Солевой мостик (белковый и супрамолекулярный) - Salt bridge (protein and supramolecular) - Wikipedia

В химия, а соляной мост это комбинация двух нековалентные взаимодействия: водородная связь и ионная связь (Рисунок 1). Ионное спаривание - одно из самых важных нековалентный силы в химии, в биологических системах, в различных материалах и во многих приложениях, таких как ионно-парная хроматография. Это наиболее часто наблюдаемый вклад в стабильность энтропийно неблагоприятной свернутой конформации белков. Хотя нековалентные взаимодействия, как известно, являются относительно слабыми взаимодействиями, небольшие стабилизирующие взаимодействия могут вносить важный вклад в общую стабильность конформера.[1] Солевые мостики встречаются не только в белках, но и в супрамолекулярная химия. Термодинамика каждого из них исследуется с помощью экспериментальных процедур для определения вклада свободной энергии солевого мостика в общую свободную энергию состояния.
Солевые мостики в химической связи
В воде образование солевых мостиков или ионные пары в основном определяется энтропией, обычно сопровождаемой неблагоприятным вкладом ΔH из-за десольватации взаимодействующих ионов при ассоциации.[2] Водородные связи способствуют стабильности ионные пары например, с протонированный ионы аммония, а с анионами образуется депротонированием, как и в случае карбоксилат, фосфат так далее; тогда константы ассоциации зависят от pH. Энтропийные движущие силы для образования пар ионов (при отсутствии значительных вкладов водородных связей) также обнаруживаются в метаноле в качестве растворителя. В неполярных растворителях контактные ионные пары с очень высокими константами ассоциации,[3];[4] в газовой фазе энергии ассоциации, например галогениды щелочных металлов достигают до 200 кДж / моль.[5] В Bjerrum или уравнение Фуосса описывает ассоциацию ионных пар как функцию зарядов ионов zA и zB и диэлектрической проницаемости среды ε; соответствующий график зависимости ΔG от zAzB показывает для более чем 200 пар ионов ожидаемую линейную корреляцию для большого разнообразия ионов.[6]Неорганические и органические ионы демонстрируют при умеренной ионной силе I сходные значения ΔG ассоциации солевого мостика от 5 до 6 кДж / моль для 1: 1 комбинации аниона и катиона, почти независимо от природы (размера, поляризуемости и т. .[7][8] Значения ΔG являются аддитивными и приблизительно линейно зависят от зарядов, взаимодействие, например, двухзарядный фосфат-анион с однозарядным катионом аммония составляет примерно 2x5 = 10 кДж / моль. Значения ΔG зависят от ионной силы I раствора, как описано Уравнение Дебая-Хюккеля, при нулевой ионной силе ΔG = 8 кДж / моль. Стабильность пар щелочных ионов как функция заряда аниона z может быть описана более подробным уравнением.[9]
Солевые мостики в белках
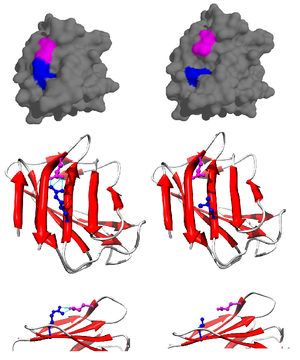
Солевой мостик чаще всего возникает из анионного карбоксилата (RCOO−) либо аспарагиновая кислота или же глютаминовая кислота и катионный аммоний (RNH3+) из лизин или гуанидиний (RNHC (NH2)2+) из аргинин (Фигура 2).[1] Хотя это наиболее распространенные, другие остатки с ионизируемыми боковыми цепями, такие как гистидин, тирозин, и серин также могут участвовать в зависимости от внешних факторов, влияющих на их pKас. Расстояние между остатками, участвующими в солевом мостике, также считается важным. Требуемое расстояние менее 4 Å (400 мкм). Аминокислоты, расстояние между которыми превышает это расстояние, не считаются образующими солевой мостик.[11] Из-за многочисленных ионизируемых боковых цепей аминокислот, присутствующих в белке, pH, при котором находится белок, имеет решающее значение для его стабильности.
Методы количественной оценки стабильности солевого мостика в белках

Вклад солевого мостика в общую стабильность в свернутом состоянии белка можно оценить с помощью термодинамических данных, собранных из исследований мутагенеза и методов ядерного магнитного резонанса.[12] Используя мутировавший белок псевдодикого типа, специфически мутировавший для предотвращения преципитации при высоком pH, вклад солевого мостика в общую свободную энергию свернутого состояния белка можно определить путем выполнения точечной мутации, изменения и, следовательно, разрушения соли мост. Например, было обнаружено, что в лизоциме Т4 существует солевой мостик между аспарагиновой кислотой (Asp) на остатке 70 и гистидином (His) на остатке 31 (Фиг.3). Сайт-направленный мутагенез с аспарагином (Asn) (рис. 4) было получено три новых мутанта: Asp70Asn His31 (мутант 1), Asp70 His31Asn (мутант 2) и Asp70Asn His31Asn (двойной мутант).

