Пролин - Proline
 | |||
| |||
| Имена | |||
|---|---|---|---|
| Название ИЮПАК Пролин | |||
| Систематическое название ИЮПАК Пирролидин-2-карбоновая кислота[1] | |||
| Идентификаторы | |||
3D модель (JSmol ) | |||
| 80812 | |||
| ЧЭБИ | |||
| ЧЭМБЛ | |||
| ChemSpider | |||
| DrugBank | |||
| ECHA InfoCard | 100.009.264 | ||
| Номер ЕС |
| ||
| 26927 | |||
| КЕГГ | |||
| MeSH | Пролин | ||
PubChem CID | |||
| Номер RTECS |
| ||
| UNII | |||
| |||
| |||
| Характеристики | |||
| C5ЧАС9NО2 | |||
| Молярная масса | 115.132 г · моль−1 | ||
| Внешность | Прозрачные кристаллы | ||
| Температура плавления | От 205 до 228 ° C (от 401 до 442 ° F, от 478 до 501 K) (разлагается) | ||
| Растворимость | 1,5 г / 100 г этанола 19 ° C[2] | ||
| бревно п | -0.06 | ||
| Кислотность (пKа) | 1,99 (карбоксил), 10,96 (амино)[3] | ||
| Опасности | |||
| Паспорт безопасности | Видеть: страница данных | ||
| S-фразы (устарело) | S22, S24 / 25 | ||
| Страница дополнительных данных | |||
| Показатель преломления (п), Диэлектрическая постоянная (εр), так далее. | |||
Термодинамический данные | Фазовое поведение твердое тело – жидкость – газ | ||
| УФ, ИК, ЯМР, РС | |||
Если не указано иное, данные для материалов приводятся в их стандартное состояние (при 25 ° C [77 ° F], 100 кПа). | |||
| Ссылки на инфобоксы | |||

Пролин (символ Pro или же п)[4] органическая кислота классифицируется как протеиногенная аминокислота (используется в биосинтез белков ), хотя он не содержит аминогруппа -NH
2 но скорее вторичный амин. Азот вторичного амина находится в протонированном NH2+ образуются в биологических условиях, а карбоксильная группа находится в депротонированный −COO− форма. "Боковая цепь" от α углерод соединяется с азотом, образуя пирролидин цикл, классифицируя его как алифатический аминокислота. Он не является незаменимым для человека, то есть организм может синтезировать его из незаменимой аминокислоты. L-глутамат. это закодированный всеми кодоны начиная с CC (CCU, CCC, CCA и CCG).
Пролин - единственная протеиногенная аминокислота, которая является вторичным амином, поскольку атом азота присоединен к α-углероду цепочкой из трех других атомов углерода.
История и этимология
Пролин был впервые выделен в 1900 г. Ричард Вильштеттер который получил аминокислоту при изучении N-метилпролина. Год спустя Эмиль Фишер опубликовали синтез пролина из эфира пропилмалоновой кислоты фталимида.[5] Название пролин происходит от пирролидин, один из его компонентов.[6]
Биосинтез
Пролин биосинтетически полученный из аминокислоты L-глутамат. Глутамат-5-полуальдегид сначала сформирован глутамат-5-киназа (АТФ-зависимый) и глутамат-5-полуальдегиддегидрогеназа (который требует НАДН или НАДФН). Затем он может либо спонтанно циклизоваться с образованием 1-пирролин-5-карбоновая кислота, который восстанавливается до пролина пирролин-5-карбоксилатредуктаза (используя НАДН или НАДФН), или превратился в орнитин к орнитинаминотрансфераза с последующей циклизацией орнитинциклодезаминаза с образованием пролина.[7]

Биологическая активность
L-Пролин действует как слабый агонист из рецептор глицина и обоих NMDA и не-NMDA (AMPA /каинат ) ионотропные рецепторы глутамата.[8][9][10] Было предложено стать потенциальным эндогенный эксайтотоксин.[8][9][10] В растения, накопление пролина является обычной физиологической реакцией на различные стрессы, но также является частью программы развития в генеративные ткани (например. пыльца ).[11]
Свойства в структуре белка
Отличительная циклическая структура боковой цепи пролина придает пролину исключительную конформационную жесткость по сравнению с другими аминокислотами. Это также влияет на скорость образования пептидной связи между пролином и другими аминокислотами. Когда пролин связан как амид в пептидной связи, его азот не связан с водородом, а это означает, что он не может действовать как водородная связь донор, но может быть акцептором водородной связи.
