Зауропода - Sauropoda
| Зауроподы | |
|---|---|
 | |
| Навесной каркас Апатозавр луизский, Музей Карнеги | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Подотряд: | †Зауроподоморфа |
| Clade: | †Анхизаврия |
| Clade: | †Зауропода Марш, 1878 |
| Подгруппы[1] | |
| Синонимы | |
| |
Зауропода /sɔːˈрɒпədə/, члены которого известны как зауроподы /ˈsɔːrəпɒdz/[2][3] (от сауро- + -под, "ящерица -оногий "), является клады из заурисхий ("ящерица") динозавры. У зауроподов были очень длинные шеи, длинные хвосты, маленькие головы (по сравнению с остальной частью их тела) и четыре толстые, похожие на столбы ноги. Они отличаются огромными размерами, достигнутыми некоторыми видами, и в эту группу входят самые большие животные, когда-либо обитавшие на суше. Хорошо известный роды включают Брахиозавр, Диплодок, Апатозавр и Бронтозавр.[4][5]
Самые старые известные динозавры-завроподы известны с Раннеюрский период.[6] Исанозавр и Антетонитрус первоначально были описаны как Триасовый зауроподы,[7][8] но их возраст, а в случае Антетонитрус впоследствии был поставлен под сомнение также его статус зауроподов.[9][6][10] Следы зауроподоморфа, похожие на следы зауроподоморфа из Формация Флеминг-фьорд (Гренландия ) может, однако, указывать на вхождение группы в Поздний триас.[6] Посредством Поздняя юра (150 миллионов лет назад) завроподы получили широкое распространение (особенно диплодоциды и брахиозавриды ). Посредством Поздний мел, одна группа зауроподов, титанозавры, заменил все остальные и имел почти глобальное распространение. Однако, как и все другие нептичьи динозавры, жившие в то время, титанозавры вымер в Меловое – палеогеновое вымирание. Окаменелые останки зауроподов были найдены на всех континентах, в том числе Антарктида.[11][12][13][14]
Название Sauropoda было придумано O.C. Марш в 1878 г. и происходит от Греческий, что означает «нога ящерицы».[15] Зауроподы - одна из самых узнаваемых групп динозавров, которая стала неотъемлемой частью популярной культуры из-за своих внушительных размеров.
Полные ископаемые находки зауроподов редки. Многие виды, особенно самые крупные, известны только по изолированным и разрозненным костям. У многих почти полных экземпляров отсутствуют головы, кончики хвостов и конечности.
Описание
Зауроподы были травоядный (поедание растений), обычно довольно длинношеие[16] четвероногие (четвероногие), часто с лопатчатыми (лопатообразными: широкими на конце, узкими на шейке) зубами. У них были крошечные головы, массивные тела и у большинства были длинные хвосты. Их задние лапы были толстыми, прямыми и мощными, заканчивались булавовидными ступнями с пятью пальцами, хотя только на внутренних трех (или в некоторых случаях четырех) были когти. Их передние конечности были более тонкими и обычно заканчивались столпоподобными руками, рассчитанными на поддержку веса; часто коготь был только на большом пальце. Многие иллюстрации зауроподов во плоти упускают эти факты, неточно изображая зауроподов с копытами, покрывающими пальцы без когтей, или с более чем тремя когтями или копытами на руках. В проксимальный хвостовые позвонки чрезвычайно диагностический для зауроподов.[17]
Размер
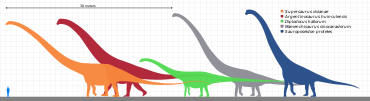
Наиболее определяющей характеристикой зауроподов был их размер. Даже карликовые зауроподы (от 5 до 6 метров или 20 футов в длину) считались одними из самых крупных животных в их экосистема. Их единственные реальные конкуренты по размеру - это Rorquals, такой как синий кит. Но, в отличие от китов, зауроподы были в первую очередь наземные животные.
Их строение тела не отличалось так сильно, как у других динозавров, возможно, из-за ограничений по размеру, но они были достаточно разнообразными. Некоторые, как диплодоциды, обладали чрезвычайно длинными хвостами, которые они, возможно, были в состоянии трещина как хлыст в качестве сигнала или для отпугивания или ранения хищников,[18] или сделать звуковые удары.[19][20] Суперзавр от 33 до 34 метров (от 108 до 112 футов) в длину,[21] был самым длинным зауроподом, известным из достаточно полных останков, но другие, такие как старый рекордсмен, Диплодок, также были очень длинными. В голотип (и теперь потеряно) позвонок из Amphicoelias fragillimus (сейчас же Maraapunisaurus) могло произойти от животного длиной 58 метров (190 футов);[22] его позвоночник был бы значительно длиннее, чем у синего кита. Однако исследование, опубликованное в 2015 году, предполагает, что оценки размеров A. fragillimus возможно, было сильно преувеличено.[23] Самый длинный динозавр, известный из разумных окаменелостей, вероятно, Аргентинозавр huinculensis с оценкой длины от 35 метров (115 футов) до 36 метров (118 футов) согласно последним исследованиям.[24][25] Однако гигант Барозавр Образец BYU 9024 мог быть даже больше, достигая длины 45-48 метров (148-157 футов).[24][25][26]
Самое длинное наземное животное из ныне живущих, сетчатый питон, достигает длины всего 6,95 метра (22,8 фута).[27]
Другие, как брахиозавриды, были чрезвычайно высокими, с высокими плечами и очень длинными шеями. Завропосейдон был, вероятно, самым высоким, достигая высоты около 18 метров (60 футов), с предыдущим рекордом самой длинной шеи, который держал Маменчизавр. Для сравнения: жирафа, самый высокий из всех ныне живущих наземных животных, имеет рост от 4,8 до 5,5 метров (от 16 до 18 футов).
Лучшее свидетельство указывает на то, что самые массовые были Аргентинозавр (65-80 метрических тонн[28][24][25]), Маменчизавр синоканадорум (60-80 метрических тонн[25]), гигант Барозавр образец (60-80 + метрические тонны[24][25][26]) и Патаготитан с участием Пуэртазавр (50-55 метрических тонн [24][25]). Существовали скудные (и теперь отсутствующие) доказательства того, что так называемые Bruhathkayosaurus, мог бы весить более 175 метрических тонн, но это ставится под сомнение.[29] Вес Amphicoelias fragillimus оценивается в 122,4 метрических тонны[22] но исследование 2015 года показало, что эти оценки могли быть сильно преувеличены.[23] Самое крупное наземное животное, живущее сегодня, кустовой слон, весит не более 10,4 метрических тонн (11,5 коротких тонн).[30]
Среди самых маленьких зауроподов были примитивные Омденозавр (4 м или 13 футов в длину), карлик титанозавр Мадьярозавр (6 м или 20 футов в длину), а карлик брахиозаврид Европазавр, длина которого у взрослого человека составляла 6,2 метра.[31] Его небольшой рост, вероятно, был результатом островная карликовость встречается в популяции зауроподов, изолированных на острове в конце юрского периода на территории современной области Лангенберг на севере Германия.[32][33] В диплодокоид зауропод Брахитрахелопан был самым коротким членом своей группы из-за необычно короткой шеи. В отличие от других зауроподов, чьи шеи могут вырасти в четыре раза длиннее их спины, шея Брахитрахелопан был короче его позвоночника.
