Нейроанатомия - Neuroanatomy
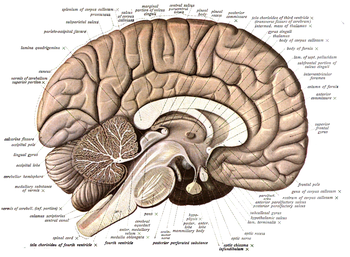
Нейроанатомия это исследование структуры и организации нервная система. В отличие от животных с радиальная симметрия, нервная система которого состоит из распределенной сети клеток, животные с двусторонняя симметрия имеют обособленную нервную систему. Таким образом, их нейроанатомия лучше изучена. В позвоночные, нервная система отделена от внутренней структуры мозг и спинной мозг (вместе называли Центральная нервная система, или ЦНС) и пути нервов, которые соединяются с остальной частью тела (известные как периферическая нервная система, или ПНС). Выделение отдельных структур и областей нервной системы имело решающее значение для исследования того, как она работает. Например, многое из того, что узнали нейробиологи, основано на наблюдении за тем, как повреждения или «поражения» определенных областей мозга влияют на поведение или другие нейронные функции.
Для получения информации о составе нервных систем животных, кроме человека, см. нервная система. Для получения информации о типичной структуре нервной системы Homo sapiens см. человеческий мозг или же периферическая нервная система. В этой статье обсуждается информация, относящаяся к изучать нейроанатомии.
История

Первая известная письменная запись об исследовании анатомии человеческого мозга - это древнеегипетский документ, Эдвин Смит Папирус.[1] Следующее крупное развитие нейроанатомии было сделано греком Алкмеоном, который определил, что мозг, а не сердце управляет телом, и что чувства зависят от мозга.[2]
После открытий Алкмеона многие ученые, философы и врачи со всего мира продолжали вносить свой вклад в понимание нейроанатомии, в частности: Гален, Герофил, Разес и Эрасистрат. Герофил и Эрасистрат Александрийские были, пожалуй, самыми влиятельными греческими нейробиологами, занимавшимися препарированием мозга.[2] В течение нескольких сотен лет после этого, в условиях культурного табу на вскрытие, в нейробиологии не произошло значительного прогресса. Однако папа Сикст IV эффективно возродил изучение нейроанатомии, изменив политику папы и разрешив человеческое вскрытие. Это привело к буму исследований в области нейроанатомии художников и ученых эпохи Возрождения.[3]
В 1664 г. Томас Уиллис, врач и профессор Оксфордского университета, ввел термин «неврология», когда опубликовал свой текст «Анатомия мозга», который считается основой нейроанатомии.[4] Последующие триста пятьдесят с лишним лет привели к большому количеству документации и исследований нейронной системы.
Сочинение
На тканевом уровне нервная система состоит из нейроны, глиальные клетки, и внеклеточный матрикс. И нейроны, и глиальные клетки бывают разных типов (см., Например, раздел нервной системы список различных типов клеток в теле взрослого человека ). Нейроны - это клетки нервной системы, обрабатывающие информацию: они ощущают окружающую среду, общаются друг с другом с помощью электрических сигналов и химических веществ, называемых нейротрансмиттерами, которые обычно действуют через синапсы (тесные контакты между двумя нейронами или между нейроном и мышечной клеткой; обратите внимание, также возможны внесинаптические эффекты, а также высвобождение нейротрансмиттеров во внеклеточное нервное пространство) и вызывают наши воспоминания, мысли и движения. Глиальные клетки поддерживают гомеостаз, производят миелин (олигодендроциты), и обеспечивают поддержку и защиту нейронов мозга. Некоторые глиальные клетки (астроциты ) может даже распространяться межклеточно волны кальция на большие расстояния в ответ на стимуляцию и высвобождение глиотрансмиттеры в ответ на изменение концентрации кальция. Шрамы от ран в головном мозге в основном содержат астроциты. В внеклеточный матрикс также обеспечивает поддержку на молекулярном уровне клеток мозга, переносящих вещества в кровеносные сосуды и из них.
