитин, что приводит к различным типам полиубиквитиновых цепей.[54] Цепи, в которых каждый дополнительный убиквитин связан с лизином 48 предыдущего убиквитина, играют роль в нацеливании на протеасомы, тогда как другие типы цепей могут участвовать в других процессах.[55][56]

Разворачивание и перемещение
После того, как белок был убиквитинирован, он распознается регуляторной частицей 19S на стадии АТФ-зависимого связывания.[15][28] Затем белок-субстрат должен войти внутрь частицы 20S, чтобы войти в контакт с протеолитическими активными центрами. Поскольку центральный канал частицы 20S является узким и закрывается N-концевыми хвостами субъединиц α кольца, субстраты должны быть по крайней мере частично развернуты, прежде чем они войдут в ядро.[15] Прохождение развернутой подложки в сердцевину называется перемещение и обязательно происходит после деубиквитинирования.[15][28] Однако порядок, в котором субстраты деубиквитинируются и разворачиваются, еще не ясен.[57] Какой из этих процессов является ограничивающий шаг в целом реакция протеолиза зависит от конкретного субстрата; для некоторых белков процесс разворачивания ограничивает скорость, в то время как деубиквитинирование является самым медленным шагом для других белков.[27] Предполагается, что степень, в которой субстраты должны быть развернуты перед транслокацией, составляет около 20 аминокислотных остатков из-за атомной структуры вовлеченной в субстрат 26S протеасомы в состоянии, совместимом с деубиквитилированием.[15] но существенный третичная структура, и, в частности, нелокальные взаимодействия, такие как дисульфидные связи, достаточны для подавления разложения.[58] Наличие внутренне неупорядоченный белок сегменты достаточного размера, либо на конце белка, либо внутри, также были предложены для облегчения эффективного инициирования деградации.[59][60]
Гейт, образованный субъединицами α, предотвращает попадание пептидов длиной более четырех остатков внутрь частицы 20S. Молекулы АТФ, связанные до начальной стадии распознавания, являются гидролизованный до транслокации. Хотя энергия необходима для развертывания субстрата, она не требуется для транслокации.[27][28] Собранная протеасома 26S может разрушать развернутые белки в присутствии негидролизуемого Аналог АТФ, но не может разлагать свернутые белки, указывая на то, что энергия гидролиза АТФ используется для развертывания субстрата.[27] Прохождение развернутой подложки через открытый затвор происходит через облегченное распространение если крышка 19S находится в АТФ-связанном состоянии.[61]
Механизм раскладывания глобулярные белки обязательно общий, но в некоторой степени зависит от аминокислотная последовательность. Длинные последовательности чередования глицина и аланин было показано, что они ингибируют развертывание субстрата, снижая эффективность протеасомной деградации; это приводит к высвобождению частично разложившихся побочных продуктов, возможно, из-за разделения стадий гидролиза и разворачивания АТФ.[62] Такие глицин-аланиновые повторы также встречаются в природе, например, в шелк фиброин; в частности, некоторые Вирус Эпштейна-Барра генные продукты, несущие эту последовательность, могут остановить протеасому, помогая вирусу размножаться, предотвращая презентация антигена по главному комплексу гистосовместимости.[63]
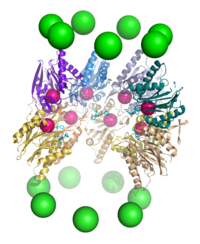
Протеолиз
Протеасома функционирует как эндопротеаза.[64][65][66][67] Механизм протеолиза β-субъединицами коровой частицы 20S осуществляется через треонин-зависимую нуклеофильная атака. Этот механизм может зависеть от связанного воды молекула для депротонирования реактивного треонина гидроксил. Деградация происходит в центральной камере, образованной объединением двух β-колец, и обычно не высвобождает частично разрушенные продукты, вместо этого восстанавливая субстрат до коротких полипептидов, обычно длиной 7–9 остатков, хотя они могут составлять от 4 до 25 остатков, в зависимости от организм и субстрат. Биохимический механизм, определяющий длину продукта, полностью не охарактеризован.[68] Хотя три каталитических β-субъединицы имеют общий механизм, они имеют немного разные субстратные специфичности, которые считаются химотрипсиноподобными, трипсиноподобными и пептидилглутамилпептидными (PHGH) -подобными. Эти вариации специфичности являются результатом межатомных контактов с локальными остатками около активных сайтов каждой субъединицы. Каждая каталитическая β-субъединица также имеет консервативный остаток лизина, необходимый для протеолиза.[22]
Хотя протеасома обычно продуцирует очень короткие пептидные фрагменты, в некоторых случаях эти продукты сами являются биологически активными и функциональными молекулами. Определенный факторы транскрипции регулирование экспрессии определенных генов, включая один компонент комплекса млекопитающих NF-κB, синтезируются как неактивные предшественники, убиквитинирование которых и последующая протеасомная деградация превращают их в активную форму. Такая активность требует, чтобы протеасома внутренне расщепляла субстратный белок, а не процессивно разрушала его с одного конца. Было высказано предположение, что долго петли на поверхности этих белков служат протеасомными субстратами и проникают в центральную полость, в то время как большая часть белка остается снаружи.[69] Подобные эффекты наблюдались в дрожжевых белках; этот механизм избирательной деградации известен как регулируемый процессинг, зависящий от убиквитина / протеасомы (RUP).[70]
Убиквитин-независимая деградация
Хотя большинство протеасомных субстратов необходимо убиквитинировать перед деградацией, есть некоторые исключения из этого общего правила, особенно когда протеасома играет нормальную роль в пост-переводной переработка белка. Протеасомная активация NF-κB путем обработки p105 в p50 посредством внутреннего протеолиза - один из основных примеров.[69] Некоторые белки, которые предположительно нестабильны из-за внутренне неструктурированный регионы,[71] деградируют убиквитин-независимым образом. Наиболее известным примером убиквитин-независимого протеасомного субстрата является фермент орнитиндекарбоксилаза.[72] Убиквитин-независимые механизмы нацеливания на ключ клеточный цикл регуляторы, такие как p53 также сообщалось, хотя р53 также подвержен убиквитин-зависимой деградации.[73] Наконец, структурно аномальные, неправильно свернутые или сильно окисленные белки также подвержены убиквитин-независимой и 19S-независимой деградации в условиях клеточного стресса.[74]
Эволюция

Протеасома 20S встречается повсеместно и незаменима у эукариот. Немного прокариоты, в том числе многие археи и бактериальный порядок Актиномицеты, также имеют общие гомологи протеасомы 20S, тогда как большинство бактерий обладают тепловой удар гены hslV и hslU, генными продуктами которого являются мультимерная протеаза, расположенная в двухслойном кольце, и АТФаза.[75] Было высказано предположение, что белок hslV похож на вероятного предка протеасомы 20S.[76] В целом, HslV не важен для бактерий, и не все бактерии обладают им, в то время как некоторые протисты обладают как 20S, так и hslV системами.[75] Многие бактерии также обладают другими гомологами протеасомы и связанной с ней АТФазы, в первую очередь ClpP и ClpX. Эта избыточность объясняет, почему система HslUV не важна.
Анализ последовательности предполагает, что каталитические β-субъединицы расходились раньше в эволюции, чем преимущественно структурные α-субъединицы. У бактерий, экспрессирующих протеасому 20S, субъединицы β имеют высокий идентичность последовательности с субъединицами β архей и эукариот, тогда как идентичность последовательности α намного ниже. Наличие протеасом 20S в бактериях может быть результатом боковой перенос гена, а диверсификация субъединиц среди эукариот объясняется множеством дупликация гена События.[75]
Контроль клеточного цикла
Прогрессирование клеточного цикла контролируется упорядоченным действием циклин-зависимые киназы (CDK), активируется определенным циклины которые разграничивают фазы клеточный цикл. Митотические циклины, которые сохраняются в клетке всего несколько минут, имеют один из самых коротких периодов жизни среди всех внутриклеточных белков.[1] После того, как комплекс CDK-циклин выполнил свою функцию, связанный циклин полиубиквитинируется и разрушается протеасомой, которая обеспечивает направленность клеточного цикла. В частности, выход из митоз требует протеасомозависимой диссоциации регуляторного компонента циклин B от фактор, способствующий митозу сложный.[77] В позвоночное животное клетки, «проскальзывание» через митотическую контрольную точку, приводящее к преждевременному Фаза M выход может произойти, несмотря на задержку этого выхода из-за КПП шпинделя.[78]
Более ранние контрольные точки клеточного цикла, такие как пост-точка ограничения проверить между грамм1 фаза и S фаза аналогичным образом включают протеасомную деградацию циклин А, убиквитинизации которого способствует комплекс, способствующий анафазе (APC), E3 убиквитинлигаза.[79] APC и белковый комплекс Skp1 / Cul1 / F-бокс (СКФ комплекс ) являются двумя ключевыми регуляторами деградации циклинов и контроля контрольных точек; сам SCF регулируется APC посредством убиквитинирования адаптерного белка Skp2, который предотвращает активность SCF перед переходом G1-S.[80]
Отдельные компоненты частицы 19S имеют свои собственные регулирующие роли. Ганкырин, недавно обнаруженный онкопротеин, является одним из подкомпонентов 19S, который также прочно связывает циклин-зависимая киназа CDK4 и играет ключевую роль в распознавании убиквитинированных p53, через его сродство к убиквитинлигазе MDM2. Ганкирин анти-апоптотический и было показано, что в некоторых опухоль типы клеток, такие как гепатоцеллюлярная карцинома.[81]
Регулирование роста растений
В растения, сигнализация ауксины, или же фитогормоны что порядок направления и тропизм роста растений, вызывает нацеливание на класс фактор транскрипции репрессоры, известные как белки Aux / IAA для протеасомной деградации. Эти белки убиквитинируются SCFTIR1 или SCF в комплексе с рецептором ауксина TIR1. Деградация белков Aux / IAA дерепрессирует факторы транскрипции в семействе факторов ауксинового ответа (ARF) и индуцирует ARF-направленную экспрессию генов.[82] Клеточные последствия активации ARF зависят от типа растения и стадии развития, но участвуют в управлении ростом корней и жилок листа. Считается, что специфический ответ на дерепрессию ARF опосредуется специфичностью пары отдельных белков ARF и Aux / IAA.[83]
Апоптоз
Как внутренние, так и внешние сигналы может привести к индукции апоптоз, или запрограммированная гибель клеток. Результирующая деконструкция клеточных компонентов в основном осуществляется специализированными протеазами, известными как каспасы, но протеасома также играет важные и разнообразные роли в процессе апоптоза. На участие протеасомы в этом процессе указывает как усиление убиквитинирования белка, так и ферментов E1, E2 и E3, которое наблюдается задолго до апоптоза.[84][85][86] Во время апоптоза протеасомы, локализованные в ядре, также могут перемещаться на внешнюю мембрану. пузыри характеристика апоптоза.[87]
Ингибирование протеасом по-разному влияет на индукцию апоптоза в разных типах клеток. В общем, протеасома не требуется для апоптоза, хотя ее ингибирование проапоптозно для большинства типов клеток, которые были изучены. Апоптоз опосредуется нарушением регулируемой деградации белков клеточного цикла, способствующих росту.[88] Однако некоторые клеточные линии, в частности, первичные культуры из неподвижный и дифференцированный клетки, такие как тимоциты и нейроны - предотвращается апоптоз при воздействии ингибиторов протеасом. Механизм этого эффекта не ясен, но предполагается, что он специфичен для клеток в состоянии покоя или является результатом различной активности проапоптотических киназа JNK.[89] Способность ингибиторов протеасом индуцировать апоптоз в быстро делящихся клетках была использована в нескольких недавно разработанных химиотерапия такие агенты, как бортезомиб и салиноспорамид А.