Незадолго до этого 29 марта 2017 года след зауропода длиной около 5,6 футов (1,7 метра) был обнаружен на Валмадани в регионе Кимберли Западной Австралии.[34] В сообщении говорится, что это был самый крупный из известных случаев. В 2020 году Молина-Перес и Ларраменди оценили размер животного в 31 метр (102 фута) и 72 тонны (79,4 коротких тонны), основываясь на длине следа 1,75 метра (5,7 фута).[24]
Конечности и ступни
Как массивный четвероногие, зауроподы развили специализированные гравипортальный (несущие) конечности. Задние лапы были широкими и у большинства видов сохраняли по три когтя.[35] Особенно необычными по сравнению с другими животными были сильно измененные передние лапы (манус ). Передние лапы зауроподов очень отличались от передних лап современных крупных четвероногих животных, таких как слоны. Вместо того, чтобы расширяться в стороны, чтобы создать широкую ногу, как у слонов, манус кости зауроподов были расположены в полностью вертикальные колонны с чрезвычайно редкими костями пальцев (хотя неясно, были ли самые примитивные зауроподы, такие как Вулканодон и Барапазавр, были такие передние лапы).[36] Передние лапы были настолько модифицированы в эусауроподы что отдельные цифры не были бы видны при жизни.
Расположение кости переднего отдела стопы (пястный ) столбики у евзауроподов были полукруглыми, поэтому отпечатки передних ног зауроподов имеют подковообразную форму. В отличие от слонов, печатные свидетельства показывают, что у зауроподов отсутствовала какая-либо мясистая подкладка для поддержки передних лап, что делало их вогнутыми.[36] Единственным когтем, видимым у большинства зауроподов, был характерный коготь большого пальца (связанный с пальцем I). Такой коготь был почти у всех зауроподов, но неизвестно, для чего он служил. Коготь был самым большим (а также высоким и сплющенным с боков) у диплодоцидов и очень маленьким у брахиозавридов, некоторые из которых, судя по свидетельствам следов,, похоже, полностью утратили коготь.[37]
Титанозавры могли полностью потерять коготь большого пальца (за исключением ранних форм, таких как Janenschia ). Титанозавры были самыми необычными среди зауроподов, так как помимо внешнего когтя они полностью утратили пальцы передней лапы. У продвинутых титанозавров не было пальцев или костей пальцев, и они ходили только на подковообразных «культях», составленных из столбчатых пястных костей.[38]
Распечатать доказательства из Португалия показывает, что, по крайней мере, у некоторых зауропод (вероятно, брахиозаврид) нижняя и боковые части передней части стопы, вероятно, были покрыты небольшими колючими чешуйками, которые оставляли следы на отпечатках.[39] У титанозавров концы пястных костей, соприкасавшиеся с землей, были необычно широкими и прямоугольными, а у некоторых экземпляров сохранились остатки мягкой ткани, покрывающей эту область, что позволяет предположить, что передние лапы у этих видов были окаймлены какой-то набивкой.[38]
Мэтью Боннан[40][41] показал, что длинные кости динозавров зауроподов выросли изометрически: то есть практически не было изменений в форме, когда молодые зауроподы превратились в гигантских взрослых особей. Боннан предположил, что этот странный характер масштабирования (у большинства позвоночных наблюдаются значительные изменения формы длинных костей, связанные с увеличением поддержки веса) может быть связан с принципом ходулочника (предложенным ученым-любителем Джимом Шмидтом), в котором длинные ноги взрослых зауроподов позволяли им легко преодолевать большие расстояния без изменения общей механики.
Воздушные мешочки
Наряду с другими заурисхий динозавры (такие как тероподы, включая птиц), зауроподы имели систему воздушные мешки, о чем свидетельствуют вмятины и полые полости на большей части их позвонки которые были захвачены ими. Пневматические, полые кости являются характерной чертой всех зауропод.[42] Эти воздушные пространства уменьшали общий вес массивных шей, которые были у зауроподов, и система воздушного мешка в целом, позволяя односторонний поток воздуха через жесткие легкие, позволяла зауроподам получать достаточно кислорода.[43]
Птичья выемка костей зауроподов была обнаружена на раннем этапе изучения этих животных, и, по сути, по крайней мере один экземпляр зауроподов был обнаружен в XIX веке (Орнитопсис ) изначально был ошибочно идентифицирован как птерозавр (летающая рептилия) из-за этого.[44]
Броня

У некоторых зауроподов броня. Были роды с маленькими клубы на их хвостах, как Шунозавр, и несколько титанозавры, такие как Сальтазавр и Ампелозавр, имел маленькие костлявые остеодермы покрывая части их тел.
Зубы
Исследование Майкла Д'Эмика и его коллег из Университета Стоуни-Брук показало, что зауроподы эволюционировали с высокой степенью замещения зубов, чтобы не отставать от своего большого аппетита. Исследование показало, что Нигерзавр, например, заменяли каждый зуб каждые 14 дней, Камаразавр заменяли каждый зуб каждые 62 дня, и Диплодок заменяли каждый зуб раз в 35 дней.[45] Ученые обнаружили, что качества зуба влияют на то, сколько времени требуется для роста нового зуба. Камаразаврзубы росли дольше, чем у Диплодок потому что они были больше.[46]
Д'Эмик и его команда также отметили, что различия между зубами зауроподов также указывают на разницу в питании. Диплодок ел растения низко к земле и Камаразавр просматривали листья с верхней и средней ветвей. По словам ученых, специализация их диет помогала сосуществовать разным травоядным динозаврам.[45][46]
Шеи
Шеи зауроподов были найдены более 15 метров (49 футов) в длину, что в шесть раз длиннее шеи жирафа, ставшего мировым рекордом.[47] Обеспечение этого было рядом важных физиологических особенностей. В целом большой размер тела динозавров и положение на четвероногих ногах обеспечили стабильную основу для поддержки шеи, а голова стала очень маленькой и легкой, потеряв способность переваривать пищу орально. Из-за того, что их головы превращались в простые орудия сбора урожая, с помощью которых растения попадали в тело, зауроподам требовалось меньше энергии для подъема головы, и, таким образом, они могли развить шеи с менее плотными мышцами и соединительной тканью. Это резко уменьшило общую массу шеи, что позволило продолжить удлинение.
У зауроподов также было множество приспособлений в строении скелета. У некоторых зауроподов насчитывалось до 19 шейные позвонки, тогда как почти все млекопитающие ограничиваются семью. Кроме того, каждый позвонок был чрезвычайно длинным и имел множество пустот, которые могли быть заполнены только воздухом. Система воздушных мешков, соединенная с пространствами, не только облегчила длинные шеи, но и эффективно увеличила поток воздуха через трахею, помогая существам дышать достаточным количеством воздуха. Развивая позвонки, состоящие на 60% из воздуха, зауроподы смогли минимизировать количество плотных и тяжелых костей, не жертвуя способностью делать достаточно большие вдохи для снабжения всего тела кислородом.[47] По словам Кента Стивенса, компьютерная реконструкция скелетов, сделанных из позвонков, показывает, что шеи зауроподов были способны подметать большие области кормления без необходимости перемещать их тела, но не могли быть втянуты в положение намного выше плеч для исследования площадь или выше.[48]
Другая предполагаемая функция длинной шеи зауроподов заключалась в том, чтобы отводить огромное количество тепла, производимого их большой массой тела. Принимая во внимание, что метаболизм выполнял бы огромную работу, он определенно генерировал бы большое количество тепла, и устранение этого избыточного тепла было бы важным для выживания.[49]Также предполагалось, что длинные шеи охлаждали вены и артерии, идущие к мозгу, избегая попадания чрезмерно нагретой крови в голову. Фактически было обнаружено, что увеличение скорости метаболизма, вызванное шеей зауроподов, было немного более чем компенсировано дополнительной площадью поверхности, с которой могло рассеиваться тепло.[50]
Палеобиология
Экология
Когда зауроподы были впервые обнаружены, их огромный размер побудил многих ученых сравнить их с современными. киты. Большинство исследований XIX и начала XX веков пришли к выводу, что зауроподы были слишком большими, чтобы выдерживать свой вес на суше, и, следовательно, они должны были в основном водный. Большинство реставраций жизни зауроподов в искусстве первых трех четвертей 20 века изображали их полностью или частично погруженными в воду.[51] Это раннее представление было поставлено под сомнение в 1950-х годах, когда исследование Кермака (1951) продемонстрировало, что если бы животное погрузили в воду на несколько метров, давления было бы достаточно, чтобы фатально схлопнуть легкие и дыхательные пути.[52] Однако это и другие ранние исследования зауроподов экология были ошибочными в том, что они игнорировали значительный объем доказательств того, что тела зауроподов были сильно пропитаны воздушные мешки. В 1878 году палеонтолог Э. Коуп даже называл эти конструкции «поплавками».
