Альфа-спираль - Alpha helix

В альфа спираль (α-спираль) является обычным мотив в вторичная структура из белки и является правая рука -спираль строение, в котором каждый позвоночник N − H группа водородные связи к позвоночнику C = O группа аминокислота расположены три или четыре остатки ранее по последовательности белка.
Альфа-спираль еще называют классической Α-спираль Полинга – Кори – Брэнсона. Название 3.613-спираль также используется для этого типа спирали, обозначающего среднее количество остатков на виток спирали, при этом 13 атомов участвуют в кольце, образованном водородной связью.
Среди типов локальной структуры белков α-спираль является наиболее экстремальной и наиболее предсказуемой по последовательности, а также наиболее распространенной.

Открытие


В начале 1930-х гг. Уильям Эстбери показали, что произошли кардинальные изменения в рентгеновский снимок дифракция волокна влажных волокон шерсти или волос при значительном растяжении. Данные свидетельствуют о том, что нерастянутые волокна имели спиральную молекулярную структуру с характерным повторением ≈5,1. Ангстремс (0.51 нанометры ).
Первоначально Эстбери предложила для волокон структуру изогнутой цепи. Позже он присоединился к другим исследователям (особенно к американскому химику Морис Хаггинс ) предлагая следующее:
- нерастянутые белковые молекулы образовали спираль (которую он назвал α-формой)
- растяжение заставляло спираль разматываться, образуя расширенное состояние (которое он назвал β-формой).
Несмотря на некорректность деталей, модели этих форм, представленные Эстбери, были правильными по сути и соответствовали современным элементам вторичная структура, α-спираль и β-цепь (Номенклатура Астбери была сохранена), которые были разработаны Линус Полинг, Роберт Кори и Герман Брэнсон в 1951 г. (см. ниже); в этой статье показаны как правые, так и левые спирали, хотя в 1960 г. кристаллическая структура миоглобина[1] показал, что правая форма является распространенной. Ганс Нейрат был первым, кто показал, что модели Астбери не могут быть точными в деталях, потому что они включают столкновения атомов.[2] Статья Нейрата и данные Эстбери вдохновили Х. С. Тейлор,[3] Морис Хаггинс[4] и Брэгг и соавторы[5] предлагать модели кератин которые чем-то напоминают современную α-спираль.
Двумя ключевыми достижениями в моделировании современной α-спирали были: правильная геометрия связки благодаря определение кристаллической структуры из аминокислоты и пептиды и предсказание Полинга планарный пептидные связи; и его отказ от предположения о целом числе остатков на один виток спирали. Переломный момент наступил ранней весной 1948 года, когда Полинг простудился и лег спать. Ему было скучно, и он нарисовал на полоске бумаги полипептидную цепь примерно правильных размеров и свернул ее в спираль, стараясь сохранить плоские пептидные связи. После нескольких попыток он создал модель с физически вероятными водородными связями. Затем Полинг работал с Кори и Брэнсоном, чтобы подтвердить свою модель перед публикацией.[6] В 1954 году Полинг был удостоен своей первой Нобелевской премии «за исследование природы химической связи и ее применение для выяснения структуры сложных веществ».[7] (например, белки), в том числе структуру α-спирали.
