Спинофорозавр - Spinophorosaurus
| Спинофорозавр | |
|---|---|
 | |
| 3D-печать реконструкция голотип скелет в Naturkundemuseum Брауншвейг; окаменелости на полу подлинные | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Подотряд: | †Зауроподоморфа |
| Clade: | †Зауропода |
| Clade: | †Грависаурия |
| Род: | †Спинофорозавр Ремес и другие., 2009 |
| Разновидность: | †С. nigerensis |
| Биномиальное имя | |
| †Spinophorosaurus nigerensis Ремес и другие., 2009 | |
Спинофорозавр это род из зауропод динозавр что жил в том, что сейчас Нигер вовремя Средняя юра период. Первые два экземпляра были раскопаны в 2000-х годах немецкими и испанскими командами в сложных условиях. Скелеты были доставлены в Европу и воспроизведены в цифровом виде, что сделало Спинофорозавр первый зауропод, у которого был скелет 3D-печать, и должны были быть возвращены в Нигер в будущем. Вместе эти два экземпляра представляли большую часть скелета рода и один из наиболее известных базальный зауроподы своего времени и места. Первый скелет был изготовлен образец голотипа нового рода и вида Spinophorosaurus nigerensis в 2009; общее название («хребетная ящерица») относится к тому, что изначально считалось шипами остеодермы, а конкретное название (Niger и -ensis) указывает на то, где оно было найдено. Позднее к этому роду был отнесен молодой зауропод из того же района.
Субзарядный голотип, по оценкам, был около 13 м (43 фута) в длину, в то время как паратип был около 14 м (46 футов) в длину. Высота плеч, достигнутая этими людьми, оценивалась примерно в 4 м (13 футов), а вес - примерно в 7 метрических тонн (7,7 коротких тонн). В черепная коробка был коротким, глубоким и широким, а нейроанатомия был в некотором роде промежуточным между базальным зауроподоморфы и более того полученный неозауроподы. Зубы лопатообразные (ложкообразные) с большими промежутками. зубчики в верхней части короны, наследственный признак зауроподов. Шея Спинофорозавр один из самых известных среди зауроподов, содержащий 13 позвонки. Спинные позвонки имели несколько небольших заполненных воздухом внутренних камер, что было характерно для более поздних зауроподов. Хвост имел сильную мускулатуру и имел довольно жесткую заднюю часть из-за длинной и перекрывающейся друг друга. шевронные кости. Кости, которые первоначально считались остеодермами с шипами на кончике хвоста, позже были предложены ключицы.
Спинофорозавр был классифицирован как очень базальный зауропод, или как внутри Eusauropoda, более производная группа. Анатомия, возраст и расположение особей указывают на то, что важные изменения в эволюции зауроподов могли произойти в Северная Африка, возможно, контролируется климатические зоны и посадить биогеография. Особенности вестибулярный аппарат предполагают, что зрение и скоординированные движения глаз, головы и шеи были важны в Спинофорозавр. 3D-модели скелета использовались для проверки его диапазон движения. Одно исследование предполагает, что это могло быть браузер, а другой исследовал возможные позы для спаривания. Швы между нервными дугами с центрами позвонков были более сложными в передней части позвоночника. хобот из Спинофорозавр, поскольку в этом регионе, вероятно, были наибольшие нагрузки. Спинофорозавр известно из Irhazer Shale, а геологическая формация считается среднеюрским по возрасту. Он был образован отложениями из рек и озер в большой речной долинной системе.
Открытие

Богатая фауна динозавров Нигер был доведен до сведения ученых в результате французских и итальянских раскопок в 1960-х и 1970-х годах. Это привело к описанию новых роды из Нижний мел скалы, прежде всего игуанодонт Уранозавр. Более старая последовательность скал, Формация Тиурарен, был исследован американским палеонтологом Пол Серено, который провел широкомасштабную кампанию раскопок в Нигере в период с 1999 по 2003 год. Хотя первоначально считалось, что это образование относится к нижнему меловому периоду, теперь считается, что это образование гораздо более древнего. Средняя юра возраст. Серено назвал новых динозавров, таких как зауропод Jobaria и теропод Афровенатор из Тиурарена; большинство находок было обнаружено у обрыва, известного как Falaise de Tiguidit на юге Регион Агадес. В Марендет, Серено левая частичная Jobaria скелеты в поле как достопримечательность.[2]:11–13[1] Начиная с 2003 г., проект PALDES (Paleontología y Desarrollo - «Палеонтология и развитие») вела раскопки в южном регионе Агадес. Совместные усилия испанской науки и гуманитарных учреждений, PALDES были направлены на объединение палеонтологических исследований с программой развития региона. Это включало улучшение инфраструктуры, образовательных структур и развитие туризма, включая запланированное строительство нового палеонтологического музея в Тадибене.[3]
В начале 2005 года немецкие исследователи Ульрих Йогер и Эдгар Зоммер исследовали полупустыни к югу от Агадеса после местных Туареги сообщил Зоммеру о появлении крупных костей в этом регионе. Соммер - основатель CARGO, организации по оказанию помощи, специализирующейся на улучшении местной системы образования для туарегов, а Джогер - биолог и директор Государственный музей естественной истории, Брауншвейг, Германия. На обратном пути они поболтали с группой туарегов, которые затем направили их в холмистую местность поблизости, усеянную небольшими фрагментами костей. Это место (где раньше не проводились раскопки динозавров) находится примерно в 30 км (19 миль) к северу от Falaise de Tiguidit и недалеко от города Адербиссинат в регионе Агадес и часть Irhazer Shale (или же Argiles de l'Irhazer), а геологическая формация ниже (и, следовательно, немного старше) Tiourarén. После часового поиска Джогер обнаружил округлый костяной наконечник, торчащий из поверхности, который после дальнейших раскопок оказался целым бедренная кость (бедренная кость) того, что позже станет образец голотипа из Спинофорозавр. Связанный лопатка (лопатка) и позвонок были обнаружены вскоре после этого. В осадок в местности твердый, но хрупкий алевролит, можно было удалить с костей легкими ударами молотка.[2]:17–27, 38[1][4]