Начиная с 1970-х годов, начали изучаться эффекты воздушных мешков зауроподов на их предполагаемый водный образ жизни. Палеонтологи, такие как Кумбс и Баккер, использовали это, а также доказательства из седиментология и биомеханика, чтобы показать, что зауроподы были в основном наземными животными. В 2004 году Д. Хендерсон заметил, что из-за их обширной системы воздушных мешков зауроподы были бы плавучими и не могли бы полностью погрузить свое туловище под поверхность воды; Другими словами, они будут плавать, и им не грозит коллапс легких из-за давления воды во время плавания.[51]
Доказательства плавания у зауроподов получены по ископаемым следам, которые, как иногда находили, сохраняли отпечатки только передних лап (рук). Хендерсон показал, что такие следы могут быть объяснены зауроподами с длинными передними конечностями (такими как макронарианцы ) плавает на относительно мелкой воде, достаточно глубокой, чтобы короткие задние лапы не касались дна, и используя передние конечности для плоскодонка вперед.[51] Однако из-за своих пропорций тела плавающие зауроподы также были бы очень нестабильными и неадаптированными в течение длительного времени в воде. Этот режим водного движение в сочетании с его нестабильностью, Хендерсон назвал зауроподов в воде «подвыпившими игроками».[51]
Таким образом, хотя зауроподы не могли быть водными, как это исторически изображалось, есть свидетельства того, что они предпочитали влажные и прибрежные места обитания. Следы зауроподов обычно обнаруживаются вдоль береговой линии или пересекают поймы, а окаменелости зауроподов часто встречаются во влажной среде или смешиваются с окаменелостями морских организмов.[51] Хорошим примером этого может быть массивный Юрский следы зауроподов найдены в лагуна депозиты на Шотландия с Остров Скай.[53]
Выпас и родительская забота

Многие линии окаменелостей как из костных пластов, так и из тропинок указывают на то, что зауроподы были стадными животными, которые сформировали стада. Однако состав стада варьировался у разных видов. Некоторые костные пласты, например участок из Средняя юра из Аргентина, похоже, показывают стада, состоящие из особей разных возрастных групп, смешанных с молодыми и взрослыми особями. Однако ряд других ископаемых участков и троп указывает на то, что многие виды зауропод передвигались стадами, разделенными по возрасту, причем молодые особи составляли стада отдельно от взрослых особей. Такие стратегии сегрегированного выпаса были обнаружены у таких видов, как Аламозавр, Беллузавр и немного диплодоциды.[54]
В обзоре данных о различных типах стад Майерс и Фиорилло попытались объяснить, почему зауроподы, по-видимому, часто образовывали раздельные стада. Исследования микроскопического изнашивания зубов показывают, что у молодых зауроподов была диета, отличавшаяся от их взрослых собратьев, поэтому совместное выпас скота не было бы таким продуктивным, как раздельное, когда отдельные члены стада могли кормиться скоординированно. Огромная разница в размерах между молодыми и взрослыми особями, возможно, также сыграла свою роль в различных стратегиях кормления и выпаса.[54]

Поскольку сегрегация молоди и взрослых особей должна была произойти вскоре после вылупления и в сочетании с тем фактом, что вылупившиеся детеныши зауроподов, скорее всего, были ранний, Майерс и Фиорилло пришли к выводу, что виды с сегрегированными по возрасту стадами не проявили бы особой родительской заботы.[54] С другой стороны, ученые, изучавшие смешанные по возрасту стада зауроподов, предположили, что эти виды могли заботиться о своих детенышах в течение длительного периода времени, прежде чем молодые достигли взрослого возраста.[55] Исследование 2014 года показало, что время от откладки яйца до момента вылупления могло составлять от 65 до 82 дней.[56] Неизвестно, насколько именно сегрегированное и смешанное по возрасту стадо варьировалось в разных группах зауроподов. Дальнейшие примеры стадного поведения необходимо будет обнаружить у большего количества видов зауроподов, чтобы начать обнаружение возможных закономерностей распространения.[54]
Задняя позиция

С самого начала истории их исследования ученые, такие как Осборн, предположили, что зауроподы могут вставать на задние лапы, используя хвост как третью «ногу» треноги.[57] Скелетная гора, изображающая диплодоцид Барозавр лентус вставая на задние лапы в Американский музей естественной истории одна из иллюстраций этой гипотезы. В статье 2005 года Ротшильд и Мольнар рассуждали, что, если бы зауроподы время от времени принимали позу на двух ногах, были бы доказательства стрессовых переломов в «руках» передних конечностей. Однако после того, как они исследовали большое количество скелетов зауроподов, их не обнаружили.[58]

Генрих Маллисон (в 2009 г.) был первым, кто изучил физический потенциал различных зауроподов, чтобы они занимали триподную стойку. Мэллисон обнаружил, что некоторые персонажи, ранее связанные с приспособлениями для выращивания, на самом деле не связаны (например, широко расставленные бедренные кости титанозавры ) или препятствовали бы выращиванию. Например, у титанозавров был необычайно гибкий позвоночник, который снизил бы устойчивость в позе трипода и увеличил бы нагрузку на мышцы. Точно так же маловероятно, чтобы брахиозавриды могли вставать на задние лапы, поскольку их центр тяжести был намного дальше вперед, чем у других зауроподов, что могло бы сделать такую стойку нестабильной.[59]
С другой стороны, диплодоциды, по-видимому, были хорошо приспособлены для того, чтобы встать на триподную стойку. У диплодоцидов центр масс находился прямо над бедрами, что давало им больший баланс на двух ногах. У диплодоцидов также были самые подвижные шеи зауроподов, мускулистый тазовый пояс и хвостовые позвонки особой формы, которые позволяли хвосту выдерживать вес в точке касания земли. Мэллисон пришел к выводу, что диплодоциды лучше приспособлены к выращиванию, чем слоны, которые иногда случаются в дикой природе. Он также утверждает, что стрессовые переломы в дикой природе не возникают в результате повседневного поведения,[59] такие как деятельность, связанная с кормлением (против Ротшильда и Мольнара).[58]
Поза головы и шеи
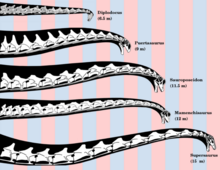
Существуют разногласия по поводу того, как зауроподы держали голову и шею, а также позы, которых они могли достичь при жизни. Различные исследования рассматривают проблему с разных сторон, таких как нейтральное сочленение шейного позвонка и оценивают диапазон движений, метаболический и потребность в энергии, связанная с невероятно длинной шеей, и сравнение с живыми животными, пришли к разным выводам. Утверждение о том, что длинные шеи зауроподов использовались для просмотра высоких деревьев, было поставлено под сомнение на основании расчетов энергии, необходимой для создания артериального давления для головы, если ее держать вертикально.[60] Эти расчеты показывают, что это заняло бы примерно половину потребляемой энергии.[61] Кроме того, для подачи крови к голове, вертикально удерживаемой высоко, потребовалось бы артериальное давление около 700. мм рт. ст. (= 0.921 бар ) в сердце. Для этого потребовались бы сердца в 15 раз больше сердец китов такого же размера.[62] Это использовалось, чтобы доказать, что более вероятно, что длинная шея обычно держалась горизонтально, чтобы они могли питаться растениями на очень большой площади без необходимости перемещать свое тело - потенциально значительная экономия энергии для животных весом от 30 до 40 тонн. . В подтверждение этого реконструкции шеи Диплодок и Апатозавр покажите, что они в основном прямые с легким наклоном, ориентируя голову и шею в «нейтральной, неотклоненной позе».[63]