Образование пептидной связи с входящей про-тРНКPro значительно медленнее, чем с любыми другими тРНК, что является общей особенностью N-алкиламинокислот.[12] Образование пептидной связи между входящей тРНК и цепью, оканчивающейся на пролине, также происходит медленно; медленнее всего с образованием пролин-пролиновых связей.[13]
Исключительная конформационная жесткость пролина влияет на вторичная структура белков около остатка пролина и может объяснять более высокую распространенность пролина в белках теплолюбивый организмы. Вторичная структура белка можно описать с точки зрения двугранные углы φ, ψ и ω белкового остова. Циклическая структура боковой цепи пролина фиксирует угол φ примерно на -65 °.[14]
Пролин действует как структурный разрушитель в середине регулярного вторичная структура такие элементы, как альфа спирали и бета-листы; однако пролин обычно обнаруживается как первый остаток альфа спираль а также в краевых прядях бета-листы. Пролин также часто встречается в повороты (другой вид вторичной структуры) и помогает в образовании бета-поворотов. Это может объяснить любопытный факт, что пролин обычно подвергается воздействию растворителей, несмотря на то, что он полностью алифатический боковая цепь.
Множественные пролины и / или гидроксипролины подряд может создать полипролиновая спираль, преобладающая вторичная структура в коллаген. В гидроксилирование пролина пролилгидроксилаза (или другие добавки электроноакцепторных заместителей, таких как фтор ) увеличивает конформационную стабильность коллаген существенно.[15] Следовательно, гидроксилирование пролина является критическим биохимическим процессом для поддержания соединительная ткань высших организмов. Тяжелые заболевания, такие как цинга может быть результатом дефектов гидроксилирования, например, мутаций фермента пролилгидроксилазы или отсутствия необходимых аскорбат (витамин С) кофактор.
Цис-транс-изомеризация
Пептидные связи к пролину и другим N-замещенные аминокислоты (такие как саркозин ), могут заполнять как СНГ и транс изомеры. Большинство пептидных связей в подавляющем большинстве принимают транс изомер (обычно 99,9% в недеформированных условиях), главным образом потому, что амидный водород (транс изомер) предлагает меньшее стерическое отталкивание по сравнению с предыдущим Cα атом, чем следующий Cα атом (СНГ изомер). Напротив, СНГ и транс изомеры пептидной связи X-Pro (где X представляет собой любую аминокислоту) испытывают стерические конфликты с соседним замещением и имеют гораздо меньшую разницу в энергии. Следовательно, доля пептидных связей X-Pro в СНГ изомер в недеформированных условиях значительно повышен, с СНГ фракции обычно находятся в пределах 3-10%.[16] Однако эти значения зависят от предыдущей аминокислоты, при этом Gly[17] и ароматный[18] остатки, дающие повышенную долю СНГ изомер. СНГ фракции до 40% были идентифицированы для пептидных связей Aromatic-Pro.[19]
С кинетической точки зрения, СНГ-транс пролин изомеризация это очень медленный процесс, который может помешать развитию сворачивание белка путем улавливания одного или нескольких остатков пролина, важных для укладки в ненативный изомер, особенно когда для нативного белка требуется СНГ изомер. Это связано с тем, что остатки пролина синтезируются исключительно в рибосома как транс изомерная форма. Все организмы обладают пролилизомераза ферменты чтобы катализировать эту изомеризацию, а некоторые бактерии имеют специализированные пролилизомеразы, связанные с рибосомой. Однако не все пролины необходимы для сворачивания, и сворачивание белка может происходить с нормальной скоростью, несмотря на наличие ненативных конформеров многих пептидных связей X-Pro.
