Лизин - Lysine
 L-лизин | |
| Имена | |
|---|---|
| Название ИЮПАК (2S) -2,6-Диаминогексановая кислота (L-лизин) (2р) -2,6-Диаминогексановая кислота (D-лизин) | |
| Другие имена Лизин, D-лизин, L-лизин, LYS, h-Lys-OH | |
| Идентификаторы | |
3D модель (JSmol ) |
|
| ЧЭБИ | |
| ЧЭМБЛ | |
| ChemSpider | |
| ECHA InfoCard | 100.000.673 |
| КЕГГ | |
PubChem CID | |
| UNII | |
| |
| |
| Свойства | |
| C6ЧАС14N2О2 | |
| Молярная масса | 146.190 г · моль−1 |
| 1,5 кг / л | |
| Фармакология | |
| B05XB03 (КТО) | |
| Страница дополнительных данных | |
| Показатель преломления (п), Диэлектрическая постоянная (εр), так далее. | |
Термодинамический данные | Фазовое поведение твердое тело – жидкость – газ |
| УФ, ИК, ЯМР, РС | |
Если не указано иное, данные для материалов приводятся в их стандартное состояние (при 25 ° C [77 ° F], 100 кПа). | |
| Ссылки на инфобоксы | |
Лизин (символ Lys или K)[1] является α-аминокислота который используется в биосинтезе белки. Он содержит α-аминогруппу (которая находится в протонированный −NH3+ образуются в биологических условиях), группа α-карбоновой кислоты (которая находится в депротонированной -COO− образуются в биологических условиях) и лизил боковой цепи ((CH2)4NH2), классифицируя его как основной заряженный (при физиологическом pH), алифатический аминокислота. это закодированный посредством кодоны AAA и AAG. Как почти все другие аминокислоты, α-углерод хиральный и лизин может относиться к энантиомер или рацемическая смесь обоих. Для целей данной статьи лизин будет относиться к биологически активному энантиомеру L-лизина, где α-углерод находится в S-конфигурации.
Организм человека не может синтезировать лизин. это необходим для людей и должны быть получены из рациона. У организмов, которые синтезируют лизин, он имеет два основных биосинтетические пути, то диаминопимелат и α-аминоадипат пути, которые используют различные ферменты и субстраты и встречаются в различных организмах. Лизин катаболизм происходит одним из нескольких путей, наиболее распространенным из которых является сахаропиновый путь.
Лизин играет несколько ролей в организме человека, в первую очередь протеиногенез, но также при сшивании коллаген полипептиды, поглощение основных минеральных питательных веществ и производство карнитин, что является ключевым в метаболизм жирных кислот. Лизин также часто участвует в модификации гистонов, и, таким образом, влияет на эпигеном. В ε -аминогруппа часто участвует в водородных связях и в качестве общего основания в катализ. Ε-аммоний группа (NH3+) присоединен к четвертому атому углерода от α-углерода, который присоединен к карбоксильной (C = OOH) группе.[2]
Из-за его важности в нескольких биологических процессах недостаток лизина может привести к нескольким болезненным состояниям, включая дефекты соединительной ткани, нарушение метаболизма жирных кислот, анемию и системную белково-энергетическую недостаточность. Напротив, переизбыток лизина, вызванный неэффективным катаболизмом, может вызвать тяжелые неврологические расстройства.
Лизин был впервые выделен немецким химиком-биологом Фердинандом Генрихом Эдмундом Дрекселем в 1889 году из белка. казеин в молоке.[3] Он назвал это "лизин".[4] В 1902 году немецкие химики Эмиль Фишер и Фриц Вейгерт определили химическую структуру лизина, синтезируя его.[5]
Биосинтез
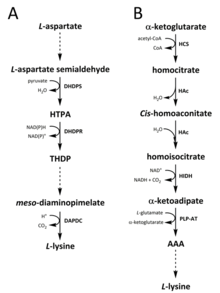
В природе идентифицированы два пути синтеза лизина. В диаминопимелатный (DAP) путь принадлежит к аспартат производное биосинтетическое семейство, которое также участвует в синтезе треонин, метионин и изолейцин.[6][7] В то время как α-аминоадипатный (AAA) путь является частью глутамат биосинтетическая семья.[8][9]
Путь DAP находится в обоих прокариоты и растений и начинается с дигидродипиколинатсинтаза (DHDPS) (E.C 4.3.3.7) катализированный реакция конденсации между производным аспартата, L-аспартат полуальдегид и пируват сформировать (4S) -4-гидрокси-2,3,4,5-тетрагидро- (2S) -дипиколиновая кислота (HTPA).[10][11][12][13][14] Затем продукт уменьшенный от дигидродипиколинатредуктаза (DHDPR) (E.C 1.3.1.26), с НАД (P) H в качестве донора протона с получением 2,3,4,5-тетрагидродипиколината (THDP).[15] С этого момента были обнаружены четыре варианта пути, а именно пути ацетилазы, аминотрансферазы, дегидрогеназы и сукцинилазы.[6][16] Пути как ацетилазы, так и варианта сукцинилазы используют четыре фермент В случае катализируемых стадий аминотрансферазный путь использует два фермента, а дегидрогеназный путь использует один фермент.[17] Эти четыре варианта пути сходятся при образовании предпоследнего продукта, мезо‑Диаминопимелат, который впоследствии ферментативно декарбоксилированный в необратимой реакции, катализируемой диаминопимелатдекарбоксилаза (DAPDC) (E.C 4.1.1.20) для производства L-лизин.[18][19] Путь DAP регулируется на нескольких уровнях, включая вышестоящие ферменты, участвующие в процессинге аспартата, а также на начальной стадии конденсации, катализируемой DHDPS.[19][20] Лизин придает сильный негативный отзыв петля на этих ферментах и, следовательно, регулирует весь путь.[20]
Путь AAA включает конденсацию α-кетоглутарат и ацетил-КоА через промежуточный AAA для синтеза L-лизин. Было показано, что этот путь присутствует в нескольких дрожжи виды, а также простейшие и высшие грибы.[9][21][22][23][24][25][26] Также сообщалось, что альтернативный вариант маршрута AAA был обнаружен в Термус термофильный и Pyrococcus horikoshii, что может указывать на то, что этот путь более широко распространен у прокариот, чем предполагалось изначально.[27][28][29] Первый и ограничивающий шаг в пути AAA является реакция конденсации между ацетил-CoA и α ‑ кетоглутаратом, катализируемая гомоцитрат-синтаза (HCS) (E.C 2.3.3.14), чтобы получить промежуточный гомоцитрил ‑ КоА, который является гидролизованный тем же ферментом для производства гомоцитрат.[30] Гомоцитрат ферментативно обезвоженный от гомоаконитаза (HAc) (E.C 4.2.1.36), чтобы получить СНГ-homoaconitate.[31] Затем HAc катализирует вторую реакцию, в которой СНГ-homoaconitate подвергается регидратация производить гомоизоцитрат.[9] Полученный продукт подвергается окислительный декарбоксилирование гомоизоцитратдегидрогеназа (HIDH) (E.C 1.1.1.87) с образованием α ‑ кетоадипата.[9] Затем AAA формируется через пиридоксаль-5'-фосфат (PLP) -зависимый аминотрансфераза (PLP-AT) (E.C 2.6.1.39), используя глутамат в качестве донора аминогруппы.[30] С этого момента путь AAA меняется в зависимости от [чего-то здесь не хватает? -> по крайней мере, заголовок раздела! ] о королевстве. У грибов AAA восстанавливается до α ‑ аминоадипат-полуальдегида посредством AAA-редуктазы (E.C 1.2.1.95) в уникальном процессе, включающем как аденилирование и сокращение, которое активируется фосфопантетеинилтрансфераза (E.C 2.7.8.7).[9] Как только полуальдегид образуется, сахаропин редуктаза (E.C 1.5.1.10) катализирует реакцию конденсации с глутаматом и NAD (P) H в качестве донора протонов, а я добываю восстанавливается с образованием предпоследнего продукта - сахаропина.[29] Последний этап пути развития грибов включает: сахаропиндегидрогеназа (SDH) (E.C 1.5.1.8) катализируемый окислительный дезаминирование сахаропина, в результате чего L-лизин.[9] В варианте пути AAA, обнаруженном у некоторых прокариот, AAA сначала превращается в N‑Ацетил-α-аминоадипат, который является фосфорилированный а затем редуктивно дефосфорилированный к ε-альдегиду.[29][30] Тогда альдегид трансаминированный к N‑Ацетил-лизин, который деацетилируется с образованием L-лизин.[29][30] Однако ферменты, участвующие в этом вариантном пути, нуждаются в дальнейшей проверке.
