Диамантиназавр - Diamantinasaurus
| Диамантиназавр | |
|---|---|
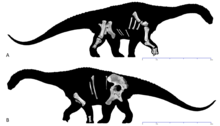 | |
| Голотип скелет на (а) правом и (б) левом видах | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Подотряд: | †Зауроподоморфа |
| Clade: | †Зауропода |
| Clade: | †Титанозаврия |
| Род: | †Диамантиназавр Hocknull и другие. 2009 |
| Разновидность: | †D. matildae |
| Биномиальное имя | |
| †Diamantinasaurus matildae Hocknull и другие. 2009 | |
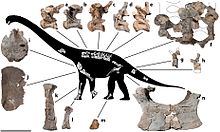
Диамантиназавр это род титанозавр зауропод из Австралия которые жили в первые Поздний мел около 94 миллионов лет назад. В типовой вид рода это D. matildae, впервые описанный и названный в 2009 к Скотт Хокнулл и его коллеги, основанные на ископаемых находках в Формация Винтон. Название означает «ящерица диамантина», название происходит от местонахождения поблизости Река Диамантина и Греческий слово sauros, "ящерица". Специфический эпитет из австралийской песни Waltzing Matilda, также местность голотип и паратип. Известный скелет включает большую часть передняя конечность, плечевой пояс, таз, задняя конечность и ребра голотипа и одной плечевой кости, а радиус и немного позвонки паратипа.
История открытия

В голотип из Диамантиназавр был впервые обнаружен за четыре сезона раскопок около Винтон, Квинсленд, Австралия. Кости, найденные рядом с голотипом Австраловенатор и крокодиломорфы и моллюски.[1] Два найденных динозавра, известные по образцам, занесенным в каталог как AODF 603 и 604 были описаны в 2009 г. Скотт Хокнулл и его коллеги. Образец AODF 603 стал основой для рода Диамантиназавр, а виды D. matildae. Название вида - отсылка к песне «Waltzing Matilda», написанной Банджо Патерсон в Винтоне, а родовое имя происходит от Река Диамантина, проходящая рядом с типовой местностью в сочетании с Греческий sauros, что означает «ящерица». AODF 603, голотип, включает право лопатка, обе плеча, верно локтевая кость, оба неполные Руки, спинной ребра и гастралия, частичное таз, а правая задняя конечность отсутствует.[3] В паратип на том же образце включает спинные и крестцовые позвонки, правая пластина грудины, как сейчас считается, представляет собой остаток коракоидный, а радиус, и одно руководство фаланга. Все эти кости происходят из AODL 85, получившего прозвище «Место Матильды» на Станция Elderslie Sheep, расположенный примерно в 60 км (37 миль) к западу-северо-западу от Винтона в центральной Квинсленд. Этот населенный пункт находится в верхней части средней полосы Формация Винтон, который датируется Сеноманский из Поздний мел.[1][3]
Открытие Диамантиназавр закончилась пауза в открытии новых динозавров в Австралии, названной первым зауроподом за более чем 75 лет. Вместе с Австраловенатор, Диамантиназавр получил прозвище в честь австралийской песни "Waltzing Matilda", Австраловенатор называют "Банджо" и Диамантиназавр по прозвищу «Матильда». Винтонотитан, также с сайта, получил название "Клэнси".[4][5] Эта находка была, по-видимому, крупнейшим открытием динозавров в Австралии, которое было задокументировано со времен Муттабурразавр в 1981 г.[5]
Еще один образец, AODF 836, был описан в 2016 году. Он включает части черепа, включая левый чешуйчатый, почти завершено черепная коробка, верно надугловой, и различные фрагменты. Кроме того, образец также включает атлас, ось, пять других шейных позвонков, три спинных позвонка, дополнительные спинные ребра, части бедра и еще одна правая лопатка.[6]
Описание

