Морфология насекомых - Insect morphology

2. глазки (ниже)
3. глазки (верхние)
4. сложный глаз
5. мозг (церебральный ганглии )
6. переднегрудь
7. дорсальный кровеносный сосуд
8. трахея трубы (ствол с дыхальце )
9. среднегрудь
10. заднегрудь
11. переднее крыло
12. заднее крыло
13. средний кишечник (желудок)
14. спинная труба (сердце)
15. яичник
16. задняя кишка (кишечник, прямая кишка и анус)
17. анус
18. яйцевод
19. нервный шнур (брюшные ганглии)
20. Мальпигиевы трубы
21. подушечки предплюсны
22. когти
23. предплюсна
24. большеберцовая кость
25. бедренная кость
26. вертел
27. передняя кишка (зоб, желудок)
28. грудной ганглий
29. тазик
30. слюнная железа
31. подэзофагеальный ганглий
32. ротовые органы
Морфология насекомых это исследование и описание физическая форма из насекомые. Терминология, используемая для описания насекомых, аналогична терминологии, используемой для других членистоногие из-за их общей эволюционной истории. Насекомых от других членистоногих отделяют три физические особенности: их тело разделено на три части (голова, грудная клетка и брюшко), есть три пары ног и расположены ротовые части. за пределами из головная капсула. Это положение ротового аппарата и отличает их от ближайших родственников, не насекомых. гексаподы, который включает Протура, Diplura, и Коллембола.
Строение тела насекомых сильно различается. Частные лица могут колебаться от 0,3 мм (сказки ) до 30 см в поперечнике (мотылек );[1]:7 нет глаз или их много; хорошо развитые крылья или их нет; и ноги, модифицированные для бега, прыжков, плавания или даже копания. Эти модификации позволяют насекомым заселять практически все экологическая ниша на планете, кроме глубокого океана. В этой статье описывается основное тело насекомого и некоторые из основных вариаций различных частей тела; в процессе он определяет многие из технических терминов, используемых для описания тел насекомых.
Резюме анатомии
У насекомых, как и у всех членистоногих, нет внутреннего скелета; вместо этого у них есть экзоскелет, жесткий внешний слой, состоящий в основном из хитин который защищает и поддерживает тело. Тело насекомого делится на три части: голова, грудная клетка, и брюшная полость.[2] Голова предназначена для сенсорного ввода и приема пищи; грудная клетка, которая является точкой привязки для ног и крылья (если есть), специализируется на передвижении; и живот для пищеварение, дыхание, выделение, и размножение.[1]:22–48 Хотя общая функция трех частей тела одинакова для всех видов насекомых, есть существенные различия в базовой структуре, включая крылья, ноги и усики, а ротовой аппарат сильно варьируется от группы к группе.[3]
Внешний
Экзоскелет
Внешний скелет насекомого, кутикула, состоит из двух слоев; в эпикутикула, который представляет собой тонкий, воскообразный, водостойкий внешний слой, не содержащий хитина, а слой под ним называется прокутикула. Это хитин, он намного толще, чем эпикутикула, и состоит из двух слоев: внешний - это экзокутикула, а внутренний - эндокутикула. Жесткая и гибкая эндокутикула состоит из множества слоев волокнистого хитина и белков, перекрещивающихся друг с другом в виде сэндвича, в то время как экзокутикула жесткая и склеротизированный.[1]:22–24 Экзокутикула значительно уменьшена у многих мягкотелых насекомых, особенно у личинка этапы (например, гусеницы ). Химически хитин представляет собой длинноцепочечную полимер из N-ацетилглюкозамин, производное глюкозы. В неизмененном виде хитин полупрозрачный, податливый, упругий и довольно прочный. В членистоногие однако его часто видоизменяют, встраивая в затвердевший белковый матрица, которая формирует большую часть экзоскелет. В чистом виде он кожистый, но при инкрустации карбонат кальция, становится намного сложнее.[4] Разницу между неизмененной и модифицированной формами можно увидеть, сравнив стенку тела гусеница (без изменений) в жук (изменено).
Начиная с самой эмбриональной стадии, слой столбчатых или кубовидных эпителиальных клеток дает начало внешней кутикуле и внутренней базальной мембране. Большая часть материала насекомых находится в эндокутикуле. Кутикула обеспечивает мышечную поддержку и действует как защитный щит по мере развития насекомого. Однако, поскольку он не может расти, внешняя склеротизированная часть кутикулы периодически сбрасывается в процессе, называемом «линькой». По мере приближения времени линьки большая часть материала экзокутикулы реабсорбируется. При линьке сначала старая кутикула отделяется от эпидермиса (аполис ). Ферментативная жидкость для линьки выделяется между старой кутикулой и эпидермисом, которая отделяет экзокутикулу, переваривая эндокутикулу и изолируя ее материал для новой кутикулы. Когда новая кутикула сформировалась в достаточной степени, эпикутикула и редуцированная экзокутикула теряют шелушение.[5]:16–20
Четыре основных области сегмента тела насекомого: тергум или спинной, грудина или вентрально и два плевра или боковые. Закаленные пластины в экзоскелете называются склеритами, которые представляют собой подразделения основных областей - тергитов, стернитов и плевритов для соответствующих областей тергума, грудины и плевры.[6]
Голова



В голова у большинства насекомых заключен в твердый, сильно склеротизированный экзоскелетный головная капсула '. Основное исключение составляют те виды, личинки которых не полностью склеротизированы, в основном это некоторые голометаболы; но даже у большинства не склеротизированных или слабо склеротизированных личинок, как правило, имеются хорошо склеротизированные головные капсулы, например у личинок Coleoptera и Hymenoptera. Личинки Циклоррафа однако, как правило, практически не имеют головной капсулы.
На головной капсуле находится большинство основных органов чувств, включая усики, глазки и сложные глаза. Он также несет на себе ротовой аппарат. У взрослого насекомого головная капсула, по-видимому, не сегментирована, хотя эмбриологические исследования показывают, что она состоит из шести сегментов, которые несут парные головные придатки, включая ротовые части, каждая пара на определенном сегменте.[7] Каждая такая пара занимает один сегмент, хотя не все сегменты у современных насекомых несут видимые отростки.
Из всех отрядов насекомых, Прямокрылые наиболее удобно отображать самые разнообразные черты головы насекомых, включая швы и склериты.[6] Здесь вершина, или верхушка (спинная область), расположена между сложными глазами у насекомых с гипогнатный и опистогнатный головы. В прогнатический насекомых вершина находится не между сложными глазами, а там, где глазки обычно встречаются. Это связано с тем, что первичная ось головы повернута на 90 °, чтобы стать параллельной первичной оси тела. У некоторых видов эта область видоизменена и получает другое название.[8]:13
В экдизиальный шов состоит из венечных, лобных и эпикраниальных швов, а также экдизиальных линий и линий декольте, которые различаются у разных видов насекомых. Экдизиальный шов продольно накладывается на макушку и разделяет надпочечниковые половины головы на левую и правую стороны. В зависимости от насекомого шовный материал может иметь разную форму: Y, U или V. Эти расходящиеся линии, образующие экдизиальный шов, называются лобными или лобными. лобный швы. Не у всех видов насекомых есть лобные швы, но у тех, у которых они есть, швы расщепляются во время шелушение, который помогает обеспечить выход нового возраста из покровов.
В лоб это та часть головной капсулы, которая лежит Вентрад или же предшествующий вершины. Лоб различается по размеру относительно насекомого, и у многих видов определение его границ произвольно, даже у некоторых таксонов насекомых, у которых есть четко выраженные головные капсулы. Однако у большинства видов лоб на переднем крае граничит с лобно-липеальной или эпистомальной бороздой над наличником. Боковой он ограничен лобно-щечной бороздой, если она есть, и границей с макушкой, линией экдизиального расщепления, если она видна. Если есть срединный глазок, то он обычно находится на лбу, хотя у некоторых насекомых, таких как многие перепончатокрылые, все три глазка появляются на макушке. Более формальное определение состоит в том, что это склерит, из которого возникают мышцы-расширители глотки, но во многих контекстах это тоже бесполезно.[7] В анатомии некоторых таксонов, например многих Цикадоморфа, передняя часть головы довольно четко выделяется и имеет тенденцию быть широкой и субвертикальной; эта срединная область обычно принимается за лоб.[9]
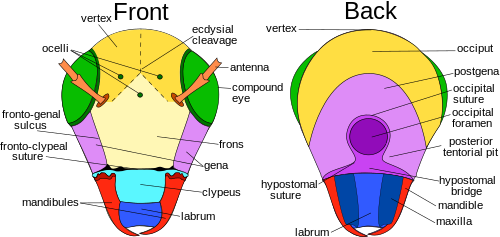
В наличник представляет собой склерит между лицом и верхней губой, который у примитивных насекомых отделяется от лба дорсально лобным швом. Клипеогенный шов латерально ограничивает наличник, при этом наличник вентрально отделен от верхней губы клипеолабральным швом. Наличник различается по форме и размеру, например у видов чешуекрылых с большим наличником с удлиненным ротовым аппаратом. Щека или гена образует склеротизированные области на каждой стороне головы под сложными глазами, доходящими до горлового шва. Как и многие другие части, составляющие голову насекомого, щека у разных видов различается, и ее границы трудно установить. Например, в стрекозы и стрекозы, он находится между сложными глазами, наличником и ротовым аппаратом. В Postgena область непосредственно кзади, сзади или ниже щеки крылатые насекомые, образует боковую и вентральную части затылочной дуги. Затылочная дуга - это узкая полоса, образующая задний край головной капсулы, изгибающийся дорсально над отверстием. Субгенальная область обычно узкая, расположена над ротовым аппаратом; эта область также включает гипостома и плевростома.[8]:13–14 Темя выступает вперед над основаниями усиков в виде выступающего заостренного вогнутого рострума. Задняя стенка головной капсулы пронизана большим отверстием - foramen. Через него проходят системы органов, такие как нервный шнур, пищевод, слюнные протоки, и мускулатура, соединяя голову с грудная клетка.[10]
На задней стороне головы расположены затылок, Postgena, затылочное отверстие, задняя тенториальная ямка, гула, постгенальный мост, гипостомальный шов и мост, а челюсти, губа, и верхняя челюсть. Затылочный шов хорошо заложен у видов Orthoptera, но не так сильно у других отрядов. Затылочный шов - это дугообразная подковообразная бороздка на затылке, оканчивающаяся на задней части каждой нижней челюсти. Постзатылочный шов является ориентиром на задней поверхности головы и обычно находится рядом с затылочным мастером. У крыловидных отростков задняя задняя часть образует крайнюю заднюю, часто U-образную форму, которая образует ободок головы, доходящий до постзатылочного шва. У крыловидных животных, например у прямокрылых, затылочное отверстие и рот не разделены. Три типа затылочных закрытий или точек под затылочным отверстием, которые разделяют две нижние половины постгены, это: гипостомальный мост, постгенальный мост и гула. Гипостомальный мост обычно встречается у насекомых с гипогнатной ориентацией. Постгенальный мостик встречается у взрослых особей высших Двукрылые и острый Перепончатокрылые, а гула встречается на некоторых Жесткокрылые, Neuroptera, и Изоптера, которые обычно имеют ротовой аппарат, ориентированный на прогнат.[8]:15
Сложные глаза и глазки
У большинства насекомых есть одна пара больших выступающих сложные глаза состоит из единиц, называемых омматидиями (омматидий, в единственном числе), возможно, до 30 000 в одном сложном глазу, например, у больших стрекоз. Этот тип глаза дает меньшее разрешение, чем глаза у позвоночных, но он дает острое восприятие движения и обычно обладает чувствительностью к ультрафиолету и зеленому цвету и может иметь дополнительные пики чувствительности в других областях зрительного спектра. Часто существует возможность обнаружить E-вектор поляризованного света. поляризация света.[11] Также могут быть дополнительные два или три глазка, которые помогают обнаруживать слабое освещение или небольшие изменения интенсивности света. Воспринимаемое изображение представляет собой комбинацию входных сигналов от многочисленных омматидий, расположенных на выпуклой поверхности и, таким образом, направленных в несколько разных направлениях. По сравнению с простыми глазами, сложные глаза обладают очень большими углами обзора и лучшей остротой, чем спинные глазки насекомых, но имеют несколько стволовых маток (= глаза личинок), например, у личинок пилильщика (Tenthredinidae ) с остротой зрения 4 градуса и очень высокой поляризационной чувствительностью, соответствует характеристикам сложного глаза.[12] [13]