Катаболизм
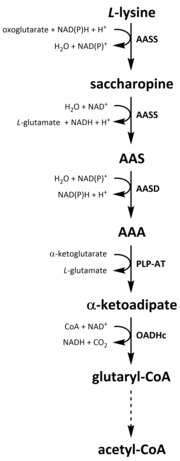
Как и все аминокислоты, катаболизм лизина образуется в результате поглощения пищевого лизина или расщепления внутриклеточный белок. Катаболизм также используется как средство контроля внутриклеточной концентрации свободного лизина и поддержания устойчивое состояние для предотвращения токсического действия чрезмерного количества свободного лизина.[32] В катаболизме лизина участвует несколько путей, но наиболее часто используется путь сахаропина, который в основном происходит в печень (и эквивалентные органы) у животных, особенно в пределах митохондрии.[33][32][34][35] Это противоположно ранее описанному пути AAA.[33][36] У животных и растений первые две стадии пути сахаропина катализируются бифункциональным ферментом, α-аминоадипиновая полуальдегидсинтаза (AASS), которые обладают как лизин-кетоглутаратредуктазой (LKR) (E.C 1.5.1.8), так и активностями SDH, тогда как у других организмов, таких как бактерии и грибы, оба этих фермента кодируются отдельными гены.[37][38] Первый шаг включает катализируемое LKR восстановление L-лизин в присутствии α-кетоглутарата для производства сахаропина, при этом НАД (Ф) Н действует как донор протонов.[39] Затем сахаропин подвергается реакции дегидратации, катализируемой SDH в присутствии НАД+, чтобы производить ААС и глутамат.[40] Дегидрогеназа AAS (AASD) (E.C 1.2.1.31) затем дополнительно дегидратирует молекулу до AAA.[39] Впоследствии PLP-AT катализирует реакцию, обратную реакции пути биосинтеза AAA, в результате чего AAA превращается в α-кетоадипат. Продукт, α ‑ кетоадипат, декарбоксилируется в присутствии НАД.+ и кофермент А с образованием глутарил-КоА, однако фермент, участвующий в этом, еще полностью не выяснен.[41][42] Некоторые данные свидетельствуют о том, что комплекс 2-оксоадипатдегидрогеназы (OADHc), который структурно гомологичен субъединице E1 оксоглутаратдегидрогеназный комплекс (OGDHc) (E.C 1.2.4.2), отвечает за реакцию декарбоксилирования.[41][43] Наконец, глутарил-КоА окислительно декарбоксилируется до кротон-КоА посредством глутарил-КоА дегидрогеназа (E.C 1.3.8.6), который затем подвергается дальнейшей переработке с помощью нескольких ферментативных стадий с образованием ацетил-КоА; необходимый углерод метаболит участвует в цикл трикарбоновых кислот (TCA).[39][44][45][46]
Пищевая ценность
Лизин - одна из девяти незаменимых аминокислот человека.[47] Потребности человека в питании варьируются от ~ 60 мг · кг.−1· D−1 в младенчестве до ~ 30 мг · кг−1· D−1 у взрослых.[33] Это требование обычно выполняется в западное общество с приемом лизин из мясных и овощных источников значительно превышает рекомендуемые требования.[33] В вегетарианских диетах потребление лизина меньше из-за ограниченного количества лизина в зерновые культуры по сравнению с мясными источниками.[33]
Учитывая ограниченную концентрацию лизина в зерновых культурах, долгое время предполагалось, что содержание лизина можно увеличить за счет генетическая модификация практики.[48][49] Часто эти методы включают преднамеренное нарушение регуляции пути DAP посредством введения нечувствительных к лизину обратной связи. ортологи фермента DHDPS.[48][49] Эти методы имели ограниченный успех, вероятно, из-за токсичный побочные эффекты увеличения свободного лизина и косвенные эффекты на цикл TCA.[50] Растения накапливают лизин и другие аминокислоты в виде семян. запасные белки, находящийся в семенах растения, и представляет собой съедобный компонент зерновых культур.[51] Это подчеркивает необходимость не только увеличения свободного лизина, но и направления лизина на синтез стабильных запасных белков семян и, следовательно, увеличения питательной ценности потребляемого компонента сельскохозяйственных культур.[52][53] Хотя методы генетической модификации имели ограниченный успех, более традиционные селекция методы позволили изолировать "Качественная протеиновая кукуруза ", что значительно повысило уровень лизина и триптофан, также незаменимая аминокислота. Это увеличение содержания лизина объясняется непрозрачный-2 мутация, которая уменьшила транскрипция недостатка лизина зеин связанных запасных белков семян и, как следствие, увеличилось количество других белков, богатых лизином.[53][54] Обычно, чтобы преодолеть ограниченное содержание лизина в домашний скот корм, добавлен лизин промышленного производства.[55][56] Производственный процесс включает ферментативный культивирование Коринебактерии глутамикум и последующая очистка лизина.[55]
Диетические источники
Хорошими источниками лизина являются продукты с высоким содержанием белка, такие как яйца, мясо (особенно красное мясо, баранина, свинина и птица), соя, фасоль и горох, сыр (особенно пармезан) и определенная рыба (например, треска и сардины ).[57] Лизин - это ограничивающая аминокислота (незаменимая аминокислота, содержащаяся в наименьшем количестве в конкретном продукте питания) в большинстве злаки, но в большинстве импульсы (бобовые).[58] Вегетарианская диета или диета с низким содержанием животного белка может быть достаточной для белка, включая лизин, если она включает как зерновые, так и бобовые, но нет необходимости употреблять эти две группы продуктов в одном приеме пищи.