Однако исследования живых животных показали, что большинство живых четвероногие при настороженности обычно приподнимают основание шеи. Вывод из костей о «нейтральных позах», предполагающих более горизонтальное положение,[63] может быть ненадежным.[64][65] В опубликованном в 2013 г. исследовании страус шеи, однако, поставили под сомнение предполагаемую гибкость шеи зауроподов. Исследования Мэтью Кобли и др. С помощью компьютерного моделирования показали, что прикрепления мышц и хрящи шеи, вероятно, в значительной степени ограничивают гибкость. Авторы предостерегают от оценки диапазона движений, просто используя только кости. Это открытие также показывает, что зауроподам, возможно, приходилось перемещать все свое тело, чтобы получить лучший доступ к местам, где они могли пастись и бродить по растительности.[66][67][68]
Пути и передвижение



Зауропод пути и другие ископаемые следы (известные как «ихниты») известны по многочисленным свидетельствам, имеющимся на большинстве континентов. Ichnites помогли поддержать другие биологические гипотезы о зауроподах, включая общую анатомию передних и задних лап (см. Конечности и ступни над). Как правило, отпечатки передних лап намного меньше задних и часто имеют форму полумесяца. Иногда ихниты сохраняют следы когтей и помогают определить, какие группы зауропод потеряли когти или даже пальцы на передних лапах.[69]
Следы зауроподов из Формация Вильяр-дель-Арсобиспо раннего Берриасский возраст в Испании поддерживает стадное поведение группы. Возможно, треки больше похожи на Зауроподихнус гигантский чем любые другие ихногенеры, хотя предполагалось, что они происходят от базальных титанозавров. Гусеницы ширококолейные, а группировка максимально приближена к Зауроподихнус также подтверждается расстоянием от кисти до стопы, морфологией кисти, имеющей форму фасоли, и морфологией стопы субтреугольной формы. Невозможно определить, были ли следы стада нанесены молодыми или взрослыми особями, из-за отсутствия предварительной идентификации индивидуального возраста следа.[70]

Обычно следы зауроподов делятся на три категории в зависимости от расстояния между противоположными конечностями: узкая колея, средняя колея и широкая колея. Калибр пути может помочь определить, насколько широко расставлены конечности различных зауроподов и как это могло повлиять на их походку.[69] Исследование 2004 г., проведенное Дей и его коллегами, показало, что среди групп продвинутых зауроподов можно найти общую закономерность, при этом каждое семейство зауроподов характеризуется определенными толщинами колеи. Они обнаружили, что большинство зауроподов, кроме титанозавры конечности узкие, с сильными отпечатками большого пальца на передних лапах. Следы средней колеи с отпечатками когтей на передних лапах, вероятно, принадлежат брахиозавридам и другим примитивным титанозаврообразные, у которых развились более широкие конечности, но сохранились когти. Первобытные истинные титанозавры также сохранили когти переднего отдела стопы, но развили конечности полностью широкой толщины. Конечности с широкой шириной были сохранены продвинутыми титанозаврами, следы от которых показывают широкую колею и отсутствие каких-либо когтей или пальцев на передних лапах.[71]
Иногда встречаются только следы от передних лап. Фолкингем и другие. [72] использовали компьютерное моделирование, чтобы показать, что это может быть связано со свойствами подложки. Они должны быть в самый раз, чтобы сохранить следы.[73] Различия в площади поверхности задних и передних конечностей и, следовательно, контактного давления с субстратом могут иногда приводить к сохранению только путей следа передних конечностей.
Биомеханика и скорость

В исследовании, опубликованном в PLoS ONE 30 октября 2013 г., Билл Селлерс, Родольфо Кориа, Ли Маргеттс и другие., Аргентинозавр был реконструирован в цифровом виде, чтобы впервые проверить его передвижение. До исследования наиболее распространенным способом оценки скорости было изучение костной ткани. гистология и ихнология. Обычно исследования гистологии и скорости костей зауроподов сосредоточены на посткраниальном скелете, который обладает многими уникальными особенностями, такими как увеличенный отросток на локтевая кость, широкий лепесток на ilia, наклоненная внутрь верхняя треть бедренная кость, и очень яйцевидный стержень бедренной кости. Эти функции полезны при попытке объяснить паттерны следов гравипортальных животных. При изучении ихнологии для расчета скорости зауроподов возникает несколько проблем, таких как предоставление оценок только для определенных походок из-за предвзятости к сохранению и наличие гораздо большего количества проблем с точностью.[74]

Оценить походку и скорость Аргентинозавр, Исследование провели анализ опорно-двигательного аппарата. Единственный предыдущие мышечно-скелетные исследования были проведены на гоминоиды, ужасные птицы, и другие динозавры. Прежде чем они смогли провести анализ, команда должна была создать цифровой скелет рассматриваемого животного, показать, где будет расслоение мышц, определить местонахождение мышц и суставов и, наконец, определить свойства мышц, прежде чем определить походку и скорость. Результаты исследования биомеханики показали, что Аргентинозавр был механически компетентен на максимальной скорости 2 м / с (5 миль в час), учитывая большой вес животного и нагрузку, которую могли выдерживать его суставы. [75] Результаты также показали, что возможно появление гораздо более крупных наземных позвоночных, но для предотвращения коллапса суставов потребуется значительная реконструкция тела и возможное достаточное изменение поведения.[74]
Размер тела

Зауроподы были гигантскими потомками удивительно маленьких предков. Базальные динозаврообразные, такие как Псевдолагозух и Марасух из среднего триаса Аргентины, весил приблизительно 1 кг (2,2 фунта) или меньше. Они превратились в заурисхии, которые быстро увеличили количество бауплан размер, хотя более примитивные члены любят Эораптор, Панфагия, Pantydraco, Сатурналии и Гуайбазавр все еще сохранил умеренный размер, возможно, менее 10 кг (22 фунта). Даже с этими небольшими примитивными формами среди зауроподоморфов наблюдается заметное увеличение размеров, хотя скудные остатки этого периода делают интерпретацию предположительной. Есть один конкретный пример небольшого производного зауроподоморфа: Анхизавр, менее 50 кг (110 фунтов), хотя он ближе к зауроподам, чем Платеозавр и Риохазавр, которые имели вес более 1 т (0,98 длинных тонн; 1,1 коротких тонны).[49]