На уровне органов нервная система состоит из областей мозга, таких как гиппокамп у млекопитающих или грибовидные тела из плодовая муха.[5] Эти области часто имеют модульную структуру и играют особую роль в общих системных путях нервной системы. Например, гиппокамп имеет решающее значение для формирования воспоминаний, связанных со многими другими церебральными областями. Периферическая нервная система также содержит афферентные или эфферентные нервы, которые представляют собой пучки волокон, которые исходят из головного и спинного мозга или из сенсорных или моторных периферических ганглиев и многократно разветвляются, иннервируя каждую часть тела. Нервы состоят в основном из аксоны или дендриты нейронов (аксоны в случае эфферентных моторных волокон и дендриты в случае афферентных сенсорных волокон нервов) вместе с множеством мембран, которые обертывают и разделяют их на нервные пучки.
Нервная система позвоночных делится на центральную и периферическую нервную систему. В Центральная нервная система (ЦНС) состоит из мозг, сетчатка, и спинной мозг, в то время как периферическая нервная система (ПНС) состоит из всех нервов и ганглиев (пакетов периферических нейронов) вне ЦНС, которые соединяют ее с остальной частью тела. ПНС подразделяется на соматическую и вегетативную нервные системы. В соматическая нервная система состоит из «афферентных» нейронов, которые переносят сенсорную информацию от соматических (телесных) органов чувств в ЦНС, и «эфферентных» нейронов, которые передают двигательные инструкции произвольным мышцам тела. В автономная нервная система может работать с контролем ЦНС или без него (поэтому он называется «автономным»), а также имеет два подразделения, называемые сочувствующий и парасимпатический, которые важны для передачи двигательных приказов основным внутренним органам тела, таким образом контролируя такие функции, как сердцебиение, дыхание, пищеварение и слюноотделение. Вегетативные нервы, в отличие от соматических, содержат только эфферентные волокна. Сенсорные сигналы, поступающие из внутренних органов, проходят в ЦНС через соматические сенсорные нервы (например, висцеральная боль) или через некоторые конкретные черепные нервы (например, химиочувствительные или механические сигналы).
Ориентация в нейроанатомии

В анатомии в целом и нейроанатомии в частности для обозначения ориентации и местоположения используется несколько наборов топографических терминов, которые обычно относятся к оси тела или мозга (см. Анатомические условия расположения ). Ось ЦНС часто ошибочно считают более или менее прямой, но на самом деле она всегда показывает два вентральных изгиба (шейный и головной изгиб) и дорсальный изгиб (изгиб моста), все из-за различного роста во время эмбриогенеза. В нейроанатомии чаще всего используются следующие пары терминов:
- Дорсальный и вентральный: дорсальный свободно относится к верхней или верхней стороне, представленной верхней пластинкой мозга, и вентрально к нижней или нижней стороне, представленной пластиной пола. Эти дескрипторы изначально использовались для спина и вентрум - спина и живот - тела; живот большинства животных ориентирован к земле; прямая поза человека помещает наш вентральный аспект вперед, а спинной аспект становится задним. Случай головы и мозга особенный, поскольку живот не переходит в голову должным образом, если только мы не предположим, что рот представляет собой вытянутый элемент живота. Поэтому обычно те части мозга, которые лежат близко к основанию черепа и через него к ротовой полости, называются вентральными, то есть на его нижней или нижней стороне, как определено выше, тогда как дорсальные части ближе к ограждающий свод черепа. Ссылка на верхнюю и нижнюю пластины мозга менее подвержена путанице, также позволяет нам следить за осевыми изгибами, упомянутыми выше. Таким образом, дорсальный и вентральный - относительные термины в головном мозге, точное значение которых зависит от конкретного места.