Поскольку отдельные линзы настолько малы, эффекты дифракция накладывают ограничение на возможное разрешение, которое может быть получено (при условии, что они не работают как фазированные решетки ). Противостоять этому можно, только увеличив размер и количество линз. Чтобы видеть с разрешением, сопоставимым с нашими простыми глазами, людям потребовались бы сложные глаза, каждый из которых достигал бы размера их головы. Сложные глаза делятся на две группы: аппозиционные глаза, которые формируют несколько перевернутых изображений, и суперпозиционные глаза, которые образуют одно прямое изображение.[14][15] Сложные глаза растут по краям за счет добавления новых омматидиев.[16]
Антенны

Антенны, иногда называемые «щупальцами», представляют собой гибкие придатки, расположенные на голове насекомого, которые используются для восприятия окружающей среды. Насекомые находятся могут чувствовать антеннами из-за тонких волосков (щетинки ), которые их покрывают.[17]:8–11 Однако прикосновение - не единственное, что могут обнаружить антенны; многочисленные крошечные сенсорные структуры на антеннах позволяют насекомым ощущать запахи, температуру, влажность, давление и даже потенциально чувствовать себя в космосе.[17]:8–11[18][19] Некоторые насекомые, в том числе пчелы и некоторые группы мух, также могут улавливать звук с помощью своих антенн.[20]
Количество сегментов в антенне значительно варьируется среди насекомых, причем высшие мухи имеющий всего 3-6 сегментов,[21] пока взрослый тараканы может иметь более 140.[22] Общая форма усиков также весьма разнообразна, но первый сегмент (прикрепленный к голове) всегда называется скапусом, а второй сегмент - ножкой. Остальные членики усиков или жгутики называются жгутиком.[17]:8–11
Общие типы антенн насекомых показаны ниже:
 Aristate |  Capitate |  Клавиша |  Filiform |  Flabellate |  Коленчатый |  Щетинистый |
 Пластинчатый |  Монилиформ |  Пектинат |  Перистый |  Зубчатый |  Стилат |
Ротовой полости
Ротовой аппарат насекомых состоит из верхней челюсти, губ, а у некоторых видов - нижней челюсти.[8]:16[23] Верхняя губа представляет собой простой сросшийся склерит, часто называемый верхней губой, движется в продольном направлении и прикрепляется к наличнику. Нижние челюсти (челюсти) - это пара сильно склеротизированных структур, которые движутся под прямым углом к телу и используются для кусания, жевания и разрезания пищи. Верхнечелюстные кости - это парные образования, которые также могут двигаться под прямым углом к телу и имеют сегментированные щупики. Губная губа (нижняя губа) представляет собой сросшуюся структуру, движущуюся в продольном направлении и имеющую пару сегментированных щупиков.[24]

Легенда: а - усики
c - сложный глаз
фунт - губа
lr - верхняя губа
md - нижние челюсти
mx - максиллы
Ротовой аппарат вместе с остальной частью головы может быть сочленен по крайней мере в трех различных положениях: прогнатическом, опистогнатном и гипогнатическом. У видов с прогнатическим сочленением голова расположена вертикально на одном уровне с телом, например у видов Formicidae; в то время как у гипогнатного типа голова выровнена по горизонтали рядом с телом. Опистогнатная голова расположена по диагонали, например, у видов Blattodea и немного Жесткокрылые.[25] Ротовой аппарат сильно различается у насекомых разных отрядов, но две основные функциональные группы - нижнечелюстные и бугорчатые. Ротовой аппарат Haustellate используется для всасывания жидкостей и может быть дополнительно классифицирован по наличию стилеты, которые включают в себя пирсинг-сосание, обтирание и сифонирование. Стилеты представляют собой игольчатые выступы, используемые для проникновения в ткани растений и животных. Стилеты и питательный зонд образуют модифицированные нижние челюсти, верхнюю челюсть и гипофаринкс.[24]
- Мандибулят Ротовой аппарат, один из самых распространенных у насекомых, используется для откусывания и измельчения твердой пищи.
- Пирсинг-сосание В ротовой полости есть стилеты, которые используются для проникновения в твердые ткани и последующего всасывания жидкой пищи.
- Обтирание Ротовые органы используются для губки и всасывания жидкости и не имеют стилетов (например, у большинства двукрылых).
- Сифонирование в ротовом аппарате отсутствуют стилеты, они используются для сосания жидкостей и обычно встречаются среди видов Чешуекрылые.
Ротовой аппарат нижней челюсти встречается у видов Одоната, взрослый Neuroptera, Жесткокрылые, Перепончатокрылые, Blattodea, Прямокрылые, и Чешуекрылые. Однако у большинства взрослых чешуекрылых ротовой аппарат перекачивает, в то время как их личинки (обычно называемые гусеницы ) имеют челюсти.
Мандибулят
В верхняя губа представляет собой широкую лопасть, образующую крышу преоральной полости, подвешенную к наличнику перед ртом и образующую верхнюю губу.[1]:22–24 На внутренней стороне он перепончатый и может образовывать срединную долю, эпифаринкс, несущие некоторые сенсилла. Верхняя губа поднимается от нижних челюстей двумя мышцами, возникающими в голове, и вставляется медиально в передний край верхней губы. Он частично закрывается от нижних челюстей двумя мышцами, отходящими от головы и вставленными на задних боковых краях двух небольших склеритов, тормов, и, по крайней мере, у некоторых насекомых, резилиновой пружиной в кутикуле на стыке верхняя губа с наличником. [26] До недавнего времени считалось, что верхняя губа связана с первым сегментом головы. Однако недавние исследования эмбриологии, экспрессии генов и нервного питания верхней губы показывают, что она иннервируется тритоцеребрумом головного мозга, который представляет собой сросшиеся ганглии третьего сегмента головы. Он образован из слияния частей пары предковых придатков, обнаруженных на третьем сегменте головы, что показывает их взаимосвязь.[1]:22–24 Его вентральная, или внутренняя, поверхность обычно перепончатая и образует лопастевидный эпифаринкс, на котором расположены механосенсилла и хемосенсилла.[27][28]
У жевательных насекомых две челюсти, по одной с каждой стороны головы. Нижние челюсти расположены между верхней губой и максиллы. Челюсти режут и раздавливают пищу и могут использоваться для защиты; как правило, они имеют апикальную режущую кромку, а более базальная молярная область измельчает пищу. Они могут быть очень тяжелыми (около 3 Моос, или твердость вдавливания около 30 кг / мм2); таким образом, многие термиты и жуки не испытывают физических трудностей при просверливании фольги, сделанной из таких обычных металлов, как медь, свинец, олово и цинк.[1]:22–24 Режущие кромки обычно укрепляются добавлением цинка, марганца или, реже, железа в количестве до 4% от сухого веса.[27] Как правило, они являются самыми крупными ротовыми аппаратами жевательных насекомых, которые используются для пережевывания (разрезания, разрыва, дробления, жевания) пищевых продуктов. Они открываются наружу (по бокам головы) и сходятся кнутри. У плотоядных жевательных насекомых нижние челюсти могут быть изменены, чтобы стать более похожими на ножи, тогда как у травоядных жевательных насекомых они, как правило, широкие и плоские на противоположных лицах (например, гусеницы ). У мужчин жуки-олени, челюсти видоизменены до такой степени, что не выполняют никакой функции питания, а вместо этого используются для защиты мест спаривания от других самцов. В муравьи, нижние челюсти также выполняют защитную функцию (особенно в солдатских кастах). В муравьи-быки челюсти удлиненно-зубчатые, используются как охотничьи (и оборонительные) придатки.
Расположены под нижними челюстями, парные максиллы манипулировать едой во время жевание. Максиллы могут иметь волоски и «зубы» по внутреннему краю. На внешнем поле Галеа представляет собой чашевидную или совковидную структуру, которая находится над внешним краем губы. У них также есть щупальца, которые используются для определения характеристик потенциальных продуктов. Верхнечелюстные кости занимают боковые позиции, по одной с каждой стороны головы за нижними челюстями. Проксимальная часть верхней челюсти состоит из базального кардо, имеющего одно сочленение с головкой, и плоской пластинки, ножки, шарнирно соединенной с ней. Кардо. Кардо и ножки свободно соединены с головой мембраной, поэтому они могут двигаться. Дистально на ножках расположены две доли, внутренняя лацинея и внешняя галея, одна или обе из которых могут отсутствовать. Латеральнее на ножках находится сочлененный, похожий на ногу щупик, состоящий из нескольких сегментов; у прямокрылых их пять. Передние и задние вращающие мышцы вставляются на кардо, а вентральные приводящие мышцы, возникающие на тенториуме, вставляются как на кардо, так и на ножки. На ножках возникают мышцы-сгибатели lacinea и galea, а в черепе возникает еще один lacineal flexor, но ни lacinea, ни galea не имеют мышцы-разгибателя. Щупик имеет поднимающую и депрессорную мышцы, возникающие в ножках, и каждый сегмент щупика имеет одну мышцу, вызывающую сгибание следующего сегмента.[26]
В ротовом аппарате нижней челюсти губа представляет собой четвероногую структуру, хотя она образована двумя сросшимися вторичными челюстями. Его можно охарактеризовать как дно рта. С помощью верхних челюстей он помогает при манипуляциях с едой во время жевание или жевание, или, в необычном случае нимфа стрекозы, простирается, чтобы схватить добычу обратно к голове, где челюсти могут съесть ее. Губа похожа по строению на верхняя челюсть, но с придатками двух сторон, сливающимися по средней линии, так что они образуют срединную пластину. Базальная часть губ, эквивалентная кардинам верхней челюсти и, возможно, включающая часть грудины губного сегмента, называется постментумом. Его можно подразделить на проксимальный субментум и дистальный субментум. Дистальнее постментума и эквивалентно сросшимся ножкам верхней челюсти находится прементум. Прементум сзади закрывает предротовую полость. На конце он несет четыре лопасти, два внутренних лоскута и два внешних параглоссы, которые вместе известны как язычок. Одна или обе пары долей могут отсутствовать или они могут сливаться, образуя единый срединный отросток. С каждой стороны прементума возникает щупик, часто трехчлениковый.[26]
Гипофаринкс представляет собой срединную долю сразу за ртом, выступающую вперед из задней части предротовой полости; это доля неясного происхождения, но, возможно, связана с нижнечелюстным сегментом;[26] у аптеригот, уховерток и нимфальных поденок в гипофаринксе есть пара боковых долей - superlinguae (единственное число: superlingua). Он делит полость на дорсальный пищевой мешок, или цибарий, и вентральный слюнарий, в который открывается слюнный проток.[1]:22–24 Обычно он сливается с либием.[27] Большая часть гипофаринкса перепончатая, но адоральное лицо склеротизовано дистально и проксимально содержит пару поддерживающих склеритов, идущих вверх и заканчивающихся боковой стенкой устья. Мышцы, возникающие на лбу, вставляются в эти склериты, которые дистально соединены с парой язычных склеритов. Они, в свою очередь, вставили в них антагонистические пары мышц, возникающие на тенториуме и губе. Различные мышцы служат для раскачивания гипофаринкса вперед и назад, а у таракана еще две мышцы проходят через гипофаринкс и расширяют слюнное отверстие и расширяют слюнный канал.[26]
- Примеры нижних челюстей