Произошедшие от зауроподоморфов, зауроподы были огромными. Их гигантский размер, вероятно, был результатом увеличения скорости роста, что стало возможным благодаря тахиметаболическая эндотермия, черта, которая эволюционировала у зауроподоморфов. Разветвившись на зауроподов, зауроподоморфы продолжали неуклонно увеличиваться в размерах с более мелкими зауроподами, такими как раннеюрские. Барапазавр и Котасавр, эволюционируя в еще более крупные формы, такие как среднеюрские Маменчизавр и Патагозавр. Отвечая на рост зауроподов, их хищники-тероподы также росли, как показано на рисунке. Аллозавр размер целофизоид от Германия.[49]
Размер у Neosauropoda
Neosauropoda вполне вероятно, что это кладовая динозавров с самыми большими размерами тела из когда-либо существовавших. Предполагается, что несколько исключений меньшего размера вызваны островная карликовость, хотя есть тенденция Титанозаврия в сторону меньшего размера. Однако титанозавры были одними из самых крупных зауроподов в истории. Помимо титанозавров, клада диплодокоидов, группа гигантов, называемая Dicraeosauridae, определяется небольшим размером тела. Однако ни один зауропод не был очень маленьким, поскольку даже «карликовые» зауроподы крупнее 500 кг (1100 фунтов), что составляет лишь около 10% всех видов млекопитающих.[49]
Независимый гигантизм
Хотя в целом зауроподы были крупными, гигантский размер (40 т (39 длинных тонн; 44 коротких тонны) или более) достигался независимо друг от друга несколько раз в их эволюции. Многие гигантские формы существовали в поздней юре (в частности, Кимериджский и Туронский ), такой как туриазавр Туриазавр и диплодокоиды Амфикоэлии, Диплодок и Суперзавр. В период от раннего до позднего мелового периода гиганты Завропосейдон, Паралититан, Аргентинозавр, Пуэртазавр, Антарктозавр гигантеус, Дредноутус Schrani, Notocolossus и Футалогкозавр жили, самый ранний из которых был брахиозавридом, а все последние - титанозаврами. Один малоизвестный возможный гигант - это Хуангетитан ruyangensis, известен только по ребрам длиной 3 м (9,8 футов). Эти гигантские виды жили в период от поздней юры до позднего мела, появляясь независимо друг от друга в течение 85 миллионов лет.[49]
Островная карликовость у зауроподов
Два хорошо известных островных карликовых вида зауроподов относятся к меловому периоду. Мадьярозавр (в какой-то момент его идентичность как карлика была оспорена) и юрский период Европазавр, оба из Европы. Несмотря на то, что эти зауроподы маленькие, единственный способ доказать, что они настоящие карлики, - это изучение их гистологии костей. В исследовании, проведенном Мартином Сандером и его коллегами в 2006 году, было изучено одиннадцать человек Europasaurus holgeri используя гистологию костей, и продемонстрировал, что небольшие островные виды эволюционировали за счет снижения скорости роста длинных костей по сравнению со скоростью роста предковых видов на материке.[76] Два других возможных карлика Рапетозавр, существовавший на острове Мадагаскар, изолированный остров в меловом периоде, и Ампелозавр, титанозавр, обитавший на Пиренейском полуострове на юге Испании и Франции. Возможный Цетиозаврикус из Швейцарии тоже может быть карликом, но это еще предстоит доказать.[49] Один из самых крайних случаев островной карликовости встречается у Европазавр, родственник гораздо большего Камаразавр и Брахиозавр: он был всего около 6,2 м (20 футов) в длину, что является отличительной чертой этого вида. Как и у всех карликовых видов, их низкая скорость роста привела к их небольшому размеру.[77][49]
История открытия
Первые обрывки окаменелостей, которые теперь признаны зауроподами, произошли от Англия и изначально интерпретировались по-разному. Их родство с другими динозаврами было признано только после их первоначального открытия.


Первой окаменелостью зауроподов, которая была описана с научной точки зрения, был единственный зуб, известный неверующим.Линнеевский дескриптор Рутеллум импликатум.[78] Эта окаменелость была описана Эдвард Лхойд в 1699 году, но в то время не был признан гигантской доисторической рептилией.[79] Динозавры будут признаны группой только спустя столетие.
Ричард Оуэн опубликовал первые современные научные описания зауроподов в 1841 году в книге и статье, называя Кардиодон и Цетиозавр. Кардиодон был известен только по двум необычным зубам в форме сердца (от которых и получил свое название), которые нельзя было идентифицировать, кроме того факта, что они произошли от ранее неизвестного большого рептилия. Цетиозавр был известен немного лучше, но все еще остается лоскутным. Оуэн тогда думал, что Цетиозавр была гигантской морской рептилией, родственной современной крокодилы отсюда и его название, которое означает «китовая ящерица». Год спустя, когда Оуэн придумал название Динозаврия, он не включил Цетиозавр и Кардиодон в этой группе.[80]
В 1850 г. Гидеон Мантелл признал динозавровую природу нескольких костей, относящихся к Цетиозавр пользователя Owen. Мантелл заметил, что кости ног содержат мозговая полость, характеристика наземных животных. Он отнес эти образцы к новому род Пелорозавр, и сгруппировал его вместе с динозаврами. Однако Мантелл так и не узнал родство с Цетиозавр.[44]
Следующей находкой зауроподов, которая была описана и ошибочно идентифицирована как нечто иное, чем динозавр, был набор тазобедренных позвонков. Гарри Сили в 1870 году. Сили обнаружил, что позвонки имеют очень легкую конструкцию для своего размера и содержат отверстия для воздушные мешки (пневматизация). Такие воздушные мешки в то время были известны только в птицы и птерозавры, а Сили считал, что позвонки произошли от птерозавра. Он назвал новый род Орнитопсис, или "птичье лицо" из-за этого.[44]
Когда более полные образцы Цетиозавр были описаны Филлипсом в 1871 году, он наконец узнал в животном динозавра, родственного Пелорозавр.[81] Однако только после описания новых, почти полных скелетов зауроподов из Соединенные Штаты (представляющий Апатозавр и Камаразавр ) позже в том же году появилась полная картина зауроподов. Приблизительная реконструкция полного скелета зауроподов была произведена художником Джоном А. Райдером, нанятым палеонтологом Э. Коуп, основанный на останках Камаразавр, хотя, согласно более поздним находкам и биомеханическим исследованиям, многие особенности все еще были неточными или неполными.[82] Также в 1877 г. Ричард Лидеккер назвал имя другого родственника Цетиозавр, Титанозавр, на основе изолированного позвонка.[44]

В 1878 г. наиболее полный зауропод был обнаружен и описан Отниэль Чарльз Марш, кто назвал это Диплодок. С этой находкой Марш также создал новую группу, чтобы содержать Диплодок, Цетиозавр, и их увеличивающийся список родственников, чтобы отличить их от других основных групп динозавров. Марш назвал эту группу зауроподами, или «лапы ящерицы».[44]
Классификация
Классификация численности зауроподов в значительной степени стабилизировалась в последние годы, хотя все еще существуют некоторые неопределенности, такие как размещение Euhelopus, Гаплокантозавр, Jobaria и Nemegtosauridae.
Кладограмма после анализа, представленного Сандером и его коллегами в 2011 году.[49]
| † Зауропода |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
использованная литература
- ^ Хольц, Томас Р. младший (2012). Динозавры: наиболее полная и современная энциклопедия для любителей динозавров всех возрастов (PDF). Случайный дом.
- ^ «зауропод». Словарь Merriam-Webster.
- ^ «зауропод». Dictionary.com Несокращенный. Случайный дом.