Ответ на клеточный стресс
В ответ на клеточные стрессы, такие как инфекционное заболевание, тепловой удар, или же окислительное повреждение – белки теплового шока которые идентифицируют неправильно свернутые или развернутые белки и нацелены на их протеасомную деградацию. Обе Hsp27 и Hsp90 —сопровождающий белки участвуют в увеличении активности убиквитин-протеасомной системы, хотя они не являются прямыми участниками этого процесса.[90] Hsp70, с другой стороны, связывает открытые гидрофобный пятна на поверхности неправильно свернутых белков и рекрутирует E3 ubiquitin ligases, такие как CHIP, чтобы пометить белки для протеасомной деградации.[91] Белок CHIP (карбоксильный конец белка, взаимодействующего с Hsp70) сам регулируется посредством ингибирования взаимодействий между ферментом CHIP E3 и его партнером по связыванию E2.[92]
Подобные механизмы существуют, чтобы способствовать деградации окислительно поврежденный белки через протеасомную систему. В частности, протеасомы, локализованные в ядре, регулируются PARP и активно разлагают неправильно окисленные гистоны.[93] Окисленные белки, которые часто образуют большие аморфные агрегаты в клетке, могут быть разложены непосредственно ядром 20S частицы без регуляторного колпачка 19S и не требуют гидролиза АТФ или мечения убиквитином.[74] Однако высокий уровень окислительного повреждения увеличивает степень сшивки между фрагментами белка, делая агрегаты устойчивыми к протеолизу. Большее количество и размер таких сильно окисленных агрегатов связаны с старение.[94]
Нарушение регуляции протеасомной системы убиквитина может способствовать возникновению нескольких нервных заболеваний. Это может привести к опухолям головного мозга, таким как: астроцитомы.[95] В некоторых случаях с поздним началом нейродегенеративный заболевания, при которых агрегация неправильно свернутых белков является общим признаком, например болезнь Паркинсона и Болезнь Альцгеймера, могут образовываться большие нерастворимые агрегаты неправильно свернутых белков, что приводит к нейротоксичность через механизмы, которые еще недостаточно изучены. Снижение активности протеасом было предложено как причина агрегации и Тело Леви образование при болезни Паркинсона.[96] Эта гипотеза подтверждается наблюдением, что дрожжи модели болезни Паркинсона более восприимчивы к токсичности от α-синуклеин, основной белковый компонент телец Леви, в условиях низкой протеасомной активности.[97] Нарушение протеасомной активности может лежать в основе когнитивных расстройств, таких как расстройства аутистического спектра, а также мышечные и нервные заболевания, такие как миопатия с тельцами включения.[95]
Роль в иммунной системе
Протеасома играет прямую, но решающую роль в функции адаптивная иммунная система. Пептид антигены отображаются главный комплекс гистосовместимости белки класса I (MHC) на поверхности антигенпрезентирующие клетки. Эти пептиды являются продуктами протеасомной деградации белков, возникших в результате вторжения. возбудитель. Хотя конститутивно экспрессируемые протеасомы могут участвовать в этом процессе, специализированный комплекс, состоящий из белков, чьи выражение индуцируется интерферон гамма, являются основными продуцентами пептидов, которые оптимальны по размеру и составу для связывания MHC. Эти белки, экспрессия которых увеличивается во время иммунного ответа, включают регуляторную частицу 11S, основная известная биологическая роль которой регулирует продукцию лигандов MHC, и специализированные субъединицы β, называемые β1i, β2i и β5i, с измененной субстратной специфичностью. Комплекс, образованный специализированными субъединицами β, известен как иммунопротеасома.[16] Другая субъединица варианта β5i, β5t, экспрессируется в тимусе, приводя к тимус-специфической «тимопротеасоме», функция которой пока неясна.[98]
Сила связывания лиганда MHC класса I зависит от состава лиганда. C-конец, поскольку пептиды связываются водородная связь и путем тесного контакта с областью, называемой «карманом B» на поверхности MHC. Многие аллели MHC класса I предпочитают гидрофобные С-концевые остатки, и иммунопротеасомный комплекс с большей вероятностью генерирует гидрофобные С-концы.[99]
Благодаря своей роли в генерации активированной формы NF-κB, анти-апоптотический и про-воспалительный регулятор цитокин экспрессии протеасомная активность была связана с воспалительными и аутоиммунные заболевания. Повышенные уровни протеасомной активности коррелируют с активностью заболевания и связаны с аутоиммунными заболеваниями, включая системная красная волчанка и ревматоидный артрит.[16]
Протеасома также участвует в Внутриклеточный протеолиз, опосредованный антителами связанных с антителами вирионов. На этом пути нейтрализации TRIM21 (белок из семейства трехчастных мотивов) связывается с иммуноглобулин G направить вирион к протеасоме, где он расщепляется.[100]
Ингибиторы протеасом
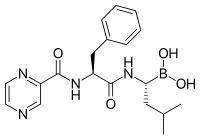
Ингибиторы протеасом иметь эффективные анти-опухоль деятельность в культура клеток, побуждая апоптоз путем нарушения регулируемой деградации белков клеточного цикла, способствующих росту.[88] Этот подход избирательного индуцирования апоптоза в опухолевых клетках доказал свою эффективность на животных моделях и в испытаниях на людях.
Лактацистин, натуральный продукт, синтезированный Streptomyces бактерии, был первым обнаруженным непептидным ингибитором протеасом.[101] и широко используется в качестве исследовательского инструмента в биохимии и клеточной биологии. Лицензия на лактацистин была передана компании Myogenics / Proscript, которая была приобретена Millennium Pharmaceuticals, теперь часть Takeda Pharmaceuticals. Лактацистин ковалентно модифицирует амино-концевой треонин каталитических β-субъединиц протеасомы, особенно β5-субъединицу, отвечающую за химотрипсиноподобную активность протеасомы. Это открытие помогло установить протеасому как новый механистически новый класс протеаз: аминоконцевой треониновая протеаза.