Использует
Пролин и его производные часто используются в качестве асимметричных катализаторов в органокатализ пролина реакции. В Сокращение CBS и пролин катализированный альдольная конденсация являются яркими примерами.
При пивоварении белки, богатые пролином, соединяются с полифенолами, вызывая помутнение (помутнение).[20]
L-Пролайн - это осмозащитное средство и поэтому используется во многих фармацевтических и биотехнологических приложениях.
В среда роста используется в культура ткани растений можно дополнить пролином. Это может увеличить рост, возможно, потому, что помогает растениям переносить стрессы, связанные с культурой тканей.[21][нужен лучший источник ] О роли пролина в стрессовой реакции растений см. § Биологическая активность.
Специальности
Пролин - одна из двух аминокислот, которые не соответствуют типичным Рамачандран сюжет, вместе с глицин. Из-за образования кольца, связанного с бета-углеродом, углы ψ и φ вокруг пептидной связи имеют меньше допустимых градусов вращения. В результате он часто встречается в «поворотах» белков, поскольку его свободная энтропия (ΔS) не так велика по сравнению с другими аминокислотами, и, таким образом, в свернутой форме по сравнению с развернутой формой изменение энтропии меньше. Кроме того, пролин редко встречается в α- и β-структурах, так как он снижает стабильность таких структур, поскольку α-N его боковой цепи может образовывать только одну азотную связь.
Кроме того, пролин - единственная аминокислота, которая не окрашивается в красный / пурпурный цвет при проявлении путем опрыскивания нингидрин для использования в хроматография. Пролин, напротив, дает оранжево-желтый цвет.
История
Ричард Вильштеттер синтезировал пролин по реакции натриевой соли диэтилмалонат с 1,3-дибромпропан в 1900 г. В 1901 г. Герман Эмиль Фишер изолированный пролин из казеина и продуктов разложения γ-фталимидо-пропилмалонового эфира.[22]
Синтез
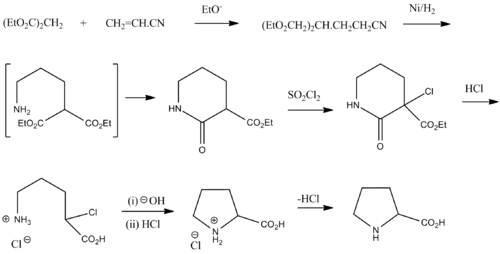
Рацемический пролин может быть синтезирован из диэтилмалонат и акрилонитрил:[23]
Эволюция
Несколько независимых эволюционных исследований с использованием различных типов данных показали, что пролин принадлежит к группе аминокислот, составляющих ранний генетический код.[24][25][26][27] Например, регионы низкой сложности (в белках), которые могут напоминать протопептиды ранних генетический код сильно обогащены пролином.[27]
Смотрите также
Рекомендации
- ^ Pubchem. «Пролайн». pubchem.ncbi.nlm.nih.gov. В архиве из оригинала 16 января 2014 г.. Получено 8 мая 2018.
- ^ Х.-Д. Белиц; В. Грош; П. Шиберле (15 января 2009 г.). Пищевая химия. п. 15. ISBN 978-3-540-69933-0. В архиве из оригинала от 15.05.2016.
- ^ Нельсон Д.Л., Кокс М.М. Принципы биохимии. NY: W.H. Фримен и компания.
- ^ «Номенклатура и символика аминокислот и пептидов». Совместная комиссия IUPAC-IUB по биохимической номенклатуре. 1983. Архивировано с оригинал 9 октября 2008 г.. Получено 5 марта 2018.