Диамантиназавр был относительно небольшим для титанозавра, возможно, достигая 15–16 м (49–52 футов) в длину и 15–20 т (17–22 коротких тонны) в весе. Некоторые из его родственников известны как одержимые доспехи остеодермы хотя неизвестно, был ли Диамантиназавр были эти.[4] Как и другие зауроподы, Диамантиназавр было бы большим четвероногий травоядное животное.[7] С момента первоначального описания единственные существенные изменения включают неправильную идентификацию "грудной пластины", неправильное расположение пальцевых фаланг III-1 и IV-1 как III-1 и V-1 соответственно, а также идентификацию отсутствующей части малоберцовой кости. .[1]
Череп Диамантиназавр изучен не полностью, сохранились только задняя крыша черепа и черепная коробка. Аналогично Сальтазавр и Рапетозавр и в отличие от Немегтозавр, то надвисочное окно граничил с лобная кость. В отличие от обоих последних родов, Диамантиназавр имеет низкий надзатылочный над черепным отверстием, которое впоследствии меньше чем в 1,5 раза превышает высоту базальные трубки (имеющий отверстие). Однако все эти черты присущи Сальтазавр. У производных титанозавров обнаруживается множество других черт, в том числе наклон черепа вниз, боковые отростки головного мозга в форме зубцов, ненарушенная гипофизарная ямка, и более центральное отверстие для внутренняя сонная артерия.[6]

Что характерно для Титанозаврообразные, все шейные и спинные позвонки Диамантиназавр находятся опистоцельный и камелевый (много маленьких внутренних камер). В ось позвонка рода короткая, потенциальная характеристика Saltasauridae. Контрастный Сальтазавр и Рапетозавр Тем не менее презигапофизы из Диамантиназавр простираются перед центром. Только некоторые известные средние спинные кости постспинальная пластинка (гребень на задней поверхности позвоночника) проходит ниже самого позвоночника.[6] Как и другие базальные зауроподы Европазавр и Euhelopus, спинные позвонки имеют выемку в верхней части заднего центра лица, что придает ей вид сердца, контрастируя с более производными титанозаврами или Жирафатитан которые обладают уплощенными центрами. Хотя они отличаются формой центра, Опистоцеликаудия и Диамантиназавр являются единственными титанозаврами, у которых общий брюшной киль находится внутри резко выраженной впадины под спинной костью. Дорсальные презигапофизы связаны с позвоночником спинопрезигапофизная пластинка, которого нет в Опистоцеликаудия и большинство спинных Рапетозавр, а постзигодиапофизарная пластинка нашел в Диамантиназавр также отсутствует у большинства производных титанозавров. Нет никаких указаний на гипосфен-гипантрум, диагностический признак производных титанозавров. Совместно с Опистоцеликаудия, Аламозавр и Лираинозавр за исключением других титанозавров, Диамантиназавр имеет простой нераздельный гребень между задним центром и диапофизом (задняя центродиапофизарная пластинка). Плохо сохранившийся элемент между презигапофизом и центром может быть задняя центропрезигапофизная пластинка, встречается у некоторых брахиозавридов, базальных титанозавров и Опистоцеликаудия. Диамантиназавр обладал по крайней мере пятью, возможно шестью, крестцовые позвонки.[1]
Передняя конечность

Практически вся правая передняя конечность известна по Диамантиназавр, хотя левая плечевая кость известна в дополнение к правой, а левая первая пястная кость известна, в то время как правая не сохранилась. Диагностика Диамантиназавр, то гленоид (humerus) сочленение лопатки повернуто наружу, в отличие от всех других сомфоспондилов. Похожий на Аламозавр и таксонов вокруг основания Titanosauria известен по крайней мере один вентральный отросток, хотя он плохо сохранился. В лопатка из Диамантиназавр прочен, имеет более круглое поперечное сечение, чем другие сомфоспондиланы. Коракоид, ошибочно идентифицированный как грудинный в первоначальном описании, простой и без признаков, контрастирующий Huabeisaurus, Лираинозавр и Опистоцеликаудия. В проксимальный поверхность плечевой кости 1,068 м (3,50 фута) заметно изогнута, как и у титанозавров. Опистоцеликаудия и Сальтазавр. Боковой угол также имеет квадрат, помещая его в Сомфоспондили. Как и у большинства сомфоспондиланов, но в отличие от Euhelopus и Рапетозавр, Диамантиназавр имеет средний сдвиг дельто-грудной гребень. Гребни для прикрепления мышц менее развиты, чем у Опистоцеликаудия и Мадьярозавр. В отличие от производных титанозавров мыщелки, соединяющиеся с предплечьем, не выражены. Диамантиназавр имеет локтевую кость по сравнению с производными титанозаврами по уровню прочности, а также имеет очень выраженный олекранон. Аналогично радиус Диамантиназавр крепче всех титанозавров, кроме Опистоцеликаудия. Локтевая кость имеет длину 70 см (28 дюймов), а радиус - 67,5 см (26,6 дюйма).[1]