Бортезомиб (Боронированный MG132), молекула, разработанная Millennium Pharmaceuticals и продаваемый как Velcade, является первым ингибитором протеасом, который получил клиническое применение в качестве химиотерапия агент.[102] Бортезомиб используется при лечении: множественная миелома.[103] Примечательно, что множественная миелома приводит к увеличению уровней пептидов, производных от протеасом, в сыворотка крови которые снижаются до нормального уровня в ответ на успешную химиотерапию.[104] Исследования на животных показали, что бортезомиб также может оказывать клинически значимое воздействие на панкреатический рак.[105][106] Были начаты доклинические и ранние клинические исследования для изучения эффективности бортезомиба при лечении других заболеваний. В-клетка -связанные раки,[107] особенно некоторые виды неходжкинской лимфомы.[108] Клинические результаты также, кажется, оправдывают использование ингибитора протеасом в сочетании с химиотерапией при В-клеточном остром лимфобластном лейкозе. [109] Ингибиторы протеасом могут убивать некоторые типы культивируемых лейкозных клеток, устойчивых к глюкокортикоидам.[110]
Молекула ритонавир, продаваемый как Norvir, был разработан как ингибитор протеазы и используется для нацеливания ВИЧ инфекционное заболевание. Однако было показано, что он ингибирует протеасомы, а также свободные протеазы; чтобы быть конкретным, химотрипсин -подобная активность протеасомы ингибируется ритонавиром, в то время как трипсин -подобная активность несколько усиливается.[111] Исследования на животных моделях показывают, что ритонавир может оказывать ингибирующее действие на рост глиома клетки.[112]
Ингибиторы протеасом также показали себя многообещающими при лечении аутоиммунных заболеваний на животных моделях. Например, исследования на мышах, несущих человека кожные трансплантаты обнаружили уменьшение размеров поражений от псориаз после лечения ингибитором протеасом.[113] Ингибиторы также оказывают положительное влияние на грызун модели астма.[114]
Мечение и ингибирование протеасомы также представляет интерес в лабораторных условиях для обоих in vitro и in vivo изучение протеасомальной активности в клетках. Наиболее часто используемые лабораторные ингибиторы: лактацистин и пептидный альдегид MG132 изначально разработан лабораторией Голдберга. Флуоресцентный ингибиторы также были разработаны для специфической маркировки активных сайтов собранной протеасомы.[115]
Клиническое значение
Протеасома и ее субъединицы имеют клиническое значение по крайней мере по двум причинам: (1) нарушенная комплексная сборка или дисфункциональная протеасома может быть связана с патофизиологией конкретных заболеваний, и (2) они могут использоваться в качестве мишеней для лекарств для терапевтических целей. вмешательства. Совсем недавно были предприняты дополнительные усилия по рассмотрению протеасомы для разработки новых диагностических маркеров и стратегий. Улучшенное и всестороннее понимание патофизиологии протеасомы должно привести к клиническому применению в будущем.
Протеасомы образуют ключевой компонент убиквитин-протеасомной системы (UPS). [116] и соответствующий контроль качества клеточного белка (PQC). Протеин убиквитинирование и последующие протеолиз и деградация протеасомами являются важными механизмами в регуляции клеточный цикл, рост клеток и дифференцировка, транскрипция генов, сигнальная трансдукция и апоптоз.[117] Впоследствии нарушение сборки и функции протеасомного комплекса ведет к снижению протеолитической активности и накоплению поврежденных или неправильно свернутых белков. Такое накопление белка может способствовать патогенезу и фенотипическим характеристикам нейродегенеративных заболеваний,[118][119] сердечно-сосудистые заболевания,[120][121][122] воспалительные реакции и аутоиммунные заболевания,[123] и системные реакции на повреждение ДНК, приводящие к злокачественные новообразования.[124]
Несколько экспериментальных и клинических исследований показали, что аберрации и нарушение регуляции UPS вносят вклад в патогенез нескольких нейродегенеративных и миодегенеративных заболеваний, включая Болезнь Альцгеймера,[125] болезнь Паркинсона[126] и Болезнь Пика,[127] боковой амиотрофический склероз (ALS),[127] болезнь Хантингтона,[126] Болезнь Крейтцфельдта-Якоба,[128] болезни мотонейронов, полиглутаминовые (PolyQ) заболевания, мышечные дистрофии[129] и несколько редких форм нейродегенеративных заболеваний, связанных с слабоумие.[130] Как часть убиквитин-протеасомной системы (UPS), протеасома поддерживает гомеостаз сердечного белка и, таким образом, играет важную роль в сердечном белке. ишемический травма, повреждение,[131] гипертрофия желудочков[132] и сердечная недостаточность.[133] Кроме того, накапливаются доказательства того, что UPS играет важную роль в злокачественной трансформации. Протеолиз UPS играет важную роль в ответах раковых клеток на стимулирующие сигналы, которые имеют решающее значение для развития рака. Соответственно, экспрессия гена за счет деградации факторы транскрипции, Такие как p53, с-июн, c-Fos, NF-κB, c-Myc, HIF-1α, MATα2, STAT3, стерол-регулируемые связывающие элементы белки и рецепторы андрогенов Все они контролируются ИБП и, таким образом, участвуют в развитии различных злокачественных новообразований.[134] Кроме того, UPS регулирует деградацию продуктов гена-супрессора опухолей, таких как аденоматозный полипоз кишечной палочки (APC) при колоректальном раке, ретинобластома (Rb). и опухолевый супрессор фон Хиппеля – Линдау (ВХЛ), а также ряд протоонкогены (Раф, Мой с, Myb, Rel, Src, Мос, ABL ). ИБП также участвует в регуляции воспалительных реакций. Эта активность обычно объясняется ролью протеасом в активации NF-κB, который дополнительно регулирует экспрессию провоспалительных цитокины Такие как TNF-α, ИЛ-β, Ил-8, молекулы адгезии (ICAM-1, VCAM-1, Р-селектин ) и простагландины и оксид азота (НЕТ).[123] Кроме того, UPS также играет роль в воспалительных реакциях в качестве регуляторов пролиферации лейкоцитов, в основном за счет протеолиза циклинов и деградации CDK ингибиторы.[135] Наконец, аутоиммунное заболевание пациенты с SLE, Синдром Шегрена и ревматоидный артрит (RA) преимущественно демонстрируют циркулирующие протеасомы, которые можно использовать в качестве клинических биомаркеров.[136]
Смотрите также
- Карта протеолиза
- Экзосомный комплекс
- Расщепление белков, связанное с эндоплазматическим ретикулумом
- JUNQ и IPOD
Рекомендации
- ^ а б c d Лодиш Х, Берк А., Мацудаира П., Кайзер Калифорния, Кригер М., Скотт М. П., Зипурски С. Л., Дарнелл Дж. (2004). "3". Молекулярная клеточная биология (5-е изд.). Нью-Йорк: W.H. Freeman and CO. Стр.66–72. ISBN 978-0-7167-4366-8.
- ^ Петерс JM, Franke WW, Kleinschmidt JA (март 1994). «Отличительные 19 S и 20 S субкомплексы 26 S протеасомы и их распределение в ядре и цитоплазме». Журнал биологической химии. 269 (10): 7709–18. PMID 8125997.
- ^ Нассиф, Николас Д .; Cambray, Samantha E .; Краут, Даниэль А. (май 2014 г.). «Скольжение: частичная деградация субстрата АТФ-зависимыми протеазами». IUBMB Life. 66 (5): 309–317. Дои:10.1002 / iub.1271. PMID 24823973. S2CID 29860298.
- ^ а б Комитет по Нобелевской премии (2004). «Лауреаты Нобелевской премии по химии 2004 г.». Получено 11 декабря 2006.
- ^ Этлингер Дж. Д., Гольдберг А. Л. (январь 1977 г.). «Растворимая АТФ-зависимая протеолитическая система, ответственная за деградацию аномальных белков в ретикулоцитах». Труды Национальной академии наук Соединенных Штатов Америки. 74 (1): 54–8. Bibcode:1977PNAS ... 74 ... 54E. Дои:10.1073 / пнас.74.1.54. ЧВК 393195. PMID 264694.
- ^ Ciehanover A, Hod Y, Hershko A (апрель 1978 г.). «Термостойкий полипептидный компонент АТФ-зависимой протеолитической системы из ретикулоцитов». Сообщения о биохимических и биофизических исследованиях. 81 (4): 1100–5. Дои:10.1016 / 0006-291X (78) 91249-4. PMID 666810.
- ^ Goldknopf IL, Busch H (март 1977 г.). «Изопептидная связь между полипептидами негистонового гистона и гистона 2А хромосомного конъюгата-протеина А24». Труды Национальной академии наук Соединенных Штатов Америки. 74 (3): 864–8. Bibcode:1977ПНАС ... 74..864Г. Дои:10.1073 / пнас.74.3.864. ЧВК 430507. PMID 265581.
- ^ Цехановер А (сентябрь 2005 г.). «Ранняя работа над системой убиквитиновых протеасом, интервью с Аароном Чехановером. Интервью CDD». Гибель клеток и дифференциация. 12 (9): 1167–77. Дои:10.1038 / sj.cdd.4401691. PMID 16094393.
- ^ Вилк С., Орловский М. (ноябрь 1980 г.). «Катион-чувствительная нейтральная эндопептидаза: выделение и специфичность фермента гипофиза крупного рогатого скота». Журнал нейрохимии. 35 (5): 1172–82. Дои:10.1111 / j.1471-4159.1980.tb07873.x. PMID 6778972. S2CID 9028201.
- ^ Арриго А.П., Танака К., Гольдберг Ф., Уэлч В.Дж. (1988). «Идентичность частицы просомы 19S с большим многофункциональным протеазным комплексом клеток млекопитающих». Природа. 331 (6152): 192–94. Дои:10.1038 / 331192a0. PMID 3277060. S2CID 97688.Танака К., Ваксман Л., Гольдберг А.Л. (июнь 1983 г.). «АТФ выполняет две разные роли в деградации белка в ретикулоцитах, одна требует и другая независима от убиквитина». Журнал клеточной биологии. 96 (6): 1580–5. Дои:10.1083 / jcb.96.6.1580. ЧВК 2112434. PMID 6304111.
- ^ Hough R, Pratt G, Rechsteiner M (июнь 1987 г.). «Очистка двух высокомолекулярных протеаз из лизата ретикулоцитов кролика». Журнал биологической химии. 262 (17): 8303–13. PMID 3298229.
- ^ Гершко А. (сентябрь 2005 г.). «Ранняя работа над системой убиквитиновых протеасом, интервью с Аврамом Хершко. Интервью CDD». Гибель клеток и дифференциация. 12 (9): 1158–61. Дои:10.1038 / sj.cdd.4401709. PMID 16094391.