- Рострально-каудальное: ростральный относится в общей анатомии к передней части тела (к носу или трибуна на латыни), и каудальный относится к хвостовой части тела (по направлению к хвосту; кауда на латыни). Рострокаудальный размер головного мозга соответствует его длине оси, которая проходит через указанные изгибы от каудального конца спинного мозга к ростральному концу примерно на перекрестье зрительных нервов. В прямом Человеке термины направления «высший» и «низший» по существу относятся к этому рострокаудальному измерению, потому что оси нашего тела и мозга примерно ориентированы вертикально в вертикальном положении. Однако у всех позвоночных в нервной трубке развивается очень заметный вентральный излом нервной трубки, который все еще обнаруживается в центральной нервной системе взрослых, известный как головной изгиб. Последний изгибает ростральную часть ЦНС на 180 градусов относительно каудальной части, на переходе между передний мозг (ось заканчивается рострально на перекрестье зрительного нерва) и мозговой ствол и спинной мозг (ось примерно вертикальна, но включает дополнительные незначительные перегибы на мостике и шейных изгибах) Эти изгибные изменения осевого размера проблематичны при попытке описать относительное положение и плоскости сечения в головном мозге. Существует множество литературы, в которой ошибочно игнорируются осевые изгибы и предполагается относительно прямая ось мозга.
- Медиальный и латеральный: медиальный означает, что он находится близко или относительно ближе к средней линии (дескриптор медиана означает положение точно по средней линии). Боковой наоборот (позиция более или менее отделена от средней линии).
Обратите внимание, что такие дескрипторы (дорсальный / вентральный, ростральный / каудальный; медиальный / латеральный) являются относительными, а не абсолютными (например, можно сказать, что латеральная структура лежит медиальнее чего-то еще, что лежит еще более латерально).
Обычно используемые термины для плоскостей ориентации или плоскостей сечения в нейроанатомии - «сагиттальный», «поперечный» или «коронарный», а также «осевой» или «горизонтальный». Опять же, в этом случае ситуация для плавающих, ползающих или четвероногих (лежачих) животных отличается от ситуации для человека или других прямостоячих видов из-за измененного положения оси. Из-за осевых изгибов головного мозга ни одна плоскость сечения никогда не достигает полной серии сечений в выбранной плоскости, потому что некоторые сечения неизбежно приводят к разрезанию под наклоном или даже перпендикулярно к нему, когда они проходят через изгибы. Опыт позволяет различать части, которые в результате сокращаются по желанию.
- В средней сагиттальной плоскости тело и мозг делятся на левую и правую половины; сагиттальные сечения, как правило, параллельны этой срединной плоскости, перемещаясь по медиально-латеральному измерению (см. изображение выше). Период, термин сагиттальный этимологически относится к срединному шву между правой и левой теменными костями черепа, известному классически как сагиттальный шов, поскольку он выглядит примерно как стрелка в месте слияния с другими швами (сагитта; стрелка на латыни).