Считается, что пища содержит достаточное количество лизина, если в ней содержится не менее 51 мг лизина на грамм белка (так что белок составляет 5,1% лизина).[59] L-лизин HCl используется в качестве пищевая добавка, обеспечивая 80,03% L-лизина.[60] Таким образом, 1 г L-лизина содержится в 1,25 г L-лизина HCl.
| Еда | Лизин (% белка) |
|---|---|
| Рыбы | 9.19% |
| Говядина, фарш, 90% постная / 10% жирная, приготовленная | 8.31% |
| Курица, жарение, мясо и кожа, приготовленные, жареные | 8.11% |
| Фасоль адзуки (фасоль адзуки), зрелые семена, сырые | 7.53% |
| Молоко, обезжиренное | 7.48% |
| Соя, зрелые семена, сырые | 7.42% |
| Яйцо, целое, сырое | 7.27% |
| Горох, расколотые, зрелые семена, сырые | 7.22% |
| Чечевица, розовый, сырой | 6.97% |
| Фасоль, зрелые семена, сырые | 6.87% |
| Нута, (бобы гарбанзо, бенгальский грамм), зрелые семена, сырые | 6.69% |
| Фасоль, зрелые семена, сырые | 5.73% |
Биологические роли
Наиболее частая роль лизина - протеиногенез. Лизин часто играет важную роль в структура белка. Поскольку его боковая цепь содержит положительно заряженную группу на одном конце и длинную гидрофобный углеродный хвост близко к позвоночнику, лизин считается несколько амфипатический. По этой причине лизин можно найти скрытым, а также чаще всего в каналах растворителя и на внешней стороне белков, где он может взаимодействовать с водной средой.[61] Лизин также может способствовать стабильности белка, поскольку его ε-аминогруппа часто участвует в водородная связь, соляные мосты и ковалентный взаимодействия, чтобы сформировать База Шиффа.[61][62][63][64]
Вторая важная роль лизина заключается в эпигенетический регулирование посредством гистон модификация.[65][66] Существует несколько типов ковалентных модификаций гистонов, которые обычно включают остатки лизина, обнаруженные в выступающем хвосте гистонов. Модификации часто включают добавление или удаление ацетил (-CH3CO) формирование ацетиллизин или возвращаясь к лизину, до трех метил (‑CH3), убиквитин или белок сумо группа.[65][67][68][69][70] Различные модификации влияют на генная регуляция, в котором гены могут быть активированы или подавлены.
Лизин также играет ключевую роль в других биологических процессах, включая; структурные белки соединительной ткани, кальций гомеостаз, и метаболизм жирных кислот.[71][72][73] Было показано, что лизин участвует в сшивание между тремя спиральные полипептиды в коллаген, что обеспечивает его стабильность и предел прочности.[71][74] Этот механизм сродни роли лизина в стенки бактериальных клеток, в котором лизин (и мезо-диаминопимелат) имеют решающее значение для образования поперечных связей и, следовательно, стабильности клеточной стенки.[75] Эта концепция ранее рассматривалась как средство предотвращения нежелательного выпуска потенциально патогенный генетически модифицированные бактерии. Было предложено, чтобы ауксотрофный напряжение кишечная палочка (Икс1776) может использоваться для всех практик генетической модификации, поскольку штамм не может выжить без добавления DAP и, следовательно, не может существовать вне лабораторной среды.[76] Было высказано предположение, что лизин участвует в абсорбции кальция в кишечнике и задержке в почках и, таким образом, может играть роль в кальциевый гомеостаз.[72] Наконец, было показано, что лизин является предшественником карнитин, который переносит жирные кислоты в митохондрии, где они могут окисляться с выделением энергии.[73][77] Карнитин синтезируется из триметиллизин, который является продуктом распада определенных белков, так как такой лизин сначала должен быть включен в белки и метилирован, прежде чем он превратится в карнитин.[73] Однако у млекопитающих основным источником карнитина являются пищевые источники, а не превращение лизина.[73]
В опсины любить родопсин и зрительные опсины (кодируемые генами OPN1SW, OPN1MW, и OPN1LW ), ретинальдегид образует База Шиффа с консервативным остатком лизина, и взаимодействие света с ретинилиден группа вызывает передачу сигнала в цветовое зрение (Увидеть визуальный цикл подробнее).
Спорные роли
Давно обсуждается, что лизин при внутривенном или пероральном введении может значительно увеличить высвобождение гормоны роста.[78] Это привело к тому, что спортсмены стали использовать лизин как средство, способствующее росту мышц во время тренировок, однако на сегодняшний день не было обнаружено никаких существенных доказательств в поддержку этого применения лизина.[78][79]
Потому что Вирус простого герпеса (HSV) белки богаче аргинином и беднее лизином, чем клетки, которые они заражают, добавки лизина были опробованы в качестве лечения. Поскольку две аминокислоты поглощаются кишечником, регенерируются почками и перемещаются в клетки одним и тем же переносчики аминокислот, избыток лизина теоретически ограничивает количество аргинина, доступного для репликации вируса.[80] Клинические исследования не дают убедительных доказательств эффективности в качестве профилактический или при лечении вспышек ВПГ.[81][82] В ответ на утверждения продукта о том, что лизин может улучшать иммунный ответ на ВПГ, Европейское агентство по безопасности пищевых продуктов не обнаружил доказательств причинно-следственной связи. В том же обзоре, опубликованном в 2011 году, не было обнаружено никаких доказательств в поддержку утверждений о том, что лизин может снижать уровень холестерина, повышать аппетит, способствовать синтезу белка в любой роли, кроме обычного питательного вещества, или увеличивать абсорбцию или удержание кальция.[83]
Роли в болезни
Заболевания, связанные с лизином, являются результатом последующей обработки лизина, то есть включения в белки или модификации альтернативных биомолекул. Роль лизина в коллагене была изложена выше, однако недостаток лизина и гидроксилизин вовлеченные в сшивание пептидов коллагена, связаны с болезненным состоянием соединительной ткани.[84] Поскольку карнитин является ключевым метаболитом, полученным из лизина, участвующим в метаболизме жирных кислот, некачественная диета, в которой отсутствует достаточное количество карнитина и лизина, может привести к снижению уровня карнитина, что может иметь значительный каскадный эффект на здоровье человека.[77][85] Было также показано, что лизин играет роль в анемия, поскольку предполагается, что лизин влияет на поглощение утюг и, как следствие, концентрация ферритин в плазма крови.[86] Однако точный механизм действия еще предстоит выяснить.[86] Чаще всего дефицит лизина наблюдается в незападных обществах и проявляется как белково-энергетическая недостаточность, который оказывает глубокое и системное воздействие на здоровье человека.[87][88] Также есть наследственный генетическое заболевание, которое включает мутации в ферментах, ответственных за катаболизм лизина, а именно в бифункциональном ферменте AASS сахаропинового пути.[89] Из-за отсутствия катаболизма лизина аминокислота накапливается в плазме и у пациентов развивается гиперлизинемия, которые могут проявляться как бессимптомные, так и тяжелые неврологические нарушения, в том числе эпилепсия, атаксия, спастичность, и психомоторное нарушение.[89][90] Клиническое значение гиперлизинемии является предметом дискуссий в этой области, при этом некоторые исследования не обнаружили корреляции между физическими или умственными недостатками и гиперлизинемией.[91] В дополнение к этому, мутации в генах, связанных с метаболизмом лизина, участвуют в нескольких болезненных состояниях, включая пиридоксин-зависимая эпилепсия (Ген ALDH7A1 ), α-кетоадипическая и α-аминоадипическая ацидурия (Ген DHTKD1 ), и глутаровая ацидурия 1 типа (Ген GCDH ).[41][92][93][94][95]