- ^ Tschopp, E .; Mateus, O .; Бенсон, Р. Б. Дж. (2015). «Филогенетический анализ на уровне образцов и таксономическая ревизия Diplodocidae (Dinosauria, Sauropoda)». PeerJ. 3: e857. Дои:10.7717 / peerj.857. ЧВК 4393826. PMID 25870766.
- ^ blogs.scientificamerican.com тетрапод-зоология 2015-04-24 Эта вещь бронтозавра
- ^ а б c Йенс Н. Лалленсак; Хендрик Кляйн; Джеспер Милан; Оливер Уингс; Октавио Матеус; Ларс Б. Клемменсен (2017). «Следы динозавров зауроподоморфа из формации Флеминг-фьорд в Восточной Гренландии: свидетельства существования зауроподов позднего триаса». Acta Palaeontologica Polonica. 62 (4): 833–843. Дои:10.4202 / app.00374.2017.
- ^ Эрик Баффето; Варавуд Сутехорн; Жиль Куни; Хайян Тонг; Жан Ле Лёфф; Сасидхорн Кхансубха; Сути Джонгаутчариякул (2000). «Самый ранний из известных динозавров зауроподов». Природа. 407 (6800): 72–74. Bibcode:2000Натура 407 ... 72Б. Дои:10.1038/35024060. PMID 10993074. S2CID 4387776.
- ^ Адам М. Йейтс; Джеймс В. Китчинг (2003). «Самый ранний из известных динозавров зауроподов и первые шаги к перемещению зауроподов». Труды Королевского общества B: биологические науки. 270 (1525): 1753–1758. Дои:10.1098 / rspb.2003.2417. ЧВК 1691423. PMID 12965005.
- ^ Блэр В. Макфи; Адам М. Йейтс; Иона Н. Шуаньер; Фернандо Абдала (2014). «Полная анатомия и филогенетические взаимоотношения Антетонитрус ингенипес (Sauropodiformes, Dinosauria): значение для происхождения зауроподы ". Зоологический журнал Линнеевского общества. 171 (1): 151–205. Дои:10.1111 / zoj.12127.
- ^ Блэр В. Макфи; Эмес М. Борди; Lara Sciscio; Иона Н. Чойнер (2017). "Биостратиграфия завроподоморф формации Эллиот на юге Африки: отслеживание эволюции зауроподоморф через границу триаса и юры". Acta Palaeontologica Polonica. 62 (3): 441–465. Дои:10.4202 / app.00377.2017.
- ^ Фернандо Э. Новас (2009). Эпоха динозавров в Южной Америке. Издательство Индианского университета. п. 181. ISBN 978-0-253-35289-7.
- ^ Примечания по геологии Оклахомы. Геологическая служба Оклахомы. 2003. с. 40.
- ^ Бо Риффенбург (2007). Энциклопедия Антарктики. Тейлор и Фрэнсис. п. 415. ISBN 978-0-415-97024-2.
- ^ Дж. Дж. Алистер Крейм; Лондонское геологическое общество (1989). Происхождение и эволюция антарктической биоты. Геологическое общество. п. 132. ISBN 978-0-903317-44-3.
- ^ Марш, О. (1878). «Основные персонажи американских динозавров юрского периода. Часть I"". Американский журнал науки и искусства. 16 (95): 411–416. Дои:10.2475 / ajs.s3-16.95.411. HDL:2027 / hvd.32044107172876. S2CID 219245525.
- ^ Майкл П. Тейлор, Мэтью Дж. Ведель (2013). «Почему у зауроподов длинные шеи и почему у жирафов короткие шеи». PeerJ. 1: e36. Дои:10.7717 / peerj.36. ЧВК 3628838. PMID 23638372.
Шеи динозавров зауроподов были безусловно самыми длинными из всех животных ...
CS1 maint: использует параметр авторов (ссылка на сайт) - ^ Тидуэлл В., Карпентер К. и Мейер С. 2001. Новая форма титанозавра (зауропода) из пачки ядовитой полосы формации Кедр-Маунтин (нижний мел), Юта. В кн .: Мезозойская жизнь позвоночных. Д. Х. Танке и К. Карпентер (ред.). Издательство Индианского университета, ред. Д. Х. Танке и К. Карпентер. Издательство Индианского университета. 139-165.
- ^ Баккер, Роберт (1994). «Укус бронто». Земля. 3 (6): 26–33.
- ^ Петерсон, Иварс (март 2000 г.). "Кнуты и хвосты динозавров". Новости науки. Архивировано из оригинал на 2007-07-14. Получено 2007-07-07.
- ^ Myhrvold, Nathan P .; Карри, Филип Дж. (1997). «Сверхзвуковые зауроподы? Хвостовая динамика у диплодоцидов». Палеобиология. 23 (4): 393–409. Дои:10.1017 / S0094837300019801. ISSN 0094-8373.
- ^ Лавлейс, Дэвид М .; Хартман, Скотт А .; Валь, Уильям Р. (2007). «Морфология экземпляра Суперзавр (Dinosauria, Sauropoda) из формации Моррисон, штат Вайоминг, и переоценка филогении диплодоцидов ". Arquivos do Museu Nacional. 65 (4): 527–544.
- ^ а б Карпентер, К. (2006). "Крупнейшее из большого: критическая переоценка мега-зауроподов Amphicoelias fragillimus" (PDF). Бюллетень Музея естественной истории и науки Нью-Мексико. 36: 131–138. S2CID 56215581.
- ^ а б Кэри Вудрафф и Джон Р. Фостер (15 июля 2015 г.). «Хрупкое наследие Amphicoelias fragillimus (Dinosauria: Sauropoda; формация Моррисон - новейшая юра)» (PDF). PeerJ PrePrints. 3: e1037.
- ^ а б c d е ж Молина-Перес и Ларраменди (2020). Факты и цифры о динозаврах: зауроподы и другие зауроподоморфы. Нью-Джерси: Издательство Принстонского университета. С. 42–267.
- ^ а б c d е ж Пол, Грегори С. (2019). «Определение самого крупного известного наземного животного: критическое сравнение различных методов восстановления объема и массы вымерших животных» (PDF). Летопись музея Карнеги. 85 (4): 335–358. Дои:10.2992/007.085.0403. S2CID 210840060.
- ^ а б Тейлор, Майк (2019). «Суперзавр, ультразавр и дистилозавр в 2019 году, часть 2b: размер животного BYU 9024».
- ^ Фредрикссон, Г. М. (2005). «Охота на солнечных медведей сетчатым питоном в Восточном Калимантане, Индонезийское Борнео». Зоологический бюллетень Raffles. 53 (1): 165–168.
- ^ Mazzetta, G.V .; Christiansen, P .; Фаринья, Р.А. (2004). "Гиганты и причуды: размер тела некоторых южно-южноамериканских динозавров мелового периода" (PDF). Историческая биология. 16 (2–4): 71–83. CiteSeerX 10.1.1.694.1650. Дои:10.1080/08912960410001715132. S2CID 56028251.
- ^ "Гигант, которого никогда не было: Бруаткайозавр". 1 января 2012 г.
- ^ Ларраменди, А. (2015). «Высота плеч, масса тела и форма хоботков» (PDF). Acta Palaeontologica Polonica. 60. Дои:10.4202 / app.00136.2014. S2CID 2092950.
- ^ Мартин Сандер, П .; Матеуш, Октавио; Лавен, Томас; Knötschke, Нильс (2006). «Гистология костей указывает на островную карликовость у нового позднеюрского динозавра зауроподов». Природа. 441 (7094): 739–41. Bibcode:2006Натура 441..739М. Дои:10.1038 / природа04633. PMID 16760975. S2CID 4361820.