Структура
Геометрия и водородная связь
Аминокислоты в α-спирали расположены правосторонним образом. спиральный структура, где каждый аминокислотный остаток соответствует повороту спирали на 100 ° (т. е. спираль имеет 3,6 остатка на оборот) и переносу на 1,5 Å (0,15 нм) вдоль оси спирали. Дуниц[8] описывает, как первая статья Полинга по этой теме фактически показывает левую спираль, энантиомер истинной структуры. Иногда встречаются короткие отрезки левой спирали с большим содержанием ахирального глицин аминокислоты, но неблагоприятны для других нормальных, биологических L-аминокислоты. Шаг альфа-спирали (расстояние по вертикали между последовательными витками спирали) составляет 5,4 Å (0,54 нм), что является произведением 1,5 и 3,6. Самое главное, что N-H группа аминокислоты образует водородная связь с C = O группа аминокислоты четыре остатки ранее; это повторилось я + 4 → я водородная связь является наиболее заметной характеристикой α-спирали. Официальная международная номенклатура[9][10] определяет два способа определения α-спиралей, правило 6.2 с точки зрения повторения φ, ψ торсионные углы (см. ниже) и правило 6.3 с точки зрения комбинированного рисунка пека и водородных связей. Α-спирали можно идентифицировать в структуре белка с помощью нескольких вычислительных методов, одним из которых является DSSP (ОпределятьВторичная структура белка).[11]
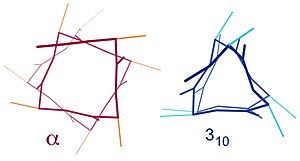
Подобные структуры включают 310 спираль (я + 3 → я водородная связь) и π-спираль (я + 5 → я водородная связь). Α-спираль можно описать как 3,613 спираль, поскольку я +4 интервал добавляет три атома к H-связанной петле по сравнению с более плотной 310 helix, и в среднем в одно кольцо α-спирали вовлечено 3,6 аминокислот. Нижние индексы относятся к числу атомов (включая водород) в замкнутом контуре, образованном водородной связью.[12]

Остатки в α-спиралях обычно принимают каркас (φ, ψ) двугранные углы около (-60 °, -45 °), как показано на изображении справа. В более общем плане они используют двугранные углы, так что ψ двугранный угол одного вычета и φ двугранный угол следующий сумма остатков составляет примерно -105 °. Как следствие, двугранные углы α-спирали, как правило, попадают в диагональную полосу на поверхности. Диаграмма Рамачандрана (с наклоном -1), в диапазоне от (-90 °, -15 °) до (-35 °, -70 °). Для сравнения, сумма двугранных углов для 310 спираль составляет примерно -75 °, тогда как для π-спирали примерно -130 °. Общая формула для угла поворота Ω на остаток любой полипептидной спирали с транс изомеры задаются уравнением[14][15]
- 3 cos Ω = 1 - 4 cos2 φ + ψ/2
Α-спираль плотно упакована; внутри спирали почти нет свободного места. Боковые цепи аминокислот находятся на внешней стороне спирали и направлены примерно «вниз» (то есть к N-концу), как ветви вечнозеленого дерева (Рождественская елка эффект). Эта направленность иногда используется в предварительных картах электронной плотности с низким разрешением для определения направления белкового остова.[16]
Стабильность
Спирали, наблюдаемые в белках, могут иметь длину от четырех до более чем сорока остатков, но типичная спираль содержит около десяти аминокислот (около трех витков). В общем, коротко полипептиды не проявляют значительной α-спиральной структуры в растворе, поскольку энтропийный Стоимость, связанная с укладкой полипептидной цепи, не компенсируется достаточным количеством стабилизирующих взаимодействий. В общем, позвоночник водородные связи α-спиралей считаются немного слабее, чем в β-листы, и легко атакуются окружающими молекулами воды. Однако в более гидрофобных средах, таких как плазматическая мембрана, или в присутствии сорастворителей, таких как трифторэтанол (ТФЭ) или выделенный из растворителя в газовой фазе,[17] олигопептиды легко принимают стабильную α-спиральную структуру. Кроме того, в пептиды могут быть включены поперечные сшивки для конформационной стабилизации спиральных складок. Сшивки стабилизируют спиральное состояние, энтропийно дестабилизируя развернутое состояние и удаляя энтальпийно стабилизированные «ложные» складки, которые конкурируют с полностью спиральным состоянием.[18] Было показано, что α-спирали более стабильны, устойчивы к мутациям и поддаются конструированию, чем β-цепи в природных белках,[19] а также в искусственно созданных белках.[20]

Экспериментальное определение
Поскольку α-спираль определяется ее водородными связями и конформацией основной цепи, наиболее подробные экспериментальные доказательства α-спиральной структуры получены с помощью атомного разрешения. Рентгеновская кристаллография как в примере, показанном справа. Ясно, что все карбонильные атомы кислорода основной цепи направлены вниз (к С-концу), но слегка расширяются, а Н-связи приблизительно параллельны оси спирали. Белковые структуры из ЯМР-спектроскопия также хорошо показывают спирали, с характерными наблюдениями ядерный эффект Оверхаузера (NOE) связи между атомами на соседних витках спирали. В некоторых случаях отдельные водородные связи можно наблюдать непосредственно как небольшую скалярную связь в ЯМР.
Есть несколько методов с более низким разрешением для задания общей спиральной структуры. В ЯМР химические сдвиги (в частности, Cα, Сβ и C ′) и остаточные диполярные связи часто характерны для спиралей. Дальний УФ (170–250 нм) круговой дихроизм Спектр спиралей также идиосинкразический, с ярко выраженным двойным минимумом в области 208 и 222 нм. Инфракрасный спектроскопия используется редко, так как α-спиральный спектр напоминает спектр случайный катушки (хотя они могут быть различимы, например, водород-дейтериевый обмен ). Наконец, крио электронная микроскопия теперь способен различать отдельные α-спирали в белке, хотя их отнесение к остаткам все еще является активной областью исследований.