Пирсинг-сосание
Ротовой аппарат может выполнять несколько функций. Некоторые насекомые сочетают проникающие части с губками, которые затем используются для прокалывания тканей растений и животных. Самки комаров питаются кровью (гемофаг ) делая их переносчиками болезней. Ротовой аппарат комара состоит из хоботка, парных нижних челюстей и верхних челюстей. Верхнечелюстные кости образуют игольчатые образования, называемые стилеты, которые окружены губами. Когда комар кусает, верхнечелюстные кости проникают в кожу и фиксируют ротовой аппарат, позволяя вставить другие части. Губная губа, похожая на оболочку, отодвигается назад, а оставшиеся части рта проходят через ее кончик в ткани. Затем через гипофаринкс комар вводит слюна, который содержит антикоагулянты чтобы предотвратить свертывание крови. И, наконец, верхняя губа (верхняя губа) используется для всасывания крови. Виды рода Анофелес для них характерны длинные щупики (две части с расширяющимся концом), почти доходящие до конца верхней губы.[29]
- Примеры пирсинга в ротовой полости

Слепень (женский)

Поставить галочку (Ixodes ricinus ), которое не насекомое, а паукообразный, данные для сравнения





Сифонирование
В хоботок сформирован из верхнечелюстные галеи и есть приспособление, обнаруженное у некоторых насекомых для сосания.[30] Мышцы цибария или глотки сильно развиты и образуют насос.У Hemiptera и многих Diptera, которые питаются жидкостями растений или животных, некоторые компоненты ротового аппарата модифицированы для прокалывания, и удлиненные структуры называются стилетами. Комбинированные трубчатые структуры называют хоботком, хотя в некоторых группах используется специальная терминология.
У видов чешуекрылых он состоит из двух трубок, соединенных крючками и отделяемых для очистки. Каждая трубка вогнута внутрь, образуя центральную трубку, через которую всасывается влага. Всасывание осуществляется за счет сжатия и расширения мешка в голове.[31] Хоботок свернут под головой, когда насекомое находится в состоянии покоя, и расширяется только при кормлении.[30] Верхнечелюстные щупики уменьшены или даже рудиментарны.[32] Они бросаются в глаза и имеют пять сегментов в некоторых базальных семействах и часто складчатые.[8] Форма и размеры хоботка эволюционировали, чтобы дать разным видам более широкий и, следовательно, более выгодный рацион.[30] Существует аллометрический масштабная взаимосвязь между массой тела Lepidoptera и длиной хоботка[33] от которого интересным адаптивным отличием является необычно длинноязычная ястребиная моль Xanthopan morganii praedicta. Чарльз Дарвин предсказал существование и длину хоботка этой бабочки до ее открытия, основываясь на своих знаниях о мадагаскарской звездной орхидее с длинными шипами. Angraecum sesquipedale.[34]
- Примеры сифонирования ротовой полости




Обтирание
Ротовой аппарат насекомых, питающихся жидкостями, по-разному видоизменяется, образуя трубку, через которую жидкость может втягиваться в рот, и обычно еще одну, через которую проходит слюна. Мышцы цибария или глотки сильно развиты и образуют насос.[26] У не кусающих мух нижние челюсти отсутствуют, а другие структуры редуцированы; лабиальные щупики видоизменяются, образуя губную губу, и верхнечелюстные щупики присутствуют, хотя иногда и короткие. У Brachycera губа особенно заметна и используется для обтирания жидкой или полужидкой пищи.[35] В Лабелла представляют собой сложную структуру, состоящую из множества бороздок, называемых псевдотрахеями, которые всасывают жидкости. Слюнные выделения лабеллы способствуют растворению и сбору частиц пищи, так что они могут легче поглощаться псевдотрахеями; Считается, что это происходит за счет капиллярного действия. Жидкая пища затем всасывается из псевдотрахей через пищевой канал в пищевод.[36]
Ротовой аппарат пчелы бывают жевательного и притирочно-сосущего типа. Притирка - это режим кормления, при котором жидкая или полужидкая пища, приставшая к выступающему органу или «языку», переносится с субстрата в рот. в пчела (Перепончатокрылые: Apidae: Apis mellifera ), удлиненные и слившиеся губные лоскуты образуют волосатый язык, который окружен верхнечелюстными галеями и губными щупиками, образуя трубчатый хоботок, содержащий канал питания. Во время кормления язык окунается в нектар или мед, который прилипает к волоскам, а затем втягивается, так что приставшая жидкость переносится в пространство между галлеями и губными щупиками. Это возвратно-поступательное движение происходит неоднократно. Движение жидкости ко рту, по-видимому, является результатом действия цибариального насоса, которому способствует каждое втягивание языка, выталкивающее жидкость вверх по пищевому каналу.[1]:22–24
- Примеры обтирания ротовой полости


Грудная клетка
Насекомое грудная клетка состоит из трех сегментов: переднегрудь, среднегрудь, и заднегрудь. В передний ближайший к голове сегмент - переднегрудь; его основные особенности - первая пара ног и переднеспинка. Средний сегмент - это среднегрудь; его основные особенности - вторая пара ног и передние крылья, если таковые имеются. Третий, задний, грудной сегмент, упирающийся в брюшко, - это заднегрудь, несущая третью пару ног и задние крылья. Каждый сегмент расширен межсегментарным швом. Каждый сегмент состоит из четырех основных регионов. Дорсальная поверхность называется тергумом (или notum, чтобы отличать его от терги брюшка).[1]:22–24 Две боковые области называются плеврой (единственное число: плеврон), а вентральная часть называется грудиной. В свою очередь, задняя часть переднегруди называется переднеспинкой, задняя часть среднегруди - среднеспинкой, а заднегрудь - заднегруди. Продолжая эту логику, есть также мезоплевра и метаплевра, а также мезостерна и заднегрудь.[8]
Тергальные пластинки грудной клетки представляют собой простые структуры у аптериго и многих неполовозрелых насекомых, но у крылатых взрослых особей они видоизменяются по-разному. Каждая птероторакальная нота имеет два основных отдела: передний, несущий крыло алинотум, и задний, несущий фрагму постнотум. Phragmata (единственное число: phragma) - это пластинчатые аподемы, которые проходят внутрь под анекостальными швами, отмечая первичные межсегментарные складки между сегментами; phragmata обеспечивают прикрепление мышц продольного полета. Каждый алинотум (иногда ошибочно называемый «notum») может быть пересечен швами, отмечающими положение внутренних укрепляющих гребней, и обычно делит пластину на три области: передний прескутум, скутум и меньший задний щиток. Считается, что боковые плевральные склериты произошли от субкоксального сегмента ноги предкового насекомого. Эти склериты могут быть отдельными, как у чешуйницы, или слитыми в почти непрерывную склеротическую область, как у большинства крылатых насекомых.[1]:22–24
Переднегрудь
Переднеспинка переднегруди может быть простой по строению и небольшой по сравнению с другими нотами, но у жуков, богомолов, многих клопов и некоторых прямокрылых переднеспинка расширена, а у тараканов она образует щит, закрывающий часть тела. голова и среднегрудь.[8][1]:22–24
Птероторакс
Поскольку мезоторакс и заднегрудь удерживают крылья, у них есть общее название, называемое птеротораксом (птерон = крыло). Переднее крыло, которое носит разные названия в разных отрядах (например, надкрылья у прямокрылых и надкрылья у жесткокрылых), возникает между мезонотумом и мезоплеврой, а заднее крыло сочленяется между заднеспинкой и метаплеврой. Ноги возникают из мезоплевры и метаплевры. Мезоторакс и заднегрудь имеют плевральный шов (мезоплевральный и метаплевральный швы), идущий от основания крыла до тазика ноги. Склерит перед плевральным швом называется эпистерном (поочередно мезэпистерна и метэпистерна). Склерит кзади от шва называется эпимироном (поочередно мезепимироном и метепимироном). Дыхальца, внешние органы дыхательной системы, находятся на птеротораксе, обычно одно между про- и мезоплеороном, а также одно между мезо- и метаплевроном.[8]
Вентральный вид, или грудина, соответствует тому же условию: переднегрудь находится под переднегрудью, среднегрудь - под среднегрудью, а заднегрудь - под заднегруди. На спине, плевре и грудины каждого сегмента имеется множество различных склеритов и швов, которые сильно различаются от порядка к порядку, и они не будут подробно обсуждаться в этом разделе.[8]
Крылья
У большинства филогенетически развитых насекомых есть две пары крылья расположены на втором и третьем грудных сегментах.[1]:22–24 Насекомые - единственные беспозвоночные развили летные способности, и это сыграло важную роль в их успехе. Полет насекомых не очень хорошо изучен, в значительной степени полагаясь на турбулентные аэродинамические эффекты. Примитивные группы насекомых используют мышцы, которые действуют непосредственно на структуру крыльев. Более продвинутые группы, составляющие Neoptera имеют складные крылья, а их мышцы действуют на стенку грудной клетки и опосредованно приводят в действие крылья.[1]:22–24 Эти мышцы способны сокращаться несколько раз для каждого нервного импульса, позволяя крыльям биться быстрее, чем это было бы обычно.
Полет насекомых может быть чрезвычайно быстрым, маневренным и универсальным, возможно, из-за меняющейся формы, исключительного контроля и переменного движения крыла насекомого. Отряды насекомых используют разные механизмы полета; например, полет бабочки можно объяснить с помощью стационарного, непреходящего аэродинамика, и тонкий профиль теория.