- Плоскость сечения, ортогональная оси любой удлиненной формы, в принципе считается поперечной (например, поперечное сечение пальца или позвоночника); если нет оси длины, нет возможности определить такие сечения, или есть бесконечные возможности). Следовательно, поперечные сечения тела у позвоночных параллельны ребрам, которые ортогональны позвоночнику, который представляет ось тела как у животных, так и у человека. Мозг также имеет собственную продольную ось - продольную ось первичной удлиненной нервной трубки - которая становится в значительной степени вертикальной при вертикальном положении человека, так же, как ось тела, за исключением ее рострального конца, как отмечалось выше. Это объясняет, что поперечные срезы спинного мозга примерно параллельны нашим ребрам или земле. Однако это верно только для спинного мозга и ствола мозга, поскольку передний мозговой конец нервной оси изгибается как изгиб во время раннего морфогенеза в хиазматический гипоталамус, где и заканчивается; соответственно изменяется ориентация истинных поперечных сечений, и они перестают быть параллельны ребрам и земле, а перпендикулярны им; Незнание этой морфологической особенности мозга (присутствующей во всех без исключения позвоночных головного мозга) вызывало и до сих пор вызывает множество ошибочных представлений о частях мозга переднего мозга. Признавая особенность ростральных поперечных сечений, традиция ввела для них другой дескриптор, а именно: венечный разделы. Венечные срезы разделяют передний мозг от рострального (переднего) до каудального (заднего), образуя серию, ортогональную (поперечную) к локальной оси изгиба. Эту концепцию нельзя осмысленно применять к стволу головного мозга и спинному мозгу, так как там коронковые секции становятся горизонтальными по отношению к осевому измерению, будучи параллельными оси. В любом случае концепция «корональных» сечений менее точна, чем «поперечные», поскольку часто используются плоскости корональных сечений, которые не являются истинно ортогональными к ростральному концу оси мозга. Этот термин этимологически связан с коронарный шов кранев и это положение, в котором носят коронки (лат. корона означает корона). Неясно, какой вид короны имелся в виду изначально (возможно, просто диадема), и это, к сожалению, приводит к неоднозначности в плоскости сечения, определяемой просто как коронка.
- Коронарная плоскость, пересекающая человеческую голову и мозг, по современным представлениям должна быть параллельна лицу (плоскость, в которой корона короля сидит на его голове, не совсем параллельна лицу, и эта концепция была перенесена на животных с менее развитыми фронтальными способностями, чем у нас. очевидно, даже более противоречивый, но есть неявная ссылка на коронарный шов черепа, который образуется между лобной и височной / теменной костями, давая своего рода конфигурацию диадемы, которая примерно параллельна лицу). Таким образом, плоскости коронарного сечения относятся только к голове и мозгу, где диадема имеет смысл, а не к шее и телу ниже.
- Горизонтальные участки по определению выровнены (параллельны) горизонту. У плавающих, ползучих и четвероногих животных ось тела сама по себе горизонтальна, и поэтому горизонтальные участки проходят по длине спинного мозга, отделяя брюшную часть от спинной. Горизонтальные сечения ортогональны как поперечным, так и сагиттальным сечениям и теоретически параллельны оси длины. Из-за осевого изгиба головного мозга (переднего мозга) истинные горизонтальные сечения в этой области ортогональны венечным (поперечным) сечениям (как и горизонт относительно лица).
Согласно этим соображениям, три направления пространства представлены именно сагиттальной, поперечной и горизонтальной плоскостями, тогда как коронарные сечения могут быть поперечными, наклонными или горизонтальными, в зависимости от того, как они соотносятся с осью мозга и ее изгибами.
Инструменты
Современные разработки в нейроанатомии напрямую связаны с технологиями, используемыми для выполнения исследование. Следовательно, необходимо обсудить различные доступные инструменты. Многие из гистологический методы, используемые для изучения других тканей, также могут быть применены к нервной системе. Однако есть некоторые методы, которые были разработаны специально для изучения нейроанатомии.
Окрашивание клеток
В биологических системах окрашивание - это метод, используемый для увеличения контрастности отдельных деталей на микроскопических изображениях.
Окрашивание по Нисслю использует анилиновые основные красители для интенсивного окрашивания кислых полирибосом в шероховатой эндоплазматической сети, которого много в нейронах. Это позволяет исследователям различать разные типы клеток (например, нейроны и глия ), а также формы и размеры нейронов в различных областях нервной системы. цитоархитектура.
Классический Пятно Гольджи использует дихромат калия и нитрат серебра чтобы выборочно заполнить хроматом серебра осадок нескольких нервных клеток (нейронов или глии, но в принципе любые клетки могут реагировать аналогичным образом). Эта так называемая процедура пропитки хроматом серебра полностью или частично окрашивает тела клеток и нейриты некоторых нейронов -дендриты, аксон - коричневого и черного цвета, что позволяет исследователям проследить свой путь до самых тонких конечных ветвей в срезе нервной ткани благодаря прозрачности, обусловленной отсутствием окрашивания в большинстве окружающих клеток. В настоящее время материал, пропитанный методом Гольджи, адаптирован для электронной микроскопии визуализации неокрашенных элементов, окружающих окрашенные отростки и тела клеток, тем самым добавляя дополнительную разрешающую способность.