Длинные гомополимеры аминокислот, если они растворимы, часто образуют спирали. Такие длинные изолированные спирали также можно обнаружить другими методами, такими как диэлектрическая релаксация, двойное лучепреломление потока, и измерения постоянная диффузии. Строго говоря, эти методы обнаруживают только характеристику вытянутый (длинная сигаровидная) гидродинамическая форма спирали или ее большая дипольный момент.
Аминокислотные склонности
Различные аминокислотные последовательности имеют разную склонность к образованию α-спиральной структуры. Метионин, аланин, лейцин, глутамат, и лизин незаряженный ("МАЛЕК" в аминокислота Однобуквенные коды) все имеют особенно высокую склонность к спиралевидному образованию, тогда как пролин и глицин имеют плохую склонность к образованию спиралей.[21] Пролин либо ломает, либо изгибает спираль, потому что он не может отдавать амид водородная связь (не имеющий амидного водорода), а также потому, что его боковая цепь стерически вмешивается в основу предыдущего поворота - внутри спирали это вызывает изгиб примерно на 30 ° оси спирали.[12] Однако пролин часто рассматривается как первый остаток спирали, предполагается из-за его структурной жесткости. С другой стороны, глицин также имеет тенденцию разрушать спирали, потому что его высокая конформационная гибкость делает энтропийно дорогостоящим принятие относительно ограниченной α-спиральной структуры.
Таблица предрасположенности стандартных аминокислот к альфа-спирали
Предполагаемые различия в свободная энергия, Δ (Δграмм), оцененная в ккал / моль на остаток в α-спиральной конфигурации относительно аланина, произвольно установленного равным нулю. Более высокие числа (более положительные свободные энергии) менее предпочтительны. Возможны значительные отклонения от этих средних чисел в зависимости от идентичности соседних остатков.
Различия в свободной энергии на остаток[22] Аминокислота 3-
письмо1-
письмоВинтовой штраф ккал / моль кДж / моль Аланин Ала А 0.00 0.00 Аргинин Arg р 0.21 0.88 Аспарагин Asn N 0.65 2.72 Аспарагиновая кислота Жерех D 0.69 2.89 Цистеин Cys C 0.68 2.85 Глютаминовая кислота Glu E 0.40 1.67 Глутамин Gln Q 0.39 1.63 Глицин Gly грамм 1.00 4.18 Гистидин Его ЧАС 0.61 2.55 Изолейцин Иль я 0.41 1.72 Лейцин Лея L 0.21 0.88 Лизин Lys K 0.26 1.09 Метионин Встретились M 0.24 1.00 Фенилаланин Phe F 0.54 2.26 Пролин Pro п 3.16 13.22 Серин Сер S 0.50 2.09 Треонин Thr Т 0.66 2.76 Триптофан Trp W 0.49 2.05 Тирозин Тюр Y 0.53 2.22 Валин Вал V 0.61 2.55
Дипольный момент
Спираль имеет общий дипольный момент за счет совокупного эффекта отдельных микродиполей от карбонил группы пептидной связи, указывающие вдоль оси спирали.[23] Эффекты этого макродиполя вызывают некоторые разногласия. α-спирали часто встречаются с N-концом, связанным с отрицательно заряженной группой, иногда аминокислота боковая цепь, такая как глутамат или же аспартат, или иногда фосфат-ион. Некоторые считают, что спиральный макродиполь электростатически взаимодействует с такими группами. Другие считают, что это вводит в заблуждение, и более реалистично сказать, что потенциал водородной связи свободных групп NH на N-конце α-спирали может быть удовлетворен за счет водородной связи; это также можно рассматривать как набор взаимодействий между локальными микродиполями, такими как С = O ··· H − N.[24][25]
Спиральные катушки
Спирали типа "спиральная спираль" представляют собой высокостабильные формы, в которых две или более спиралей наматываются друг на друга, образуя структуру "суперспираль". Спиральные катушки содержат очень характерный мотив последовательности известный как гептад повтор, в которой мотив повторяется через каждые семь остатков по последовательности (аминокислота остатки, а не пары оснований ДНК). Первый и особенно четвертый остатки (известные как а и d позиции) почти всегда гидрофобный; четвертый остаток обычно лейцин - отсюда и название структурный мотив называется лейциновая молния, который является разновидностью спиральной катушки. Эти гидрофобные остатки собираются вместе внутри спирального пучка. В общем, пятый и седьмой остатки ( е и грамм позиции) имеют противоположные заряды и образуют солевой мостик, стабилизированный электростатический взаимодействия. Волокнистые белки Такие как кератин или "стебли" миозин или же кинезин часто используют спиральные конструкции, как и несколько димеризация белки. Пара катушек - четырехконтурная.пучок спиралей - очень распространенный структурный мотив в белках. Например, это происходит у человека гормон роста и несколько разновидностей цитохром. В Роп протеин, который способствует репликации плазмиды в бактериях, представляет собой интересный случай, в котором один полипептид образует спиральную спираль, а два мономера собираются, чтобы сформировать пучок из четырех спиралей.