Внутренний
Каждое крыло состоит из тонкой мембраны, поддерживаемой системой жилок. Мембрана образована двумя плотно прилегающими слоями покровов, в то время как вены образуются там, где два слоя остаются отдельными, а кутикула может быть толще и более склеротизированной. Внутри каждой из основных вен находится нерв и трахея, и, поскольку полости вен связаны с гемоцель, гемолимфа может течь в крылья.[26] По мере развития крыла дорсальный и вентральный покровные слои на большей части своей площади сближаются, образуя перепонку крыла. Остальные участки образуют каналы, будущие вены, в которых могут проходить нервы и трахеи. Кутикула, окружающая вены, утолщается и склеротизируется в большей степени, что обеспечивает прочность и жесткость крыла. На крыльях могут встречаться волоски двух типов: микротрихии, которые мелкие и неравномерно разбросаны, и макротрихии, которые более крупные, с углублениями и могут ограничиваться жилками. Чешуйки Lepidoptera и Trichoptera - сильно модифицированные макротрихии.[27]
Вен

У некоторых очень мелких насекомых жилкование может быть значительно сокращено. В халкидные осы, например, присутствуют только подреберья и часть лучевой кости. И наоборот, увеличение жилкования может происходить из-за разветвления существующих жилок с образованием дополнительных жилок или за счет развития дополнительных вставочных жилок между исходными, как в крыльях Прямокрылые (кузнечики и сверчки). У некоторых насекомых имеется большое количество поперечных жилок, и они могут образовывать сетку, как на крыльях Одоната (стрекозы и стрекозы) и у основания передних крыльев Tettigonioidea и Acridoidea (катидиды и кузнечики соответственно).[26]
Арчедиктион - это название гипотетической схемы жилкования крыльев, предложенной для самого первого крылатого насекомого. Он основан на сочетании предположений и данных об ископаемых. Поскольку считается, что все крылатые насекомые произошли от общего предка, аркедикация представляет собой «шаблон», который был изменен (и упорядочен) естественным отбором в течение 200 миллионов лет. Согласно современным догмам, аркедиктион содержал от шести до восьми продольных жилок. Эти жилы (и их ветви) названы в соответствии с системой, разработанной Джоном Комстоком и Джорджем Нидхэмом - системой Комстока-Нидхэма:[37]
- Коста (К) - передняя кромка крыла
- Subcosta (Sc) - вторая продольная жилка (за костой), как правило, неразветвленная
- Радиус (R) - третья продольная жилка, от одной до пяти ветвей доходят до края крыла
- СМИ (M) - четвертая продольная жилка, от одной до четырех ветвей доходят до края крыла
- Кубитус (Cu) - пятая продольная жилка, от одной до трех ветвей доходят до края крыла
- Анальные вены (A1, A2, A3) - неразветвленные вены за локтем
Коста (C) является ведущей краевой жилкой у большинства насекомых, хотя небольшая жилка, прекоста, иногда встречается над костой. Практически у всех существующих насекомых[1]:41–42 прекоста сливается с костой; коста редко когда-либо разветвляется, потому что она находится на переднем крае, который у своего основания связан с плечевой пластиной. Трахея реберной вены - это, возможно, ветвь подреберной трахеи. После косты расположена третья вена, subcosta, которая разветвляется на две отдельные вены: переднюю и заднюю. Основание подреберья связано с дистальным концом шейки первой подмышечной впадины. Четвертая вена - лучевая, которая разветвляется на пять отдельных жилок. Радиус, как правило, является самой сильной жилкой крыла. Ближе к середине крыла он разветвляется на первую нераздельную ветвь (R1) и вторую ветвь, называемую радиальным сектором (Ra), которая дихотомически подразделяется на четыре дистальных ветви (R2, R3, R4, R5). В основном радиус гибко соединяется с передним концом второй подмышечной впадины (2Ax).[38]
Пятая жила крыла - СМИ. В шаблоне архетипа (A) среда разветвляется на две основные ветви: переднюю среду (MA), которая делится на две дистальные ветви (MA1, MA2), и срединный сектор, или заднюю среду (MP), которая имеет четыре концевые ответвления (М1, М2, М3, М4). У большинства современных насекомых передняя среда утрачена, и обычная «среда» представляет собой четырехветвленную заднюю среду с общим базальным стеблем. У Ephemerida, согласно современным интерпретациям жилкования крыла, сохраняются обе ветви медиальной части, в то время как у Odonata сохраняющаяся медиа является примитивной передней ветвью. Ствол медианы часто соединяется с лучевой костью, но когда он встречается как отдельная вена, его основание связано с дистальной срединной пластиной (m ') или непрерывно склеротизируется с последней. Cubitus, шестая жилка крыла, в основном двухветвистая. Первичная развилка происходит около основания крыла, образуя две основные ветви (Cu1, Cu2). Передняя ветвь может распадаться на несколько вторичных ветвей, но обычно она разветвляется на две дистальные ветви. Вторую ветвь кубита (Cu2) у перепончатокрылых, трихоптерых и чешуекрылых Комсток и Нидхэм ошибочно приняли за первый анальный. Проксимально главный ствол кубита связан с дистальной срединной пластиной (m ') основания крыла.[38]
Посткубитус (Pcu) - первый анал системы Комстока и Нидхэма. Postcubitus, однако, имеет статус самостоятельной жилки крыла и должен быть признан таковой. В крыльях нимфы ее трахея возникает между локтевой трахеей и группой ваннальных трахей. В зрелых крыльях более распространенных насекомых postcubitus всегда связан проксимально с локтевым суставом и никогда близко не связан со склеритом-сгибателем (3Ax) основания крыла. У Neuroptera, Mecoptera и Trichoptera посткубитус может быть более тесно связан с ванными венами, но его основание всегда свободно от последних. Посткубитус обычно неразветвленный; примитивно он двухветвленный. Ваннальные вены (от lV до nV) - это анальные вены, непосредственно связанные с третьей подмышечной впадиной, и на которые непосредственно влияет движение этого склерита, вызывающее сгибание крыльев. Число сосудистых жилок варьирует от одной до 12 в зависимости от расширения ваннальной площади крыла. Ваннальные трахеи обычно возникают из общего трахеального ствола у нимфальных насекомых, а вены считаются ветвями одной анальной вены. Дистально сосудистые вены либо простые, либо разветвленные. Югальная вена (J) скуловой доли крыла часто занята сетью неправильных вен или может быть полностью перепончатой; иногда он содержит одну или две отдельные маленькие вены, первую скуловую вену, или дуговую вену, и вторую скуловую вену, или кардинальную вену (2J).[38]
- C-Sc поперечные вены - пробегать между костой и субкостой
- R поперечные вены - пробежать между соседними ветвями радиуса
- Поперечные вены R-M - бегать между радиусом и носителем
- Поперечные вены M-Cu - бегать между медиа и кубитусом
Все жилки крыла подвержены вторичному раздвоению и соединению поперечными жилками. У некоторых отрядов насекомых поперечные жилки настолько многочисленны, что весь рисунок жилкования превращается в плотную сеть ветвящихся и поперечных жилок. Однако обычно встречается определенное количество поперечных жилок, имеющих определенные места. Более постоянные поперечные вены - это плечевая поперечная вена (h) между ребрами и подреберьями, радиальная поперечная вена (r) между R и первой вилкой Rs, секториальная поперечная вена (и) между двумя развилками. из R8, средняя поперечная вена (мм) между M2 и M3 и медиокубитальная поперечная вена (m-cu) между средой и локтевым суставом.[38]
Вены на крыльях насекомых характеризуются выпукло-вогнутым расположением, как у поденок (т. Е. Вогнутое - «вниз», а выпуклое - «вверх»), которые регулярно чередуются и ветвятся; всякий раз, когда вена разветвляется, между двумя ветвями всегда есть вставная вена противоположного положения. Вогнутая жила разделится на две вогнутые жилы (при этом интерполированная жила будет выпуклой), и регулярное изменение вен сохраняется.[39] Кажется, что жилки крыла образуют волнообразный узор в зависимости от того, имеют ли они тенденцию складываться вверх или вниз, когда крыло расслаблено. Базальные стержни вен выпуклые, но каждая жилка дистально разветвляется на переднюю выпуклую ветвь и заднюю вогнутую ветвь. Таким образом, реберные и субкоста считаются выпуклой и вогнутой ветвями первичной первой жилки, Rs - вогнутой ветвью радиуса, задняя среда - вогнутой ветвью среды, Cu1 и Cu2 соответственно выпуклой и вогнутой, в то время как примитивный посткубитус и первый ваннал имеет переднюю выпуклую ветвь и заднюю вогнутую ветвь. Выпуклая или вогнутая природа жилок использовалась в качестве доказательства при определении идентичности сохраняющихся дистальных ветвей жилок современных насекомых, но не было продемонстрировано, что она одинакова для всех крыльев.[26][38]
Поля