Гистохимия
Гистохимия использует знания о свойствах биохимических реакций химических компонентов мозга (включая, в частности, ферменты), чтобы применять селективные методы реагирования для визуализации того, где они происходят в мозге, а также любых функциональных или патологических изменений. Это важно в отношении молекул, связанных с производством и метаболизмом нейротрансмиттеров, но также применимо во многих других направлениях химиоархитектуры или химической нейроанатомии.
Иммуноцитохимия представляет собой особый случай гистохимии, который использует селективные антитела против различных химических эпитопов нервной системы для избирательного окрашивания определенных типов клеток, аксональных пучков, нейропилей, глиальных отростков или кровеносных сосудов, или определенных внутрицитоплазматических или внутриядерных белков и других иммуногенетических молекул, например , нейротрансмиттеры. Иммунореагированные белки факторов транскрипции показывают считывание генома в терминах транслированного белка. Это значительно увеличивает способность исследователей различать разные типы клеток (например, нейроны и глия ) в различных областях нервной системы.
Гибридизация in situ использует синтетические РНК-зонды, которые селективно прикрепляются (гибридизуются) к комплементарным транскриптам мРНК экзонов ДНК в цитоплазме, чтобы визуализировать считывание генома, то есть различать активную экспрессию гена с точки зрения мРНК, а не белка. Это позволяет гистологически (in situ) идентифицировать клетки, участвующие в производстве генетически закодированных молекул, которые часто представляют собой дифференцировку или функциональные признаки, а также молекулярные границы, разделяющие отдельные домены мозга или популяции клеток.
Генетически закодированные маркеры
Выражая в мозге переменное количество красных, зеленых и синих флуоресцентных белков, так называемые "мозговой лук «Мутантная мышь позволяет комбинаторно визуализировать множество разных цветов в нейронах. Это помечает нейроны достаточно уникальными цветами, чтобы их можно было часто отличить от своих соседей. флуоресцентная микроскопия, позволяя исследователям отображать локальные связи или взаимное расположение (мозаику) между нейронами.
Оптогенетика использует трансгенную конститутивную и сайт-специфическую экспрессию (обычно у мышей) заблокированных маркеров, которые можно избирательно активировать при освещении световым лучом. Это позволяет исследователям очень разборчивым образом изучать связи аксонов в нервной системе.
Неинвазивная визуализация мозга
Магнитно-резонансная томография широко используется для исследования мозга структура и функция неинвазивно у здоровых людей. Важным примером является диффузионная тензорная визуализация, который основан на ограниченной диффузии воды в ткани для получения изображений аксонов. В частности, вода движется быстрее в направлении, совпадающем с аксонами, что позволяет сделать вывод об их структуре.
Вирусные методы
Некоторые вирусы могут реплицироваться в клетках мозга и перекрестных синапсах. Таким образом, вирусы, модифицированные для экспрессии маркеров (таких как флуоресцентные белки), могут использоваться для отслеживания связи между областями мозга через несколько синапсов.[6] Два индикаторных вируса, которые реплицируются и распространяются транснейронально / транссинаптически, являются Вирус простого герпеса тип 1 (HSV)[7] и Рабдовирусы.[8] Вирус простого герпеса использовался для отслеживания связей между мозгом и желудком, чтобы исследовать области мозга, участвующие в висцеро-сенсорной обработке.[9] В другом исследовании вирус простого герпеса вводился в глаз, что позволяло визуализировать оптический путь от сетчатка в зрительная система.[10] Примером трассирующего вируса, который реплицируется из синапса в сому, является вирус псевдобешенства.[11] Используя вирусы псевдобешенства с разными флуоресцентными репортерами, модели двойного заражения могут анализировать сложную синаптическую архитектуру.[12]
Методы на основе красителей
Аксональный транспорт методы используют различные красители (варианты пероксидазы хрена, флуоресцентные или радиоактивные маркеры, лектины, декстраны), которые более или менее активно поглощаются нейронами или их отростками. Эти молекулы селективно транспортируются антероградно (от сомы к окончанию аксона) или ретроградно (от окончаний аксонов до сомы), что свидетельствует о первичных и побочных связях в головном мозге. Эти «физиологические» методы (поскольку используются свойства живых, неповрежденных клеток) могут быть объединены с другими процедурами и существенно заменили более ранние процедуры, изучающие дегенерацию поврежденных нейронов или аксонов. Подробные синаптические связи могут быть определены с помощью корреляционной электронной микроскопии.