Из-за полноты материала передних конечностей отсутствие кости запястья среди сохранившихся материалов предположил Поропат и другие. (2014) быть связано с их истинным отсутствием в жизни, как в Опистоцеликаудия и Аламозавр. Манус Диамантиназавр сравнительно отображает некоторые плезиоморфные черты, в том числе: середину пястный самый длинный (41,2 см (16,2 дюйма) Mc III по сравнению со следующим по длине 37,5 см (14,8 дюйма) Mc II); наличие когтя большого пальца; и наличие множественных фаланг, имеющих формулу фаланг 2–1–1–1–1. Однако манус Диамантиназавр полностью цилиндрический и вертикальный, как и другие титанозавры. Наличие большого количества фаланг у Диамантиназавр использовался Poropat и другие. (2014), чтобы предположить, что у всех титанозавров действительно были окостеневшие фаланги, в отличие от более ранних исследований. Следуя этой логике, они предложили для Опистоцеликаудия и Эпахтозавр, которые оба сохраняют одну фалангу безымянного пальца, отсутствие других было связано с тем, что они были потеряны до окаменения предыдущих пальцев, а не отсутствовали. Полное отсутствие сохранившихся фаланг у Аламозавр, Рапетозавр, Neuquensaurus и Сальтазавр возможно, из-за дезартикуляции, а не из-за отсутствия окостенения.[1]
Задняя конечность

Слева подвздошная кость, Лево и право лобковые волосы, Лево и право искья, и вся правая нога без стопы сохраняется для Диамантиназавр, хотя некоторые кости сильно фрагментированы и плохо сохранились. Внешняя сторона подвздошной кости хорошо сохранилась, но из-за ее размера и хрупкости внутренняя сторона не видна по анатомическим особенностям. Верхний край подвздошной кости сломан, обнажая многочисленные маленькие внутренние камеры, которые есть у титанозавров. Аламозавр, Эпахтозавр, Лираинозавр, Сальтазавр и Сонидозавр. Как и у других производных титанозавров, передний отросток подвздошной кости расширяется в сторону и вращается, так что вертикальное тело подвздошной кости становится горизонтальной полкой. Диамантиназавр также демонстрирует производные черты зауроподов в виде округлой подвздошной кости, уменьшенной суставной поверхности седалищной кости и выпуклости над седалищным сочленением (общая только с Опистоцеликаудия среди Titanosauriformes). Лобковая кость, как и у продвинутых зауроподов, представляет собой уплощенную кость без переднего крючка, как у диплодокоидов, но с потенциально аутапоморфными бороздками, окружающими ее. запирательное отверстие. Сочленение с седалищной костью занимает 46% длины лобка, как у большинства макронариев, но контрастирует с Аламозавр и Опистоцеликаудия, где она уменьшена. Вся седалищная кость составляет всего 68% длины лобка, как у других титанозавров, а также расширяется кнутри, так что все дно таза закрыто. В отличие от некоторых титанозавров седалищная кость Диамантиназавр не отображает ни сужения по ширине, ни выступающих внутрь фланцев. Диамантиназавр также отсутствует заметный мышечный шрам для M. flexor tibialis internus 3 на стороне дистальной седалищной кости, которая является диагностической для таксона среди Neosauropoda.[1]