Зоны крыла разделены и подразделяются на линии сгиба, вдоль которого складываются крылья, и линии сгибания, которые прогибаются во время полета. Основное различие между линиями сгибания и сгиба часто нечеткое, поскольку линии сгиба могут допускать некоторую гибкость или наоборот. Две константы, присутствующие почти во всех крыльях насекомых, - это ключица (линия сгибания) и скуловая складка (или линия сгиба), образующие изменчивые и неудовлетворительные границы. Складывание крыльев может быть очень сложным, при этом поперечное складывание происходит в задних крыльях. Dermaptera и Coleoptera, а у некоторых насекомых анальная область может складываться веером.[1]:41–42На крыльях насекомых есть четыре разных поля:
- Ремигиум
- Анальная область (vannus)
- Югальная область
- Подмышечная область
- Алула
Большинство вен и поперечных вен встречается в передней области ремигиум, который отвечает за большую часть полета, приводится в действие грудными мышцами. Заднюю часть ремигиума иногда называют булавой; два других задних поля - анальное и скуловые области.[1]:41–42 Когда сосудистая складка занимает обычное положение впереди группы анальных вен, ремигиум содержит реберные, подреберные, лучевые, медиальные, локтевые и посткубитальные вены. В согнутом крыле ремигиум поворачивается кзади на гибком базальном соединении лучевой кости со второй подмышечной впадиной, а основание медиокубитального поля загибается медиально на подмышечной области вдоль базальной складки (bf) между срединными пластинами (m, м ') основания крыла.[38]
Ваннус окаймлен ванной складкой, которая обычно находится между посткубитом и первой ванной жилкой. У прямокрылых оно обычно занимает такое положение. Однако у переднего крыла Blattidae единственная складка в этой части крыла лежит непосредственно перед посткубитусом. У Plecoptera ваннальная складка находится кзади от postcubitus, но проксимально пересекает основание первой ванальной жилки. У цикад ваннальная складка лежит сразу за первой сосудистой жилкой (lV). Эти небольшие вариации фактического положения сосудистой складки, однако, не влияют на единство действия сосудистых вен, контролируемых сгибающим склеритом (3Ax), при сгибании крыла. У большинства прямокрылых на задних крыльях вторичная делительная вена образует ребро в ваннальной складке. Ваннус обычно имеет треугольную форму, а его жилки обычно расходятся от третьей подмышечной впадины, как ребра веера. Некоторые из сосудистых вен могут быть разветвленными, а вторичные вены могут чередоваться с первичными. Vannal область, как правило, лучше всего развита в hindwing, в котором она может быть увеличена, чтобы сформировать поверхность на поддержание производства, как и в Plecoptera и Orthoptera. Большие веерообразные расширения задних крыльев Acrididae, несомненно, являются ванными областями, поскольку все их жилки поддерживаются на третьих подмышечных склеритах на основаниях крыльев, хотя Мартынов (1925) приписывает большую часть веерных областей у Acrididae скуловым областям. крыльев. Истинный jugum крыла саранчовых представлен только мезадом малой мембраны (Ju) последней вочной жилки. Jugum более развита у некоторых других прямокрылых, например у мантид. У большинства высших насекомых с узкими крыльями vannus становится редуцированным, и ваннальная складка утрачивается, но даже в таких случаях изогнутое крыло может изгибаться по линии между postcubitus и первой сосудистой жилкой.[38]
Скуловая область, или neala, - это область крыла, которая обычно представляет собой небольшую перепончатую область проксимальнее основания ваннуса, усиленную несколькими небольшими, неправильными веноподобными утолщениями; но когда он хорошо развит, он представляет собой отдельную часть крыла и может содержать одну или две скуловые вены. Когда скуловая область переднего крыла развита как свободная доля, она выступает под углом плечевой кости заднего крыла и, таким образом, служит для соединения двух крыльев вместе. В группе Lepidoptera Jugatae имеет длинную пальцевидную лопасть. Скуловая область была названа neala («новое крыло»), потому что это, очевидно, вторичная и недавно разработанная часть крыла.[38]
Вспомогательная область, содержащая подмышечные склериты, в целом имеет форму разностороннего треугольника. Основание треугольника (a-b) - шарнир крыла с корпусом; вершина (с) - дистальный конец третьего подмышечного склерита; более длинная сторона находится кпереди от вершины. Точка d на передней стороне треугольника отмечает сочленение лучевой вены со вторым подмышечным склеритом. Линия между d и c - это plica basalis (bf), или складка крыла у основания медиокубитального поля.[38]
На заднем углу основания крыла у некоторых двукрылых есть пара перепончатых лопастей (чешуйчатых или чашеобразных), известных как alula. Алюла хорошо развита у домашней мухи. Наружная чешуйка (c) возникает от основания крыла за третьим подмышечным склеритом (3Ax) и, очевидно, представляет собой скуловую лопасть других насекомых (A, D); более крупная внутренняя чешуя (d) отходит от заднего щиткового края тергума крыловидного сегмента и образует защитный капюшон-навес над недоуздком. В согнутом крыле внешняя чешуйка alula перевернута вверх дном над внутренней чешуей, на которую движение крыла не влияет. У многих двукрылых глубокий разрез анальной области перепонки крыла позади единственной ваннальной жилки выделяет проксимальную крыловую лопасть, дистальнее внешней чешуи крыльев.[38]
Суставы