- ^ Копп Ф., Штайнер Р., Дальманн Б., Куен Л., Рейнауэр Х. (август 1986 г.). «Размер и форма поликаталитической протеиназы из скелетных мышц крысы». Biochimica et Biophysica Acta (BBA) - Структура белка и молекулярная энзимология. 872 (3): 253–60. Дои:10.1016/0167-4838(86)90278-5. PMID 3524688.
- ^ Лёве Дж., Сток Д., Яп Б., Цвикль П., Баумейстер В., Хубер Р. (апрель 1995 г.). «Кристаллическая структура протеасомы 20S из архей T. acidophilum при разрешении 3,4 A». Наука. 268 (5210): 533–9. Дои:10.1126 / science.7725097. PMID 7725097.
- ^ а б c d е ж грамм час я j k Донг И, Чжан С., Ву З., Ли Х, Ван В.Л., Чжу И, Стойлова-Макфи С., Лу И, Финли Д., Мао Ю. (ноябрь 2018 г.). «Крио-ЭМ структуры и динамика субстрат-задействованной протеасомы 26S человека». Природа. 565 (7737): 49–55. Дои:10.1038 / s41586-018-0736-4. ЧВК 6370054. PMID 30479383.
- ^ а б c d Ван Дж, Мальдонадо, Массачусетс (август 2006 г.). «Убиквитин-протеасомная система и ее роль в воспалительных и аутоиммунных заболеваниях». Клеточная и молекулярная иммунология. 3 (4): 255–61. PMID 16978533.
- ^ а б c Stadtmueller, BM; Хилл, КП (7 января 2011 г.). «Активаторы протеасом». Молекулярная клетка. 41 (1): 8–19. Дои:10.1016 / j.molcel.2010.12.020. ЧВК 3040445. PMID 21211719.
- ^ а б c Смит Д.М., Чанг С.К., Парк С., Финли Д., Ченг Ю., Голдберг А. Л. (сентябрь 2007 г.). «Стыковка карбоксильных концов протеасомных АТФаз в альфа-кольце протеасомы 20S открывает ворота для входа в субстрат». Молекулярная клетка. 27 (5): 731–44. Дои:10.1016 / j.molcel.2007.06.033. ЧВК 2083707. PMID 17803938.
- ^ "МЕРОПС Семейный Т1". EMBL-EBI. Получено 16 февраля 2019.
- ^ Вилк С., Орловский М. (март 1983 г.). «Доказательства того, что нейтральная эндопептидаза, чувствительная к катионам гипофиза, представляет собой многокаталитический протеазный комплекс». Журнал нейрохимии. 40 (3): 842–9. Дои:10.1111 / j.1471-4159.1983.tb08056.x. PMID 6338156. S2CID 23508675.
- ^ а б Нанди Д., Тахилиани П., Кумар А., Чанду Д. (март 2006 г.). «Убиквитин-протеасомная система» (PDF). Журнал биологических наук. 31 (1): 137–55. Дои:10.1007 / BF02705243. PMID 16595883. S2CID 21603835.
- ^ а б Хайнемайер В., Фишер М., Криммер Т., Стахон Ю., Вольф Д.Х. (октябрь 1997 г.). «Активные центры 20 S протеасомы эукариот и их участие в процессинге субъединиц-предшественников». Журнал биологической химии. 272 (40): 25200–9. Дои:10.1074 / jbc.272.40.25200. PMID 9312134.
- ^ а б Падманабхан А., Вуонг С.А., Хохштрассер М. (март 2016 г.). «Сборка эволюционно консервативной альтернативной изоформы протеасомы в клетках человека». Отчеты по ячейкам. 14 (12): 2962–74. Дои:10.1016 / j.celrep.2016.02.068. ЧВК 4828729. PMID 26997268.
- ^ Величутина И., Коннерли П.Л., Арендт С.С., Ли Х, Хохштрассер М. (февраль 2004 г.). «Пластичность сборки 20S протеасомного кольца эукариот, выявленная делецией субъединицы в дрожжах». Журнал EMBO. 23 (3): 500–10. Дои:10.1038 / sj.emboj.7600059. ЧВК 1271798. PMID 14739934.
- ^ Kusmierczyk AR, Kunjappu MJ, Funakoshi M, Hochstrasser M (март 2008 г.). «Мультимерный фактор сборки контролирует образование альтернативных протеасом 20S». Структурная и молекулярная биология природы. 15 (3): 237–44. Дои:10.1038 / nsmb.1389. PMID 18278055. S2CID 21181637.
- ^ Цвикл П., Нг Д., Ву К.М., Кленк Х.П., Голдберг А.Л. (сентябрь 1999 г.). «Архебактериальная АТФаза, гомологичная АТФазам в эукариотической протеасоме 26 S, активирует расщепление белка 20 S протеасомами». Журнал биологической химии. 274 (37): 26008–14. Дои:10.1074 / jbc.274.37.26008. PMID 10473546.
- ^ а б c d Смит Д.М., Кафри Дж., Ченг Й., Нг Д., Вальц Т., Голдберг А.Л. (декабрь 2005 г.). «Связывание АТФ с PAN или 26S АТФазами вызывает ассоциацию с протеасомой 20S, открытие ворот и транслокацию развернутых белков». Молекулярная клетка. 20 (5): 687–98. Дои:10.1016 / j.molcel.2005.10.019. PMID 16337593.
- ^ а б c d е Лю Ч.В., Ли Х, Томпсон Д., Вудинг К., Чанг Т.Л., Тан З., Ю Х., Томас П.Дж., ДеМартино Г.Н. (октябрь 2006 г.). «Связывание АТФ и гидролиз АТФ играют разные роли в функции протеасомы 26S». Молекулярная клетка. 24 (1): 39–50. Дои:10.1016 / j.molcel.2006.08.025. ЧВК 3951175. PMID 17018291.
- ^ Лам Ю.А., Лоусон Т.Г., Велаютам М., Цвейер Дж.Л., Пикарт С.М. (апрель 2002 г.). «Субъединица протеасомной АТФазы распознает сигнал деградации полиубиквитина». Природа. 416 (6882): 763–7. Bibcode:2002Натурал.416..763Л. Дои:10.1038 / 416763a. PMID 11961560. S2CID 4421764.
- ^ а б Beck F, Unverdorben P, Bohn S, Schweitzer A, Pfeifer G, Sakata E, Nickell S, Plitzko JM, Villa E, Baumeister W, Förster F (сентябрь 2012 г.). «Структурная модель 26S протеасомы дрожжей с почти атомным разрешением». Труды Национальной академии наук Соединенных Штатов Америки. 109 (37): 14870–5. Bibcode:2012PNAS..10914870B. Дои:10.1073 / pnas.1213333109. ЧВК 3443124. PMID 22927375.
- ^ а б Lander GC, Estrin E, Matyskiela ME, Bashore C, Nogales E, Martin A (февраль 2012 г.). «Полная субъединичная архитектура регуляторной частицы протеасомы». Природа. 482 (7384): 186–91. Bibcode:2012Натура 482..186л. Дои:10.1038 / природа10774. ЧВК 3285539. PMID 22237024.
- ^ Lasker K, Förster F, Bohn S, Walzthoeni T, Villa E, Unverdorben P, Beck F, Aebersold R, Sali A, Baumeister W. (январь 2012 г.). «Молекулярная архитектура голокомплекса 26S протеасомы, определенная интегративным подходом». Труды Национальной академии наук Соединенных Штатов Америки. 109 (5): 1380–7. Дои:10.1073 / pnas.1120559109. ЧВК 3277140. PMID 22307589.
- ^ а б c Chen S, Wu J, Lu Y, Ma YB, Lee BH, Yu Z, Ouyang Q, Finley DJ, Kirschner MW, Mao Y (ноябрь 2016 г.). «Структурная основа динамической регуляции протеасомы 26S человека». Труды Национальной академии наук Соединенных Штатов Америки. 113 (46): 12991–12996. Дои:10.1073 / pnas.1614614113. ЧВК 5135334. PMID 27791164.
- ^ Хуан X, Луань Б., Ву Дж, Ши И (сентябрь 2016 г.). «Атомная структура протеасомы человека 26S». Структурная и молекулярная биология природы. 23 (9): 778–785. Дои:10.1038 / nsmb.3273. PMID 27428775. S2CID 21909333.
- ^ Швейцер А., Ауфдерхайде А., Рудак Т., Бек Ф., Пфейфер Дж., Плитцко Дж. М., Саката Е., Шультен К., Фёрстер Ф., Баумейстер В. (июль 2016 г.). «Структура протеасомы 26S человека с разрешением 3,9 Å». Труды Национальной академии наук Соединенных Штатов Америки. 113 (28): 7816–7821. Дои:10.1073 / pnas.1614614113. ЧВК 5135334. PMID 27791164.
- ^ а б c d Чжу И, Ван В.Л., Ю Д, Оуян Цюй, Лу И, Мао И (апрель 2018 г.). «Структурный механизм регулируемого нуклеотидами ремоделирования разворачивания ААА-АТФазы в активированной протеасоме 26S человека». Nature Communications. 9 (1): 1360. Bibcode:2018НатКо ... 9.1360Z. Дои:10.1038 / s41467-018-03785-w. ЧВК 5893597. PMID 29636472.
- ^ а б c Unverdorben P, Beck F, Śledź P, Schweitzer A, Pfeifer G, Plitzko JM, Baumeister W, Förster F (апрель 2014 г.). «Глубокая классификация большого набора данных крио-ЭМ определяет конформационный ландшафт протеасомы 26S». Труды Национальной академии наук Соединенных Штатов Америки. 111 (15): 5544–9. Bibcode:2014ПНАС..111.5544У. Дои:10.1073 / pnas.1403409111. ЧВК 3992697. PMID 24706844.