После того, как мутанты установлены, можно использовать два метода для расчета свободной энергии, связанной с солевым мостиком. Один метод включает наблюдение температуры плавления белка дикого типа по сравнению с температурой плавления трех мутантов. За денатурацией можно следить по изменению круговой дихроизм. Снижение температуры плавления указывает на снижение стабильности. Это количественно определяется с помощью метода, описанного Бектелем и Шеллманом, где разница в свободной энергии между ними рассчитывается через ΔТΔS.[13] С этим расчетом есть некоторые проблемы, и его можно использовать только с очень точными данными.[нужна цитата ] В примере лизоцима Т4 ΔS псевдо-дикого типа ранее сообщалось при pH 5,5, поэтому средняя разница температур 11 ° C при этом pH, умноженная на сообщенное значение ΔS 360 кал / (моль · К) (1,5 кДж / (моль · К)) дает изменение свободной энергии примерно -4 ккал / моль (-17 кДж / моль). Это значение соответствует количеству свободной энергии, вносимой солевым мостиком в стабильность белка.
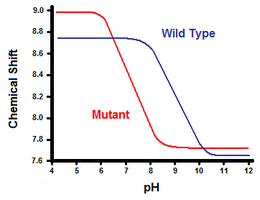
Второй метод использует спектроскопия ядерного магнитного резонанса для расчета свободной энергии солевого мостика. Выполняют титрование, регистрируя химический сдвиг, соответствующий протонам углерода, примыкающего к карбоксилатной или аммониевой группе. Середина кривой титрования соответствует pKа, или pH, при котором соотношение протонированных: депротонированных молекул составляет 1: 1. Продолжая пример лизоцима Т4, кривую титрования получают путем наблюдения за сдвигом протона С2 гистидина 31 (Фиг.5). На рис. 5 показан сдвиг кривой титрования между диким типом и мутантом, в котором Asp70 представляет собой Asn. Образовавшийся солевой мостик находится между депротонированным Asp70 и протонированным His31. Это взаимодействие вызывает сдвиг, видимый в pKа. В развернутом белке дикого типа, где отсутствует солевой мостик, His31, как сообщается, имеет pKа 6,8 дюймов H20 буферов средней ионной силы. На рисунке 5 показан pKа дикого типа 9.05. Эта разница в pKа поддерживается взаимодействием His31 с Asp70. Чтобы поддерживать солевой мостик, His31 будет пытаться удерживать протон как можно дольше. Когда солевой мостик разрушен, как у мутанта D70N, pKа возвращается к значению 6,9, что намного ближе к значению His31 в развернутом состоянии.
Разница в pKа может быть определен количественно, чтобы отразить вклад солевого мостика в свободную энергию. С помощью Свободная энергия Гиббса: Δграмм = −RT ln (Kэкв), куда р - универсальная газовая постоянная, Т это температура в кельвинах, а Kэкв - константа равновесия реакции в равновесии. Депротонирование His31 представляет собой кислотную равновесную реакцию со специальным Kэкв известный как константа диссоциации кислоты, Kа: His31-H+ ⇌ His31 + H+. РKа тогда связано с Kа следующим образом: pKа = −log (Kа). Расчет разности свободной энергии мутанта и дикого типа теперь может быть выполнен с использованием уравнения свободной энергии, определение pKа, наблюдаемая pKа значения и отношения между натуральными логарифмами и логарифмами. В примере с лизоцимом Т4 этот подход дал расчетный вклад примерно 3 ккал / моль в общую свободную энергию.[12] Аналогичный подход может быть использован с другим участником солевого мостика, таким как Asp70 в примере лизоцима Т4, путем мониторинга его сдвига в pKа после мутации His31.
Предупреждение при выборе подходящего эксперимента связано с расположением солевого мостика внутри белка. Окружающая среда играет большую роль во взаимодействии.[14] При высокой ионной силе солевой мостик может быть полностью замаскирован, поскольку участвует электростатическое взаимодействие. Солевой мостик His31-Asp70 в лизоциме Т4 был похоронен внутри белка. Энтропия играет большую роль в поверхностных солевых мостиках, где остатки, которые обычно обладают способностью двигаться, ограничиваются их электростатическим взаимодействием и водородными связями. Было показано, что это уменьшает энтропию настолько, чтобы почти стереть вклад взаимодействия.[15] Поверхностные солевые мостики можно изучать так же, как и заглубленные солевые мостики, используя циклы двойных мутантов и ЯМР-титрование.[16] Хотя существуют случаи, когда заглубленные соляные мосты, как и все остальное, способствуют стабильности, существуют исключения, и заглубленные соляные мосты могут оказывать дестабилизирующий эффект.[11] Кроме того, поверхностные солевые мостики при определенных условиях могут проявлять стабилизирующий эффект.[14][16] Стабилизирующий или дестабилизирующий эффект следует оценивать в индивидуальном порядке, и можно сделать несколько общих заявлений.