Затем Джогер и Соммер наняли местных туарегов для поддержки и через два дня обнаружили большую часть образца, который включал практически полностью сочлененный позвоночный столб и несколько костей конечностей и таза. Позвоночный столб образовывал почти полный круг, кончик хвоста находился там, где можно было бы ожидать череп, но не был найден. Не имея оборудования и разрешения на раскопки, они засыпали образец обломками для защиты и вернулись в Германию, где теперь планируют полномасштабные научные раскопки, которые будут проведены музеем Брауншвейга. Официальное разрешение на раскопки было обещано музею в 2006 году Республикой Нигер; Взамен музей должен был построить и оборудовать новую школу для местных детей туарегов в поселке Инджитане. Осенью 2006 года Зоммер и Йогер вместе с другими сотрудниками музея Брауншвейга повторно посетили место в рамках подготовки к раскопкам, поместив одну из тазовых костей в гипс для проверки оборудования и методики. Команда также обнаружила следы тероподов примерно в 1 км от этого места. Спонсоры для финансирования школы и раскопок были найдены в начале 2007 года. Официальная кампания под названием «Projekt Dino» началась 1 марта 2007 года, когда два грузовика с оборудованием выехали из Брауншвейга в Нигер, следуя маршрутом через Испанию, Марокко, Мавританию. , и Мали (более короткий маршрут через Сахара было невозможно из-за риска террористических атак). Другая часть команды, состоящая из десяти постоянных членов, прилетела самолетом.[2]:17–27, 38[1][5][6] Это была первая почти за столетие немецкая экспедиция динозавров в Африку.[1]:29[7]
А пока команда испанского проекта PALDES во главе с Палеонтологический музей Эльче, работал в области. В начале 2007 года Мохамед Эчика, мэр Адербиссината, разрешил команде PALDES раскопать скелет, ранее обнаруженный немцами; впоследствии скелет был отправлен в Испанию. Не подозревая об этих действиях, авангард немецкой команды по прибытии 16 марта обнаружил пустое место раскопок (с признаками профессиональных раскопок); грузовики прибыли 20 марта. Несмотря на разочарование, немецкая команда обнаружила второй Спинофорозавр образец, будущее паратип В 15 м (49 футов) от первого, 17 марта. В исследовательской траншее на территории, усеянной небольшими фрагментами костей, вскоре были обнаружены фрагменты челюсти и зубов; на следующий день ребра, позвонки, плечевая кость (кость плеча) и лопатку. Восемь местных помощников по раскопкам присоединились к группе 19 марта. 20 марта, до прибытия грузовиков, запас пресной воды, первоначально составлявший 200 л (53 галлона США), был исчерпан, поскольку местные помощники использовали ее для мытья накануне вечером, в результате чего члены команды упали в обморок. Раскопки обычно прерывались между 12:00 и 15:00, когда температура достигала 43–45 ° C (109–113 ° F). 25 марта все немецкие члены команды, кроме двух, заболели, страдали понос и проблемы с кровообращением. На протяжении раскопок прогресс документировался фотографиями и полевыми заметками.[2]:29–45[8]

К 27 марта плечевая кость, лопатка и большинство ребер будущего паратипа уже были замотаны защитным пластырем и извлечены. Хотя под скелетом не было видно других костей, команда удалила еще 60–80 см (24–31 дюйм) осадка, чтобы убедиться, что все окаменелости собраны.[2]:60 Раскопки были завершены 2 апреля, и окаменелости были упакованы для транспортировки в порт. Котону 3 апреля. В тот же день Эчика сообщил группе, что первый скелет был раскопан испанской группой с его разрешения. В качестве компенсации он пообещал провести команду к другому месту окаменелости, расположенному примерно в 80 км к югу от Агадеса, на утесе Тигуидит. Там команда нашла заднюю часть возможного Jobaria скелет, но вынужден был оставить самый большой блок на поле до следующего сезона. Чтобы отговорить других собирать блок, был изготовлен взрывной манекен, прикрепленный к окаменелости с предупреждением на испанском языке.[2]:68–73[9] Немецкая команда забрала блок в следующем сезоне 2008 года; команда PALDES отменила свои планы раскопок на тот год после вспышки Восстание туарегов (2007–2009).[2]:100, 107

Два Спинофорозавр образцы были временно размещены в музеях Испании и Германии. По контракту с Республикой Нигер они должны были быть возвращены в страну в будущем под управлением Национальный музей естественной истории в Ниамей а также небольшой, недавно построенный местный музей.[2]:143 Образец будущего паратипа прибыл в Германию 18 марта 2007 г .; Для его подготовки, занявшей два с половиной года, музей Брауншвейга арендовал отдельное здание фабрики. Параллельно с этим совместно работающие немецкие и испанские группы подготовили совместный документ. Немецкая команда оцифровала подготовленные кости и фрагменты обоих образцов в 3D с помощью лазерного сканирования. Поскольку скелет в Брауншвейге был завершен только на 70%, образец в Испании использовался для заполнения недостающих частей; Во время этого процесса было обнаружено, что скелет в Испании был тем скелетом, который немцы первоначально обнаружили и потеряли. 3D-сканы были отремонтированы цифровым способом и деформированы, напечатано в 3D, и собранный в смонтированный скелет для выставки музея Брауншвейга (первый скелет зауропода, воспроизведенный с помощью 3D-печати). Модель жизни в натуральную величину Спинофорозавр, получивший прозвище «Наму» (по названию музея), был поставлен перед главным входом в музей.[2]:79–85[10][11] Испанская команда создала отдельные 3D-модели из фотографий голотипа с использованием фотограмметрия (где снимаются объекты с разных ракурсов для их картирования);[12] хвостовой позвонок был выставлен в музее Эльче в 2018 году.[13] В тезисе конференции 2018 года Гарсиа-Мартинес и его коллеги объявили, что они реконструировали морфологию второго заднего позвонка, который плохо сохранился, на основе лучше сохранившихся первого и пятого задних позвонков. Это было сделано с использованием геометрическая морфометрия на основе ориентиров, где соответствующие 3D-координаты собираются для каждого позвонка и анализируются статистически.[14]
Первый скелет (разделенный между музеем Эльче, где он был каталогизирован как GCP-CV-4229, и музеем Брауншвейга, каталогизированным как NMB-1699-R[15]) был сделан голотипом нового рода и вида Spinophorosaurus nigerensis немецким палеонтологом Кристианом Ремесом и его коллегами из немецкой и испанской групп в 2009 году. Общее название состоит из позвоночник, Латинское слово "шип"; фороПо-гречески «нести»; и sauros, что означает ящерица (полностью "хребетная ящерица"): это имя относится к тому, что изначально интерпретировалось как остеодермы несущие шипы, которые позже предложили представлять ключицы вместо. Конкретное название относится к Нигеру, где таксон был открыт.[16][1][17][18] Образец голотипа состоит из черепная коробка, а заглазничная кость, а чешуйчатый, а квадратный, а крыловидный, а надугловой, и почти полный посткраниальный скелет, в котором отсутствует грудина, antebrachium, manus и pes. Второй, образец паратипа (NMB-1698-R), состоит из частичного черепа и неполного посткраниального скелета. Элементы, сохранившиеся в этом экземпляре, но не голотип, включают предчелюстная кость, верхняя челюсть, слезный, зубной, угловатый, спинные ребра правой стороны, плечевая кость и педальная фаланга. Считается, что эти два экземпляра принадлежат к одному таксону, поскольку элементы скелета, перекрывающиеся между ними, были идентичными, а также из-за близости скелетов в одном стратиграфическом слое.[1] В то время, когда это было описано, Спинофорозавр был наиболее известным среднеюрским зауроподом из Северной Африки,[1] и к 2018 году считался одним из самых полных ранних (не-неозауропод ) зауроподы.[19]
В 2012 году Адриан Парамо и Франсиско Ортега из команды PALDES сообщили о небольшом скелете зауропода (образец GCP-CV-BB-15), который был обнаружен на земле, в нескольких метрах от двух Спинофорозавр образцы; все окаменелости, вероятно, были из одних и тех же пластов. Небольшой скелет состоит из 14 позвонков (некоторые из них сочлененные), включая все шейные позвонки, а также некоторые позвонки задней части. Центры его позвонков на 20% меньше, чем у него. Спинофорозавр и нейроцентральный шов открыт, что указывает на то, что это несовершеннолетний. Несколько отличительных черт скелета разделяют Спинофорозавр, и хотя некоторые признаки этого рода отсутствуют, он, скорее всего, представляет собой молодь Спинофорозавр (различия, вероятно, объясняются онтогенез, меняется в процессе роста).[20][15]
Описание