- ^ «Пролайн». В архиве из оригинала 27.11.2015.
- ^ "пролин". Словарь английского языка American Heritage, 4-е издание. В архиве из оригинала от 15.09.2015. Получено 2015-12-06.
- ^ Lehninger, Albert L .; Нельсон, Дэвид Л .; Кокс, Майкл М. (2000). Принципы биохимии (3-е изд.). Нью-Йорк: У. Х. Фриман. ISBN 1-57259-153-6..
- ^ а б Справочник по ионным каналам: каналы, управляемые внеклеточными лигандами. Академическая пресса. 16 ноября 1995. С. 126–. ISBN 978-0-08-053519-7. В архиве из оригинала от 26 апреля 2016 г.
- ^ а б Хенци В., Райхлинг Д. Б., Хелм С. В., МакДермотт А. Б. (1992). «L-пролин активирует рецепторы глутамата и глицина в культивируемых нейронах дорсального рога крысы». Мол. Pharmacol. 41 (4): 793–801. PMID 1349155.
- ^ а б Орхан Э. Арслан (7 августа 2014 г.). Нейроанатомические основы клинической неврологии, второе издание. CRC Press. С. 309–. ISBN 978-1-4398-4833-3. В архиве из оригинала 14 мая 2016 г.
- ^ Verbruggen N, Hermans C (2008). «Накопление пролина в растениях: обзор» (PDF). Аминокислоты. 35 (4): 753–759. Дои:10.1007 / s00726-008-0061-6. PMID 18379856. S2CID 21788988.
- ^ Павлов Михаил Юрьевич; Уоттс, Ричард Э; Тан, Чжунпин; Корниш, штат Вирджиния (W); Эренберг, Монс; Форстер, Энтони С. (2010), «Медленное образование пептидной связи пролином и другими N-алкиламинокислотами при трансляции», PNAS, 106 (1): 50–54, Дои:10.1073 / pnas.0809211106, ЧВК 2629218, PMID 19104062.
- ^ Buskirk, Allen R .; Грин, Рэйчел (2013). «Преодоление полипролиновых пауз». Наука. 339 (6115): 38–39. Bibcode:2013Наука ... 339 ... 38Б. Дои:10.1126 / наука.1233338. ЧВК 3955122. PMID 23288527.
- ^ Моррис, Энн (1992). «Стереохимическое качество координат структуры белка». Белки: структура, функции и биоинформатика. 12 (4): 345–364. Дои:10.1002 / prot.340120407. PMID 1579569. S2CID 940786.
- ^ Шпак, Пол (2011). «Химия и ультраструктура костей рыб: значение для тафономии и анализа стабильных изотопов». Журнал археологической науки. 38 (12): 3358–3372. Дои:10.1016 / j.jas.2011.07.022. В архиве из оригинала от 18.01.2012.
- ^ Alderson, T.R .; Lee, J.H .; Charlier, C .; Ин, Дж. И Бакс, А. (2017). «Склонность к образованию цис-пролина в развернутых белках». ChemBioChem. 19 (1): 37–42. Дои:10.1002 / cbic.201700548. ЧВК 5977977. PMID 29064600.
- ^ Саркар, С.К .; Young, P.E .; Салливан С.Э. и Торчиа Д.А. (1984). «Обнаружение цис- и транс-пептидных связей X-Pro в белках с помощью 13C ЯМР: приложение к коллагену». Труды Национальной академии наук США. 81 (15): 4800–4803. Bibcode:1984PNAS ... 81.4800S. Дои:10.1073 / пнас.81.15.4800. ЧВК 391578. PMID 6589627.
- ^ Thomas, K.M .; Надутхамби, Д. и Зондло, Нью-Джерси (2006). «Электронный контроль цис-транс-изомерии амида через взаимодействие ароматических пролилов». Журнал Американского химического общества. 128 (7): 2216–2217. Дои:10.1021 / ja057901y. PMID 16478167.