- ^ Николь Кляйн (2011). Биология динозавров зауроподов: понимание жизни гигантов. Издательство Индианского университета. п. 73. ISBN 978-0-253-35508-9.
- ^ Рейтнер, Иоахим; Ян, Цюнь; Ван, Йонгдон; Райх, Майк (6 сентября 2013 г.). Палеобиология и геобиология ископаемых лагерштеттенов в истории Земли: совместная конференция «Paläontologische Gesellschaft» и «Палеонтологического общества Китая», Геттинген, Германия, 23-27 сентября 2013 г.. Universitätsverlag Göttingen. п. 21. ISBN 978-3-86395-135-1.
- ^ Палаццо, Кьяра (28 марта 2017 г.). "Самые большие в мире следы динозавров найдены в парке Юрского периода в Австралии.'". Телеграф.
- ^ Боннан, М.Ф. 2005. Анатомия Pes у динозавров-завропод: значение для функциональной морфологии, эволюции и филогении; стр. 346-380 в К. Карпентер и В. Тидвелл (ред.), Громовые ящерицы: динозавры-зауроподоморфы. Блумингтон, Индиана: Издательство Индианского университета.
- ^ а б Боннан, Мэтью Ф. (2003). «Эволюция формы кисти у завропод динозавров: последствия для функциональной морфологии, ориентации передних конечностей и филогении». Журнал палеонтологии позвоночных. 23 (3): 595–613. Дои:10.1671 / A1108. S2CID 85667519.
- ^ Апчерч, П. (1994). «Функция когтя мануса у динозавров зауроподов». Гайя. 10: 161–171.
- ^ а б Апестегия, С. (2005). «Эволюция пястной кости титанозавра». Стр. 321-345 в Tidwell, V. and Carpenter, K. (eds.) Громовые ящерицы: динозавры зауроподоморфа. Индианаполис: Издательство Индианского университета.
- ^ Milàn, J .; Christiansen, P .; Матеус, О. (2005). «Сохраненный в трехмерном виде слепок зауроподов из верхней юры Португалии: значение для формы зауроподов и двигательной механики». Каупия. 14: 47–52.
- ^ Боннан, М. Ф. (2004). "Морфометрический анализ формы плечевой кости и бедра у моррисонов зауроподов: значение для функциональной морфологии и палеобиологии". Палеобиология. 30 (3): 444–470. Дои:10.1666 / 0094-8373 (2004) 030 <0444: maohaf> 2.0.co; 2. JSTOR 4096900.
- ^ Боннан, Мэтью Ф. (2007). «Линейный и геометрический морфометрический анализ моделей масштабирования длинных костей у юрских динозавров-неозауроподов: их функциональное и палеобиологическое значение». Анатомический рекорд. 290 (9): 1089–1111. Дои:10.1002 / ar.20578. PMID 17721981. S2CID 41222371.
- ^ Ведель, М.Дж. (2009). "Свидетельства существования птичьих воздушных мешков у заурисхийских динозавров ". (pdf) Журнал экспериментальной зоологии, 311A: 18 стр.
- ^ Taylor, M.P .; Ведель, М.Дж. (2013). «Почему у зауроподов длинные шеи и почему у жирафов короткие шеи». PeerJ. 1: e36. Дои:10.7717 / peerj.36. ЧВК 3628838. PMID 23638372.
- ^ а б c d е Тейлор, М. (2010). «Исследование динозавров зауроподов: исторический обзор». Ричард Муди, Эрик Баффето, Дэвид М. Мартил и Даррен Нейш (ред.), Динозавры (и другие вымершие ящеры): историческая перспектива. HTML аннотация.
- ^ а б Д'Эмик, Майкл Д., Уитлок, Джон А., Смит, Кэтлин М., Фишер, Дэниел К., Уилсон, Джеффри А. (17 июля 2013 г.). «Эволюция высоких показателей замещения зубов у динозавров зауроподов». PLOS ONE. 8 (7): e69235. Bibcode:2013PLoSO ... 869235D. Дои:10.1371 / journal.pone.0069235. ЧВК 3714237. PMID 23874921.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ а б Парикмахер, Элизабет (2004-06-09). «Зубная щетка не требуется: динозавры меняют улыбку каждый месяц». Christian Science Monitor. Получено 2013-08-14.
- ^ а б Taylor, M.P .; Ведель, М.Дж. (2013). «Почему у зауроподов длинные шеи и почему у жирафов короткие шеи». PeerJ. 1: e36. Дои:10.7717 / peerj.36. ЧВК 3628838. PMID 23638372.
- ^ Стивенс, К. (2013). "Сочленение шеи зауроподов: методология и мифология". PLOS ONE. 8 (10): e78572. Bibcode:2013PLoSO ... 878572S. Дои:10.1371 / journal.pone.0078572. ЧВК 3812995. PMID 24205266.
- ^ а б c d е ж г час Сандер, П. Мартин; Кристиан, Андреас; Клаус, Маркус; Фехнер, Регина; Джи, Кэрол Т .; Грибелер, Ева-Мария; Гунга, Ханнс-Кристиан; Хуммель, Юрген; Маллисон, Генрих; Перри, Стивен Ф .; и другие. (2011). «Биология динозавров зауроподов: эволюция гигантизма». Биологические обзоры. 86 (1): 117–155. Дои:10.1111 / j.1469-185X.2010.00137.x. ISSN 1464-7931. ЧВК 3045712. PMID 21251189.
- ^ Хендерсон, Д. (2013). «Шеи зауроподов: действительно ли они из-за потери тепла?». PLOS ONE. 8 (10): e77108. Bibcode:2013PLoSO ... 877108H. Дои:10.1371 / journal.pone.0077108. ЧВК 3812985. PMID 24204747.
- ^ а б c d е Хендерсон, Д. (2004). "Типичные игроки: пневматичность, плавучесть и водные привычки динозавров-зауроподов". Труды Лондонского королевского общества B. 271: S180 – S183. Дои:10.1098 / rsbl.2003.0136. ЧВК 1810024. PMID 15252977.
- ^ Кермак, К. (1951). «Заметка о повадках зауроподов». Анна. Mag. Nat. Hist. 4 (44): 830–832. Дои:10.1080/00222935108654213.
- ^ «Открытие гигантских болотных зауроподов, сделанных на острове Скай (Wired UK)». Проводная Великобритания. 2015-12-02. Получено 2016-03-22.
- ^ а б c d Майерс, Т.С.; Фиорилло, А. (2009). «Доказательства стадного поведения и возрастной сегрегации у динозавров зауроподов». Палеогеография, палеоклиматология, палеоэкология. 274 (1–2): 96–104. Bibcode:2009ГПП ... 274 ... 96М. Дои:10.1016 / j.palaeo.2009.01.002.
- ^ Кориа, Р.А. (1994). «На моноспецифическом сообществе динозавров-зауроподов из Патагонии: последствия для стадного поведения». GAIA. 10: 209–213.
- ^ Ruxton, Graeme D .; Бирчард, Джеффри Ф .; Диминг, Д. Чарльз (2014). «Время инкубации как важное влияние на яйценоскость и распространение в кладках динозавров зауроподов». Палеобиология. 40 (3): 323–330. Дои:10.1666/13028. S2CID 84437615.
- ^ Осборн, Х. Ф. (1899). "Скелет диплодока, недавно установленный в Американском музее". Наука. 10 (259): 870–4. Bibcode:1899Sci .... 10..870F. Дои:10.1126 / science.10.259.870. PMID 17788971.