Аранжировки лица
Аминокислоты, составляющие определенную спираль, могут быть нанесены на косозубое колесо, представление, которое иллюстрирует ориентацию составляющих аминокислот (см. статью для лейциновая молния для такой диаграммы). Часто в глобулярные белки, а также в специализированных конструкциях, таких как бухты и лейциновые молнии, α-спираль будет иметь две «грани», одна из которых содержит преимущественно гидрофобный аминокислоты, ориентированные внутрь белка, в гидрофобное ядро, и один, содержащий преимущественно полярный аминокислоты, ориентированные на растворитель -открытая поверхность белка.
Изменения в ориентации связывания также происходят для олигопептидов с лицевой структурой. Этот паттерн особенно распространен в антимикробные пептиды, и было разработано множество моделей, чтобы описать, как это соотносится с их функцией. Общим для многих из них является то, что гидрофобная сторона антимикробного пептида образует поры в плазматической мембране после связывания с жировыми цепями в ядре мембраны.[26][27]
Сборки большего размера

Миоглобин и гемоглобин, первые два белка, структура которых была решена с помощью рентгеновского кристаллография, имеют очень похожие складки, состоящие примерно на 70% из α-спирали, а остальные представляют собой неповторяющиеся области или «петли», соединяющие спирали. При классификации белков по их доминирующей складке Структурная классификация белков база данных поддерживает большую категорию специально для белков всех альфа.
Тогда гемоглобин имеет еще больший масштаб. четвертичная структура, в котором функциональная кислородсвязывающая молекула состоит из четырех субъединиц.
Функциональные роли


Связывание ДНК
α-Спирали имеют особое значение в ДНК связывающие мотивы, в том числе спираль-поворот-спираль мотивы лейциновая молния мотивы и цинковый палец мотивы. Это связано с удобным структурным фактом, что диаметр α-спирали составляет около 12 Å (1,2 нм), включая средний набор боковых цепей, примерно такой же, как ширина большой канавки в B-форме. ДНК, а также потому, что спиральная катушка (или лейциновая застежка-молния) димеры спиралей могут легко позиционировать пару взаимодействующих поверхностей, чтобы контактировать с симметричным повтором, обычным для двойной спирали ДНК.[28] Примером обоих аспектов является фактор транскрипции Макс (см. Изображение слева), который использует спиральную спиральную катушку для димеризации, помещая другую пару спиралей для взаимодействия в двух последовательных витках большой бороздки ДНК.
Мембранный охват
α-Спирали также являются наиболее распространенным элементом структуры белка, проникающим через биологические мембраны (трансмембранный белок ),[29] это предполагается, потому что спиральная структура может удовлетворять всем водородным связям основной цепи внутри, не оставляя полярных групп, открытых для мембраны, если боковые цепи гидрофобны. Белки иногда закрепляются одной спиралью, проходящей через мембрану, иногда парой, а иногда и пучком спиралей, наиболее классически состоящим из семи спиралей, расположенных вверх и вниз в кольцо, например, для родопсины (см. изображение справа) или для G-белковые рецепторы (GPCR).
Механические свойства
α-Спирали при осевой деформации растяжения, характерное условие нагружения, которое проявляется во многих богатых альфа-спиралями нитях и тканях, приводит к характерному трехфазному поведению касательного модуля жесткость-мягкость-жесткость.[30] Фаза I соответствует режиму малых деформаций, во время которого спираль однородно растягивается, за ней следует фаза II, в которой альфа-спиральные витки разрываются за счет разрыва групп Н-связей. Фаза III обычно связана с растяжением ковалентной связи с большой деформацией.