- ^ Ledź P, Unverdorben P, Beck F, Pfeifer G, Schweitzer A, Förster F, Baumeister W (апрель 2013 г.). «Структура протеасомы 26S со связанным АТФ-γS дает представление о механизме нуклеотид-зависимой транслокации субстрата». Труды Национальной академии наук Соединенных Штатов Америки. 110 (18): 7264–7269. Bibcode:2013ПНАС..110.7264С. Дои:10.1073 / pnas.1305782110. ЧВК 3645540. PMID 23589842.
- ^ Matyskiela ME, Lander GC, Martin A (июль 2013 г.). «Конформационное переключение протеасомы 26S делает возможной деградацию субстрата». Структурная и молекулярная биология природы. 20 (7): 781–788. Дои:10.1038 / nsmb.2616. ЧВК 3712289. PMID 23770819.
- ^ Лу И, Ву Дж, Донг И, Чен С., Сун С., Ма Й.Б., Оуян К., Финли Д., Киршнер М.В., Мао Й. (июль 2017 г.). "Конформационный ландшафт регулирующей частицы протеасомы человека, связанной с p28". Молекулярная клетка. 67 (2): 322–333.e6. Дои:10.1016 / j.molcel.2017.06.007. ЧВК 5580496. PMID 28689658.
- ^ Кёлер А., Кашио П., Леггетт Д.С., Ву К.М., Голдберг А.Л., Финли Д. (июнь 2001 г.). «Осевой канал коровой частицы протеасомы контролируется АТФазой Rpt2 и контролирует как поступление субстрата, так и высвобождение продукта». Молекулярная клетка. 7 (6): 1143–52. Дои:10.1016 / S1097-2765 (01) 00274-X. PMID 11430818.
- ^ Ферстер А., Мастерс Э.И., Уитби Ф.Г., Робинсон Х., Хилл С.П. (май 2005 г.). «1.9A структура комплекса активатора протеасомы-11S и значение для взаимодействий протеасома-PAN / PA700». Молекулярная клетка. 18 (5): 589–99. Дои:10.1016 / j.molcel.2005.04.016. PMID 15916965.
- ^ Витт С., Квон Ю.Д., Шарон М., Фельдерер К., Бейттлер М., Робинсон К.В., Баумейстер В., Япончик Б.К. (июль 2006 г.). «Сборка протеасомы запускает переключатель, необходимый для созревания активного сайта». Структура. 14 (7): 1179–88. Дои:10.1016 / j.str.2006.05.019. PMID 16843899.
- ^ Крюгер Э., Клётцель П.М., Эненкель С. (2001). «Биогенез 20S протеасомы». Биохимия. 83 (3–4): 289–93. Дои:10.1016 / S0300-9084 (01) 01241-X. PMID 11295488.
- ^ Мурата С., Яширода Х., Танака К. (февраль 2009 г.). «Молекулярные механизмы сборки протеасом». Обзоры природы Молекулярная клеточная биология. 10 (2): 104–115. Дои:10.1038 / nrm2630. PMID 19165213. S2CID 21263837.
- ^ Саката Э., Стенгель Ф., Фукунага К., Чжоу М., Саэки Й., Фёрстер Ф., Баумейстер В., Танака К., Робинсон CV (июнь 2011 г.). «Каталитическая активность Ubp6 увеличивает созревание протеасомной регуляторной частицы». Молекулярная клетка. 42 (5): 637–649. Дои:10.1016 / j.molcel.2011.04.021. PMID 21658604.
- ^ Фукунага К., Кудо Т., Тох-э А., Танака К., Саеки Ю. (июнь 2010 г.). «Рассечение пути сборки крышки протеасомы в Saccharomyces cerevisiae». Сообщения о биохимических и биофизических исследованиях. 396 (4): 1048–1053. Дои:10.1016 / j.bbrc.2010.05.061. PMID 20471955.
- ^ Хаас А.Л., Вармс СП, Хершко А., Роуз И.А. (март 1982 г.). «Убиквитин-активирующий фермент. Механизм и роль в конъюгации белок-убиквитин». Журнал биологической химии. 257 (5): 2543–8. PMID 6277905.
- ^ Thrower JS, Hoffman L, Rechsteiner M, Pickart CM (январь 2000 г.). «Распознавание протеолитического сигнала полиубиквитина». Журнал EMBO. 19 (1): 94–102. Дои:10.1093 / emboj / 19.1.94. ЧВК 1171781. PMID 10619848.
- ^ Risseeuw EP, Daskalchuk TE, Banks TW, Liu E, Cotelesage J, Hellmann H, Estelle M, Somers DE, Crosby WL (июнь 2003 г.). "Анализ взаимодействия белков субъединиц SCF убиквитин E3 лигазы из Arabidopsis". Журнал растений. 34 (6): 753–67. Дои:10.1046 / j.1365-313X.2003.01768.x. PMID 12795696.
- ^ Эльзассер С., Финли Д. (август 2005 г.). «Доставка убиквитинированных субстратов в машины для разворачивания белков». Природа клеточной биологии. 7 (8): 742–9. Дои:10.1038 / ncb0805-742. PMID 16056265. S2CID 21069699.
- ^ Саданандом А., Бейли М., Эван Р., Ли Дж., Нелис С. (октябрь 2012 г.). «Убиквитин-протеасомная система: центральный модификатор передачи сигналов растений». Новый Фитолог. 196 (1): 13–28. Дои:10.1111 / j.1469-8137.2012.04266.x. PMID 22897362.
- ^ Sharp PM, Ли WH (1987). «Гены убиквитина как парадигма согласованной эволюции тандемных повторов». Журнал молекулярной эволюции. 25 (1): 58–64. Bibcode:1987JMolE..25 ... 58S. Дои:10.1007 / BF02100041. PMID 3041010. S2CID 7929162.
- ^ Pickart CM, Fushman D (декабрь 2004 г.). «Полиубиквитиновые цепи: сигналы полимерных белков». Современное мнение в области химической биологии. 8 (6): 610–16. Дои:10.1016 / j.cbpa.2004.09.009. PMID 15556404.
- ^ Xu P, Duong DM, Seyfried NT, Cheng D, Xie Y, Robert J, Rush J, Hochstrasser M, Finley D, Peng J (апрель 2009 г.). «Количественная протеомика раскрывает функцию нестандартных цепей убиквитина в протеасомной деградации». Клетка. 137 (1): 133–45. Дои:10.1016 / j.cell.2009.01.041. ЧВК 2668214. PMID 19345192.
- ^ Pickart CM (ноябрь 2000 г.). «Убиквитин в цепочках». Тенденции в биохимических науках. 25 (11): 544–8. Дои:10.1016 / S0968-0004 (00) 01681-9. PMID 11084366.
- ^ Чжу К., Вани Дж., Ван К. Е., Эль-Махди М., Снапка Р. М., Вани А. А. (июль 2005 г.). «Деубиквитинирование протеасомами координируется с транслокацией субстрата для протеолиза in vivo». Экспериментальные исследования клеток. 307 (2): 436–51. Дои:10.1016 / j.yexcr.2005.03.031. PMID 15950624.
- ^ Венцель Т., Баумейстер В. (март 1995 г.). «Конформационные ограничения в деградации белка протеасомой 20S». Структурная биология природы. 2 (3): 199–204. Дои:10.1038 / nsb0395-199. PMID 7773788. S2CID 41599619.
- ^ Инобе Т., Фишбейн С., Пракаш С., Матушек А. (март 2011 г.). «Определение геометрии двухкомпонентного протеасомного дегрона». Природа Химическая Биология. 7 (3): 161–7. Дои:10.1038 / nchembio.521. ЧВК 3129032. PMID 21278740.
- ^ ван дер Ли Р., Ланг Б., Круз К., Гспонер Дж., Санчес де Гроот Н., Хьюнен М.А., Матушек А., Фуксрайтер М., Бабу М.М. (сентябрь 2014 г.). «Внутренне неупорядоченные сегменты влияют на период полужизни белка в клетке и в процессе эволюции». Отчеты по ячейкам. 8 (6): 1832–44. Дои:10.1016 / j.celrep.2014.07.055. ЧВК 4358326. PMID 25220455.
- ^ Смит Д.М., Бенарудж Н., Голдберг А. (октябрь 2006 г.). «Протеасомы и связанные с ними АТФазы: деструктивная комбинация». Журнал структурной биологии. 156 (1): 72–83. Дои:10.1016 / j.jsb.2006.04.012. PMID 16919475.
- ^ Хойт М.А., Зич Дж., Такеучи Дж., Чжан М., Говертс С., Коффино П. (апрель 2006 г.). «Глицин-аланинские повторы нарушают правильное разворачивание субстрата протеасомой». Журнал EMBO. 25 (8): 1720–9. Дои:10.1038 / sj.emboj.7601058. ЧВК 1440830. PMID 16601692.
- ^ Чжан М., Коффино П. (март 2004 г.). «Повторяющаяся последовательность белка ядерного антигена 1, кодируемого вирусом Эпштейна-Барра, прерывает процессинг протеасомного субстрата». Журнал биологической химии. 279 (10): 8635–41. Дои:10.1074 / jbc.M310449200. PMID 14688254.