Образец голотипа был около 13 метров (43 фута) в длину при измерении вдоль позвоночного столба, в то время как паратип был примерно на 13 процентов больше, размером около 14 метров (46 футов).[1][2] Трехмерная фотограмметрическая модель скелета голотипа имеет размеры 11,7 м (38 футов) от головы до хвоста, пропорции отличаются от оценок, основанных на двухмерных реконструкциях скелета.[12] Высота плеч, достигнутая этими людьми, оценивалась примерно в 4 м (13 футов),[2] и вес около 7 метрических тонн (7,7 коротких тонн).[1][21] Эндокраниальная и нейроцентральная части образца голотипа. швы в черепе и позвонках, соответственно, не слиты, что указывает на то, что это был суб-взрослый, тогда как образец паратипа имеет полностью сросшиеся нейроцентральные швы.[1]
В 2020 году Видаль и его коллеги пересмотрели позицию Спинофорозавр, основанный на установке цифрового каркаса. В то время как оригинальная реконструкция скелета 2009 года показала динозавра с горизонтальной позой, цифровая реконструкция показала более вертикальную осанку с высокими плечами и поднятой шеей.[22]
Череп

Лобные кости крыша черепа были сращены по средней линии, в отличие от других швов черепа. Между лобными частями был небольшой шишковидное отверстиепримерно в 10 мм (0,4 дюйма) перед швом между лобной и теменной костями. На заднем крае черепа была открытая пост теменная выемка - особенность, известная только в дикреозавриды и Аброзавр. Вырез, следующий по средней линии в передней части соединенных лобных костей, указывает на то, что, возможно, был зубец, проходящий от каждой носовой кости между лобными костью, как это могло быть в случае Нигерзавр но необычен среди зауропод в целом. Верхние височные оконные проемы, пара крупных отверстий на крыше черепа, были в четыре раза шире, чем длинные; такие расширенные оконные проемы были характерны для Шунозавр и больше полученный (или «продвинутые») зауроподы. В затылочный мыщелок задняя часть черепа была вогнутой по бокам, как у Шунозавр. В базальная тубера (пара расширений на нижней стороне основания черепа, которые служили прикреплениями мышц) были увеличены и направлены в стороны, что является уникальным среди известных зауроподов. Квадрат не имел вогнутости на задней стороне, наследственный условие иначе известно только в Тазудазавр среди зауроподов. Лопатчатые (в форме ложки) зубы были уникальны тем, что обладали увеличенными зубчики вокруг вершины коронки, с большим количеством зубчиков на переднем крае коронки.[1][23]
Мозговой Спинофорозавр был широким и коротким спереди назад, умеренно глубоким и в целом относительно большим. Он значительно отличался от таковых других юрских зауроподов, возможно, кроме Атласавр. Они разделили базиптеригоидный отросток на их базисфеноиды которые были похожи в том, что были направлены строго назад. ДЕЙСТВОВАТЬ эндокаст полости мозга голотипа показали, что он был похож на большинство зауроподов тем, что имел хорошо выраженные изгибы моста и головного мозга, гипофизарную ямку, которая была большой и продолговатой, а структура мозга была скрыта пространствами, в которых располагались относительно толстые мозговые оболочки а также дуральные венозные синусы. В лабиринт из внутреннее ухо было характерно иметь длинные и стройные полукружные каналы, аналогичные тем из Массоспондил и Жирафатитан. Его нейроанатомия был в некотором роде промежуточным между базальный (или «примитивные») зауроподоморфы и более производные неозауроподы.[23]
Позвонки и ребра

Позвоночный столб почти полностью известен, а голотип - один из немногих экземпляров зауроподов, у которых есть полная шея.[24] Шея состояла из 13 шейные позвонки. В багажнике было 12 спинной и четыре крестцовые позвонки. Хвост насчитывал более 37 хвостовые позвонки.[1] Сложные элементы, отдельные позвонки состоят из нижней части, центр, а верхняя часть - нервная дуга. Важные ориентиры нервной дуги включают выступающую вверх нервный позвоночник (остистый отросток) и выступающий вбок диапофизы, которые вместе придают позвонку Т-образную форму на виде спереди и сзади. Пара суставные отростки соединяющиеся с соседними позвонками выступают спереди (презигапофизы) и задний (постзигапофизы).[25]
Шейные позвонки были похожи на позвонки Jobaria и Цетиозавр. Их центральная часть была примерно в 3,1 раза длиннее ширины; поэтому они были умеренно удлиненными по сравнению с зауроподами в целом, но в целом длиннее, чем у других базальных форм. В шейных центрах были большие выемки по бокам, углублявшиеся к передней части; такой плевроцели были также разработаны в Jobaria и Патагозавр. В отличие от Jobariaплевроцели не были разделены косым костным гребнем. Срединный киль имелся на нижней стороне переднего конца центра, который отсутствовал у Цетиозавр. Концы презигапофизов имели треугольное удлинение, которое также видно на Jobaria, хотя он глубже в этом роде. Выше постзигапофизы были сравнительно большими. эпипофизы, костные выступы для прикрепления мышц. Шейные позвонки отличались от базальных зауроподов из Южной Америки и Индии. Диапофизы (боковые отростки нервной дуги) были наклонены немного вниз и имели треугольные выступы на задних краях - черты, невидимые у этих южных форм. Кроме того, нервные отростки были морщинистыми (морщинистыми) на их задней и передней поверхностях и близко к основанию шеи, более широкими при виде сбоку и менее высокими. На виде сбоку между центром и нервной дугой имелось U-образное углубление, которое является аутапоморфия (уникальная особенность) Спинофорозавр.[1]