Различные движения крыльев, особенно у насекомых, которые в состоянии покоя сгибают крылья горизонтально над спиной, требуют более сложной суставной структуры у основания крыла, чем простое соединение крыла с телом. Каждое крыло прикреплено к телу перепончатой базальной областью, но суставная перепонка содержит ряд небольших суставных склеритов, вместе известных как птералии. Птералии включают переднюю плечевую пластинку у основания реберной вены, группу подмышечных впадин (Ax), связанных с подреберной, лучевой и ваннальной венами, и две менее выраженные срединные пластинки (m, m ') у основания реберной вены. медиокубитальная область. Подмышечные впадины особенно развиты только у насекомых, сгибающих крыло, где они составляют сгибательный механизм крыла, управляемый сгибающей мышцей, возникающей на плевре. Для основания крыла также характерна небольшая лопасть на переднем крае суставной области проксимальнее плечевой пластинки, которая в переднем крыле некоторых насекомых превращается в большой плоский чешуйчатый лоскут, тегула, перекрывающий друг друга. основание крыла. Сзади суставная перепонка часто образует обширную лопасть между крылом и телом, а ее край обычно утолщен и гофрирован, создавая вид связки, так называемого подмышечного канатика, непрерывного мезально с задней маргинальной складкой щитка сустава. тергальная пластинка, несущая крыло.[38]
Суставные склериты, или птералии, основания крыльев насекомых с изгибающимися крыльями и их отношения к телу и жилкам крыльев, показанные схематически, следующие:
- Плечевые пластины
- Первая подмышечная
- Второй подмышечный
- Третья подмышечная
- Четвертый подмышечный
- Срединные пластины (м, м ')
Плечевая пластинка обычно представляет собой небольшой склерит на переднем крае основания крыла, подвижный и сочлененный с основанием реберной вены. У Odonata плечевые пластинки сильно увеличены,[38] с двумя мышцами, отходящими от эпистерна, вставленными в плечевые пластинки, и двумя мышцами, отходящими от края эпимера, вставленными в подмышечную пластинку.[26]
Первый подмышечный склерит (lAx) представляет собой переднюю шарнирную пластину основания крыла. Его передняя часть опирается на передний нотальный крыловой отросток тергума (ПНП); его задняя часть сочленяется с краем тергала. Передний конец склерита обычно образуется в виде тонкого плеча, вершина которого (е) всегда связана с основанием подреберной вены (Sc), хотя и не соединяется с последней. Тело склерита латерально сочленяется со второй подмышечной впадиной. Второй подмышечный склерит (2Ax) более изменчив по форме, чем первый подмышечный, но его механические отношения не менее определенны. Она наклонно шарнирно прикреплена к внешнему краю тела первой подмышечной впадины, а радиальная вена (R) всегда гибко прикрепляется к ее переднему концу (d). Вторая подмышечная впадина представляет собой как дорсальную, так и вентральную склеротизацию в основании крыла; его вентральная поверхность опирается на опорный отросток крыла плеврона. Вторая подмышечная впадина, таким образом, является стержневым склеритом основания крыла, и она специально воздействует на радиальную жилку.[38]
Третий подмышечный склерит (3Ax) лежит в задней части суставной области крыла. Его форма очень вариабельна и часто нерегулярна, но третья подмышечная впадина - это склерит, в который вставлен сгибатель крыла (D). Мезально он сочленяется кпереди (f) с задним концом второй подмышечной впадины и сзади (b) с задним крыловым отростком тергума (PNP) или с небольшой четвертой подмышечной впадиной, когда последний присутствует. Дистально третья подмышечная впадина продолжается в отростке, всегда связанном с основанием группы жилок в анальной области крыла, здесь называемых ванными жилками (V). Таким образом, третья подмышечная впадина обычно является задней шарнирной пластиной основания крыла и является активным склеритом сгибательного механизма, который непосредственно воздействует на сосудистые вены. Сокращение мышцы-сгибателя (D) вращает третью подмышечную мышцу на ее мезальных сочленениях (b, f) и тем самым поднимает ее дистальную руку; это движение вызывает сгибание крыла. Четвертый подмышечный склерит не является постоянным элементом основания крыла. Когда он присутствует, он обычно представляет собой небольшую пластинку между третьей подмышечной частью и задним нотальным отростком крыла и, вероятно, является отдельной частью последнего.[38]
Срединные пластинки (m, m ') также являются склеритами, которые не так однозначно дифференцируются как специфические пластины, как три основных подмышечных мышцы, но они являются важными элементами сгибающего аппарата. Они лежат в средней области основания крыла, дистальнее второй и третьей подмышек, и отделены друг от друга наклонной линией (bf), которая образует заметную выпуклую складку во время сгибания крыла. Проксимальная пластинка (m) обычно прикрепляется к дистальному концу третьей подмышечной впадины и, возможно, должна рассматриваться как часть последней. Дистальная пластинка (m ') менее постоянно присутствует как отдельный склерит и может быть представлена общей склеротизацией основания медиокубитального поля крыла. Когда жилки этой области отчетливы в основании, они связаны с внешней срединной пластиной.[38]
Сцепление, складывание и другие особенности
У многих видов насекомых переднее и заднее крыло соединены вместе, что улучшает аэродинамическую эффективность полета. Наиболее распространенный механизм сцепления (например, Перепончатокрылые и Трихоптера ) представляет собой ряд маленьких крючков на переднем крае заднего крыла, или «хамули», которые фиксируются на переднем крыле, удерживая их вместе (хамулатное сцепление). У некоторых других видов насекомых (например, Mecoptera, Чешуекрылые, и немного Трихоптера ) скуловая доля переднего крыла покрывает часть заднего крыла (соединение скуловой кости), или края переднего крыла и заднего крыла перекрываются широко (соединение в форме амплификации), или щетинки заднего крыла, или уздечка, зацепляются под удерживающей структурой или сетчаткой на сетчатке. переднее крыло.[1]:43
В состоянии покоя крылья у большинства насекомых удерживаются над спиной, что может включать продольное складывание перепонки крыла, а иногда и поперечное складывание. Иногда может происходить складывание по линиям сгибания.Хотя линии сгиба могут быть поперечными, как в задних крыльях жуков и уховерток, они обычно радиальны по отношению к основанию крыла, что позволяет сгибать соседние части крыла друг над другом или под ним. Самая распространенная линия складки - скуловая складка, расположенная сразу за третьей анальной веной.[27] хотя у большинства Neoptera имеется скуловая складка сразу за жилкой 3A на передних крыльях. Иногда он также присутствует на задних крыльях. Там, где анальная область заднего крыла велика, как у Orthoptera и Blattodea, вся эта часть может быть загнута под передней частью крыла вдоль ваннальной складки немного позади ключичной борозды. Кроме того, у Orthoptera и Blattodea анальная область свернута веером вдоль вен, причем анальные вены выпуклые, на гребнях складок, а добавочные вены вогнуты. В то время как булавочная борозда и скуловая складка, вероятно, гомологичны у разных видов, ваннальная складка различается по положению у разных таксонов. Складывание происходит за счет мышцы, возникающей на плевроне и вставляемой в третий подмышечный склерит таким образом, что при сокращении склерит поворачивается вокруг точек сочленения с задним нотальным отростком и вторым подмышечным склеритом.[26]
В результате дистальное плечо третьего подмышечного склерита поворачивается вверх и внутрь, так что в конечном итоге его положение полностью меняется. Анальные вены соединяются с этим склеритом таким образом, что, когда он движется, они переносятся с ним и сгибаются над спиной насекомого. Активность одной и той же мышцы в полете влияет на мощность крыла, поэтому она также важна для управления полетом. У ортоптероидных насекомых эластичность кутикулы заставляет ванную часть крыла складываться вдоль жилок. Следовательно, при перемещении крыльев в положение полета на раскрытие этой области расходуется энергия. В общем, разгибание крыла, вероятно, является результатом сокращения мышц, прикрепленных к базилярному склериту или, у некоторых насекомых, к субалярному склериту.[26]
Ноги
Типичные и обычные сегменты нога насекомого делятся на тазики, один вертел бедра, голени, предплюсны и претарсус. Тазик в своей более симметричной форме имеет форму короткого цилиндра или усеченного конуса, хотя обычно он яйцевидной формы и может быть почти сферическим. Проксимальный конец тазика опоясан субмаргинальной базисный шов, образующий внутри гребень, или Basicosta, и выделяет крайний фланец, Coxomarginale, или же базикоксит. В Basicosta укрепляет основание тазика и обычно увеличивается на внешней стенке для прикрепления мышц; однако на мезальной половине тазика он обычно слаб и часто сливается с краем тазика. Вертельные мышцы, берущие свое начало в тазике, всегда прикрепляются дистальнее базикосты. Тазик прикреплен к телу суставной перепонкой, кориумом тазика, который окружает его основание. Эти два сочленения, возможно, являются основными дорсальными и вентральными суставными точками подкоксо-коксального шарнира. Кроме того, тазик насекомых часто имеет переднее сочленение с передним брюшным концом трочантина, но трокантинальное сочленение не сосуществует с грудным сочленением. Плевральная суставная поверхность тазика ложится на мезальный перегиб стенки тазика. Если тазик подвижен только на плевральном сочленении, то суставная поверхность тазика обычно изгибается на достаточную глубину, чтобы задействовать отводящие мышцы, вставленные на внешний край основания тазика. Дистально тазик имеет переднее и заднее сочленение с вертелом. Наружная стенка тазика часто обозначена швом, идущим от основания до переднего вертлужного сочленения. У некоторых насекомых тазовый шов совпадает с плевральным швом, и в таких случаях тазик, по-видимому, разделен на две части, соответствующие эпистерну и эпимерону плевры. Коксальный шов у многих насекомых отсутствует.[38]:163–164
Перегиб стенки тазика, несущей плевральную суставную поверхность, разделяет боковую стенку базикоксита на предсуставную часть и постартикулярную часть, и эти две области часто выглядят как две маргинальные доли на основании тазика. Задняя доля обычно больше и называется мерон. Мерон может быть значительно увеличен за счет расширения дистально в задней стенке тазика; у Neuroptera, Mecoptera, Trichoptera и Lepidoptera мерон настолько велик, что кажется, что тазик разделен на переднюю часть, так называемую «coxa genuina» и мерон, но мерон никогда не включает область заднее вертельное сочленение и ограничивающая его борозда всегда является частью базикостального шва. Тазик с увеличенным мероном имеет внешний вид, похожий на тот, который разделен швом тазика, совпадающим с плевральным швом, но эти два состояния в корне совершенно разные, и их не следует путать. Мерон достигает предела своего отклонения от обычного состояния двукрылых. У некоторых более распространенных мух, таких как Tipulidae, мерон средней ноги выглядит как большая лопасть тазика, выступающая вверх и кзади от основания тазика; у высших представителей отряда он полностью отделяется от тазика и образует пластинку боковой стенки среднегрудного отдела.[38]:164
Вертел - базальный сегмент телоподита; это всегда небольшой сегмент ноги насекомого, свободно перемещаемый посредством горизонтального шарнира на тазике, но более или менее прикрепленный к основанию бедра. При перемещении на бедренной кости шарнир вертела бедра обычно вертикальный или наклонный в вертикальной плоскости, что дает небольшое движение производства и сокращения в суставе, хотя присутствует только редукторная мышца. У Odonata, как у нимфы, так и у взрослых особей, есть два вертлужных сегмента, но они неподвижны друг относительно друга; вторая содержит мышцу-редуктор бедренной кости. Обычный единственный вертельный сегмент насекомых, поэтому, вероятно, представляет собой два вертела других членистоногих, слитых в один видимый сегмент, поскольку маловероятно, что первичный тазотреловой шарнир был утерян с ноги. У некоторых перепончатокрылых базальная часть бедренной кости имитирует второй вертел, но прикрепление мышцы-редуктора к его основанию свидетельствует о том, что она принадлежит к бедренному сегменту, поскольку, как показано на зубчатой ножке, редуктор берет свое начало в истинный второй вертел.[38]:165
В бедренная кость это третий сегмент ноги насекомого, обычно самая длинная и самая сильная часть конечности, но он варьируется по размеру от огромного заднего бедра прыгающих прямокрылых до очень маленького сегмента, который присутствует у многих личиночных форм. Объем бедренной кости обычно коррелирует с размером большеберцовых мышц, содержащихся в ней, но иногда она увеличивается и изменяется по форме для других целей, кроме целей размещения большеберцовых мышц. В большеберцовая кость обычно представляет собой тонкий сегмент у взрослых насекомых, лишь немного короче бедренной кости или бедренной кости вместе с вертелом. Его проксимальный конец образует более или менее отчетливую головку, изогнутую к бедренной кости, устройство, позволяющее большеберцовой кости сгибаться вплотную к нижней поверхности бедренной кости.[38]:165
Термины profemur, mesofemur и metafemur относятся к бедрам передних, средних и задних ног насекомого соответственно.[40] Аналогичным образом протибия, мезотибия и метатибия относятся к большеберцовой кости передних, средних и задних ног.[41]
В предплюсна насекомых соответствует предпоследнему сегменту конечности обобщенного членистоногого, который у ракообразных называется проподитом. У взрослых насекомых он обычно подразделяется на от двух до пяти подсегментов, или тарсомеров, но у Protura, некоторых Collembola и большинства голометаболических личинок насекомых он сохраняет примитивную форму простого сегмента. Подотрезки предплюсны взрослого насекомого обычно свободно перемещаются друг по другу за счет перегиба соединительных перепонок, но предплюсны никогда не имеют внутренних мышц. Предплюсна взрослых крылатых насекомых, имеющих менее пяти субучастков, вероятно, специализируется на потере одного или нескольких субучастков или в результате слияния смежных субучастков. В лапках Acrididae длинная базальная часть, очевидно, состоит из трех объединенных члеников, оставляя четвертый и пятый. Базальный тарсомер иногда заметно увеличен и отличается от базитарзуса. На нижней поверхности субсегментов предплюсны у некоторых прямокрылых есть небольшие подушечки, тарзальные пульвиллы или эуплантулы. У личинок насекомых предплюсна иногда сливается с голенью, образуя тибиотарзальный сегмент; в некоторых случаях кажется, что он исчез или превратился в зачаток между большеберцовой костью и претарсусом.[38]:165–166
По большей части бедренная и большеберцовая кость - самые длинные сегменты ног, но различия в длине и прочности каждого сегмента связаны с их функциями. Например, грессориальные и бегающие насекомые или насекомые ходящего и бегущего типов соответственно обычно имеют хорошо развитые бедра и большеберцовые кости на всех ногах, тогда как прыгающие (скачковые) насекомые, такие как кузнечики, имеют непропорционально развитые метафеморы и метатибии. У водных жуков (Жесткокрылые ) и ошибки (Hemiptera ), большеберцовые кости и / или лапки одной или нескольких пар ног обычно модифицированы для плавания (естественные) с бахромой из длинных тонких волосков. Многие наземные насекомые, такие как слепыши (Orthoptera: Gryllotalpidae ), нимфальные цикады (Hemiptera: Цикадовые ), и жуки-скарабеи (Scarabaeidae ), голени передних ног (protibiae) увеличены и изменены для рытья (окаменелости), тогда как передние ноги некоторых хищных насекомых, таких как златоглазки богомола (Neuroptera ) и богомолов (Mantodea ), специализируются на захвате добычи или хищниках. Большеберцовая кость и базальный членик каждой задней ноги медоносных пчел модифицированы для сбора и переноса пыльцы.[26]:45
Брюшная полость
План брюшка взрослого насекомого обычно состоит из 11–12 сегментов и менее склеротизирован, чем голова или грудная клетка. Каждый сегмент живота представлен склеротизованной тергумом, грудиной и, возможно, плевритом. Терги отделены друг от друга и от прилегающей грудины или плевры перепонкой. Дыхальца расположены в плевральной области. Вариация этого плана включает слияние терги или терги и грудины с образованием непрерывных спинных или брюшных щитков или конической трубки. Некоторые насекомые несут в плевральной области склерит, называемый латеротергитом. Вентральные склериты иногда называют латеростерниты. На эмбриональной стадии многих насекомых и на постэмбриональной стадии примитивных насекомых имеется 11 сегментов брюшка. У современных насекомых наблюдается тенденция к уменьшению числа брюшных сегментов, но в эмбриогенезе сохраняется примитивное число - 11, а количество брюшных сегментов значительно варьируется. Если Apterygota считается показателем наземного плана для крыловидных животных, царит путаница: у взрослых Protura 12 сегментов, у Collembola - 6. У семейства прямокрылых Acrididae 11 сегментов, а у ископаемого экземпляра Zoraptera брюшко с 10 сегментами.[8]
В целом первые семь брюшных сегментов взрослых (прегенитальные сегменты) похожи по строению и не имеют придатков. Однако у аптериготов (щетинохвостов и чешуйниц) и многих незрелых водных насекомых есть брюшные придатки. Аптериготы обладают парой стилей; рудиментарные придатки, серийно гомологичные дистальной части грудных ног. И, мезально, одна или две пары выступающих (или эксертильных) пузырьков, по крайней мере, на некоторых сегментах брюшной полости. Эти везикулы происходят из эндитов тазобедренного и вертлужного отдела (внутренних кольчатых долей) предковых придатков брюшной полости. Водные личинки и нимфы могут иметь жабры сбоку от некоторых до большинства сегментов брюшка.[1]:49 Из остальных сегментов брюшка состоят из репродуктивной и анальной частей.
Анально-генитальная часть брюшка, известная как терминальные, обычно состоит из сегментов 8 или 9 до верхушки брюшка. Сегменты 8 и 9 несут гениталии; сегмент 10 виден как полный сегмент у многих «нижних» насекомых, но всегда без придатков; а малый сегмент 11 представлен спинным эпипроктом и парой вентральных парапроктов, отходящих от грудины. Пара придатков, церки, сочленяется латерально на 11-м сегменте; обычно они кольцевидные и нитевидные, но были модифицированы (например, щипцы уховерток) или уменьшены в зависимости от отрядов насекомых. Кольцевидная хвостовая нить, median appendix dorsalis, возникает из кончика эпипрокта у аптеригот, большинства поденок (Ephemeroptera) и некоторых ископаемых насекомых. Подобная структура у нимфальных веснянок (Plecoptera) не имеет гомологии. Эти терминальные сегменты брюшка имеют выделительную и сенсорную функции у всех насекомых, но у взрослых есть дополнительная репродуктивная функция.[1]:49
Наружные половые органы