- ^ Зеемюллер Э., Лупас А., Сток Д., Лёве Дж., Хубер Р., Баумейстер В. (апрель 1995 г.). «Протеасома из Thermoplasma acidophilum: треониновая протеаза». Наука. 268 (5210): 579–82. Bibcode:1995Научный ... 268..579S. Дои:10.1126 / science.7725107. PMID 7725107.
- ^ Coux O, Tanaka K, Goldberg AL (1996). «Структура и функции протеасом 20S и 26S». Ежегодный обзор биохимии. 65: 801–47. Дои:10.1146 / annurev.bi.65.070196.004101. PMID 8811196.
- ^ Groll M, Ditzel L, Löwe J, Stock D, Bochtler M, Bartunik HD, Huber R (апрель 1997 г.). «Структура протеасомы 20S из дрожжей при разрешении 2,4 А». Природа. 386 (6624): 463–71. Дои:10.1038 / 386463a0. PMID 9087403. S2CID 4261663.
- ^ Дик Т.П., Нуссбаум А.К., Диг М., Хайнемейер В., Гролл М., Ширле М., Кейлхольц В., Стеванович С., Вольф Д.Х., Хубер Р., Раммензее Х.Г., Шильд Х (октябрь 1998 г.). «Вклад протеасомных бета-субъединиц в расщепление пептидных субстратов, анализируемых с помощью мутантов дрожжей». Журнал биологической химии. 273 (40): 25637–46. Дои:10.1074 / jbc.273.40.25637. PMID 9748229.
- ^ Voges D, Zwickl P, Baumeister W (1999). «Протеасома 26S: молекулярная машина, разработанная для контролируемого протеолиза». Ежегодный обзор биохимии. 68 (1): 1015–68. Дои:10.1146 / annurev.biochem.68.1.1015. PMID 10872471.
- ^ а б Rape M, Jentsch S (май 2002 г.). «Откусить: процессинг протеасомного белка». Природа клеточной биологии. 4 (5): E113–6. Дои:10.1038 / ncb0502-e113. PMID 11988749. S2CID 7126477.
- ^ Rape M, Jentsch S (ноябрь 2004 г.). «Продуктивная RUPture: активация факторов транскрипции протеасомным процессингом». Biochimica et Biophysica Acta (BBA) - Исследование молекулярных клеток. 1695 (1–3): 209–13. Дои:10.1016 / j.bbamcr.2004.09.022. PMID 15571816.
- ^ Ашер Г., Реувен Н., Шауль И. (август 2006 г.). "20S протеасомы и деградация белков" по умолчанию"". BioEssays. 28 (8): 844–9. Дои:10.1002 / bies.20447. PMID 16927316.
- ^ Чжан М., Пикарт С.М., Коффино П. (апрель 2003 г.). «Детерминанты распознавания протеасомами орнитиндекарбоксилазы, убиквитиннезависимого субстрата». Журнал EMBO. 22 (7): 1488–96. Дои:10.1093 / emboj / cdg158. ЧВК 152902. PMID 12660156.
- ^ Ашер Г., Шауль И. (август 2005 г.). «Протеасомная деградация p53: полиубиквитинирование - это еще не все». Клеточный цикл. 4 (8): 1015–8. Дои:10.4161 / cc.4.8.1900. PMID 16082197.
- ^ а б Shringarpure R, Grune T, Mehlhase J, Davies KJ (январь 2003 г.). «Конъюгирование убиквитина не требуется для расщепления окисленных белков протеасомами». Журнал биологической химии. 278 (1): 311–8. Дои:10.1074 / jbc.M206279200. PMID 12401807.
- ^ а б c Gille C, Goede A, Schlöetelburg C, Preissner R, Kloetzel PM, Göbel UB, Frömmel C (март 2003 г.). «Комплексный взгляд на протеасомные последовательности: последствия для эволюции протеасомы». Журнал молекулярной биологии. 326 (5): 1437–48. Дои:10.1016 / S0022-2836 (02) 01470-5. PMID 12595256.
- ^ Бохтлер М., Дицель Л., Гролл М., Хартманн С., Хубер Р. (1999). «Протеасома». Ежегодный обзор биофизики и структуры биомолекул. 28 (1): 295–317. Дои:10.1146 / annurev.biophys.28.1.295. PMID 10410804.
- ^ Chesnel F, Bazile F, Pascal A, Kubiak JZ (август 2006 г.). «Диссоциация циклина B от CDK1 предшествует его деградации при инактивации MPF в митотических экстрактах эмбрионов Xenopus laevis». Клеточный цикл. 5 (15): 1687–98. Дои:10.4161 / cc.5.15.3123. PMID 16921258.
- ^ Брито Д.А., Ридер С.Л. (июнь 2006 г.). «Проскальзывание митотической контрольной точки у людей происходит за счет разрушения циклина B в присутствии активной контрольной точки». Текущая биология. 16 (12): 1194–200. Дои:10.1016 / j.cub.2006.04.043. ЧВК 2749311. PMID 16782009.
- ^ Хэвенс К.Г., Хо А., Йошиока Н., Дауди С.Ф. (июнь 2006 г.). «Регулирование позднего фазового перехода G1 / S и APC Cdh1 с помощью активных форм кислорода». Молекулярная и клеточная биология. 26 (12): 4701–11. Дои:10.1128 / MCB.00303-06. ЧВК 1489138. PMID 16738333.
- ^ Башир Т., Доррелло Н.В., Амадор В., Гуардаваккаро Д., Пагано М. (март 2004 г.). «Контроль убиквитинлигазы SCF (Skp2-Cks1) с помощью убиквитинлигазы APC / C (Cdh1)». Природа. 428 (6979): 190–3. Дои:10.1038 / природа02330. PMID 15014502. S2CID 4401971.
- ^ Хигасицудзи Х., Лю Ю., Майер Р.Дж., Фудзита Дж. (Октябрь 2005 г.). «Онкопротеин ганкирин негативно регулирует как p53, так и RB, усиливая протеасомную деградацию». Клеточный цикл. 4 (10): 1335–7. Дои:10.4161 / cc.4.10.2107. PMID 16177571.
- ^ Дхармасири С., Эстель М. (2002). «Роль регулируемой деградации белка в ответе на ауксин». Молекулярная биология растений. 49 (3–4): 401–9. Дои:10.1023 / А: 1015203013208. PMID 12036263. S2CID 7669386.
- ^ Weijers D, Benkova E, Jäger KE, Schlereth A, Hamann T, Kientz M, Wilmoth JC, Reed JW, Jürgens G (май 2005 г.). «Специфичность развития ауксинового ответа парами регуляторов транскрипции ARF и Aux / IAA». Журнал EMBO. 24 (10): 1874–85. Дои:10.1038 / sj.emboj.7600659. ЧВК 1142592. PMID 15889151.
- ^ Хаас А.Л., Бабошина О., Уильямс Б., Шварц Л.М. (апрель 1995 г.). «Скоординированная индукция пути конъюгации убиквитина сопровождает запрограммированную в процессе развития смерть скелетных мышц насекомых». Журнал биологической химии. 270 (16): 9407–12. Дои:10.1074 / jbc.270.16.9407. PMID 7721865.
- ^ Шварц Л. М., Майер А., Кош Л., Энгельштейн М., Майер С. (октябрь 1990 г.). «Активация экспрессии гена полиубиквитина во время запрограммированной гибели клеток». Нейрон. 5 (4): 411–9. Дои:10.1016 / 0896-6273 (90) 90080-У. PMID 2169771. S2CID 33829749.
- ^ Лёв П., Басселл К., Доусон С.П., Биллетт М.А., Майер Р.Дж., Рейнольдс С.Е. (январь 1997 г.). «Экспрессия субъединицы АТФазы 26S протеасомы, MS73, в мышцах, которые подвергаются программируемой гибели клеток, и ее контроль с помощью экдистероидных гормонов у насекомого Manduca sexta». Письма FEBS. 400 (3): 345–9. Дои:10.1016 / S0014-5793 (96) 01413-5. PMID 9009228. S2CID 10873052.
- ^ Питцер Ф, Дантес А., Фукс Т, Баумейстер В., Амстердам А (сентябрь 1996 г.). «Удаление протеасом из ядра и их накопление в апоптотических пузырьках во время запрограммированной гибели клеток». Письма FEBS. 394 (1): 47–50. Дои:10.1016/0014-5793(96)00920-9. PMID 8925925. S2CID 29256092.
- ^ а б Адамс Дж., Паломбелла В.Дж., Сосвилл Е.А., Джонсон Дж., Дестри А., Лазарус Д.Д., Маас Дж., Пьен С.С., Пракаш С., Эллиотт П.Дж. (июнь 1999 г.). «Ингибиторы протеасом: новый класс сильнодействующих и эффективных противоопухолевых средств». Исследования рака. 59 (11): 2615–22. PMID 10363983.
- ^ Орловский Р.З. (апрель 1999 г.). «Роль убиквитин-протеасомного пути в апоптозе». Гибель клеток и дифференциация. 6 (4): 303–13. Дои:10.1038 / sj.cdd.4400505. PMID 10381632.
- ^ Гарридо С., Брюнет М., Диделот С., Зермати Ю., Шмитт Е., Кремер Г. (ноябрь 2006 г.). «Белки теплового шока 27 и 70: антиапоптотические белки с канцерогенными свойствами». Клеточный цикл. 5 (22): 2592–601. Дои:10.4161 / cc.5.22.3448. PMID 17106261.