Спинные позвонки имели необычную камелистую внутреннюю структуру (содержали множество небольших заполненных воздухом камер). Эта особенность также известна в гораздо более поздних титанозавры а также в маменчизавриды, где это развились независимо из бывшей группы.[26] Хотя передние спинные позвонки имели глубокие плевроцели в центре, эти отверстия становились намного мельче по направлению к задней части туловища. Самые задние спинные позвонки также были пропорционально короткими. В Амигдалодон и Патагозавр напротив, задние спинные кости были более удлиненными и имели ярко выраженные плевроцели. Нервный канал спинных позвонков очень узкий, но высокий.[1] Гипосфен-гипантрум сочленения (добавочные суставные отростки) присутствовали во всех дорсальных отделах, делая позвоночник более жестким.[1][27] Нервные отростки имели заметные морщинистости на передней и задней сторонах, как и у других базальных зауроподов. Крайние передние хвостовые позвонки имели такую же морщинистость на нервных отростках, как и спинные позвонки, - особенность, известная только в Омейзавр. В задней части хвоста нервные отростки были сильно отклонены назад и простирались на переднюю часть последующего позвонка, как у некоторых восточноазиатских зауроподов. Барапазавр, и Jobaria.[1]
Ребра со второго по пятый спинные позвонки были уплощены и направлены назад, тогда как ребра с шестого по одиннадцатый спинных позвонков были более округлыми в поперечном сечении и более вертикально ориентированными. Таким образом, грудную клетку можно четко разделить на грудную и поясничную части; такая дифференциация была описана только у одного другого зауропод, дикреозаврида. Брахитрахелопан. Кроме того, на концах грудных ребер были места прикрепления грудных ребер, которые соединялись с грудиной. В передней части хвоста шевроны (парный кости ниже центра позвонков) были пластинчатыми, что является основным условием. В задней части хвоста шевроны были стержневидными, а левый и правый экземпляры отделены друг от друга. Эти стержневидные шевроны плотно прилегали к нижним краям центров. Они сочленялись с предыдущими и последующими шевронами на средней длине позвонков, укрепляя позвоночный сустав и ограничивая сгибание хвоста.[1]
Пояса, конечности и неправильно идентифицированные шипы на хвосте

Лопатка была уникальной среди зауроподов: она сильно изогнута с расширенным веерообразным нижним концом. В поперечном сечении он имел D-образную форму, характерную для эвзауроподов. Его верхний конец был расширен, а на нижнем крае имелся костяной выступ.[19] В этом отношении он был похож на маменчизаврид из Азии, но отличался от гондванских форм. Вулканодон, Барапазавр, и Патагозавр, у которого верхний торец был лишь слабо уширен, а задний фланец отсутствовал. В коракоидный, который сочленяется с нижним концом лопатки, имел отчетливую форму почки, считающуюся аутапоморфией. У него был большой бугорок двуглавой мышцы, к которому двуглавая мышца плеча прикреплена мышца. Ключица была крепкой, хотя и более тонкой, чем у Jobaria. Из передней конечности сохранилась только плечевая кость. Ее нижний конец был асимметричным и имел увеличенные добавочные мыщелки (направленные вперед выступы на нижнем переднем крае кости) - черты, которые в остальном наблюдаются только у маменчисаврид. Лобковая кость и седалищная кость таза крепкие, последний на конце расширен. Верхняя бедра (бедренная кость) характеризовалась наличием малый вертел на верхнем конце - костный выступ, служащий местом прикрепления мышц, тянущих заднюю ногу вперед и внутрь. В четвертый вертел, который выступал с поверхности спины и закреплял мышцы, тянущие заднюю ногу назад, был особенно большим в Спинофорозавр. Близко к четвертому вертлугу было большое отверстие, которое отсутствует у других зауроподов, и, следовательно, это аутапоморфия. Большеберцовая кость (большеберцовая кость) была подобна таковой у других базальных зауроподов, а малоберцовая кость (кость голени) была крепкой. Лодыжки, верхняя часть астрагал имел фасетки для сочленения с большеберцовой и малоберцовой костью, не разделенные костной стенкой, и целых восемь питательное отверстие (отверстия, позволяющие кровеносным сосудам проникать в кость).[1]

Элементы, первоначально интерпретированные как левая и правая остеодерма (кость, образованная в коже), были обнаружены вместе с голотипом скелета. Эти кости имели округлое основание, из которого торчал шиповидный выступ; внутренние поверхности были морщинистыми и вогнутыми. Хотя они были обнаружены в области таза, Ремес и его коллеги считали, что они расположены на кончике хвоста у живого животного, что они считали отличительной чертой этого рода. Это положение было основано на том факте, что левый и правый элементы были обнаружены близко друг к другу, что позволяет предположить, что они пришли из средней линии тела. Кроме того, усиление задней части хвоста удлиненными шевронами наблюдается и у других динозавров с булавами или шипами на хвосте. Подобные шипы были частью хвостовой булавы у родственных зауроподов. Шунозавр; такой хвостовой булавы, вероятно, не было в Спинофорозавр, так как задние хвостовые позвонки стали слишком маленькими. Предполагаемая правая остеодерма была несколько больше левой и немного отличалась по форме. Это указывает на то, что они не образовали пару; в этом случае они, вероятно, будут просто зеркально перевернутыми копиями друг друга. Скорее, эти различия указывают на то, что изначально присутствовали две пары шипов.[1]
В 2013 году палеонтологи Эмануэль Чопп и Октавио Матеус повторно исследовали предполагаемые шипы на хвосте и обнаружили, что они не имеют типичной морщинистой поверхности остеодермы, наблюдаемой у других бронированных динозавров, или клубного расширения, наблюдаемого у Шунозавр. Из-за их сломанных краев они также сомневались, были ли эти элементы разного размера, как первоначально предполагалось. Поскольку эти элементы были обнаружены под лопаткой, они вместо этого предположили, что они представляют ключицы, и что ископаемые останки должны быть пересмотрены в этом отношении.[18] Видаль и его коллеги согласились с этим предложением в 2015 и 2018 годах.[28][19]
Классификация