- ^ Gustafson, C.L .; Parsely, N.C .; Asimgil, H .; и другие. (2017). «Медленный конформационный переключатель в домене трансактивации BMAL1 модулирует циркадные ритмы». Молекулярная клетка. 66 (4): 447–457.e7. Дои:10.1016 / j.molcel.2017.04.011. ЧВК 5484534. PMID 28506462.
- ^ К.Дж. Зиберт, «Дымка и пена»,«Архивная копия». В архиве из оригинала от 11.07.2010. Получено 2010-07-13.CS1 maint: заархивированная копия как заголовок (связь) Доступ 12 июля 2010 г.
- ^ Пазуки, А; Асгари, Дж; Сохани, М; Пессаракли, М и Афлаки, Ф (2015). «Влияние некоторых источников органического азота и антибиотиков на рост каллуса сортов риса Indica». Журнал питания растений. 38 (8): 1231–1240. Дои:10.1080/01904167.2014.983118. S2CID 84495391.
- ^ R.H.A. Плиммер (1912) [1908], R.H.A. Плиммер и Ф. Хопкинс (ред.), Химический состав белков, Монографии по биохимии, Часть I. Анализ (2-е изд.), Лондон: Longmans, Green and Co., стр. 130, получено 20 сентября, 2010
- ^ Фогель, Практическая органическая химия 5-е издание
- ^ Трифонов, Э.Н. (декабрь 2000 г.). «Согласованный временной порядок аминокислот и эволюция триплетного кода». Ген. 261 (1): 139–151. Дои:10.1016 / S0378-1119 (00) 00476-5.
- ^ Хиггс, Пол Дж .; Пудриц, Ральф Э. (июнь 2009 г.). "Термодинамическая основа синтеза пребиотических аминокислот и природа первого генетического кода". Астробиология. 9 (5): 483–490. Дои:10.1089 / аст.2008.0280. ISSN 1531-1074.
- ^ Халиотис, Анаргирос; Властаридис, Панайотис; Мосиалос, Димитрис; Ибба, Майкл; Becker, Hubert D .; Статопулос, Константинос; Амуциас, Григориос Д. (17 февраля 2017 г.). «Сложная эволюционная история аминоацил-тРНК синтетаз». Исследования нуклеиновых кислот. 45 (3): 1059–1068. Дои:10.1093 / нар / gkw1182. ISSN 0305-1048. ЧВК 5388404. PMID 28180287.
- ^ а б Нтунтуми, Криса; Властаридис, Панайотис; Мосиалос, Димитрис; Статопулос, Константинос; Илиопулос, Иоаннис; Промпонас, Василиос; Оливер, Стивен Дж. Амуциас, Григорис Д. (04.11.2019). «Области низкой сложности в белках прокариот выполняют важные функциональные роли и являются высококонсервативными». Исследования нуклеиновых кислот. 47 (19): 9998–10009. Дои:10.1093 / нар / gkz730. ISSN 0305-1048. ЧВК 6821194. PMID 31504783.
дальнейшее чтение
- Balbach, J .; Schmid, F. X. (2000), "Изомеризация пролина и ее катализ в сворачивании белка", в Pain, R.H. (ed.), Механизмы сворачивания белков (2-е изд.), Oxford University Press, стр. 212–49, ISBN 978-0-19-963788-1.
- Подробный научный обзор нарушений метаболизма пролина и гидроксипролина можно найти в главе 81 OMMBID. Чарльз Скрайвер, Beaudet, A.L., Valle, D., Sly, W.S., Vogelstein, B., Childs, B., Kinzler, K.W. (Проверено в 2007 г.). Онлайн-метаболические и молекулярные основы наследственных заболеваний. Нью-Йорк: Макгроу-Хилл. - Резюме 255 глав, полный текст по многим университетам. Также есть Блог OMMBID.
внешняя ссылка
| Общие темы |  | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| По свойствам |
| ||||||||||
| |||||||||||