- ^ Park SH, Bolender N, Eisele F, Kostova Z, Takeuchi J, Coffino P, Wolf DH (январь 2007 г.). «Цитоплазматический механизм шаперона Hsp70 подвергает неправильно свернутые и некомпетентные по импорту эндоплазматического ретикулума белки к деградации через систему убиквитин-протеасома». Молекулярная биология клетки. 18 (1): 153–65. Дои:10.1091 / mbc.E06-04-0338. ЧВК 1751312. PMID 17065559.
- ^ Дай Кью, Цянь С.Б., Ли Х.Х., МакДонаф Х., Борчерс С., Хуанг Д., Такаяма С., Янгер Дж.М., Рен Х.Й., Сир Д.М., Паттерсон С. (ноябрь 2005 г.). «Регулирование пути деградации белка контроля качества цитоплазмы с помощью BAG2». Журнал биологической химии. 280 (46): 38673–81. Дои:10.1074 / jbc.M507986200. PMID 16169850.
- ^ Бадер Н., Грюн Т. (2006). «Окисление и протеолиз белков». Биологическая химия. 387 (10–11): 1351–5. Дои:10.1515 / BC.2006.169. PMID 17081106. S2CID 30385354.
- ^ Дэвис К.Дж. (2003). «Расщепление окисленных белков протеасомой 20S». Биохимия. 83 (3–4): 301–10. Дои:10.1016 / S0300-9084 (01) 01250-0. PMID 11295490.
- ^ а б Lehman NL (сентябрь 2009 г.). «Убиквитиновая протеасомная система в невропатологии». Acta Neuropathologica. 118 (3): 329–47. Дои:10.1007 / s00401-009-0560-х. ЧВК 2716447. PMID 19597829.
- ^ Макнот К.С., Джексон Т., ДжноБаптист Р., Капустин А., Оланов К.В. (май 2006 г.). «Протеасомная дисфункция при спорадической болезни Паркинсона». Неврология. 66 (10 Дополнение 4): S37–49. Дои:10.1212 / 01.wnl.0000221745.58886.2e. PMID 16717251.
- ^ Шарма Н., Брэндис К.А., Эррера С.К., Джонсон Б.Е., Вайдья Т., Шреста Р., Деббурман С.К. (2006). «Модель почкующихся альфа-синуклеиновых дрожжей: токсичность усиливается нарушением протеасомы и окислительным стрессом». Журнал молекулярной неврологии. 28 (2): 161–78. Дои:10.1385 / JMN: 28: 2: 161. PMID 16679556.
- ^ Мурата С., Сасаки К., Кишимото Т., Нива С., Хаяси Х., Такахама Ю., Танака К. (июнь 2007 г.). «Регулирование развития CD8 + Т-клеток с помощью протеасом, специфичных для тимуса». Наука. 316 (5829): 1349–53. Bibcode:2007Научный ... 316.1349M. Дои:10.1126 / science.1141915. PMID 17540904. S2CID 37185716.
- ^ Касио П., Хилтон С., Киселев А.Ф., Рок К.Л., Голдберг А.Л. (май 2001 г.). «26S протеасомы и иммунопротеасомы продуцируют в основном N-удлиненные версии антигенного пептида». Журнал EMBO. 20 (10): 2357–66. Дои:10.1093 / emboj / 20.10.2357. ЧВК 125470. PMID 11350924.
- ^ Маллери Д.Л., Макьюэн В.А., Бидгуд С.Р., Тауэрс Г.Дж., Джонсон С.М., Джеймс Л.К. (ноябрь 2010 г.). «Антитела опосредуют внутриклеточный иммунитет через тройной мотив 21 (TRIM21)». Труды Национальной академии наук Соединенных Штатов Америки. 107 (46): 19985–19990. Bibcode:2010ПНАС..10719985М. Дои:10.1073 / pnas.1014074107. ЧВК 2993423. PMID 21045130.
- ^ Fenteany G, Standaert RF, Lane WS, Choi S, Corey EJ, Schreiber SL (май 1995 г.). «Ингибирование активности протеасом и субъединично-специфической модификации аминоконцевого треонина с помощью лактацистина». Наука. 268 (5211): 726–31. Bibcode:1995Научный ... 268..726F. Дои:10.1126 / science.7732382. PMID 7732382. S2CID 37779687.
- ^ Пресс-релиз Управления по контролю за продуктами и лекарствами США В архиве 19 февраля 2007 г. Wayback Machine 13 мая 2003 г. Дата доступа 29 декабря 2006 г. См. Также Информационная страница FDA Velcade.
- ^ Фишер Р.И., Бернштейн С.Х., Каль Б.С., Джулбегович Б., Робертсон М.Дж., де Вос С., Эпнер Э., Кришнан А., Леонард Дж. П., Лониал С., Штадтмауэр Е.А., О'Коннор О.А., Ши Х., Борал А.Л., Гой А. (октябрь 2006 г. ). «Многоцентровое исследование фазы II бортезомиба у пациентов с рецидивирующей или рефрактерной лимфомой из мантийных клеток». Журнал клинической онкологии. 24 (30): 4867–74. Дои:10.1200 / JCO.2006.07.9665. PMID 17001068.
- ^ Якоб С., Эгерер К., Либиш П., Тюркмен С., Заврски И., Кукелькорн Ю., Хейдер Ю., Кайзер М., Флейсснер С., Штерц Дж., Клиберг Л., Файст Е., Бурместер Г. Р., Клётцель П. М., Сезер О. (март 2007 г.). «Уровни циркулирующих протеасом являются независимым прогностическим фактором выживания при множественной миеломе». Кровь. 109 (5): 2100–5. Дои:10.1182 / кровь-2006-04-016360. PMID 17095627.
- ^ Шах С.А., Поттер М.В., Макдейд Т.П., Риккарди Р., Перуджини Р.А., Эллиотт П.Дж., Адамс Дж., Каллери М.П. (2001). «Ингибирование протеасомы 26S вызывает апоптоз и ограничивает рост рака поджелудочной железы человека». Журнал клеточной биохимии. 82 (1): 110–22. Дои:10.1002 / jcb.1150. PMID 11400168. S2CID 21223980.
- ^ Nawrocki ST, Суини-Готч B, Takamori R, McConkey DJ (январь 2004 г.). «Ингибитор протеасом бортезомиб усиливает активность доцетаксела в ксенотрансплантатах ортотопической опухоли поджелудочной железы человека». Молекулярная терапия рака. 3 (1): 59–70. PMID 14749476.
- ^ Schenkein D (июнь 2002 г.). «Ингибиторы протеасом в лечении злокачественных новообразований В-клеток». Клиническая лимфома. 3 (1): 49–55. Дои:10.3816 / CLM.2002.n.011. PMID 12141956.
- ^ О'Коннор О.А., Райт Дж., Московиц К., Маззи Дж., МакГрегор-Кортелли Б., Стабблфилд М., Штраус Д., Портлок С., Хэмлин П., Чой Э, Думетреску О, Эсселтин Д., Треху Е., Адамс Дж., Шенкейн Д., Зеленец AD (февраль 2005 г.). «Фаза II клинического опыта с новым ингибитором протеасом бортезомибом у пациентов с вялотекущей неходжкинской лимфомой и лимфомой из клеток мантии». Журнал клинической онкологии. 23 (4): 676–84. Дои:10.1200 / JCO.2005.02.050. PMID 15613699.
- ^ Мессинджер Й.Х., Гейнон П.С., Спосто Р., Ван дер Гиссен Дж., Экрот Э., Малвар Дж., Бостром Британская Колумбия (июль 2012 г.). «Бортезомиб с химиотерапией очень активен при запущенном остром лимфобластном лейкозе, являющемся предшественником группы B: исследование терапевтических достижений в области детской лейкемии и лимфомы (TACL)». Кровь. 120 (2): 285–90. Дои:10.1182 / кровь-2012-04-418640. PMID 22653976.
- ^ Ламбру Г.И., Пападимитриу Л., Хрусос Г.П., Влахопулос С.А. (апрель 2012 г.). «Влияние глюкокортикоидов и ингибиторов протеасом на лейкемический лимфобласт: множественные, разнообразные сигналы, сходящиеся на нескольких ключевых нижестоящих регуляторах». Молекулярная и клеточная эндокринология. 351 (2): 142–51. Дои:10.1016 / j.mce.2012.01.003. PMID 22273806. S2CID 28749125.
- ^ Schmidtke G, Holzhütter HG, Bogyo M, Kairies N, Groll M, de Giuli R, Emch S, Groettrup M (декабрь 1999 г.). «Как ингибитор протеазы ВИЧ-1 модулирует активность протеасом». Журнал биологической химии. 274 (50): 35734–40. Дои:10.1074 / jbc.274.50.35734. PMID 10585454.
- ^ Laurent N, de Boüard S, Guillamo JS, Christov C, Zini R, Jouault H, Andre P, Lotteau V, Peschanski M (февраль 2004 г.). «Влияние ингибитора протеасом ритонавира на рост глиомы in vitro и in vivo». Молекулярная терапия рака. 3 (2): 129–36. PMID 14985453.
- ^ Zollner TM, Podda M, Pien C, Elliott PJ, Kaufmann R, Boehncke WH (март 2002 г.). «Ингибирование протеасом снижает опосредованную суперантигеном активацию Т-клеток и тяжесть псориаза в модели SCID-hu». Журнал клинических исследований. 109 (5): 671–9. Дои:10.1172 / JCI12736. ЧВК 150886. PMID 11877475.