Гиперлизинурия характеризуется высоким содержанием лизина в моче.[96] Часто это происходит из-за нарушение обмена веществ в котором белок участвует в распаде лизина, не работает из-за генетической мутации.[97] Это также может произойти из-за отказа почечный трубчатый транспорт.[97]
Использование лизина в кормах для животных
Производство лизина для кормов для животных - крупная мировая отрасль, объем производства в 2009 году составил почти 700 000 тонн при рыночной стоимости более 1,22 миллиарда евро.[98] Лизин является важной добавкой к корму для животных, поскольку он является ограничивающей аминокислотой при оптимизации роста некоторых животных, таких как свиньи и куры, для производства мяса. Добавка лизина позволяет использовать более дешевый растительный белок (например, кукурузу, а не соя ) при сохранении высоких темпов роста и ограничении загрязнения от выделения азота.[99] Однако, в свою очередь, загрязнение фосфатами является серьезным ущербом для окружающей среды, когда кукуруза используется в качестве корма для домашней птицы и свиней.[100]
Лизин промышленно производят путем микробной ферментации на основе, в основном, сахара. Исследования в области генной инженерии активно изучают штаммы бактерий, чтобы повысить эффективность производства и позволить получать лизин из других субстратов.[98]
В популярной культуре
Фильм 1993 года парк Юрского периода (на основе 1990 г. Майкл Крайтон роман с таким же названием ) Особенности динозавры это были генетически измененный так что они не могли производить лизин, пример инженерного ауксотрофия.[101] Это было известно как «непредвиденный случай лизина» и должно было предотвратить клонированный динозавров от выживания за пределами парка, заставляя их зависеть от добавок лизина, предоставляемых ветеринарным персоналом парка. На самом деле, никакие животные не способны продуцировать лизин (это незаменимая аминокислота ).[102]
В 1996 году лизин стал центром внимания дело об установлении цен, крупнейший в истории США. В Арчер Дэниэлс Мидленд Компани заплатила штраф в размере 100 миллионов долларов США, а трое ее руководителей были осуждены и отбыли тюремное заключение. Также виновными в деле о сговоре с ценами были признаны две японские фирмы (Аджиномото, Kyowa Hakko) и южнокорейской фирмы (Sewon).[103] Секретные видеозаписи заговорщиков, устанавливающих цену на лизин, можно найти в Интернете или запросив видео в антимонопольном отделе Министерства юстиции США. Этот случай лег в основу фильма. Информатор!, а книга с таким же названием.[104]
использованная литература
![]() Эта статья была адаптирована из следующего источника под CC BY 4.0 лицензия (2018 ) (отчеты рецензента ): «Лизин: биосинтез, катаболизм и роли» (PDF), WikiJournal of Science, 1 (1): 4, 1 июня 2018 г., Дои:10.15347 / WJS / 2018.004, ISSN 2470-6345, Викиданные Q55120301
Эта статья была адаптирована из следующего источника под CC BY 4.0 лицензия (2018 ) (отчеты рецензента ): «Лизин: биосинтез, катаболизм и роли» (PDF), WikiJournal of Science, 1 (1): 4, 1 июня 2018 г., Дои:10.15347 / WJS / 2018.004, ISSN 2470-6345, Викиданные Q55120301
- ^ «Совместная комиссия IUPAC-IUB по биохимической номенклатуре (JCBN). Номенклатура и символика аминокислот и пептидов. Рекомендации 1983 г.». Биохимический журнал. 219 (2): 345–373. 15 апреля 1984 г. Дои:10.1042 / bj2190345. ЧВК 1153490. PMID 6743224.
- ^ Лизин. Биологический проект, Департамент биохимии и молекулярной биофизики, Университет Аризоны.
- ^ Дрексель Э. (1889 г.). "Zur Kenntniss der Spaltungsprodukte des Caseïns" [[Вклад] в [наши] знания о продуктах расщепления казеина]. Journal für Praktische Chemie. 2-я серия (на немецком языке). 39: 425–429. Дои:10.1002 / prac.18890390135. На стр. 428, Дрехсел представил эмпирическую формулу хлороплатинатной соли лизина - C8ЧАС16N2О2Cl2• PtCl4 + H2O - но позже он признал, что эта формула неверна, потому что кристаллы соли содержали этанол вместо воды. Увидеть: Дрексель Э. (1891). "Der Abbau der Eiweissstoffe" [Разборка белков]. Archiv für Anatomie und Physiologie (на немецком языке): 248–278.; Дрехсель Э. "Zur Kenntniss der Spaltungsproducte des Caseïns" [Вклад] в [наши] знания о продуктах расщепления казеина] (на немецком языке): 254–260.
С п. 256:] "… Die darin enthaltene Базовая шляпа die Formel C6ЧАС14N2О2. Der anfängliche Irrthum ist dadurch veranlasst worden, dass das Chloroplatinat nicht, wie angenommen ward, Krystallwasser, sondern Krystallalkohol enthält,… " (... содержащаяся в нем база имеет [эмпирическую] формулу C6ЧАС14N2О2. Первоначальная ошибка была вызвана тем, что хлороплатинат содержал в кристалле не воду (как предполагалось), а этанол…)
Цитировать журнал требует| журнал =(Помогите) - ^ Дрексель Э. (1891). "Der Abbau der Eiweissstoffe" [Разборка белков]. Archiv für Anatomie und Physiologie (на немецком языке): 248–278.; Фишер Э (1891). "Ueber neue Spaltungsproducte des Leimes" [О новых продуктах расщепления желатина] (на немецком языке): 465–469.
С п. 469:] "… Умри База C6ЧАС14N2О2, welche mit dem Namen Lysin bezeichnet werden mag,… " (… Основание C6ЧАС14N2О2, который можно обозначить словом «лизин»,…) [Примечание: Эрнст Фишер был аспирантом Дрекселя.]
Цитировать журнал требует| журнал =(Помогите) - ^ Фишер Э, Вейгерт Ф (1902). «Synthese der α, ε - Diaminocapronsäure (Неактивный лизин)» [Синтез α, ε-диаминогексановой кислоты ([оптически] неактивный лизин)]. Berichte der Deutschen Chemischen Gesellschaft (на немецком). 35 (3): 3772–3778. Дои:10.1002 / cber.190203503211.
- ^ а б Хадсон А.О., Благословение К., Маседо П., Чаттерджи С.П., Сингх Б.К., Гилварг С., Леустек Т. (январь 2005 г.). «Биосинтез лизина в растениях: свидетельство варианта известных бактериальных путей». Biochimica et Biophysica Acta (BBA) - Общие предметы. 1721 (1–3): 27–36. Дои:10.1016 / j.bbagen.2004.09.008. PMID 15652176.
- ^ Веласко А.М., Легина Дж. И., Ласкано А. (октябрь 2002 г.). «Молекулярная эволюция путей биосинтеза лизина». Журнал молекулярной эволюции. 55 (4): 445–59. Дои:10.1007 / s00239-002-2340-2. PMID 12355264.
- ^ Миядзаки Т., Миядзаки Дж., Ямане Х., Нишияма М. (июль 2004 г.). «Альфа-аминоадипатаминотрансфераза из чрезвычайно термофильной бактерии Thermus thermophilus» (PDF). Микробиология. 150 (Pt 7): 2327–34. Дои:10.1099 / мик.0.27037-0. PMID 15256574.
- ^ а б c d е ж Сюй Х, Анди Б., Цянь Дж., Западный А. Х., Кук П. Ф. (2006). «Альфа-аминоадипатный путь биосинтеза лизина в грибах». Биохимия клетки и биофизика. 46 (1): 43–64. Дои:10.1385 / CBB: 46: 1: 43. PMID 16943623.
- ^ Аткинсон С.К., Договски С., Даунтон М.Т., Чаботар П.Е., Добсон Р.К., Джеррард Дж. А., Вагнер Дж., Перуджини М.А. (март 2013 г.). «Структурные, кинетические и компьютерные исследования Vitis vinifera DHDPS раскрывают новое понимание механизма аллостерического ингибирования, опосредованного лизином». Молекулярная биология растений. 81 (4–5): 431–46. Дои:10.1007 / s11103-013-0014-7. PMID 23354837.
- ^ Гриффин, доктор медицины, Биллаканти, Дж. М., Вейсон А., Келлер С., Мертенс HD, Аткинсон С. К., Добсон Р. К., Перуджини М. А., Джеррард Дж. А., Пирс Ф. Г. (2012). «Характеристика первых ферментов, участвующих в биосинтезе лизина у Arabidopsis thaliana». PLOS ONE. 7 (7): e40318. Дои:10.1371 / journal.pone.0040318. ЧВК 3390394. PMID 22792278.