Начальный филогенетический анализ представленный Ремесом и коллегами предложил Спинофорозавр попал в число самых основных известных зауроподов, будучи лишь немногим более производным, чем Вулканодон, Цетиозавр, и Тазудазавр. В этом положении он сформировал бы сестринский таксон из Eusauropoda, клада, включающая всех более производных зауроподов. Авторы признали, что поддержка этого самого базального положения была слабой, и обсудили несколько альтернативных размещений внутри эусавропод, которые могли бы объяснить анатомическое сходство с другими зауроподами из Северной Африки и Лавразии.[1] Подобное базальное положение вне Eusauropoda было предложено несколькими последующими исследованиями,[29][30][31] который разместил Спинофорозавр как родственный таксон Тазудазавр[29] или же Волхеймера.[30] В тезисе конференции 2013 года палеонтолог Педро Мочо и его коллеги повторно оценили филогенетические отношения рода, включив дополнительную информацию из недавно подготовленных костей, утверждая, что Спинофорозавр гнездился внутри эусавропод. Согласно этому анализу, род был более производным, чем Шунозавр и Барапазавр и близко к Патагозавр и маменчизавриды.[26] Гораздо более систематическое положение внутри Eusauropoda было также предложено в исследовании 2015 г. Спинофорозавр быть сестринским таксоном Небуласавр.[32]
Кладограмма основано на Nair and Salisbury, 2012, который поддерживает базальное положение Спинофорозавр:[29]
| Кладограмма основана на материалах Xing et al., 2015, которые подтверждают более сложную позицию внутри Eusauropoda:[32]
|
Эволюция
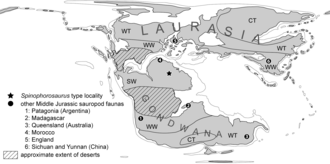
Как один из наиболее известных базальных зауроподов, Спинофорозавр помог пролить свет на раннюю эволюцию и палеобиогеография группы. Это неясно из-за редкости останков ранней и средней юры, особенно за пределами Азии. Ремес и его коллеги обнаружили, что Спинофорозавр имеет общие черты со среднеюрскими восточноазиатскими зауроподами (особенно в шейных и хвостовых позвонках, лопатке и плечевой кости), но очень отличается от южноамериканских и индийских таксонов нижней и средней юры (различия включают форму и развитие особенностей позвонков и форму лопатки). и плечевая кость). Они предположили, что это может быть объяснено среднеюрской фауной зауроподов Лавразия (северный суперконтинент того времени) и юг Гондвана (южный суперконтинент), разделенный географическими преградами. Ранее считалось, что зауроподы были распространены по суперконтиненту. Пангея (который состоял из Лавразии и Гондваны) в период ранней и ранней средней юры. У него было относительно небольшое разнообразие, пока континент распался и привел к тому, что группы зауроподов развились изолированно через викарианс в период поздней средней и поздней юры. Спинофорозавр показывает, что черты, которые когда-то считались унаследованными от зауроподов Восточной Азии, довольно плезиоморфный (наследственные черты) среди эусауропод (эусауроподы, колонизировавшие Лавразию, сохранили базальные черты, также наблюдаемые в Спинофорозавр). Эусавроподы, колонизировавшие Южную Гондвану, были особой линией группы, утратившей указанные предковые черты во время изоляции. Ремес и его коллеги отметили, что необходимо больше доказательств, чтобы поддержать эти интерпретации, но были уверены, что существует связь между юрскими зауроподами в Северной Африке, Европе и Восточной Азии.[1]
На что указывает анатомия Спинофорозавр и характер распределения зауропод в средней юре, важные изменения в эволюции зауроподов могли произойти в Северной Африке. Это было близко к экватор и имел влажный летом климат в ранней и средней юре, с высокой продуктивностью и разнообразием растений. С другой стороны, зауроподы Южной Гондваны были изолированы от экваториального региона Центральной пустыней Гондваны в ранней юре, что привело к различиям во флор Южной и Северной Гондваны. Когда пустыня сократилась в конце средней юры, появились неозауроподы, которые заменили фауну зауроподов, которая была типичной для средней юры. С Jobaria Северной Африки близко к основанию Neosauropoda, и потому что неозауроподы, такие как диплодокоиды были широко распространены в верхней юре, неозауроподы и некоторые из их подгрупп, возможно, также возникли в экваториальной области юрской Пангеи, которая, возможно, сыграла роль «горячей точки» в эволюции зауроподов. Диверсификация зауропод в юрском периоде, возможно, контролировалась климатические зоны и биогеография растений, а не просто континентальная дифференциация.[1]
Палеобиология

Спинофорозавр и некоторые другие зауроподоморфы не уменьшили вестибулярные аппараты, сенсорная система для баланса и ориентации во внутреннем ухе, хотя этого можно было ожидать от линии, которая привела к появлению тяжелых четвероногих, питающихся растениями. Неизвестно почему Спинофорозавр сохранили эту особенность, но размер и морфология лабиринтов зауроподоморфа могут быть связаны, например, с длиной и подвижностью шеи. Возможно, расширение вестибулярного аппарата является показателем важности зрения и скоординированных движений глаз, головы и шеи, хотя интерпретация вестибулярных особенностей зауроподов все еще остается неопределенной.[23]
На конференции 2018 года Бенджамин Йентген-Чешино и его коллеги сообщили о радиальной фиброламеллярной кости (RFB), типе костной ткани, характеризующейся радиально ориентированными каналами, в самой внешней части кора (внешний костный слой) плечевой кости взрослого Спинофорозавр образец и молодой человек Исанозавр. Обычно кора длинных костей зауроподов вместо этого показывает фиброламеллярный комплекс. RFB считается быстрорастущей костной тканью, о которой ранее не сообщалось у зауроподов. Его присутствие в самой внешней коре головного мозга необычно, как обычно ожидается в самой внутренней коре (что соответствует ювенильному росту). РФБ Спинофорозавр был похоронен во внешней коре головного мозга, за которым последовал нормальный фиброламеллярный комплекс, что указывает на то, что животное выжило в течение некоторого времени после его ускоренного роста, что, вероятно, предполагает патологический рост костей из-за травмы.[33]
Движение