Динамические особенности
Альфа-спирали в белках могут иметь Низкая частота аккордеонное движение, наблюдаемое Рамановская спектроскопия[31] и проанализированы с помощью квазиконтинуальной модели.[32][33] Спирали, не стабилизированные третичными взаимодействиями, демонстрируют динамическое поведение, которое в основном можно отнести к истиранию спиралей с концов.[34]
Переход спираль – катушка
Гомополимеры аминокислот (например, полилизин ) может принимать α-спиральную структуру при низкой температуре, которая «плавится» при высоких температурах. Этот переход спираль – спираль когда-то считалось аналогом белка денатурация. В статистическая механика этого перехода можно смоделировать с помощью элегантного матрица передачи метод, характеризующийся двумя параметрами: склонностью к возникновению спирали и склонностью к расширению спирали.
В искусстве

По крайней мере пять художников явно ссылались на α-спираль в своих работах: Джули Ньюдолл в живописи и Джулиан Восс-Андреэ, Вирсавия Гроссман, Байрон Рубин и Майк Тайка в скульптуре.
Художница из Сан-Франциско Джули Ньюдолл,[35] Имеет степень в области микробиологии и немного искусствовед. С 1990 года специализируется на картинах, вдохновленных микроскопическими изображениями и молекулами. На ее картине «Возвышение альфа-спирали» (2003) человеческие фигуры расположены в виде спирали α. По словам художника, «цветы отражают различные типы боковых цепей, которые каждая аминокислота передает миру».[35] Эта же метафора также повторяется со стороны ученого: «β-листы не показывают жесткой повторяющейся регулярности, но текут в изящных извилистых кривых, и даже α-спираль имеет правильную форму, больше похожую на стебель цветка, узлы ветвления которого показывают влияние окружающей среды, истории развития и эволюции каждой части в соответствии с ее собственной идиосинкразической функцией ».[12]
Джулиан Восс-Андреэ Скульптор немецкого происхождения, имеет ученую степень в области экспериментальной физики и скульптуры. С 2001 года Фосс-Андреа создает «белковые скульптуры».[36] основанный на структуре белка с α-спиралью, являющейся одним из его любимых объектов. Фосс-Андреа создал скульптуры α-спирали из различных материалов, включая бамбук и целые деревья. Памятник Voss-Andreae, созданный в 2004 году, чтобы отметить память Линус Полинг, открыватель α-спирали, образован из большой стальной балки, преобразованной в структуру α-спирали. Ярко-красная скульптура 10 футов высотой (3 м) стоит перед домом, где прошло детство Полинга в Портланд, штат Орегон.
Ленточные диаграммы α-спиралей - важный элемент в лазерных скульптурах белковых структур, созданных художником. Вирсавия Гроссман, например, из инсулин, гемоглобин, и ДНК-полимераза.[37] Байрон Рубин - бывший белковый кристаллограф, а теперь профессиональный скульптор в области металлов, белков, нуклеиновых кислот и молекул лекарств, многие из которых имеют α-спирали, такие как субтилизин, гормон роста человека, и фосфолипаза А2.[38]
Майк Тайка - вычислительный биохимик в Вашингтонский университет работаю с Дэвид Бейкер. Tyka с 2010 года делает скульптуры из белковых молекул из меди и стали, в том числе убиквитин и калиевый канал тетрамер.[39]
Смотрите также
- 310 спираль
- Бета-лист
- Давыдовский солитон
- Складывание (химия)
- Ручки в отверстия набивка
- Пи спираль
- Протеопедия Helices_in_Proteins
Рекомендации
- ^ Кендрю JC, Дикерсон Р. Э., Страндберг Б. Е., Харт Р. Г., Дэвис Д. Р., Филипс, округ Колумбия, Шор В. К. (февраль 1960 г.). «Структура миоглобина: трехмерный синтез Фурье с разрешением 2 Å». Природа. 185 (4711): 422–7. Bibcode:1960Натура.185..422K. Дои:10.1038 / 185422a0. PMID 18990802. S2CID 4167651.
- ^ Neurath H (1940). «Внутримолекулярное сворачивание полипептидных цепей в зависимости от структуры белка». Журнал физической химии. 44 (3): 296–305. Дои:10.1021 / j150399a003.
- ^ Тейлор Х.С. (1942). «Крупные молекулы через атомные очки». Труды Американского философского общества. 85 (1): 1–12. JSTOR 985121.
- ^ Хаггинс М (1943). «Строение волокнистых белков». Химические обзоры. 32 (2): 195–218. Дои:10.1021 / cr60102a002.