- ^ Соареш да Коста Т.П., Маскрофт-Тейлор А.С., Добсон Р.К., Девениш С.Р., Джеймсон Г.Б., Джеррард Дж. А. (июль 2010 г.). «Насколько важен« незаменимый »лизин активного центра в дигидродипиколинатсинтазе?». Биохимия. 92 (7): 837–45. Дои:10.1016 / j.biochi.2010.03.004. PMID 20353808.
- ^ Соарес да Коста Т.П., Кристенсен Дж.Б., Десбуа С., Гордон С.Е., Гупта Р., Хоган С.Дж., Нельсон Т.Г., Даунтон М.Т., Гарди С.К., Эбботт Б.М., Вагнер Дж., Панджикар С., Перуджини М.А. (2015). «Анализ четвертичной структуры основного олигомерного фермента». Аналитическое ультрацентрифугирование. Методы в энзимологии. 562. С. 205–23. Дои:10.1016 / bs.mie.2015.06.020. ISBN 9780128029084. PMID 26412653.
- ^ Muscroft-Taylor AC, Soares da Costa TP, Gerrard JA (март 2010 г.). «Новое понимание механизма дигидродипиколинатсинтазы с использованием изотермической калориметрии титрования». Биохимия. 92 (3): 254–62. Дои:10.1016 / j.biochi.2009.12.004. PMID 20025926.
- ^ Кристенсен Дж. Б., Соареш да Коста Т. П., Фау П., Пирс Ф. Г., Панджикар С., Перуджини М. А. (ноябрь 2016 г.). «Структура и функция цианобактериальных DHDPS и DHDPR». Научные отчеты. 6 (1): 37111. Дои:10.1038 / srep37111. ЧВК 5109050. PMID 27845445.
- ^ Маккой А.Дж., Адамс Н.Э., Хадсон А.О., Гилварг С., Леустек Т., Маурелли А.Т. (ноябрь 2006 г.). «L, L-диаминопимелатаминотрансфераза, транс-царственный фермент, используемый хламидиями и растениями для синтеза диаминопимелата / лизина». Труды Национальной академии наук Соединенных Штатов Америки. 103 (47): 17909–14. Дои:10.1073 / pnas.0608643103. ЧВК 1693846. PMID 17093042.
- ^ Хадсон А.О., Гилварг С., Леустек Т. (май 2008 г.). «Биохимическая и филогенетическая характеристика нового пути биосинтеза диаминопимелата у прокариот позволяет идентифицировать дивергентную форму LL-диаминопимелатаминотрансферазы». Журнал бактериологии. 190 (9): 3256–63. Дои:10.1128 / jb.01381-07. ЧВК 2347407. PMID 18310350.
- ^ Певерелли М.Г., Перуджини М.А. (август 2015 г.). «Оптимизированный сопряженный анализ для количественной оценки активности диаминопимелатдекарбоксилазы». Биохимия. 115: 78–85. Дои:10.1016 / j.biochi.2015.05.004. PMID 25986217.
- ^ а б Соарес да Коста Т. П., Десбуа С., Договски С., Горман М. А., Кетарен Н. Э., Паксман Дж. Дж., Сиддики Т., Заммит Л. М., Эбботт Б. М., Робинс-Браун Р. М., Паркер М. В., Джеймсон Дж. Б., Холл СВ, Панджикар С., Перуджини М. А. (август 2016). «Структурные детерминанты, определяющие аллостерическое ингибирование основной антибиотической мишени». Структура. 24 (8): 1282–1291. Дои:10.1016 / j.str.2016.05.019. PMID 27427481.
- ^ а б Джандер Г., Джоши В. (1 января 2009 г.). «Биосинтез аминокислот, полученных из аспартата в Arabidopsis thaliana». Книга об арабидопсисе. 7: e0121. Дои:10.1199 / таб.0121. ЧВК 3243338. PMID 22303247.
- ^ Энди Б., Вест АХ, Кук П.Ф. (сентябрь 2004 г.). «Кинетический механизм меченой гистидином гомоцитрат-синтазы из Saccharomyces cerevisiae». Биохимия. 43 (37): 11790–5. Дои:10.1021 / bi048766p. PMID 15362863.
- ^ Бхаттачарджи Дж. К. (1985). «Альфа-аминоадипатный путь биосинтеза лизина у низших эукариот». Критические обзоры в микробиологии. 12 (2): 131–51. Дои:10.3109/10408418509104427. PMID 3928261.
- ^ Бхаттачарджи Дж. К., Страссман М. (май 1967 г.). «Накопление трикарбоновых кислот, связанных с биосинтезом лизина в дрожжевом мутанте». Журнал биологической химии. 242 (10): 2542–6. PMID 6026248.
- ^ Gaillardin CM, Ribet AM, Heslot H (ноябрь 1982 г.). «Дикий тип и мутантные формы гомоизоцитридегидрогеназы в дрожжах Saccharomycopsis lipolytica». Европейский журнал биохимии. 128 (2–3): 489–94. Дои:10.1111 / j.1432-1033.1982.tb06991.x. PMID 6759120.
- ^ Яклич В. М., Кубичек С. П. (июль 1990 г.). «Гомоцитратсинтаза из Penicillium chrysogenum. Локализация, очистка цитозольного изофермента и чувствительность к лизину». Биохимический журнал. 269 (1): 247–53. Дои:10.1042 / bj2690247. ЧВК 1131560. PMID 2115771.
- ^ Е ЧЖ, Бхаттачарджи Дж. К. (декабрь 1988 г.). «Путь биосинтеза лизина и биохимические блоки ауксотрофов лизина Schizosaccharomyces pombe». Журнал бактериологии. 170 (12): 5968–70. Дои:10.1128 / jb.170.12.5968-5970.1988. ЧВК 211717. PMID 3142867.
- ^ Кобаши Н., Нишияма М., Танокура М. (март 1999 г.). «Независимый от аспартаткиназы синтез лизина в чрезвычайно термофильной бактерии Thermus thermophilus: лизин синтезируется через альфа-аминоадипиновую кислоту, а не через диаминопимелиновую кислоту». Журнал бактериологии. 181 (6): 1713–8. Дои:10.1128 / JB.181.6.1713-1718.1999. ЧВК 93567. PMID 10074061.
- ^ Косуге Т., Хосино Т. (1999). «Альфа-аминоадипатный путь биосинтеза лизина широко распространен среди штаммов Thermus». Журнал биологии и биоинженерии. 88 (6): 672–5. Дои:10.1016 / S1389-1723 (00) 87099-1. PMID 16232683.
- ^ а б c d Нисида Х., Нишияма М., Кобаши Н., Косуге Т., Хосино Т., Ямане Х. (декабрь 1999 г.). «Кластер прокариотических генов, участвующих в синтезе лизина через аминоадипатный путь: ключ к эволюции биосинтеза аминокислот». Геномные исследования. 9 (12): 1175–83. Дои:10.1101 / гр.9.12.1175. PMID 10613839.
- ^ а б c d Нисида Х., Нишияма М. (сентябрь 2000 г.). «Что характерно для синтеза лизина грибами через альфа-аминоадипатный путь?». Журнал молекулярной эволюции. 51 (3): 299–302. Дои:10.1007 / s002390010091. PMID 11029074.
- ^ Забриски TM, Джексон MD (февраль 2000 г.). «Биосинтез и метаболизм лизина в грибах». Отчеты о натуральных продуктах. 17 (1): 85–97. Дои:10.1039 / a801345d. PMID 10714900.