Коннектомика
Последовательная секционная электронная микроскопия получила широкое развитие для изучения нервной системы. Например, первое применение серийная растровая электронная микроскопия на корковой ткани грызунов.[13] Реконструкция схемы на основе данных, полученных с помощью этого высокопроизводительного метода, является сложной задачей, и научная игра Citizen EyeWire был разработан, чтобы помочь исследованиям в этой области.
Вычислительная нейроанатомия
Это область, в которой используются различные методы визуализации и вычислительные методы для моделирования и количественной оценки пространственно-временной динамики нейроанатомических структур как в нормальных, так и в клинических популяциях.
Модельные системы
Помимо человеческий мозг, есть много других животных, чей мозг и нервная система были тщательно изучены. модельные системы, включая мышей, данио,[14] плодовая муха,[15] и разновидность круглых червей, называемых C. elegans. У каждой из них есть свои преимущества и недостатки как модельной системы. Например, C. elegans нервная система чрезвычайно стереотипна от одного червя к другому. Это позволило исследователям использовать электронная микроскопия чтобы отобразить пути и связи всех примерно 300 нейронов этого вида. Плодовая муха широко изучается отчасти потому, что ее генетика очень хорошо изучена, и с ней легко манипулировать. Мышь используется, потому что ее мозг, как млекопитающее, больше похож по структуре на наш мозг (например, он имеет шестиуровневую структуру). кора, однако его гены можно легко модифицировать, а его репродуктивный цикл относительно быстр).
Caenorhabditis elegans

Мозг небольшой и простой у некоторых видов, таких как нематода червь, строение которого довольно простое: трубка с полой кишкой, идущей ото рта к анусу, и нервный шнур с расширением ( ганглий ) для каждого сегмента тела, с особенно большим ганглием спереди, называемым мозгом. Нематода Caenorhabditis elegans был изучен из-за его важности в генетике.[16] В начале 1970-х гг. Сидней Бреннер выбрал его в качестве модельной системы для изучения того, как гены контролируют развитие, включая развитие нейронов. Одним из преимуществ работы с этим червем является то, что нервная система гермафродит содержит ровно 302 нейрона, всегда в одних и тех же местах, создавая идентичные синаптические связи у каждого червя.[17] Команда Бреннера разрезала червей на тысячи ультратонких срезов и сфотографировала каждый срез под электронным микроскопом, затем визуально сопоставила волокна от среза к срезу, чтобы нанести на карту каждый нейрон и синапс во всем теле, чтобы получить полную картину. коннектом нематоды.[18] Для любого другого организма нет ничего, приближающегося к такому уровню детализации, и эта информация была использована для проведения множества исследований, которые без нее были бы невозможны.[19]
Drosophila melanogaster
Drosophila melanogaster является популярным экспериментальным животным, потому что его легко выращивать в массовом порядке из дикой природы, у него короткое время генерации и легко получить мутантных животных.
Членистоногие иметь центральный мозг с тремя отделениями и большой оптические лепестки за каждым глазом для визуальной обработки. Мозг плодовой мухи содержит несколько миллионов синапсов по сравнению со 100 миллиардами в человеческом мозге. Примерно две трети мозга дрозофилы посвящены визуальная обработка.