С Спинофорозавр является одним из наиболее известных базальных зауроподов, он является хорошей моделью для биомеханический исследования, которые помогают понять биологию зауроподов и функцию анатомических особенностей. Манипуляции с 3D-моделью, выполненные Видалем и его коллегами в 2015 году, показали, что каждый из 20 передних позвонков хвоста может отклоняться на 20º и 8–10º вбок. сгибание, раньше ограничивался морфологией кости. Фактический диапазон движений у живого животного был бы меньше, поскольку мягкие ткани, а также большие шевронные кости были бы ограничивающим фактором. Первые 20 хвостовых позвонков также стали длиннее к концу хвоста, как это было у диплодоцидов, и были задействованы за счет увеличенной гипаксиальной мускулатуры, на что указывают большие шевронные кости и направленные вверх поперечные отростки в первых хвостовых позвонках. Движение в задней части хвоста было ограничено, так как шевроны перекрывали друг друга, как и в случае дромеозавриды и анкилозавриды. Вместе эти особенности хвоста могли быть частью специальной функции, которая неизвестна.[28] В 2009 году Джогер и его коллеги утверждали, что предполагаемые всплески формируют тагомизатор на конце хвоста, который использовался для защиты от хищников.[2]
В 2017 году Джон Фронимос и Джеффри Уилсон использовали Спинофорозавр в качестве модели для изучения сложности нейроцентральных швов (жестких соединение соединение нервной дуги позвонка с его центром) у зауроподов, возможно, способствовало укреплению позвоночника. Зауроподы достигали зачастую огромных размеров тела как в результате быстрого, так и продолжительного роста. Пока человек рос, нейроцентральные швы были состоит из хряща чтобы разрешить этот рост. Достигнув полного размера тела, этот хрящ будет превратиться в кость, закрывая шов. Однако хрящ гораздо менее устойчив к силам, которые потенциально могут привести к вывиху нервной дуги, таким как изгиб и кручение; они становятся особенно актуальными при больших размерах тела. Зауроподы и другие архозавриформы противодействовать этой структурной слабости за счет увеличения сложности шва, что означает, что поверхности, которые соединяют нервную дугу с ее центром, имеют сложные гребни и борозды, которые сцепляются друг с другом. В Спинофорозавр, сложность швов была наиболее выражена в передней части туловища, что указывает на то, что напряжения были самыми высокими в этой области, вероятно, из-за веса длинной шеи и грудной клетки. Сложность стала меньше по отношению к черепу и крестцу. Ориентация гребней позволяет определить тип напряжения, которое повлияло на позвонок: в шейных позвонках гребни в основном препятствовали смещению нервной дуги в направлении спереди назад, в то время как гребни в позвонках туловища были больше эффективно препятствует вращению.[34]
В тезисе конференции 2018 года Видаль использовал виртуальную Спинофорозавр скелет, чтобы проверить гипотетические позы спаривания, которые были предложены для зауроподов, которые будут включать "клоакальный поцелуй "(как это делают большинство птиц), а не самец интромитентный орган. К таким гипотезам относятся «нога через спину», самец садится сзади (самка стоит или лежит), а также «спаривание назад». Позы, приводившие к вывиху или остеологической остановке, считались неприемлемыми. Поза «нога через спину» была отвергнута, поскольку стоящему самцу было бы невозможно дотянуться ногой до спины и бедра сидящей самки, даже если она была вдвое меньше самца (и неизвестно, были ли зауроподы мы сексуально диморфный по размеру). Поза, в которой самец садится на самку сзади, опираясь передними ногами на спину самки, была возможна (хвост был достаточно гибким, чтобы убираться с дороги), но даже если самка лежала, клоаки не могли подходят друг к другу достаточно близко для «клоакального поцелуя». «Клоакальный поцелуй» был бы возможен только через «спаривание назад», когда самец и самка подходили друг к другу задом и соединили свои клоаки с согнутыми в стороны хвостами, а движение ног позволило бы особям разных размеров спариваться таким образом. «Спаривание в обратном направлении» - единственный способ спаривания зауроподов без потребности в интромитентном органе самца. Такая поза распространена при спаривании. членистоногие и в "связанный "-фаза спаривания карниворан млекопитающие, такие как Canis.[35]

В 2020 году Видаль и его коллеги использовали 3D-модели как голотипа, так и ювенильного скелета, чтобы оценить диапазон движений (гибкость) шеи. Такие оценки предполагают, что исходное расстояние между позвонками можно надежно спрогнозировать и что суставные отростки все время находятся в контакте. Видаль и его коллеги продемонстрировали, что оба предположения действительно верны в современных условиях. жирафы, повышая уверенность в оценке диапазона движений вымерших животных в целом. В качестве Спинофорозавр росла, амплитуда движений шеи увеличивалась (делая возможным более крутое положение шеи); промежуток между позвонками становится больше; в нейтральной позе шея становится более наклонной. Подобные изменения в процессе роста наблюдаются и у жирафов. Шея была бы такой же гибкой, как у жирафа, благодаря большему количеству позвонков, хотя отдельные суставы были гораздо менее гибкими, чем у жирафов. Спинофорозавр мог бы питаться в тех же позах, что и жирафы, и это мог быть самый базальный зауропод, адаптированный для высокая посещаемость. Хороший обзор также подтверждается анатомическими особенностями, включая узкую морду, широкие зубы и пропорциональную длину плечевой кости по сравнению с лопаткой. Как у жирафа, как молодняк, так и взрослый Спинофорозавр человек не смог бы добраться до земли, просто опустив шею, и, возможно, расправил бы передние конечности для питья. Во время сна жирафы наклоняют к туловищу шеи набок. Хотя суставы позвонков были бы достаточно гибкими для такого изгиба в Спинофорозавр, это могло быть запрещено удлиненными шейными ребрами.[15]
Палеоокружающая среда

Спинофорозавр известен из сланцев Ирхазер в Нигере, которые, как было установлено, представляют основу Irhazer Group (на основании наличия Красная глина вот как перемежающийся песчаные пласты со следами субаэрального обнажения и следы динозавров ), который стратиграфически ниже обнажений Тегама Групп. Группа Irhazer ранее считалась от юрского до самого раннего мелового периода, поскольку вышележащая формация Tiourarén когда-то считалась относящейся к нижнему меловому периоду. Последующие исследования показали, что последняя формация относится к периоду от средней юры до начала поздней юры. Стратиграфическое и филогенетическое положение Спинофорозавр (по сравнению с другими базальными зауроподами со всего мира) указывает на то, что это может быть средняя юра (Байосский –Батонский ) в возрасте. Поскольку пласты ирхазерской серии еще не могут быть напрямую датированы, они могут быть такими же древними, как нижняя юра. Нижний предел определяется базовым Группа Агадес, который Верхний триас по возрасту (исходя из наличия Хиротерий следы окаменелостей ).[1][36]
В Спинофорозавр скелеты были обнаружены в массивном или тонкослоистом красном алевролите, чьи матрица содержал немного карбоната. Окаменелости были извлечены из верхней половины этого слоя алевролита толщиной несколько метров. Слои там субгоризонтальные, несут незначительные разломы и обычно содержат палеопочвы и карбонат в верхней части массива (примерно на метр выше уровня скелетов).[1] Отложения ирхазерской серии отлагались в условиях, которые были Fluvio -озерный (связанных с реками и озерами), в то время, когда Бассейн Юллеммеден была частью большой речной долинной системы, которая была связана с Протоатлантический побережье Гондваны через прилегающие бассейны.[36][37]