- ^ а б Чжу X, Галили Г (май 2004 г.). «Метаболизм лизина одновременно регулируется синтезом и катаболизмом как в репродуктивных, так и в вегетативных тканях». Физиология растений. 135 (1): 129–36. Дои:10.1104 / стр.103.037168. ЧВК 429340. PMID 15122025.
- ^ а б c d е Томе Д., Бос С. (июнь 2007 г.). «Потребность в лизине на протяжении жизненного цикла человека». Журнал питания. 137 (6 Прил. 2): 1642S – 1645S. Дои:10.1093 / jn / 137.6.1642S. PMID 17513440.
- ^ Blemings KP, Crenshaw TD, Swick RW, Беневенга, штат Нью-Джерси (август 1994 г.). «Лизин-альфа-кетоглутаратредуктаза и сахаропиндегидрогеназа находятся только в митохондриальном матриксе в печени крысы». Журнал питания. 124 (8): 1215–21. Дои:10.1093 / jn / 124.8.1215. PMID 8064371.
- ^ Галили Г, Тан Г, Чжу Х, Гакьер Б. (июнь 2001 г.). «Катаболизм лизина: стресс и развитие сверхрегулируемого метаболического пути». Текущее мнение в области биологии растений. 4 (3): 261–6. Дои:10.1016 / с 1369-5266 (00) 00170-9. PMID 11312138.
- ^ Арруда П., Кемпер Э.Л., Папес Ф., Лейте А (август 2000 г.). «Регулирование катаболизма лизина у высших растений». Тенденции в растениеводстве. 5 (8): 324–30. Дои:10.1016 / с 1360-1385 (00) 01688-5. PMID 10908876.
- ^ Sacksteder KA, Biery BJ, Morrell JC, Goodman BK, Geisbrecht BV, Cox RP, Gould SJ, Geraghty MT (июнь 2000 г.). «Идентификация гена альфа-аминоадипической полуальдегидсинтазы, который является дефектным при семейной гиперлизинемии». Американский журнал генетики человека. 66 (6): 1736–43. Дои:10.1086/302919. ЧВК 1378037. PMID 10775527.
- ^ Чжу X, Тан Г, Галили Г (декабрь 2002 г.). «Активность бифункционального фермента лизин-кетоглутарат редуктазы / сахаропиндегидрогеназы арабидопсиса при катаболизме лизина регулируется функциональным взаимодействием между двумя его ферментными доменами». Журнал биологической химии. 277 (51): 49655–61. Дои:10.1074 / jbc.m205466200. PMID 12393892.
- ^ а б c Киёта Э, Пена И.А., Арруда П. (ноябрь 2015 г.). «Путь сахаропина в развитии семян и стрессовая реакция кукурузы». Растения, клетки и окружающая среда. 38 (11): 2450–61. Дои:10.1111 / pce.12563. PMID 25929294.
- ^ Серрано Г.К., Резенде и Сильва Фигейра Т., Кийота Е., Заната Н., Арруда П. (март 2012 г.). «Распад лизина через сахаропиновый путь в бактериях: LKR и SDH в бактериях и его связь с ферментами растений и животных». Письма FEBS. 586 (6): 905–11. Дои:10.1016 / j.febslet.2012.02.023. PMID 22449979.
- ^ а б c Данхаузер К., Зауэр С.В., Хаак Т.Б., Виланд Т., Штауфнер С., Граф Е., Цшок Дж., Стром Т.М., Трауб Т., Окун Дж. Г., Мейтингер Т., Хоффманн Г. Ф., Прокиш Г., Кёлькер С. (декабрь 2012 г.). «Мутации DHTKD1 вызывают 2-аминоадипическую и 2-оксоадипиновую ацидурию». Американский журнал генетики человека. 91 (6): 1082–7. Дои:10.1016 / j.ajhg.2012.10.006. ЧВК 3516599. PMID 23141293.
- ^ Sauer SW, Opp S, Hoffmann GF, Koeller DM, Okun JG, Kölker S (январь 2011 г.). «Терапевтическая модуляция церебрального метаболизма L-лизина на мышиной модели глутаровой ацидурии I типа». Мозг. 134 (Pt 1): 157–70. Дои:10.1093 / мозг / awq269. PMID 20923787.
- ^ Гонсалвес Р.Л., Буник В.И., Brand MD (февраль 2016 г.). «Производство супероксида / пероксида водорода митохондриальным комплексом 2-оксоадипатдегидрогеназы». Свободная радикальная биология и медицина. 91: 247–55. Дои:10.1016 / j.freeradbiomed.2015.12.020. PMID 26708453.
- ^ Goh DL, Patel A, Thomas GH, Salomons GS, Schor DS, Jakobs C, Geraghty MT (июль 2002 г.). «Характеристика человеческого гена, кодирующего альфа-аминоадипатаминотрансферазу (AADAT)». Молекулярная генетика и метаболизм. 76 (3): 172–80. Дои:10.1016 / с1096-7192 (02) 00037-9. PMID 12126930.
- ^ Härtel U, Eckel E, Koch J, Fuchs G, Linder D, Buckel W. (1 февраля 1993 г.). «Очистка глутарил-КоА дегидрогеназы из Pseudomonas sp., Фермента, участвующего в анаэробной деградации бензоата». Архив микробиологии. 159 (2): 174–81. Дои:10.1007 / bf00250279. PMID 8439237.
- ^ Sauer SW (октябрь 2007 г.). «Биохимия и биоэнергетика дефицита глутарил-КоА дегидрогеназы». Журнал наследственных метаболических заболеваний. 30 (5): 673–80. Дои:10.1007 / s10545-007-0678-8. PMID 17879145.
- ^ Нельсон Д.Л., Кокс М.М., Ленингер А.Л. (2013). Принципы биохимии Ленингера (6-е изд.). Нью-Йорк: W.H. Фримен и компания. ISBN 978-1-4641-0962-1. OCLC 824794893.
- ^ а б Галили Г., Амир Р. (февраль 2013 г.). «Обогащение растений незаменимыми аминокислотами лизином и метионином для улучшения качества питания». Журнал биотехнологии растений. 11 (2): 211–22. Дои:10.1111 / pbi.12025. PMID 23279001.
- ^ а б Ван Г., Сюй М., Ван В., Галили Г. (июнь 2017 г.). «Обогащение садовых культур незаменимыми аминокислотами: обзор». Международный журнал молекулярных наук. 18 (6): 1306. Дои:10.3390 / ijms18061306. ЧВК 5486127. PMID 28629176.
- ^ Ангеловичи Р., Файт А., Ферни А.Р., Галили Г. (январь 2011 г.). «Признак семян с высоким содержанием лизина отрицательно связан с циклом TCA и замедляет прорастание семян Arabidopsis». Новый Фитолог. 189 (1): 148–59. Дои:10.1111 / j.1469-8137.2010.03478.x. PMID 20946418.
- ^ Эдельман М, Кольт М (2016). «Питательная ценность листа по сравнению с семенами». Границы химии. 4: 32. Дои:10.3389 / fchem.2016.00032. ЧВК 4954856. PMID 27493937.
- ^ Цзян С.Ю., Ма А., Се Л., Рамачандран С. (сентябрь 2016 г.). «Повышение содержания и качества протеина за счет чрезмерной экспрессии искусственно синтетических гибридных белков с высоким содержанием лизина и треонина в растениях риса». Научные отчеты. 6 (1): 34427. Дои:10.1038 / srep34427. ЧВК 5039639. PMID 27677708.
- ^ а б Shewry PR (ноябрь 2007 г.). «Повышение содержания белка и состава зерна злаков». Журнал зерновых наук. 46 (3): 239–250. Дои:10.1016 / j.jcs.2007.06.006.