- ^ Брэгг В.Л., Кендрю JC, Perutz MF (1950). «Конфигурации полипептидных цепей в кристаллических белках». Труды Лондонского королевского общества. Серия А. Математические и физические науки.. 203 (1074): 321–?. Bibcode:1950RSPSA.203..321B. Дои:10.1098 / rspa.1950.0142. S2CID 93804323.
- ^ Полинг Л, Кори РБ, Брэнсон HR (Апрель 1951 г.). «Структура белков; две спиральные конфигурации полипептидной цепи с водородными связями». Труды Национальной академии наук Соединенных Штатов Америки. 37 (4): 205–11. Bibcode:1951ПНАС ... 37..205П. Дои:10.1073 / pnas.37.4.205. ЧВК 1063337. PMID 14816373.
- ^ "Нобелевская премия по химии 1954 г.".
- ^ Дуниц Дж. (2001). "Левая α-спираль Полинга". Angewandte Chemie International Edition. 40 (22): 4167–4173. Дои:10.1002 / 1521-3773 (20011119) 40:22 <4167 :: AID-ANIE4167> 3.0.CO; 2-Q. PMID 29712120.
- ^ Комиссия IUPAC-IUB по биохимической номенклатуре (1970). «Аббревиатуры и символы для описания конформации полипептидных цепей». Журнал биологической химии. 245: 6489–6497.
- ^ «Конформации полипептида 1 и 2». www.sbcs.qmul.ac.uk. Получено 5 ноября 2018.
- ^ Кабш В., Сандер С. (декабрь 1983 г.). «Словарь вторичной структуры белков: распознавание образов водородных связей и геометрических элементов». Биополимеры. 22 (12): 2577–637. Дои:10.1002 / bip.360221211. PMID 6667333.
- ^ а б c Ричардсон Дж. С. (1981). «Анатомия и таксономия структуры белка». Достижения в химии белков. 34: 167–339. Дои:10.1016 / S0065-3233 (08) 60520-3. ISBN 9780120342341. PMID 7020376.
- ^ Ловелл С.К., Дэвис И.В., Арендалл В.Б., де Баккер П.И., Уорд Дж.М., Присант М.Г., Ричардсон Дж.С., Ричардсон, округ Колумбия (февраль 2003 г.). «Подтверждение структуры по геометрии Calpha: отклонения фи, пси и Сбета». Белки. 50 (3): 437–50. Дои:10.1002 / prot.10286. PMID 12557186.
- ^ Дикерсон RE, Geis I (1969), Структура и действие белков, Харпер, Нью-Йорк
- ^ Зорко, Матяж (2010). «Структурная организация белков». В Лангеле, Юло; Cravatt, Бенджамин Ф.; Грэслунд, Астрид; фон Хейне, Гуннар; Земля, Тиит; Ниссен, Шерри; Зорко, Матяж (ред.). Введение в пептиды и белки. Бока-Ратон: CRC Press. С. 36–57. ISBN 9781439882047.
- ^ Terwilliger TC (март 2010 г.). «Быстрое построение модели альфа-спиралей на картах электронной плотности». Acta Crystallographica Раздел D. 66 (Pt 3): 268–75. Дои:10.1107 / S0907444910000314. ЧВК 2827347. PMID 20179338.
- ^ Хаджинс Р.Р., Джарролд М.Ф. (1999). «Формирование спирали в несольватированных пептидах на основе аланина: спиральные мономеры и спиральные димеры». Журнал Американского химического общества. 121 (14): 3494–3501. Дои:10.1021 / ja983996a.
- ^ Кучукян П.С., Ян Я.С., Вердин Г.Л., Шахнович Е.И. (апрель 2009 г.). «Всеатомная модель стабилизации альфа-спиральной структуры пептидов углеводородными скобками». Журнал Американского химического общества. 131 (13): 4622–7. Дои:10.1021 / ja805037p. ЧВК 2735086. PMID 19334772.
- ^ Абрусан Г., Марш Дж. А. (2016). «Альфа-спирали более устойчивы к мутациям, чем бета-цепи». PLOS вычислительная биология. 12 (12): e1005242. Bibcode:2016PLSCB..12E5242A. Дои:10.1371 / journal.pcbi.1005242. ЧВК 5147804. PMID 27935949.
- ^ Rocklin GJ, et al. (2017). «Глобальный анализ сворачивания белков с использованием массового параллельного проектирования, синтеза и тестирования». Наука. 357 (6347): 168–175. Bibcode:2017Научный ... 357..168R. Дои:10.1126 / science.aan0693. ЧВК 5568797. PMID 28706065.