Было обнаружено, что четыре зуба теропод тесно связаны с Спинофорозавр голотип (по позвонку, лобку и вертлужной впадине); три имели сходство с Megalosauridae и Аллозавриды, в то время как четвертый принадлежит тому, что может быть одним из самых ранних известных членов Спинозавры. Дело в том, что зубы плохо сохранились, а Спинофорозавр Скелет хорошо сохранился, это указывает на то, что зубы были перевезены до захоронения с зауроподом.[38] Следы динозавров были найдены в сотне метров от Спинофорозавр участок, в том числе путь из шести следов от зауропода среднего размера, а также 120 хорошо сохранившихся дидактиль (двупалые) следы теропод. Первоначально считалось, что дидактильные следы были оставлены паравиан теропод (например, дромеозавры или троодонтиды, у которых есть два опорных пальца), но впоследствии было интерпретировано, что они были созданы плавающими тероподами (что объясняет, почему один палец не оставил следов).[36][39]
Рекомендации
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у Ремес, К .; Ортега, Ф .; Fierro, I .; Joger, U .; Kosma, R .; Marín Ferrer, J.M .; Ide, O.A.u; Мага, А .; Фарке, А. А. (2009). «Новый базальный динозавр зауроподов из средней юры Нигера и ранняя эволюция зауроподов». PLOS ONE. 4 (9): e6924. Bibcode:2009PLoSO ... 4.6924R. Дои:10.1371 / journal.pone.0006924. ЧВК 2737122. PMID 19756139.
- ^ а б c d е ж грамм час я j k л Joger, U .; Kosma, R .; Крюгер, Ф. Дж. (2009). Projekt Dino: Die Entdeckungsgeschichte neuer Dinosaurier в Нигере, Африка (на немецком). Брауншвейг: Cargo Verlag. ISBN 978-3-938693-17-9.
- ^ Ортега, Ф .; Fierro, I .; Chiappe, L .; и другие. (2009). Проект Палдес и наследие палеонтологии позвоночных в стране Нигер. Первый Международный конгресс по палеонтологии позвоночных Северной Африки (NAVEP1). 25–27 мая 2009 г., Марракеш - Марокко. С. 49–51.
- ^ Джогер, У. «Место открытия и раскопок 2005 г.». Google Искусство и культура. В архиве с оригинала 20 апреля 2019 г.. Получено 20 апреля 2019.
- ^ Джогер, У. «Место открытия следов». Google Искусство и культура. В архиве с оригинала 20 апреля 2019 г.. Получено 20 апреля 2019.
- ^ Джогер, У. «Экскурсионный тур 2007 - Начало пути». Google Искусство и культура. В архиве с оригинала 20 апреля 2019 г.. Получено 20 апреля 2019.
- ^ Джогер, У. (2018). "БРАУНШВЕЙГ: Государственный историко-исторический музей Брауншвейга". В Beck, L.A. (ред.). Зоологические коллекции Германии. Коллекции естествознания. Springer. С. 183–196. Дои:10.1007/978-3-319-44321-8_15. ISBN 978-3-319-44321-8.
- ^ Косма, Р. "О нет, а где наш динозавр?". Google Искусство и культура. В архиве с оригинала 20 апреля 2019 г.. Получено 20 апреля 2019.
- ^ Witzmann, F .; Hampe, O .; Ротшильд, Б. М .; Joger, U .; Kosma, R .; Schwarz, D .; Асбах, П. (2016). «Субхондральные кисты в суставах синовиальных позвонков как аналогии узлов Шморля у динозавра-завропода из Нигера». Журнал палеонтологии позвоночных. 36 (2): e1080719. Дои:10.1080/02724634.2016.1080719. S2CID 86187185.
- ^ Косма, Р. (2018). «Палеонтологическая коллекция Государственного музея естественной истории (SNHM) в Брауншвейге, Германия». In Beck, L.A .; Джогер, У. (ред.). Палеонтологические коллекции Германии, Австрии и Швейцарии. Коллекции естествознания. Springer. С. 81–92. Дои:10.1007/978-3-319-77401-5_9. ISBN 978-3-319-77401-5.
- ^ Риттер, А. «Реконструкция скелета динозавра». Google Искусство и культура. В архиве с оригинала 20 апреля 2019 г.. Получено 20 апреля 2019.
- ^ а б Vidal, D .; Aberasturi, A .; Mocho, P .; Ортега, Ф .; Санс, Дж. Л. (2016). "Сборка виртуального Спинофорозавр скелет: чему он может научить нас об эволюции эусавропод? ". VII Jornadas Internaciones sobre Paleontología de Dinosaurios y su Entorno. Салас-де-лос-Инфантес, Бургос. Бургос. С. 147–148.
- ^ "El MUPE expone por primera vez una vértebra del cuello de Spinophorosaurus nigerensis". AQU en Elche (на испанском). 2018 г.. Получено 15 июн 2019.
- ^ Гарсиа-Мартинес, Д .; Vidal, D .; Ортега, Ф. (2018). Использование трехмерной геометрической морфометрии для оценки отсутствующих позвонков в Спинофорозавр спинной отдел позвоночника (средняя юра, Нигер). 16-е заседание Европейской ассоциации палеонтологов позвоночных, Капарика (Португалия). Европейская ассоциация палеонтологов позвоночных. п. 75.
- ^ а б c Видаль, Даниэль; Мочо, Педро; Парамо, Адриан; Санс, Хосе Луис; Ортега, Франциско (13 января 2020 г.). «Онтогенетическое сходство между остеологической подвижностью шеи жирафа и зауроподов». PLOS ONE. 15 (1): –0227537. Bibcode:2020PLoSO..1527537V. Дои:10.1371 / journal.pone.0227537. ISSN 1932-6203. ЧВК 6957182. PMID 31929581. Получено 2020-01-13.
- ^ Тиммонс, Дж. (2018). «Открытия окаменелостей в Нигере с доктором Ральфом Космой». В основном мамонты, мумии и музеи. В архиве из оригинала 2 марта 2019 г.. Получено 24 февраля 2019.
- ^ Хольц, Т. Р. (2011). «Динозавры: Самая полная и современная энциклопедия для любителей динозавров всех возрастов, зимнее приложение 2011 года» (PDF). Получено 13 января 2012.
- ^ а б Tschopp, E .; Матеус, О. (2013). «Ключицы, межключицы, гастралия и грудинные ребра у зауроподовых динозавров: новые сообщения от Diplodocidae и их морфологические, функциональные и эволюционные последствия». Журнал анатомии. 222 (3): 321–340. Дои:10.1111 / joa.12012. ЧВК 3582252. PMID 23190365.