- ^ Прасанна Б., Васал С.К., Кассахун Б., Сингх Н.Н. (2001). «Качественный протеин кукурузы». Текущая наука. 81 (10): 1308–1319. JSTOR 24105845.
- ^ а б Кирхер М., Пфефферле В. (апрель 2001 г.). «Ферментативное производство L-лизина в качестве добавки к корму для животных». Атмосфера. 43 (1): 27–31. Дои:10.1016 / с0045-6535 (00) 00320-9. PMID 11233822.
- ^ Джуниор Л., Альберто Л., Летти Г. В., Соккол CR, Джуниор Л., Альберто Л., Летти Г. В., Соккол К. Р. (2016). «Разработка обогащенных L-лизином отрубей для питания животных посредством глубокой ферментации с помощью Corynebacterium glutamicum с использованием агропромышленных субстратов». Бразильский архив биологии и технологий. 59. Дои:10.1590/1678-4324-2016150519. ISSN 1516-8913.
- ^ Медицинский центр Университета Мэриленда. «Лизин». Получено 30 декабря 2009.
- ^ Молодой В.Р., Пеллетт П.Л. (1994). «Растительные белки в отношении человеческого белка и аминокислотного питания». Американский журнал клинического питания. 59 (5 Прил.): 1203S – 1212S. Дои:10.1093 / ajcn / 59.5.1203s. PMID 8172124.
- ^ Институт медицины национальных академий. «Рекомендуемая диета для макроэлементов». п. 589. Получено 29 октября 2017.
- ^ «База данных диетических добавок: информация о смеси (DSBI)».
L-ЛИЗИН HCL 10000820 80,03% лизина
- ^ а б Беттс MJ, Рассел РБ (2003). Барнс М.Р., Грей И.К. (ред.). Биоинформатика для генетиков. John Wiley & Sons, Ltd., стр. 289–316. Дои:10.1002 / 0470867302.ch14. ISBN 978-0-470-86730-3.
- ^ Blickling S, Renner C, Laber B, Pohlenz HD, Holak TA, Huber R (январь 1997 г.). «Механизм реакции дигидродипиколинатсинтазы Escherichia coli исследован методами рентгеновской кристаллографии и ЯМР-спектроскопии». Биохимия. 36 (1): 24–33. Дои:10.1021 / bi962272d. PMID 8993314.
- ^ Кумар С., Цай С.Дж., Нусинов Р. (март 2000 г.). «Факторы, повышающие термостабильность белков». Белковая инженерия. 13 (3): 179–91. Дои:10.1093 / белок / 13.3.179. PMID 10775659.
- ^ Sokalingam S, Raghunathan G, Soundrarajan N, Lee SG (9 июля 2012 г.). «Исследование влияния поверхностного лизина на мутагенез аргинина на стабильность и структуру белка с использованием зеленого флуоресцентного белка». PLOS ONE. 7 (7): e40410. Дои:10.1371 / journal.pone.0040410. ЧВК 3392243. PMID 22792305.
- ^ а б Дамбахер С., Хан М., Шотта Г. (июль 2010 г.). «Эпигенетическая регуляция развития путем метилирования гистонового лизина». Наследственность. 105 (1): 24–37. Дои:10.1038 / hdy.2010.49. PMID 20442736.
- ^ Мартин С., Чжан И (ноябрь 2005 г.). «Разнообразные функции метилирования гистонового лизина». Обзоры природы. Молекулярная клеточная биология. 6 (11): 838–49. Дои:10.1038 / nrm1761. PMID 16261189.
- ^ Блэк Дж. К., Ван Речем С., Ветстайн Дж. Р. (ноябрь 2012 г.). «Динамика метилирования лизина гистонов: установление, регуляция и биологическое воздействие». Молекулярная клетка. 48 (4): 491–507. Дои:10.1016 / j.molcel.2012.11.006. ЧВК 3861058. PMID 23200123.
- ^ Чоудхари К., Кумар С., Гнад Ф, Нильсен М.Л., Рехман М., Вальтер Т.К., Олсен Дж. В., Манн М. (август 2009 г.). «Ацетилирование лизина нацелено на белковые комплексы и ко-регулирует основные клеточные функции». Наука. 325 (5942): 834–40. Дои:10.1126 / science.1175371. PMID 19608861.
- ^ Шио Ю., Эйзенман Р. Н. (ноябрь 2003 г.). «Сумоилирование гистонов связано с репрессией транскрипции». Труды Национальной академии наук Соединенных Штатов Америки. 100 (23): 13225–30. Дои:10.1073 / pnas.1735528100. ЧВК 263760. PMID 14578449.
- ^ Ван Х, Ван Л., Эрдджумент-Бромаж Х, Видал М., Темпст П., Джонс Р. С., Чжан Ю. (октябрь 2004 г.). «Роль убиквитинирования гистона H2A в сайленсинге Polycomb». Природа. 431 (7010): 873–8. Дои:10.1038 / природа02985. HDL:10261/73732. PMID 15386022.
- ^ а б Плечи MD, Рейнс RT (2009). «Структура и стабильность коллагена». Ежегодный обзор биохимии. 78: 929–58. Дои:10.1146 / annurev.biochem.77.032207.120833. ЧВК 2846778. PMID 19344236.
- ^ а б Чивителли Р., Виллареал Д. Т., Агнусдей Д., Нарди П., Авиоли Л. В., Дженнари С. (1992). «Диетический метаболизм L-лизина и кальция у человека». Питание. 8 (6): 400–5. PMID 1486246.
- ^ а б c d Vaz FM, Wanders RJ (февраль 2002 г.). «Биосинтез карнитина у млекопитающих». Биохимический журнал. 361 (Pt 3): 417–29. Дои:10.1042 / bj3610417. ЧВК 1222323. PMID 11802770.
- ^ Ямаути М., Срихолпеч М. (25 мая 2012 г.). «Посттрансляционные модификации лизина коллагена». Очерки биохимии. 52: 113–33. Дои:10.1042 / bse0520113. ЧВК 3499978. PMID 22708567.
- ^ Воллмер В., Блано Д., де Педро М.А. (март 2008 г.). «Структура и архитектура пептидогликанов». Обзор микробиологии FEMS. 32 (2): 149–67. Дои:10.1111 / j.1574-6976.2007.00094.x. PMID 18194336.
- ^ Кертисс Р. (май 1978 г.). «Биологическое сдерживание и переносимость вектора клонирования». Журнал инфекционных болезней. 137 (5): 668–75. Дои:10.1093 / infdis / 137.5.668. PMID 351084.
- ^ а б Фланаган Дж. Л., Симмонс П. А., Вехиге Дж., Уиллкокс Мэриленд, Гаррет К. (апрель 2010 г.). «Роль карнитина в заболевании». Питание и обмен веществ. 7: 30. Дои:10.1186/1743-7075-7-30. ЧВК 2861661. PMID 20398344.
- ^ а б Хромиак Дж. А., Антонио Дж. (2002). «Использование спортсменами аминокислот в качестве высвобождающих гормон роста агентов». Питание. 18 (7–8): 657–61. Дои:10.1016 / s0899-9007 (02) 00807-9. PMID 12093449.
- ^ Corpas E, Blackman MR, Roberson R, Scholfield D, Harman SM (июль 1993 г.). «Пероральный аргинин-лизин не увеличивает гормон роста или инсулиноподобный фактор роста-I у пожилых мужчин». Журнал геронтологии. 48 (4): M128–33. Дои:10.1093 / geronj / 48.4.M128. PMID 8315224.
- ^ Габи AR (2006). «Натуральные средства от простого герпеса». Альтернативная медицина Rev. 11 (2): 93–101. PMID 16813459.