Томас Хант Морган начал работать с Drosophila в 1906 году, и эта работа принесла ему Нобелевскую премию 1933 года по медицине за определение хромосом как вектора наследования генов. Из-за большого количества инструментов, доступных для изучения генетики дрозофилы, они стали естественным предметом для изучения роли генов в нервной системе.[20] Геном был секвенирован и опубликован в 2000 году. Около 75% известных генов болезней человека имеют узнаваемое совпадение в геноме плодовых мух. Дрозофила используется в качестве генетической модели для нескольких неврологических заболеваний человека, включая нейродегенеративные расстройства Паркинсона, Хантингтона, спиноцеребеллярную атаксию и болезнь Альцгеймера. Несмотря на большую эволюционную дистанцию между насекомыми и млекопитающими, многие основные аспекты Дрозофила нейрогенетика оказалась актуальной для человека. Например, первые гены биологических часов были идентифицированы путем изучения Дрозофила мутанты, показавшие нарушенные циклы суточной активности.[21]
Смотрите также
Рекомендации
- ^ Атта, Х. М. (1999). "Хирургический папирус Эдвина Смита: старейший известный хирургический трактат". Американский хирург. 65 (12): 1190–1192. PMID 10597074.
- ^ а б Роза, F (2009). «Церебральная локализация в древности». Журнал истории неврологии. 18 (3): 239–247. Дои:10.1080/09647040802025052. PMID 20183203. S2CID 5195450.
- ^ Ginn, S. R .; Лоруссо, Л. (2008). «Мозг, разум и тело: взаимодействие с искусством в Италии эпохи Возрождения». Журнал истории неврологии. 17 (3): 295–313. Дои:10.1080/09647040701575900. PMID 18629698. S2CID 35600367.
- ^ Неер, А (2009). «Кристофер Рен, Томас Уиллис и изображение мозга и нервов». Журнал медицинских гуманитарных наук. 30 (3): 191–200. Дои:10.1007 / s10912-009-9085-5. PMID 19633935. S2CID 11121186.
- ^ Грибные тела плодовой мухи В архиве 2012-07-16 в Archive.today
- ^ Имбирь, М .; Haberl, M .; Конзельманн, К.-К .; Schwarz, M .; Фрик, А. (2013). «Раскрытие секретов нейронных цепей с помощью технологии рекомбинантного вируса бешенства». Передний. Нейронные цепи. 7: 2. Дои:10.3389 / fncir.2013.00002. ЧВК 3553424. PMID 23355811.
- ^ Макговерн, AE; Дэвис-Пойнтер, Н. Ракоци, Дж; Фиппс, S; Симмонс, Д.Г.; Маццоне, SB (2012). «Антероградное отслеживание нейронных цепей с использованием генетически модифицированного вируса простого герпеса, экспрессирующего EGFP». J Neurosci методы. 209 (1): 158–67. Дои:10.1016 / j.jneumeth.2012.05.035. PMID 22687938. S2CID 20370171.
- ^ Кайперс Х.Г., Уголини Г. (февраль 1990 г.). «Вирусы как транснейрональные трассеры». Тенденции в неврологии. 13 (2): 71–5. Дои:10.1016 / 0166-2236 (90) 90071-Н. PMID 1690933. S2CID 27938628.
- ^ Ринаман Л., Шварц Г. (март 2004 г.). «Антероградное транснейрональное вирусное отслеживание центральных висцеросенсорных путей у крыс». Журнал неврологии. 24 (11): 2782–6. Дои:10.1523 / JNEUROSCI.5329-03.2004. ЧВК 6729508. PMID 15028771.
- ^ Норгрен Р. Б., Маклин Дж. Х., Бубель Х. С., Вандер А., Бернштейн Д. И., Леман М. Н. (март 1992 г.). «Антероградный транспорт HSV-1 и HSV-2 в зрительной системе». Бюллетень исследований мозга. 28 (3): 393–9. Дои:10.1016 / 0361-9230 (92) 90038-У. PMID 1317240. S2CID 4701001.