- ^ а б Ротшильд, Б. И Мольнар Р. (2005). "Стрессовые переломы зауроподов как ключ к активности". В Карпентер, К. и Тидсуэлл, В. (ред.). Громовые ящерицы: динозавры зауроподоморфа. Издательство Индианского университета. С. 381–391. ISBN 978-0-253-34542-4.
- ^ а б Маллисон, Х. (2009). «Воспитание для еды? Кинетическое / динамическое моделирование двуногих / триподных поз у зауроподовых динозавров». С. 63 в Godefroit, P. and Lambert, O. (eds), Дань Чарльзу Дарвину и Берниссарским игуанодонам: новые взгляды на эволюцию позвоночных и раннемеловые экосистемы. Брюссель.
- ^ Бужор, Мара (2009-05-29). «Зауроподы ходили с вертикальной шеей?». ZME Science.
- ^ Сеймур, RS (июнь 2009 г.). «Поднимать шею зауропода: дороже, чтобы получить меньше». Биол. Латыш. 5 (3): 317–9. Дои:10.1098 / рсбл.2009.0096. ЧВК 2679936. PMID 19364714.
- ^ Сеймур, RS; Lillywhite, HB (сентябрь 2000 г.). «Сердца, положение шеи и интенсивность метаболизма динозавров зауроподов». Proc. Биол. Наука. 267 (1455): 1883–7. Дои:10.1098 / rspb.2000.1225. ЧВК 1690760. PMID 11052540.
- ^ а б Stevens, K.A .; Пэрриш, Дж. М. (1999). «Положение шеи и пищевые привычки двух юрских динозавров зауроподов». Наука. 284 (5415): 798–800. Bibcode:1999Научный ... 284..798С. Дои:10.1126 / science.284.5415.798. PMID 10221910.
- ^ Тейлор, М.П., Ведель, М.Дж., и Найш, Д. (2009). "Положение головы и шеи у динозавров-зауроподов, полученное от современных животных ". Acta Palaeontologica Polonica 54 (2), 2009: 213-220 Абстрактные
- ^ Ученые утверждают, что в музеях и на телевидении совершенно неправильная поза динозавров. Guardian, 27 мая 2009 г.
- ^ Кобли MJ; Rayfield EJ; Барретт PM (14 августа 2013 г.). «Межпозвоночная гибкость шеи страуса: значение для оценки гибкости шеи зауроподов». PLOS ONE. 8 (8): e72187. Bibcode:2013PLoSO ... 872187C. Дои:10.1371 / journal.pone.0072187. ЧВК 3743800. PMID 23967284.
- ^ без даты (14.08.2013). "Страусиные шеи показывают движения зауроподов и пищевые привычки". Science Daily. Получено 2013-08-16.
- ^ Гхош, Тиа (15.08.2013). «Ой! У длинношеих динозавров на самом деле могла быть жесткая шея». NBC News Live Science. Получено 2013-08-16.
- ^ а б Рига, B.J.G .; Кальво, Дж. (2009). «Новый участок следа зауроподов с широкой колеей из позднего мела Мендоса, бассейн Неукен, Аргентина» (PDF). Палеонтология. 52 (3): 631–640. Дои:10.1111 / j.1475-4983.2009.00869.x.
- ^ Castanera, D .; Barco, J. L .; Диас-Мартинес, I .; Gascón, J. S. H .; Pérez-Lorente, F. L .; Канудо, Дж. И. (2011). «Новое свидетельство стада титанозавровидных зауроподов из нижнего берриаса Иберийского хребта (Испания)». Палеогеография, палеоклиматология, палеоэкология. 310 (3–4): 227–237. Bibcode:2011ППП ... 310..227С. Дои:10.1016 / j.palaeo.2011.07.015.
- ^ Day, J.J .; Norman, D.B .; Gale, A.S .; Upchurch, P .; Пауэлл, Г. (2004). «Следы динозавров в Средней юре из Оксфордшира, Великобритания». Палеонтология. 47 (2): 319–348. Дои:10.1111 / j.0031-0239.2004.00366.x.
- ^ Falkingham, P.L .; Бейтс, К. Т .; Margetts, L .; Мэннинг, П. Л. (23 февраля 2011 г.). «Моделирование формирования следа только зауроподов с помощью анализа методом конечных элементов». Письма о биологии. 7 (1): 142–145. Дои:10.1098 / rsbl.2010.0403. ISSN 1744-9561. ЧВК 3030862. PMID 20591856.
- ^ Falkingham, P.L .; Бейтс, К. Т .; Margetts, L .; Мэннинг, П. Л. (07.08.2011). «Эффект« Златовласки »: систематическая ошибка сохранения в скоплениях следов позвоночных». Журнал интерфейса Королевского общества. 8 (61): 1142–1154. Дои:10.1098 / rsif.2010.0634. ISSN 1742-5689. ЧВК 3119880. PMID 21233145.
- ^ а б Продавцы, W. I .; Margetts, L .; Coria, R.A.B .; Мэннинг, П. Л. (2013). Carrier, Дэвид (ред.). "Марш титанов: двигательные способности динозавров зауроподов". PLOS ONE. 8 (10): e78733. Bibcode:2013PLoSO ... 878733S. Дои:10.1371 / journal.pone.0078733. ЧВК 3864407. PMID 24348896.
- ^ Сабо, Джон (2011). Динозавры. Университет Акрона: Макгроу Хилл. п. 35. ISBN 9781121093324.
- ^ Брайан К. Холл; Бенедикт Халльгримссон (1 июня 2011 г.). Эволюция Стрикбергера. Издательство "Джонс и Бартлетт". п. 446. ISBN 978-1-4496-4722-3.
- ^ Sander, P.M .; Mateus, O.V .; Laven, T .; Knötschke, Н. (2006-06-08). «Гистология костей указывает на островную карликовость у нового позднеюрского динозавра зауроподов». Природа. 441 (7094): 739–741. Bibcode:2006Натура 441..739М. Дои:10.1038 / природа04633. PMID 16760975. S2CID 4361820.
- ^ Delair, J.B .; Сарджант, W.A.S. (2002). «Самые ранние открытия динозавров: записи пересмотрены». Труды ассоциации геологов. 113 (3): 185–197. Дои:10.1016 / S0016-7878 (02) 80022-0.
- ^ Lhuyd, E. (1699). Lithophylacii Britannici Ichnographia, sive lapidium aliorumque fossilium Britannicorum singulari figura insignium. Гледич и Вайдманн: Лондон.
- ^ Оуэн, Р. (1842). «Отчет о британских ископаемых рептилиях». Часть II. Отчет Британской ассоциации содействия развитию науки, Плимут, Англия.
- ^ Филлипс, Дж. (1871). Геология Оксфорда и долины Темзы. Оксфорд: Clarendon Press, 523 стр.
- ^ Осборн, Х.Ф., Мук, К.С. (1921). "Камаразавр, Амфикоэлии и другие зауроподы Копы ». Мемуары Американского музея естественной истории, н.с. 3: 247-387 и листы LX-LXXXV.
- Боб Штраус, 2008, Зауроподы: самые большие динозавры, которые когда-либо жили, Нью-Йорк Таймс
- Кристина Карри Роджерс и Джеффри А. Уилсон, 2005, Зауроподы: эволюция и палеобиология, Калифорнийский университет Press, Беркли, ISBN 0-520-24623-3
- Апчерч П., Барретт П.М. и Додсон, П. 2004. Sauropoda. В Динозаврия, 2-е изд. Д. Вайсхампель, П. Додсон и Х. Осмольска (ред.). Калифорнийский университет Press, Беркли. Стр. 259–322.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||