Супрамолекулярная химия


Супрамолекулярная химия это область, связанная с нековалентными взаимодействиями между макромолекулами. Соляные мостики использовались химиками в этой области как разнообразными, так и творческими способами, включая определение анионов, синтез молекулярных капсул и двойных спиральных полимеров.
Комплексообразование анионов
Основной вклад супрамолекулярной химии был посвящен распознаванию и зондированию анионов.[17][18][19][20][21][22] Ионное спаривание является наиболее важной движущей силой образования комплекса анионов, но селективность, например, в пределах галогенидного ряда достигается, в основном, за счет вкладов водородных связей.
Молекулярные капсулы
Молекулярные капсулы - это химические каркасы, предназначенные для захвата и удержания гостевой молекулы (см. молекулярная инкапсуляция ). Шумна и его коллеги разработали новую молекулярную капсулу с хиральный интерьер.[23] Капсула состоит из двух половинок, как пластик. пасхальное яйцо (Рисунок 6). Взаимодействие солевого мостика между двумя половинами заставляет их самоорганизовываться в растворе (рис. 7). Они стабильны даже при нагревании до 60 ° C.
Двойные спиральные полимеры
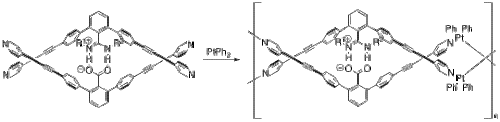
Яшима и его коллеги использовали солевые мостики для создания нескольких полимеров, которые принимают конформацию двойной спирали, как и ДНК.[24] В одном примере они включили платина для создания двойного спирального металлополимера.[25] Начиная с их мономера и бифенила платины (II) (рис. 8), их металлополимер самособирается через ряд обмен лиганда реакции. Две половины мономера скреплены вместе солевым мостиком между депротонированным карбоксилатом и протонированными атомами азота.
Рекомендации
- ^ а б Догерти, Деннис А. (2006). Современная физико-органическая химия. Саусалито, Калифорния: Университетские научные книги. ISBN 978-1-891389-31-3.
- ^ Маркус Й., Хефтер Дж. (Ноябрь 2006 г.). «Ионное спаривание». Химические обзоры. 106 (11): 4585–621. Дои:10.1021 / cr040087x. PMID 17091929.
- ^ Айзекс Н. (1996). Физическая органическая химия (2-е изд.). Англия: Лонгманс. ISBN 978-0582218635.[страница нужна ]
- ^ Ионные взаимодействия в природных и синтетических макромолекулах (А. Чиферри и А. Перико, ред.), 2012 John Wiley & Sons, Inc., стр. 35 и далее ISBN 978-0-470-52927-0.
- ^ Чейз М.В. (1998). Термохимические таблицы NIST-JANAF. J. Phys. Chem. Ref. Данные. Монография 9 (изд. Четвертое). С. 1–1951.
- ^ Шнайдер HJ, Яцимирский AK, ред. (2000). Принципы и методы супрамолекулярной химии. Чичестер: Вайли. ISBN 978-0-471-97253-2.[страница нужна ]
- ^ Бидерманн Ф., Шнайдер Х.Д. (май 2016 г.). «Экспериментальные энергии связи в супрамолекулярных комплексах». Химические обзоры. 116 (9): 5216–300. Дои:10.1021 / acs.chemrev.5b00583. PMID 27136957.
- ^ Шнайдер HJ (2009). «Механизмы связывания в супрамолекулярных комплексах». Angewandte Chemie. 48 (22): 3924–77. Дои:10.1002 / anie.200802947. PMID 19415701.
- ^ Даниэле П.Г., Фоти С., Джангуцца А., Пренести Е., Саммартано С. (2008). «Слабые комплексы щелочных и щелочноземельных металлов низкомолекулярных лигандов в водном растворе». Обзоры координационной химии. 252 (10–11): 1093–1107. Дои:10.1016 / j.ccr.2007.08.005.