Бедренная кость длиной 1,345 м (4,41 фута) примерно в два раза шире, чем длина, как у других производных зауроподов, хотя и была слегка раздавлена. Дробление не помешало сохранению linea intermuscularis cranialis гребень, также присутствует в Сальтазавр, Neuquensaurus, Бонатитан, Роказавр и Аламозавр. Как это типично для зауроподов, головка бедренной кости немного выше большой вертел, и есть легкая вертельная полка. Имеется умеренная латеральная выпуклость, выше которой бедренная кость смещена медиально, как у большинства макронариев, за исключением Опистоцеликаудия, Сальтазавр и Рапетозавр. Мыщелки для сочленения с большеберцовая кость и малоберцовая кость высоко распространяется на заднюю поверхность бедра, но в отличие от Neuquensaurus и Опистоцеликаудия не распространяются на переднюю поверхность. Впадина разделяет мыщелок малоберцовой кости, который несет небольшой гребень, также обнаруживаемый в Мадьярозавр и других титанозавров, хотя их известность уникальна для Диамантиназавр. Мыщелок малоберцовой кости больше большеберцовой и простирается дальше вниз, придавая бедренной кости скошенный вид, что потенциально является диагностическим признаком Saltasauridae, но также встречается у Рапетозавр и нетитанозавр Dongbeititan.[1]
Большеберцовая кость составляет 59% длины бедренной кости и, как обычно у новозавропод, шире, чем длина на проксимальной поверхности. Диамантиназавр имеет несколько ямок и гребней на голени, которые не наблюдались у других зауроподов, что делает их набором диагностических признаков. Как и у многих титанозавров, дистальный конец большеберцовой кости расширен до более чем двойной ширины середины диафиза, хотя тонкий фланец вдоль середины диафиза может быть диагностическим признаком Диамантиназавр. Первоначально реконструированная отсутствующая часть диафиза, малоберцовая кость имеет длину 76,9 см (30,3 дюйма) и имеет промежуточную прочность, хотя и близка к изящной. Кость плохо сохранилась, но все же демонстрирует диагностическое расширение рубца малоберцовой мышцы и диагностический медиальный гребень с окружающими бороздками. Как и у многих титанозавров, астрагал из Диамантиназавр менее чем в 1,5 раза шире длины, а проксимальная поверхность делится на восходящий отросток и ямку для голени. Также имеется неглубокая ямка для малоберцовой кости на внешней стороне астрагала, придающая кости субтреугольную форму. На переднем основании восходящего отростка нет впадин или отверстий - состояние, типичное для Eusauropoda. Отросток на задней стороне тела астрагалара уникален среди всех зауроподов, что делает его аутапоморфией Диамантиназавр.[1]
Классификация
Когда это было первоначально описано, Диамантиназавр был приписан к Литостротии incertae sedis. В обеих филогениях он был помещен в, Диамантиназавр был либо просто снаружи Saltasauridae или родственный таксон Опистоцеликаудия внутри семьи.[3] В исследовании 2014 года было обнаружено, что этот род был помещен как литотротий в обе крупные филогении в относительно производное положение в Titanosauria. Их первая филогения была изменена по сравнению с филогенией Карбадилло и Сандера (2014), матрица косвенно основана на филогении Уилсона 2002 года. На этой кладограмме Диамантиназавр оказался сестринским таксоном Тапуиазавр, их отношения за пределами Saltasauridae. В этой филогении поддержка Бремера для каждой группы была не более 1. Пять особенностей скелета поддерживали размещение Диамантиназавр в Литостротии.[1]

| Сомфоспондили |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
В том же исследовании отношения с использованием Mannion и другие. (2013) были протестированы. Эти решены с Диамантиназавр как сальтазаврид, сестра Опистоцеликаудия, с Донъянгозавр как следующий ближайший. Было обнаружено, что два символа поддерживают размещение Диамантиназавр в Lithostrotia, а третий оценить не удалось.[1]
Другой филогенетический анализ 2016 года, частично воспроизведенный ниже, показал, что это нелитостротийский титанозавр и родственный таксон современного Саванназавр.[6][8]

| Титанозаврия |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
Рост
В 2011 году был описан самый маленький положительно идентифицированный эмбрион титанозавра. Хотя это было обнаружено в Монголия, эмбрион разделяет большинство черт с Диамантиназавр и Рапетозавр. Эмбрион из относительно сферического яйца размером 87,07–91,1 миллиметра (3,428–3,587 дюйма) был идентифицирован как сохраняющийся до литохротуса. Эмбрион был немного крепким, промежуточным между устойчивостью Рапетозавр и Диамантиназавр. Яйцо является частью целого места гнездования литотротических титанозавров. Датирование региона также предполагает, что это яйцо предшествовало яйцам из Аука Махуево в Аргентине, и яйца были отложены в Раннемеловой период.[9]
Палеоэкология