Органы, непосредственно связанные со спариванием и отложением яиц, вместе известны как наружные гениталии, хотя они могут быть в основном внутренними. Компоненты наружных гениталий насекомых очень разнообразны по форме и часто имеют значительную таксономическую ценность, особенно среди видов, которые кажутся структурно похожими в других отношениях. Мужские наружные гениталии широко используются для различения видов, в то время как женские наружные гениталии могут быть более простыми и менее разнообразными.
Терминалы взрослых самок насекомых включают внутренние структуры для приема копулятивного органа самца и его сперматозоидов, а также внешние структуры, используемые для яйцекладки (откладка яиц; раздел 5.8). У большинства самок насекомых есть трубка для откладывания яиц, или яйцеклад; он отсутствует у термитов, паразитических вшей, многих Plecoptera и большинства Ephemeroptera. Яйцекладчики бывают двух видов:
- истинная, или аппендикулярная, образована от придатков 8 и 9 брюшных сегментов;
- замещающий, состоит из расширяемых задних брюшных сегментов.
Другие Придатки
Внутренний
Нервная система
В нервная система насекомого можно разделить на мозг и брюшной нервный тяж. Головная капсула состоит из шести сросшихся сегментов, каждый с парой ганглии, или кластер нервных клеток вне мозга. Первые три пары ганглиев сливаются с мозгом, а следующие три пары сливаются в структуру из трех пар ганглиев под мышечной оболочкой насекомого. пищевод, называется подэзофагеальный ганглий.[1]:57
Грудные сегменты имеют по одному узлу с каждой стороны, которые соединены в пару, по одной паре на сегмент. Такое расположение также наблюдается в брюшной полости, но только в первых восьми сегментах. Многие виды насекомых уменьшили количество ганглиев из-за слияния или сокращения.[42] У некоторых тараканов в брюшной полости всего шесть ганглиев, тогда как у осы Vespa crabro имеет только два в грудной клетке и три в брюшной полости. Некоторые насекомые, такие как домашняя муха Musca domestica, все ганглии тела слиты в один большой грудной ганглий.
По крайней мере, у некоторых насекомых есть ноцицепторы, клетки, которые обнаруживают и передают ощущения боль.[43] Это было обнаружено в 2003 году при изучении вариации реакций личинки мухи обыкновенной Дрозофила на ощупь нагретый зонд и ненагретый. Личинки реагировали на прикосновение нагретого зонда со стереотипным поведением при качении, которое не проявлялось, когда личинки касались ненагретым зондом.[44] Хотя ноцицепция была продемонстрирована у насекомых, нет единого мнения, что насекомые чувствуют боль сознательно.[45]
Пищеварительная система
Насекомое использует свою пищеварительную систему на всех этапах обработки пищи: переваривание, абсорбция, доставка и удаление фекалий.[46][47] Большая часть этой пищи попадает в организм в виде макромолекулы и другие сложные вещества, такие как белки, полисахариды, жиры, и нуклеиновые кислоты. Эти макромолекулы должны быть разбиты катаболические реакции на более мелкие молекулы, такие как аминокислоты и простые сахара до того, как он будет использован клетками тела для получения энергии, роста или воспроизводства. Этот процесс расщепления известен как пищеварение. Основная структура пищеварительной системы насекомого - длинная закрытая трубка, называемая пищеварительный тракт (или кишка), которая проходит через тело вдоль тела. Пищеварительный тракт направляет пищу в одном направлении: от рот к анус. В кишечнике происходит пищеварение почти всех насекомых. Его можно разделить на три части - передняя кишка, средняя кишка и задняя кишка - каждый из которых выполняет свой процесс пищеварения. [48] Помимо пищеварительного тракта у насекомых также есть парные слюнные железы и слюнные резервуары. Эти структуры обычно располагаются в грудной клетке рядом с передней кишкой.[1]:70–77
Передняя кишка

Первый отдел пищеварительного тракта - это передняя кишка (элемент 27 на пронумерованной схеме), или стоматотеум. Передняя кишка выстлана кутикулярной подкладкой из хитин и белки как защита от жесткой пищи. Передняя кишка включает Полость рта (рот), глотка, пищевод, и Обрезать и преджелудок (любая часть может быть сильно изменена), которые одновременно хранят пищу и указывают на то, когда следует продолжать переход к средней кишке.[1]:70 Здесь начинается пищеварение, так как частично пережеванная пища расщепляется слюной из слюнных желез. Поскольку слюнные железы производят жидкость и ферменты, переваривающие углеводы (по большей части амилазы ), сильные мышцы глотки перекачивают жидкость в ротовую полость, смазывая пищу, как это делает слюнарий, и помогая питателям крови, а также питателям ксилемы и флоэмы.
Оттуда глотка передает пищу в пищевод, который может быть простой трубкой, передающей ее зобу и провентрикулюсу, а затем по направлению к средней кишке, как у большинства насекомых. С другой стороны, передняя кишка может расшириться и превратиться в очень увеличенный зоб и преджелудок, или зоб может быть просто дивертикул, или заполненная жидкостью структура, как у некоторых видов двукрылых.[49]:30–31