- ^ Эллиотт П.Дж., Пьен С.С., Маккормак Т.А., Чепмен И.Д., Адамс Дж. (Август 1999 г.). «Ингибирование протеасом: новый механизм борьбы с астмой». Журнал аллергии и клинической иммунологии. 104 (2, ч. 1): 294–300. Дои:10.1016 / S0091-6749 (99) 70369-6. PMID 10452747.
- ^ Verdoes M, Florea BI, Menendez-Benito V, Maynard CJ, Witte MD, van der Linden WA, van den Nieuwendijk AM, Hofmann T., Berkers CR, van Leeuwen FW, Groothuis TA, Leeuwenburgh MA, Ovaa H, Neefjes JJ, Filippov Д.В., ван дер Марель Г.А., Дантума Н.П., Overkleeft HS (ноябрь 2006 г.). «Флуоресцентный ингибитор протеасом широкого спектра действия для мечения протеасом in vitro и in vivo». Химия и биология. 13 (11): 1217–26. Дои:10.1016 / j.chembiol.2006.09.013. PMID 17114003.
- ^ Клейгер Г., мэр Т. (июнь 2014 г.). «Опасное путешествие: экскурсия по убиквитин-протеасомной системе». Тенденции в клеточной биологии. 24 (6): 352–9. Дои:10.1016 / j.tcb.2013.12.003. ЧВК 4037451. PMID 24457024.
- ^ Гольдберг А. Л., Стейн Р., Адамс Дж. (Август 1995 г.). «Новое понимание функции протеасом: от архебактерий до разработки лекарств». Химия и биология. 2 (8): 503–8. Дои:10.1016/1074-5521(95)90182-5. PMID 9383453.
- ^ Сулистио Ю.А., Хиз К. (январь 2015 г.). «Убиквитин-протеасомная система и дерегуляция молекулярных шаперонов при болезни Альцгеймера». Молекулярная нейробиология. 53 (2): 905–31. Дои:10.1007 / s12035-014-9063-4. PMID 25561438. S2CID 14103185.
- ^ Ортега З, Лукас Дж.Дж. (2014). «Участие убиквитин-протеасомной системы в болезни Хантингтона». Границы молекулярной неврологии. 7: 77. Дои:10.3389 / fnmol.2014.00077. ЧВК 4179678. PMID 25324717.
- ^ Сандри М., Роббинс Дж. (Июнь 2014 г.). «Протеотоксичность: недооцененная патология при сердечных заболеваниях». Журнал молекулярной и клеточной кардиологии. 71: 3–10. Дои:10.1016 / j.yjmcc.2013.12.015. ЧВК 4011959. PMID 24380730.
- ^ Дрюс О., Тэгтмайер Х (декабрь 2014 г.). «Нацеливание на убиквитин-протеасомную систему при сердечных заболеваниях: основа новых терапевтических стратегий». Антиоксиданты и редокс-сигналы. 21 (17): 2322–43. Дои:10.1089 / ars.2013.5823. ЧВК 4241867. PMID 25133688.
- ^ Ван З.В., Хилл Дж. А. (февраль 2015 г.). «Контроль качества протеина и метаболизм: двунаправленный контроль в сердце». Клеточный метаболизм. 21 (2): 215–26. Дои:10.1016 / j.cmet.2015.01.016. ЧВК 4317573. PMID 25651176.
- ^ а б Карин М., Дельхас М. (февраль 2000 г.). «Киназа I каппа B (IKK) и NF-каппа B: ключевые элементы провоспалительной передачи сигналов». Семинары по иммунологии. 12 (1): 85–98. Дои:10.1006 / smim.2000.0210. PMID 10723801.
- ^ Ермолаева М.А., Даховник А, Шумахер Б. (январь 2015). «Механизмы контроля качества в ответах на клеточные и системные повреждения ДНК». Обзоры исследований старения. 23 (Pt A): 3–11. Дои:10.1016 / j.arr.2014.12.009. ЧВК 4886828. PMID 25560147.
- ^ Checler F, da Costa CA, Ancolio K, Chevallier N, Lopez-Perez E., Marambaud P (июль 2000 г.). «Роль протеасомы в болезни Альцгеймера». Biochimica et Biophysica Acta (BBA) - Молекулярная основа болезни. 1502 (1): 133–8. Дои:10.1016 / s0925-4439 (00) 00039-9. PMID 10899438.
- ^ а б Чунг К.К., Доусон В.Л., Доусон TM (ноябрь 2001 г.). «Роль убиквитин-протеасомного пути в болезни Паркинсона и других нейродегенеративных расстройствах». Тенденции в неврологии. 24 (11 Прил.): S7–14. Дои:10.1016 / s0166-2236 (00) 01998-6. PMID 11881748. S2CID 2211658.
- ^ а б Икеда К., Акияма Х., Араи Т., Уэно Х., Цучия К., Косака К. (июль 2002 г.). «Морфометрическая переоценка системы двигательных нейронов болезни Пика и бокового амиотрофического склероза с деменцией». Acta Neuropathologica. 104 (1): 21–8. Дои:10.1007 / s00401-001-0513-5. PMID 12070660. S2CID 22396490.
- ^ Манака Х, Като Т, Курита К., Катагири Т, Шикама Й, Кудзираи К., Каванами Т, Судзуки И, Нихей К., Сасаки Х (май 1992 г.). «Заметное увеличение убиквитина в спинномозговой жидкости при болезни Крейтцфельдта – Якоба». Письма о неврологии. 139 (1): 47–9. Дои:10.1016 / 0304-3940 (92) 90854-з. PMID 1328965. S2CID 28190967.
- ^ Мэтьюз К.Д., Мур С.А. (январь 2003 г.). «Конечностно-поясная мышечная дистрофия». Текущие отчеты по неврологии и неврологии. 3 (1): 78–85. Дои:10.1007 / s11910-003-0042-9. PMID 12507416. S2CID 5780576.
- ^ Майер Р.Дж. (март 2003 г.). «От нейродегенерации к нейрогомеостазу: роль убиквитина». Новости и перспективы наркотиков. 16 (2): 103–8. Дои:10.1358 / dnp.2003.16.2.829327. PMID 12792671.
- ^ Кализа Дж., Пауэлл С.Р. (февраль 2013 г.). «Убиквитиновая протеасомная система и ишемия миокарда». Американский журнал физиологии. Сердце и физиология кровообращения. 304 (3): H337–49. Дои:10.1152 / ajpheart.00604.2012. ЧВК 3774499. PMID 23220331.
- ^ Предмор Дж. М., Ван П., Дэвис Ф., Бартолон С., Вестфол М. В., Дайк Д. Б., Пагани Ф., Пауэлл С. Р., Дэй С.М. (март 2010 г.). «Дисфункция убиквитиновых протеасом при гипертрофических и дилатационных кардиомиопатиях». Тираж. 121 (8): 997–1004. Дои:10.1161 / CIRCULATIONAHA.109.904557. ЧВК 2857348. PMID 20159828.
- ^ Пауэлл С.Р. (июль 2006 г.). «Убиквитин-протеасомная система в физиологии и патологии сердца». Американский журнал физиологии. Сердце и физиология кровообращения. 291 (1): H1 – H19. Дои:10.1152 / ajpheart.00062.2006. PMID 16501026.
- ^ Адамс Дж (апрель 2003 г.). «Возможности ингибирования протеасомы при лечении рака». Открытие наркотиков сегодня. 8 (7): 307–15. Дои:10.1016 / с 1359-6446 (03) 02647-3. PMID 12654543.
- ^ Бен-Нерия Y (январь 2002 г.). «Регуляторные функции убиквитинирования в иммунной системе». Иммунология природы. 3 (1): 20–6. Дои:10.1038 / ni0102-20. PMID 11753406. S2CID 26973319.
- ^ Egerer K, Kuckelkorn U, Rudolph PE, Rückert JC, Dörner T., Burmester GR, Kloetzel PM, Feist E (октябрь 2002 г.). «Циркулирующие протеасомы являются маркерами повреждения клеток и иммунологической активности при аутоиммунных заболеваниях». Журнал ревматологии. 29 (10): 2045–52. PMID 12375310.
дальнейшее чтение
- Гликман М. Х., Адир Н. (январь 2004 г.). «Протеасома и тонкий баланс между разрушением и спасением». PLOS Биология. 2 (1): e13. Дои:10.1371 / journal.pbio.0020013. ЧВК 314468. PMID 14737189.
- Протеасома дрожжей 26S со списком субъединиц и изображениями
- Цехановер А (сентябрь 2005 г.). «Ранняя работа над системой убиквитиновых протеасом, интервью с Аароном Чехановером. Интервью CDD». Гибель клеток и дифференциация. 12 (9): 1167–77. Дои:10.1038 / sj.cdd.4401691. PMID 16094393.
- Гершко А. (сентябрь 2005 г.). «Ранняя работа над системой убиквитиновых протеасом, интервью с Аврамом Хершко. Интервью CDD». Гибель клеток и дифференциация. 12 (9): 1158–61. Дои:10.1038 / sj.cdd.4401709. PMID 16094391.
- Роза I (сентябрь 2005 г.). «Ранняя работа над убиквитиновой протеасомной системой, интервью с Ирвином Роузом. Интервью CDD». Гибель клеток и дифференциация. 12 (9): 1162–6. Дои:10.1038 / sj.cdd.4401700. PMID 16094392.
- Цвек Б., Дворак З. (2007). «Нацеленность на ядерный фактор-каппаВ и протеасомы с помощью дитиокарбаматных комплексов с металлами». Текущий фармацевтический дизайн. 13 (30): 3155–67. Дои:10.2174/138161207782110390. PMID 17979756. Архивировано из оригинал 29 июля 2012 г.