- ^ Pace CN, Scholtz JM (июль 1998 г.). «Шкала склонности к спирали, основанная на экспериментальных исследованиях пептидов и белков». Биофизический журнал. 75 (1): 422–7. Bibcode:1998BpJ .... 75..422N. Дои:10.1016 / S0006-3495 (98) 77529-0. ЧВК 1299714. PMID 9649402.
- ^ Пейс, К. Ник; Шольц, Дж. Мартин (1998). «Шкала склонности к спирали, основанная на экспериментальных исследованиях пептидов и белков». Биофизический журнал. 75. С. 422–427. Bibcode:1998BpJ .... 75..422N. Дои:10.1016 / с0006-3495 (98) 77529-0.
- ^ Хол WG, ван Дуйнен PT, Берендсен HJ (1978). «Диполь альфа-спирали и свойства белков». Природа. 273 (5662): 443–446. Bibcode:1978Натура.273..443H. Дои:10.1038 / 273443a0. PMID 661956. S2CID 4147335.
- ^ He JJ, Quiocho FA (октябрь 1993 г.). «Доминирующая роль локальных диполей в стабилизации нескомпенсированных зарядов на сульфате, изолированном в периплазматическом активном транспортном белке». Белковая наука. 2 (10): 1643–7. Дои:10.1002 / pro.5560021010. ЧВК 2142251. PMID 8251939.
- ^ Милнер-Уайт EJ (ноябрь 1997 г.). «Частичный заряд атома азота в пептидных связях». Белковая наука. 6 (11): 2477–82. Дои:10.1002 / pro.5560061125. ЧВК 2143592. PMID 9385654.
- ^ Кон, Эрик М .; Ширли, Дэвид Дж .; Ароцкий, Любовь; Пиччиано, Анджела М .; Риджуэй, Захари; Урбан, Майкл В .; Кароне, Бенджамин Р .; Капуто, Грегори А. (04.02.2018). «Роль катионных боковых цепей в антимикробной активности C18G». Молекулы. 23 (2): 329. Дои:10.3390 / молекулы23020329. ЧВК 6017431. PMID 29401708.
- ^ Токэ, Орсоля (2005). «Антимикробные пептиды: новые кандидаты в борьбе с бактериальными инфекциями». Биополимеры. 80 (6): 717–735. Дои:10.1002 / bip.20286. ISSN 0006-3525. PMID 15880793.
- ^ Бранден и Туз, глава 10
- ^ Бранден и Туз, глава 12.
- ^ Акбаров Т., Чен Х, Кетен С., Бюлер М.Дж. (октябрь 2007 г.). «Иерархия, множественные энергетические барьеры и надежность определяют механику разрушения альфа-спиральных и бета-листовых белковых доменов». Труды Национальной академии наук Соединенных Штатов Америки. 104 (42): 16410–5. Bibcode:2007ПНАС..10416410А. Дои:10.1073 / pnas.0705759104. ЧВК 2034213. PMID 17925444.
- ^ Painter PC, Mosher LE, Rhoads C (июль 1982 г.). «Низкочастотные моды в рамановских спектрах белков». Биополимеры. 21 (7): 1469–72. Дои:10.1002 / bip.360210715. PMID 7115900.
- ^ Chou KC (декабрь 1983 г.). «Выявление низкочастотных мод в белковых молекулах». Биохимический журнал. 215 (3): 465–9. Дои:10.1042 / bj2150465. ЧВК 1152424. PMID 6362659.
- ^ Chou KC (май 1984 г.). «Биологические функции низкочастотных колебаний (фононов). III. Спиральные структуры и микросреда». Биофизический журнал. 45 (5): 881–9. Bibcode:1984BpJ .... 45..881C. Дои:10.1016 / S0006-3495 (84) 84234-4. ЧВК 1434967. PMID 6428481.
- ^ Фирц Б., Райнер А., Кифхабер Т. (январь 2009 г.). «Локальная конформационная динамика в альфа-спиралях, измеренная с помощью быстрого триплетного переноса». Труды Национальной академии наук Соединенных Штатов Америки. 106 (4): 1057–62. Bibcode:2009PNAS..106.1057F. Дои:10.1073 / pnas.0808581106. ЧВК 2633579. PMID 19131517.
- ^ а б "Джули Ньюдолл, вдохновленное наукой искусство, музыка, настольные игры". www.brushwithscience.com. Получено 2016-04-06.
- ^ Восс-Андреэ Дж (2005). "Белковые скульптуры: строительные блоки жизни вдохновляют искусство". Леонардо. 38: 41–45. Дои:10.1162 / leon.2005.38.1.41. S2CID 57558522.