- ^ Томблин Ф.А., Лукас К.Х. (2001). «Лизин для лечения лабиального герпеса». Am J Health Syst Pharm. 58 (4): 298–300, 304. Дои:10.1093 / ajhp / 58.4.298. PMID 11225166.
- ^ Chi CC, Wang SH, Delamere FM, Wojnarowska F, Peters MC, Kanjirath PP (7 августа 2015 г.). «Вмешательства для профилактики простого герпеса на губах (герпес на губах)». Кокрановская база данных систематических обзоров (8): CD010095. Дои:10.1002 / 14651858.CD010095.pub2. ЧВК 6461191. PMID 26252373.
- ^ «Научное заключение по обоснованию заявлений о здоровье, связанных с L-лизином и иммунной защитой от вируса герпеса (ID 453), поддержанием нормальной концентрации холестерина ЛПНП в крови (ID 454, 4669), повышением аппетита, ведущим к увеличению энергии». Журнал EFSA. 9 (4): 2063. 2011. Дои:10.2903 / j.efsa.2011.2063. ISSN 1831-4732.
- ^ Пиннелл С.Р., Крейн С.М., Кензора Дж.Э., Глимчер М.Дж. (май 1972 г.). «Наследственное заболевание соединительной ткани. Коллагеновая болезнь с дефицитом гидроксилизина». Медицинский журнал Новой Англии. 286 (19): 1013–20. Дои:10.1056 / NEJM197205112861901. PMID 5016372.
- ^ Рудман Д., Сьюэлл К. В., Ансли Дж. Д. (сентябрь 1977 г.). «Дефицит карнитина у пациентов с кахектическим циррозом». Журнал клинических исследований. 60 (3): 716–23. Дои:10.1172 / jci108824. ЧВК 372417. PMID 893675.
- ^ а б Раштон Д.Х. (июль 2002 г.). «Факторы питания и выпадение волос». Клиническая и экспериментальная дерматология. 27 (5): 396–404. Дои:10.1046 / j.1365-2230.2002.01076.x. PMID 12190640.
- ^ Эмери П.В. (октябрь 2005 г.). «Метаболические изменения при недоедании». Глаз. 19 (10): 1029–34. Дои:10.1038 / sj.eye.6701959. PMID 16304580.
- ^ Гош С., Смрига М., Вувор Ф., Сури Д., Мохаммед Х., Армах С.М., Скримшоу Н.С. (октябрь 2010 г.). «Влияние добавок лизина на здоровье и заболеваемость у лиц, принадлежащих к бедным пригородным домохозяйствам в Аккре, Гана». Американский журнал клинического питания. 92 (4): 928–39. Дои:10.3945 / ajcn.2009.28834. PMID 20720257.
- ^ а б Houten SM, Te Brinke H, Denis S, Ruiter JP, Knegt AC, de Klerk JB, Augoustides-Savvopoulou P, Häberle J, Baumgartner MR, Coşkun T, Zschocke J, Sass JO, Poll-The BT, Wanders RJ, Duran M (Апрель 2013). «Генетические основы гиперлизинемии». Журнал редких заболеваний Orphanet. 8: 57. Дои:10.1186/1750-1172-8-57. ЧВК 3626681. PMID 23570448.
- ^ Хоффманн Г.Ф., Кёлькер С. (2016). Врожденные метаболические заболевания. Шпрингер, Берлин, Гейдельберг. С. 333–348. Дои:10.1007/978-3-662-49771-5_22. ISBN 978-3-662-49769-2.
- ^ Дансис Дж., Хатцлер Дж., Ампола М.Г., Ши В.Э., ван Гелдерен Х.Х., Кирби Л.Т., Вуди, Северная Каролина (май 1983 г.). «Прогноз гиперлизинемии: промежуточный отчет». Американский журнал генетики человека. 35 (3): 438–42. ЧВК 1685659. PMID 6407303.
- ^ Миллс П.Б., Струйс Э., Якобс С., Плекко Б., Бакстер П., Баумгартнер М., Виллемсен М.А., Омран Х., Тэк У., Уленберг Б., Вешке Б., Клейтон П.Т. (март 2006 г.). «Мутации антиквитина у людей с пиридоксин-зависимыми судорогами». Природа Медицина. 12 (3): 307–9. Дои:10,1038 / нм 1366. PMID 16491085.
- ^ Mills PB, Footitt EJ, Mills KA, Tuschl K, Aylett S, Varadkar S, Hemingway C, Marlow N, Rennie J, Baxter P, Dulac O, Nabbout R, Craigen WJ, Schmitt B, Feillet F, Christensen E, De Lonlay П, Пайк М.Г., Хьюз М.И., Струйс Э.А., Якобс С., Зубери С.М., Клейтон П.Т. (июль 2010 г.). «Генотипический и фенотипический спектр пиридоксин-зависимой эпилепсии (дефицит ALDH7A1)». Мозг. 133 (Pt 7): 2148–59. Дои:10.1093 / мозг / awq143. ЧВК 2892945. PMID 20554659.
- ^ Hagen J, te Brinke H, Wanders RJ, Knegt AC, Oussoren E, Hoogeboom AJ, Ruijter GJ, Becker D, Schwab KO, Franke I, Duran M, Waterham HR, Sass JO, Houten SM (сентябрь 2015 г.). «Генетические основы альфа-аминоадипической и альфа-кетоадипической ацидурии». Журнал наследственных метаболических заболеваний. 38 (5): 873–9. Дои:10.1007 / s10545-015-9841-9. PMID 25860818.
- ^ Хедлунд Г.Л., Лонго Н., Паскуали М. (май 2006 г.). «Глутаровая ацидемия 1 типа». Американский журнал медицинской генетики, часть C: Семинары по медицинской генетике. 142C (2): 86–94. Дои:10.1002 / ajmg.c.30088. ЧВК 2556991. PMID 16602100.
- ^ «Гиперлизинурия | Определите гиперлизинурию на Dictionary.com».
- ^ а б Уолтер, Джон; Джон Фернандес; Жан-Мари Саудубре; Жорж ван ден Берге (2006). Врожденные метаболические заболевания: диагностика и лечение. Берлин: Springer. п. 296. ISBN 978-3-540-28783-4.
- ^ а б «Норвежский грант на улучшение процесса производства лизина». Все о корме. 26 января 2010. Архивировано с оригинал 11 марта 2012 г.
- ^ Торид Y (2004). «Лизин и другие аминокислоты для кормов: производство и вклад в использование белка в кормлении животных». Источники белка для производства кормов для животных; Консультации экспертов и семинар ФАО по источникам белка для кормовой промышленности; Бангкок, 29 апреля - 3 мая 2002 г.. Рим: Продовольственная и сельскохозяйственная организация Объединенных Наций. ISBN 978-92-5-105012-5.
- ^ Абельсон PH (март 1999 г.). «Возможный фосфатный кризис». Наука. 283 (5410): 2015. Дои:10.1126 / science.283.5410.2015. PMID 10206902.
- ^ Койн Дж. А. (10 октября 1999 г.). "Истина не там". Нью-Йорк Таймс. Получено 6 апреля 2008.
- ^ Ву Джи (май 2009 г.). «Аминокислоты: обмен веществ, функции и питание». Аминокислоты. 37 (1): 1–17. Дои:10.1007 / s00726-009-0269-0. PMID 19301095.
- ^ Коннор Дж. М. (2008). Фиксация мировых цен (2-е изд.). Гейдельберг: Springer-Verlag. ISBN 978-3-540-78669-6.
- ^ Эйхенвальд К. (2000). Информатор: правдивая история. Нью-Йорк: Бродвейские книги. ISBN 978-0-7679-0326-4.
| Общие темы |  | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| По свойствам |
| ||||||||||
| |||||||||||