- ^ Аль-Хаггар М., Мадей-Пиларчик А., Козловски Л., Буйницки Д. М., Яхья С., Абдель-Хади Д., Шамс А., Ахмад Н., Хамед С., Пузяновска-Кузницка М. (ноябрь 2012 г.). «Новая гомозиготная мутация LMNA p.Arg527Leu в двух неродственных египетских семьях вызывает перекрывающуюся мандибулоакральную дисплазию и синдром прогерии». Европейский журнал генетики человека. 20 (11): 1134–40. Дои:10.1038 / ejhg.2012.77. ЧВК 3476705. PMID 22549407.
- ^ а б Кумар С., Нусинов Р. (июль 2002 г.). «Близкие электростатические взаимодействия в белках». ChemBioChem. 3 (7): 604–17. Дои:10.1002 / 1439-7633 (20020703) 3: 7 <604 :: AID-CBIC604> 3.0.CO; 2-X. PMID 12324994.
- ^ а б Андерсон Д.Е., Бектель В.Дж., Далквист Ф.В. (март 1990 г.). «pH-индуцированная денатурация белков: один солевой мостик вносит 3-5 ккал / моль в свободную энергию сворачивания лизоцима Т4». Биохимия. 29 (9): 2403–8. Дои:10.1021 / bi00461a025. PMID 2337607.
- ^ Becktel WJ, Schellman JA (ноябрь 1987 г.). «Кривые стабильности белков». Биополимеры. 26 (11): 1859–77. Дои:10.1002 / bip.360261104. PMID 3689874.
- ^ а б Горовиц А., Серрано Л., Аврон Б., Байкрофт М., Фершт А. Р. (декабрь 1990 г.). «Сила и совместимость вкладов поверхностных солевых мостиков в стабильность белка». Журнал молекулярной биологии. 216 (4): 1031–44. Дои:10.1016 / S0022-2836 (99) 80018-7. PMID 2266554.
- ^ Sun DP, Sauer U, Nicholson H, Matthews BW (июль 1991 г.). «Вклад сконструированных поверхностных солевых мостиков в стабильность лизоцима Т4 определяется направленным мутагенезом». Биохимия. 30 (29): 7142–53. Дои:10.1021 / bi00243a015. PMID 1854726.
- ^ а б Strop P, Mayo SL (февраль 2000 г.). «Вклад поверхностных солевых мостиков в стабильность белка» (PDF). Биохимия. 39 (6): 1251–5. Дои:10.1021 / bi992257j. PMID 10684603.
- ^ Bianchi A, Bowman-James K, García-España E, ред. (1997). Супрамолекулярная химия анионов. Нью-Йорк: Wiley-VCH. ISBN 9780471186229.
- ^ Боуман-Джеймс К., Бьянки А., Гарсия-Испания Э, ред. (2012). Координационная химия анионов. Вайнхайм: Wiley-VCH. ISBN 978-3527323708.
- ^ Сесслер Дж. Л., Гейл П. А., Чо В. С., ред. (2006). Химия анионных рецепторов. Кембридж: Королевское химическое общество. ISBN 978-0854049745.
- ^ Гейл PA, Dehaen W, ред. (2010). Распознавание анионов в супрамолекулярной химии. Springer Science. Bibcode:2010arsc.book ..... G. ISBN 978-3642264702.
- ^ Busschaert N, Caltagirone C, Van Rossom W, Gale PA (май 2015 г.). «Приложения распознавания супрамолекулярных анионов». Химические обзоры. 115 (15): 8038–155. Дои:10.1021 / acs.chemrev.5b00099. PMID 25996028.
- ^ Evans NH, Beer PD (октябрь 2014 г.). «Достижения в области супрамолекулярной химии анионов: от признания к химическим применениям» (PDF). Angewandte Chemie International Edition. 53 (44): 11716–54. Дои:10.1002 / anie.201309937. PMID 25204549.
- ^ Куберски Б., Шумна А. (апрель 2009 г.). «Самособирающаяся хиральная капсула с полярным внутренним пространством». Химические коммуникации (15): 1959–61. Дои:10.1039 / b820990a. PMID 19333456.
- ^ Лю Дж., Лам Дж. В., Тан Б.З. (ноябрь 2009 г.). «Ацетиленовые полимеры: синтезы, структуры и функции». Химические обзоры. 109 (11): 5799–867. Дои:10.1021 / cr900149d. PMID 19678641.
- ^ Икеда М., Танака Ю., Хасэгава Т., Фурушо Ю., Яшима Е. (май 2006 г.). «Конструирование двухцепочечных металлосупрамолекулярных полимеров с контролируемой спиральностью путем сочетания солевых мостиков и координации металлов». Журнал Американского химического общества. 128 (21): 6806–7. Дои:10.1021 / ja0619096. PMID 16719458.