- ^ а б c Mocho, P .; Vidal, D .; Aberasturi, A .; Kosma, R .; Ортега, Ф. (2018). Новые сведения об аппендикулярном скелете Spinophorosaurus nigerensis (Средняя юра, Нигер). 16-е заседание Европейской ассоциации палеонтологов позвоночных, Капарика (Португалия). Европейская ассоциация палеонтологов позвоночных. п. 124.
- ^ Blázquez, A.P .; Ортега, Ф. (2012). Вероятный несовершеннолетний Spinophorosaurus nigerensis (Sauropoda) из средней юры Нигера. 10-е заседание Европейской ассоциации палеонтологов позвоночных в Теруэле (Испания).
- ^ Пол, Г.С. (2016). Принстонский полевой путеводитель по динозаврам. Princeton University Press. п. 196. ISBN 978-0-691-16766-4.
- ^ Vidal, D .; Mocho, P .; Aberasturi, A .; Sanz, J. L .; Ортега, Ф. (2020). «Скелетная адаптация спинофорозавра с высокой скоростью просмотра свидетельствует об эволюционном нововведении динозавров-зауроподов» Научные отчеты. 10 (1): 6638. Bibcode:2020НатСР..10.6638В. Дои:10.1038 / s41598-020-63439-0. PMID 32313018. S2CID 215819745.
- ^ а б c Knoll, F .; Witmer, L.M .; Ортега, Ф .; Ridgely, R.C .; Шварц-Уингс, Д. (2012). Фарке, А.А. (ред.). "Мозг базального динозавра зауроподов Спинофорозавр и 3D-реконструкции краниального эндокаста и внутреннего уха ». PLOS ONE. 7 (1): e30060. Bibcode:2012PLoSO ... 730060K. Дои:10.1371 / journal.pone.0030060. ЧВК 3260197. PMID 22272273.
- ^ Тейлор, М. П. (2015). Почти все известные шеи зауроподов неполные и искаженные.. PeerJ PrePrints. Дои:10.7287 / peerj.preprints.1418v1.
- ^ Brett-Surman, M.K .; Holtz, Jr., Thomas R .; Фарлоу, Джеймс О. (2012). «Остеология динозавров». Полный динозавр, второе издание. Издательство Индианского университета. С. 135–149. ISBN 978-0-253-00849-7.
- ^ а б Mocho, P .; Ортега, Ф .; Aberasturi, A .; Эскасо, Ф. (2013). "Спинофорозавр (Sauropoda), новый взгляд на эволюцию евзауроподов ». Тезисы докладов VI Международного симпозиума по палеонтологии динозавров и их среде обитания. Салас-де-лос-Инфантес Бургос. С. 89–90.
- ^ Апестегия, С. (2005). «Эволюция комплекса гипосфен-гипантрум в зауроподах». В Tidwell, V .; Карпентер, К. (ред.). Громовые ящерицы: динозавры зауроподоморфа. Блумингтон и Индианаполис: Издательство Индианского университета. стр.248 –267. ISBN 978-0-253-34542-4.
- ^ а б Vidal, D.C .; Ортега, Ф. (2015). Специализированный хвост Spinophorosaurus nigerensis (Sauropoda. Средний юрский период) и остеологические ограничения диапазона его движений.. 63-й SVPCA (Симпозиум по палеонтологии позвоночных и сравнительной анатомии), Саутгемптон, Англия.
- ^ а б c Nair, J. P .; Солсбери, С. В. (2012). «Новые анатомические сведения о Роэтозавр Brownei Longman, 1926, гравизаврический динозавр-завроподоморф из средней юры Квинсленда, Австралия ». Журнал палеонтологии позвоночных. 32 (2): 369. Дои:10.1080/02724634.2012.622324. S2CID 85317450.
- ^ а б Holwerda, F.M .; Поль Д. (2018). «Филогенетический анализ базальных эусавропод Гондваны из ранней-средней юры Патагонии, Аргентина». Испанский журнал палеонтологии. 33 (2): 289. Дои:10.7203 / sjp.33.2.13604.
- ^ Бронзати, М .; Benson, R. B.J .; Rauhut, O.W.M .; Мэннион, П. (2018). «Быстрая трансформация головного мозга динозавров зауроподов: комплексная эволюция мозга и шеи у ранних зауроподов?». Палеонтология. 61 (2): 289–302. Дои:10.1111 / pala.12344.
- ^ а б Xing, L .; Мияшита, Т .; Карри, П. Дж .; Вы, H .; Zhang, J .; Донг, З. (2015). «Новый базальный евзауропод из средней юры Юньнани, Китай, а также состав фауны и переходы азиатских динозавров зауроподоморф». Acta Palaeontologica Polonica. 60 (1): 145–155. Дои:10.4202 / app.2012.0151. S2CID 59143277.
- ^ Jentgen-Ceschino, C .; Стейн, К. (2018). Случаи патологического роста костей в Исанозавр и Спинофорозавр (Зауропода). 5-й Международный палеонтологический конгресс - Кампус Пьера и Марии Кюри Сорбонского университета; Национальный музей естественной истории, Париж, Франция. Дои:10.13140 / RG.2.2.10914.27840.
- ^ Fronimos, J. A .; Уилсон, Дж. А. (2017). «Сложность нейроцентрального шва и распределение напряжения в позвоночнике динозавра зауроподов». Амегиниана. 54 (1): 36–49. Дои:10.5710 / AMGH.05.09.2016.3009. S2CID 132983807.
- ^ Видаль, Д. К. (2018). Могут ли зауроподы совершать «клоакальный поцелуй»? Доказательства возможностей вязки из виртуального Спинофорозавр. XVI EJIP, В Сарауце.
- ^ а б c Mudroch, A .; Richter, U .; Joger, U .; Kosma, R .; Idé, O .; Мага, А .; Фарке, А. А. (2011). «Дидактильные следы паравианских теропод (Maniraptora) из? Средней юры Африки». PLOS ONE. 6 (2): e14642. Bibcode:2011PLoSO ... 614642M. Дои:10.1371 / journal.pone.0014642. ЧВК 3038851. PMID 21339816.
- ^ Муди, Р. Т. Дж. (1997). «Бассейн Иуллеммеден». В Селли, Р. С. (ред.). Осадочные бассейны мира. 3. Elsevier Science. С. 89–103. Дои:10.1016 / S1874-5997 (97) 80008-1. ISBN 978-0444825711.
- ^ Серрано-Мартинес, А .; Vidal, D .; Sciscio, L .; Ортега, Ф .; Кнолль, Ф. (2015). «Изолированные зубы теропод из средней юры Нигера и ранняя эволюция зубов Spinosauridae». Acta Palaeontologica Polonica: 403–415. Дои:10.4202 / app.00101.2014. S2CID 53331040.
- ^ Lockley, M. G .; Harris, J.D .; Li, R .; Xing, L.D .; ван дер Люббе, Т. (2016). «Двупалые следы во времени: по следам« хищников »и их союзников». В П. Л. Фалкингеме; Да. Марти; А. Рихтер (ред.). Следы динозавров - следующие шаги. Блумингтон и Индианаполис: Издательство Индианского университета. С. 183–200. ISBN 978-0-253-02102-1.
внешняя ссылка
- Проект Дино – Google Искусство и культура серия фотографий о раскопках Брауншвейгского музея в Нигере
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||