Диамантиназавр был найден примерно в 60 км (37 миль) к северо-западу от Винтона, недалеко от станции Элдерсли.[3] Он был извлечен из богатой ископаемыми останками части Формация Винтон, который можно датировать примерно 93 миллионами лет назад.[2] Диамантиназавр был найден в слое глины между песчаник слои, интерпретируемые как Озеро старицы депозит. Также на сайте было найдено Австраловенатор, который был напрямую связан с Диамантиназавр, двустворчатые моллюски, рыбы, черепахи, крокодилы, и различные растения. В свите Винтон был фаунистический комплекс, включающий двустворчатых моллюсков, брюхоногие моллюски, насекомые, то двоякодышащая рыба Метацератодус, черепахи, крокодил Исисфордия, птерозавры, и несколько видов динозавров, таких как вышеупомянутые Австраловенатор, зауроподы Винтонотитан, Саванназавр, и Австрозавр, и безымянный анкилозавры и гипсилофодонты. Диамантиназавр Кости можно отличить от других зауроподов из-за общей прочности, а также множества специфических особенностей. Растения, известные из формации, включают: папоротники, гинкго, голосеменные, и покрытосеменные.[3]
Рекомендации
- ^ а б c d е ж грамм час я j k л Poropat, S.F .; Upchurch, P .; Mannion, P.D .; Hocknull, S.A .; Kear, B.P .; Sloan, T .; Sinapius, G.H.K .; Эллиот, Д.А. (2014). "Ревизия динозавра зауроподов Diamantinasaurus matildae Hocknull et al. 2009 из середины мелового периода Австралии: последствия для гондванского распространения титанозавров ". Исследования Гондваны. 27 (3): 995–1033. Дои:10.1016 / j.gr.2014.03.014.
- ^ а б Tucker, R.T .; Робертс, E.M .; Hu, Y .; Кемп, A.I.S .; Солсбери, С. (2013). «Ограничения возраста детрита циркона для формации Винтон, Квинсленд: контекстуализация фауны динозавров позднего мелового периода Австралии». Исследования Гондваны. 24 (2): 767–779. Bibcode:2013GondR..24..767T. Дои:10.1016 / j.gr.2012.12.009.
- ^ а б c d е Hocknull, Scott A .; Белый, Мэтт А .; Тишлер, Трэвис Р .; Повар, Alex G .; Calleja, Naomi D .; Слоан, Триш; Эллиотт, Дэвид А. (2009). Серено, Пол (ред.). «Новые среднемеловые (новейшие альбские) динозавры из Винтона, Квинсленд, Австралия». PLoS ONE. 4 (7): e6190. Bibcode:2009PLoSO ... 4.6190H. Дои:10.1371 / journal.pone.0006190. ЧВК 2703565. PMID 19584929.
- ^ а б Мюссер, А. (03.06.2010). «Виды животных: Diamantinasaurus matildae". Австралийский музей.
- ^ а б «В Австралии найдены новые динозавры». Новости BBC. 2009-07-03.
- ^ а б c d Poropat, S.F .; Mannion, P.D .; Upchurch, P .; Hocknull, S.A .; Kear, B.P .; Kundrát, M .; Tischler, T.R .; Sloan, T .; Sinapius, G.H.K .; Elliott, J.A .; Эллиотт, Д.А. (2016). «Новые австралийские зауроподы пролили свет на палеобиогеографию динозавров мелового периода». Научные отчеты. 6: 34467. Bibcode:2016НатСР ... 634467П. Дои:10.1038 / srep34467. ЧВК 5072287. PMID 27763598.
- ^ Upchurch, P .; Barrett, P.M .; Додсон, П. (2004). «Зауропода». В Weishampel, David B .; Додсон, Питер; Osmolska, Halszka (ред.). Динозаврия (второе изд.). Калифорнийский университет Press. стр.259 –322. ISBN 978-0-520-24209-8.
- ^ Санкт-Флер, Николас (20 октября 2016 г.). «Встречайте нового титанозавра. Можете называть его Уэйд». Нью-Йорк Таймс. Получено 21 октября 2016.
- ^ Grellet-Tinner, G .; Sim, C.M .; Kim, D.H .; Trimby, P .; Хига, А .; An, S.L .; О, Х.С. Kim, T.J .; Карджилов, Н. (2011). "Описание первого литотротического эмбриона титанозавра in ovo с нейтронными характеристиками и последствиями для миграции и распространения литотротического апта". Исследования Гондваны. 20 (2–3): 621–629. Bibcode:2011GondR..20..621G. Дои:10.1016 / j.gr.2011.02.007.
внешняя ссылка
![]() В Австралии обнаружены три новых динозавра в Викиновости
В Австралии обнаружены три новых динозавра в Викиновости
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||