- ^ Гроссман, Вирсавия. «О художнике». Скульптура Вирсавии. Получено 2016-04-06.
- ^ "О". молекулярная скульптура. com. Получено 2016-04-06.
- ^ Тыка, Майк. "О". www.miketyka.com. Получено 2016-04-06.
дальнейшее чтение
- Туз Дж., Брандин С. (1999). Введение в структуру белка. Нью-Йорк: Garland Pub. ISBN 0-8153-2304-2..
- Айзенберг Д. (сентябрь 2003 г.). «Открытие альфа-спирали и бета-листа, основных структурных особенностей белков». Труды Национальной академии наук Соединенных Штатов Америки. 100 (20): 11207–10. Bibcode:2003PNAS..10011207E. Дои:10.1073 / pnas.2034522100. ЧВК 208735. PMID 12966187.
- Эстбери В.Т., Вудс HJ (1931). «Молекулярные массы белков». Природа. 127 (3209): 663–665. Bibcode:1931Натура.127..663А. Дои:10.1038 / 127663b0. S2CID 4133226.
- Astbury WT, улица A (1931 г.). «Рентгеновские исследования структуры волос, шерсти и родственных волокон. I. Общие сведения». Пер. R. Soc. Лондон. A230: 75–101. Bibcode:1932РСПТА.230 ... 75А. Дои:10.1098 / рста.1932.0003.
- Эстбери В. Т. (1933). «Некоторые проблемы рентгенологического анализа структуры шерсти животных и других белковых волокон». Пер. Фарадей Соц. 29 (140): 193–211. Дои:10.1039 / tf9332900193.
- Эстбери В.Т., Вудс HJ (1934). «Рентгеновские исследования структуры волос, шерсти и родственных волокон. II. Молекулярная структура и эластические свойства кератина волос». Философские труды Лондонского королевского общества, серия A. 232 (707–720): 333–394. Bibcode:1934RSPTA.232..333A. Дои:10.1098 / рста.1934.0010.
- Эстбери В. Т., Сиссон В. А. (1935). «Рентгеновские исследования структуры волос, шерсти и родственных волокон. III. Конфигурация молекулы кератина и ее ориентация в биологической клетке». Труды Королевского общества. A150 (871): 533–551. Bibcode:1935RSPSA.150..533A. Дои:10.1098 / rspa.1935.0121.
- Сугета Х., Миядзава Т (1967). «Общий метод расчета спиральных параметров полимерных цепей на основе длин связки, углов связи и углов внутреннего вращения». Биополимеры. 5 (7): 673–679. Дои:10.1002 / bip.1967.360050708.
- Вада А (1976). «Альфа-спираль как электрический макродиполь». Достижения в биофизике: 1–63. PMID 797240.
- Чотия К., Левитт М., Ричардсон Д. (октябрь 1977 г.). «Структура белков: упаковка альфа-спиралей и гофрированные листы». Труды Национальной академии наук Соединенных Штатов Америки. 74 (10): 4130–4. Bibcode:1977PNAS ... 74.4130C. Дои:10.1073 / пнас.74.10.4130. ЧВК 431889. PMID 270659.
- Чотия К., Левитт М., Ричардсон Д. (январь 1981 г.). «Упаковка от спирали к спирали в белках». Журнал молекулярной биологии. 145 (1): 215–50. Дои:10.1016/0022-2836(81)90341-7. PMID 7265198.
- Хол WG (1985). «Роль диполя альфа-спирали в функции и структуре белка». Прогресс в биофизике и молекулярной биологии. 45 (3): 149–95. Дои:10.1016 / 0079-6107 (85) 90001-X. PMID 3892583.
- Барлоу DJ, Торнтон Дж. М. (июнь 1988 г.). «Геометрия спирали в белках». Журнал молекулярной биологии. 201 (3): 601–19. Дои:10.1016/0022-2836(88)90641-9. PMID 3418712.
- Мурзин А.Г., Финкельштейн А.В. (декабрь 1988 г.). «Общая архитектура альфа-спиральной глобулы». Журнал молекулярной биологии. 204 (3): 749–69. Дои:10.1016 / 0022-2836 (88) 90366-Х. PMID 3225849.
внешняя ссылка
- NetSurfP вер. 1.1 - Доступность поверхности белка и прогноз вторичной структуры
- Калькулятор угла поворота α-спирали
- Сайт художника Джули Ньюдолл
- Сайт художника Джулиана Вос-Андреэ
| Кривые |  | ||
|---|---|---|---|
| Спирали |
| ||
| Спирали | |||