- ^ Карта, Дж. П. (2001). «Нейроинвазивность вируса псевдобешенства: окно в функциональную организацию мозга». Достижения в вирусных исследованиях. 56: 39–71. Дои:10.1016 / S0065-3527 (01) 56004-2. ISBN 9780120398560. PMID 11450308.
- ^ Кард, Дж. П. (2011). «Подход с использованием условного репортера вируса псевдобешенства двойной инфекции для определения проекций на обеспеченные нейроны в сложных нервных цепях». PLOS ONE. 6 (6): e21141. Bibcode:2011PLoSO ... 621141C. Дои:10.1371 / journal.pone.0021141. ЧВК 3116869. PMID 21698154.
- ^ Денк, Вт; Хорстманн, Х (2004). «Последовательная сканирующая электронная микроскопия лица для восстановления трехмерной тканевой наноструктуры». PLOS Биология. 2 (11): e329. Дои:10.1371 / journal.pbio.0020329. ЧВК 524270. PMID 15514700.
- ^ Вуллиманн, Марио Ф .; Рупп, Барбар; Райхерт, Генрих (1996). Нейроанатомия мозга рыбок данио: топологический атлас. ISBN 3-7643-5120-9. В архиве из оригинала от 15.06.2013. Получено 2016-10-16.
- ^ «Атлас мозга дрозофилы». Архивировано из оригинал на 2011-07-16. Получено 2011-03-24.
- ^ "WormBook: онлайн-обзор C. elegans биология". Архивировано из оригинал на 2011-10-11. Получено 2011-10-14.
- ^ Хоберт, Оливер (2005). В C. elegans Исследовательское сообщество (ред.). «Спецификация нервной системы». WormBook: 1–19. Дои:10.1895 / wormbook.1.12.1. ЧВК 4781215. PMID 18050401. В архиве из оригинала от 17.07.2011. Получено 2011-11-05.
- ^ Белый, JG; Саутгейт, E; Томсон, Дж. Н.; Бреннер, S (1986). «Строение нервной системы нематоды Caenorhabditis elegans». Философские труды Королевского общества B. 314 (1165): 1–340. Bibcode:1986РСПТБ.314 .... 1Вт. Дои:10.1098 / рстб.1986.0056. PMID 22462104.
- ^ Ходжкин Дж (2001). "Caenorhabditis elegans". В Brenner S, Miller JH (ред.). Энциклопедия генетики. Эльзевир. С. 251–256. ISBN 978-0-12-227080-2.
- ^ "Flybrain: онлайн-атлас и база данных дрозофила нервная система". Архивировано из оригинал на 2016-05-16. Получено 2011-10-14.
- ^ Конопка, Р.Дж.; Бензер, S (1971). «Часовые мутанты Drosophila melanogaster». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 68 (9): 2112–6. Bibcode:1971ПНАС ... 68.2112К. Дои:10.1073 / pnas.68.9.2112. ЧВК 389363. PMID 5002428.
внешняя ссылка
- Нейроанатомия, ежегодный журнал клинической нейроанатомии
- Атласы мозга мышей, крыс, приматов и человека (Центр вычислительной биологии Калифорнийского университета в Лос-Анджелесе)
- brainmaps.org: Атласы мозга с нейроанатомическими аннотациями высокого разрешения
- BrainInfo для нейроанатомии
- Система управления архитектурой мозга, несколько атласов анатомии мозга
- Атлас белого вещества, Атлас диффузно-тензорной визуализации трактов белого вещества мозга

| Поля |  | |
|---|---|---|
| Бактерии | ||
| Протисты |
| |
| Растения | ||
| Беспозвоночные | ||
| Млекопитающие | ||
| Другие позвоночные | ||
| Другие темы | ||
| ||