В слюнные железы (элемент 30 на пронумерованной диаграмме) во рту насекомого выделяется слюна. Слюнные протоки ведут от желез к резервуарам, а затем через голову продвигаются к отверстию, называемому слюнарием, которое находится за гипофаринксом. Перемещая ротовой аппарат (элемент 32 на пронумерованной диаграмме), насекомое может смешивать пищу со слюной. Затем смесь слюны и пищи попадает по слюнным трубам в рот, где начинает разрушаться.[46][50] Некоторые насекомые, например мухи, имеют экстраоральное пищеварение. Насекомые, использующие экстраоральное пищеварение, выделяют пищеварительные ферменты в пищу, чтобы расщепить ее. Эта стратегия позволяет насекомым извлекать значительную часть доступных питательных веществ из источника пищи.[49]:31
Midgut
Как только пища покидает урожай, она попадает в средняя кишка (элемент 13 на пронумерованной диаграмме), также известный как мезентерон, где происходит большая часть пищеварения. Микроскопические выступы из стенки средней кишки, называемые микроворсинки, увеличивают площадь поверхности стены и позволяют впитывать больше питательных веществ; они, как правило, близки к истоку средней кишки. У некоторых насекомых роль микроворсинок и их расположение могут различаться. Например, специализированные микроворсинки, продуцирующие пищеварительные ферменты, с большей вероятностью могут находиться в конце средней кишки, а абсорбция - у истока или в начале средней кишки.[49]:32
Задняя кишка
в задняя кишка (элемент 16 на пронумерованной диаграмме) или proctodaeum, непереваренные частицы пищи соединены мочевая кислота образовывать фекальные гранулы. Прямая кишка поглощает 90% воды, содержащейся в этих фекальных гранулах, а затем сухие гранулы выводятся через задний проход (элемент 17), завершая процесс пищеварения. Мочевая кислота образуется из продуктов жизнедеятельности гемолимфы, диффундирующих из Мальпигиевы канальцы (элемент 20). Затем он опорожняется прямо в пищеварительный тракт, на стыке между средней и задней кишками. Количество мальпигиевых канальцев, которыми обладает данное насекомое, варьируется между видами, от всего лишь двух канальцев у некоторых насекомых до более 100 канальцев у других.[1]:71–72, 78–80
Дыхательные системы
Дыхание насекомых достигается без легкие. Вместо этого дыхательная система насекомых использует систему внутренних трубок и мешочков, через которые газы либо диффундируют, либо активно перекачиваются, доставляя кислород непосредственно к тканям, которые в нем нуждаются, через их трахея (элемент 8 на пронумерованной схеме). Поскольку кислород доставляется напрямую, система кровообращения не используется для переноса кислорода и поэтому значительно сокращается. Система кровообращения насекомых не имеет вены или же артерии, а вместо этого состоит из немногим более одной перфорированной спинной трубки, которая пульсирует перистальтически. По направлению к грудной клетке спинная трубка (элемент 14) делится на камеры и действует как сердце насекомого. Противоположный конец спинной трубки похож на аорту насекомого, циркулирующего по гемолимфа, жидкий аналог членистоногих кровь, внутри полости тела.[1]:61–65[51] Воздух забирается через отверстия по бокам живота, называемые дыхальца.
Есть много разных моделей газообмен демонстрируется разными группами насекомых. Характер газообмена у насекомых может варьироваться от непрерывного до непрерывного. диффузный вентиляция, чтобы прерывистый газообмен.[1]:65–68 При непрерывном газообмене кислород принимается и углекислый газ выпускается в непрерывном цикле. Однако при прерывистом газообмене насекомое поглощает кислород, пока оно активно, и небольшое количество углекислого газа выделяется, когда насекомое находится в состоянии покоя.[52] Диффузионная вентиляция - это просто форма непрерывного газообмена, который происходит при распространение вместо того, чтобы физически поглощать кислород. Некоторые виды насекомых, которые находятся под водой, также обладают адаптациями, способствующими дыханию. Как личинки, у многих насекомых есть жабры, которые могут извлекать растворенный в воде кислород, в то время как другим необходимо подниматься на поверхность воды, чтобы восполнить запасы воздуха, которые могут удерживаться или удерживаться в специальных структурах.[53][54]
Сердечно-сосудистая система
Кровь насекомого или гемолимфа является транспортной функцией и омывает органы насекомого. Составляя обычно менее 25% веса тела насекомого, он переносит гормоны, питательные вещества и отходы и играет роль в осморегуляции, контроле температуры, иммунитет, хранилище (вода, углеводы и жиры) и функции скелета. Он также играет важную роль в процессе линьки.[55][56] Дополнительная роль гемолимфы в некоторых отрядах может заключаться в защите от хищников. Он может содержать неприятные на вкус и неприятный запах химические вещества, которые отпугивают хищников.[1] Гемолимфа содержит молекулы, ионы и клетки;[1] регулирование химического обмена между ткани гемолимфа заключена в полость тела насекомого или гемоцель.[1][57] Он переносится по телу комбинированным сердечным (задним) и аорта (передние) пульсации, которые расположены дорсально прямо под поверхностью тела.[1][55][56] Он отличается от позвоночное животное кровь в том смысле, что она не содержит эритроцитов и, следовательно, не обладает высокой кислородной способностью и больше похожа лимфа встречается у позвоночных.[1][57]
Жидкости организма поступают через устья с односторонним клапаном, которые представляют собой отверстия, расположенные по длине объединенной аорты и сердечного органа. Перекачивание гемолимфы происходит за счет волн перистальтического сокращения, берущих начало в заднем конце тела, накачиваясь вперед в спинной сосуд, через аорту и затем в голову, где она вытекает в гемоцель.[1][57] Гемолимфа циркулирует в придатках однонаправленно с помощью мышечных насосов или дополнительных пульсирующих органов, обычно находящихся в основании усики или крылья, а иногда и ноги,[1] скорость откачки увеличивается с периодами повышенной активности.[56] Движение гемолимфы особенно важно для терморегуляции в таких случаях, как Одоната, Чешуекрылые, Перепончатокрылые и Двукрылые.[1]
Эндокринная система
Эти железы являются частью эндокринной системы:
1. Нейросекреторные клетки
2. Корпорация кардиака
Репродуктивная система
женский
Самки насекомых способны производить яйца, получать и хранить сперму, манипулировать спермой разных самцов и откладывать яйца. Их репродуктивные системы состоят из пары яичники, добавочные железы, одна или несколько сперматека, и воздуховоды, соединяющие эти части. Яичники вырабатывают яйца, а дополнительные железы вырабатывают вещества, которые помогают упаковать и отложить яйца. Сперматеки хранят сперму в течение различных периодов времени и вместе с порциями яйцеводы, может контролировать использование спермы. Протоки и сперматеки выстланы кутикулой.[8]:880
Яичники состоят из ряда яйцевых трубок, называемых овариолы, которые различаются по размеру и количеству в зависимости от вида. Количество яиц, которые способно произвести насекомое, зависит от количества овариол, причем скорость развития яиц также зависит от конструкции овариол. В мероистических яичниках будущие яйца многократно делятся, и большинство дочерних клеток становятся вспомогательными клетками для одного ооцит в кластере. В паноистических яичниках каждая яйцеклетка, которая должна быть произведена стволовыми зародышевыми клетками, развивается в ооцит; из зародышевой линии нет клеток-помощников. Производство яиц панорамными яичниками, как правило, медленнее, чем у мероистических яичников.[8]:880
Добавочные железы или железистые части яйцеводов производят различные вещества для поддержания спермы, транспортировки и оплодотворения, а также для защиты яйцеклеток. Они могут производить клей и защитные вещества для покрытия яиц или жесткие покрытия для партии яиц, называемой оотека. Сперматеки - это трубочки или мешочки, в которых сперма может храниться в период между спариванием и оплодотворением яйцеклетки. Тестирование на отцовство насекомых показало, что некоторые и, вероятно, многие самки насекомых используют сперматеку и различные протоки для контроля или смещения сперматозоидов в пользу одних самцов по сравнению с другими.[8]:880
Мужской
Основным компонентом мужской репродуктивной системы является яички, подвешенный в полости тела на трахеи и толстое тело. Более примитивный аптериго у насекомых один семенник, а у некоторых чешуекрылые два созревающих семенника вторично сливаются в одну структуру на более поздних стадиях развития личинок, хотя протоки, ведущие от них, остаются отдельными. Однако у большинства самцов насекомых есть пара семенников, внутри которых находятся семенные трубки или фолликулы, заключенные в перепончатый мешок. Фолликулы соединяются с семявыносящим протоком через семявыносящие протоки, а два трубчатых семявыносящих протока соединяются со срединным семяпроводящим протоком, ведущим наружу. Часть семявыносящего протока часто увеличивается с образованием семенного пузырька, в котором хранятся сперматозоиды, прежде чем они попадут в самку. Семенные пузырьки имеют железистую оболочку, которая выделяет питательные вещества для питания и поддержания спермы. Эякуляторный проток образуется в результате инвагинации эпидермальных клеток во время развития и, как следствие, имеет кутикулярную выстилку. Терминальная часть семявыбрасывающего протока может быть склеротизирована с образованием интромитентного органа, эдеагус. Остальная часть мужской репродуктивной системы происходит из эмбриональной мезодермы, за исключением половых клеток или сперматогония, которые спускаются от первичных полюсных клеток очень рано во время эмбриогенеза.[8]:885 Эдеагус может быть довольно выраженным или de minimis. Основанием эдеагуса может быть частично склеротизированная фаллотека, также называемая фаллосомой или текой. У некоторых видов фаллотека содержит пространство, называемое эндосомой (внутренний удерживающий мешок), в которое может быть втянут (втянут) верхний конец эдеагуса.Семявыносящий проток иногда втягивается (загибается) в фаллотеку вместе с семенным пузырьком.[60][61]
Внутренняя морфология разных таксонов
Blattodea
Тараканы наиболее распространены в тропических и субтропический климат. Некоторые виды тесно связаны с человеческими жилищами и широко распространены среди мусора или на кухне. Тараканы обычно всеядный за исключением дерево - поедание таких видов, как Криптоцерк; эти тараканы не переваривают целлюлоза сами, но имеют симбиотические отношения с различными простейшие и бактерии которые переваривают целлюлозу, позволяя извлекать питательные вещества. Сходство этих симбионтов в роду Криптоцерк тем в термиты таковы, что было высказано предположение, что они более тесно связаны с термиты чем другим тараканам,[62] и текущие исследования решительно поддерживают эту гипотезу отношений.[63] Все изученные до сих пор виды несут облигатную мутуалистическую эндосимбионтную бактерию. Blattabacterium, за исключением Ноктикола австралийская, австралийский вид, обитающий в пещерах, без глаз, пигмента и крыльев, который, как показывают недавние генетические исследования, является очень примитивным тараканом.[64][65]
Тараканы, как и все насекомые, дышат через систему трубок, которая называется трахеи. Трахеи насекомых прикрепляются к дыхальца, исключая голову. Таким образом, тараканы, как и все насекомые, не зависят от рта и дыхательного горла. Клапаны открываются, когда CO2 уровень у насекомого поднимается до высокого уровня; затем СО2 распространяется из трахеи наружу и в свежем виде О2 проникает внутрь. В отличие от позвоночные это зависит от кровь для перевозки O2 и CO2, трахеальная система направляет воздух прямо в клетки, трахеальные трубки, непрерывно ветвящиеся, как дерево, до тех пор, пока их тончайшие подразделения, трахеи, не будут связаны с каждой клеткой, позволяя газообразному кислороду растворяться в цитоплазме, лежащей поперек тонкой выстилки кутикулы трахеолы. CO2 диффундирует из клетки в трахеолу. В то время как у тараканов нет легких и, следовательно, они не дышат активно, как легкие позвоночных, у некоторых очень крупных видов мускулатура тела может ритмично сокращаться, чтобы принудительно вытеснять воздух из дыхательных путей; это можно рассматривать как форму дыхания.[66]
Жесткокрылые
В пищеварительная система жуков в основном базируется на растениях, которыми они по большей части питаются, в основном с передним средняя кишка выполнение пищеварения. Однако у хищных видов (например, Carabidae ) большая часть переваривания сельскохозяйственных культур происходит с помощью ферментов средней кишки. В Elateridae вида, хищные личинки испражняют ферменты на своей добыче, при этом пищеварение происходит внеорально.[8] Пищеварительный канал в основном состоит из короткого узкого глотка, расширенное расширение, обрезать и плохо развитый желудок. После есть средняя кишка, размеры которой различаются у разных видов, с большим количеством слепая кишка, с шарниром, разной длины. Обычно бывает от четырех до шести Мальпигиевы канальцы.[67]
В нервная система в жуках содержатся все типы насекомых, различающиеся у разных видов. Три грудных и семь или восемь брюшных ганглиев можно отличить от ганглиев, в которых все грудные и брюшные ганглии слиты в единую структуру. Кислород получается через трахеальная система. Воздух входит в серию трубок вдоль тела через отверстия, называемые дыхальца, а затем превращается во все более тонкие волокна.[8] Качающие движения тела заставляют воздух проходить через систему. Некоторые виды ныряющие жуки (Dytiscidae) носите с собой пузырь воздуха, когда ныряете под поверхность воды. Этот пузырь может удерживаться под надкрыльями или прижиматься к телу специальными волосками. Пузырь обычно покрывает одно или несколько дыхальцев, поэтому насекомое может дышать воздухом из пузырька, находясь под водой. Воздушный пузырек обеспечивает насекомое лишь кратковременное снабжение кислородом, но благодаря своим уникальным физическим свойствам кислород будет диффундировать в пузырек и вытеснять азот, что называется пассивная диффузия Однако объем пузыря со временем уменьшается, и жуку придется вернуться на поверхность.[68]
Как и другие виды насекомых, у жуков есть гемолимфа вместо кровь. В открытая система кровообращения Жука приводится в движение трубчатым сердцем, прикрепленным к верхней части грудной клетки.
Разные железы специализируются на разных феромонах, вырабатываемых для поиска партнеров. Феромоны из видов Рутелинея производятся из эпителиальные клетки выстилают внутреннюю поверхность апикальных сегментов брюшной полости или феромоны на основе аминокислот Melolonthinae от выворачиваемых желез на верхушке брюшной полости. Другие виды производят разные типы феромонов. Дерместиды производить сложные эфиры, и виды Elateridae производить альдегиды, производные жирных кислот и ацетаты.[8] Также для поиска партнера светлячки (Lampyridae) использовали модифицированные клетки жирового тела с прозрачными поверхностями, покрытыми отражающими кристаллами мочевой кислоты для биосинтетического производства света, или биолюминесценция. Световой продукт очень эффективен, так как его получают путем окисления люциферин ферментами люцифераза в присутствии АТФ (аденозинтрифосфат) и кислород, продуцирующий оксилюциферин, углекислый газ и свет.[8]
У значительного числа видов развились специальные железы, вырабатывающие химические вещества для отпугивания хищников (см. Защита и хищничество ). В Жужелица (у Carabidae) защитные железы, расположенные сзади, производят множество углеводороды, альдегиды, фенолы, хиноны, сложные эфиры, и кислоты выпущен из отверстия в конце живота. Африканские жужелицы (например, Anthia некоторые из которых раньше составляли род Термофилум) используют те же химические вещества, что и муравьи: муравьиная кислота.[31] Пока Жуки-бомбардиры имеют хорошо развитые, как и другие жужелицы, пигидиальные железы, выходящие за пределы боковых краев межсегментных перепонок между седьмым и восьмым сегментами брюшка. Железа состоит из двух вмещающих камер. Первый держит гидрохиноны и пероксид водорода, а во втором - только перекись водорода плюс каталазы. Эти химические вещества смешиваются и приводят к взрывному выбросу с образованием температуры около 100 ° C с разложением гидрохинона до H2 + O2 + хинон, с O2 стимулирование экскреции.[8]
Барабанные органы органы слуха. Такой орган обычно представляет собой мембрану (барабанную перепонку), натянутую на раму, поддерживаемую воздушным мешком и связанными с ним сенсорными нейронами. В отряде жесткокрылых барабанные органы описаны по крайней мере в двух семействах.[30] Несколько видов рода Cicindela в семье Cicindelidae имеют уши на дорсальной поверхности первого брюшного сегмента под крылом; два племени в семье Dynastinae (Scarabaeidae) имеют уши прямо под щитком переднеспинки или перепонкой шеи. Уши обоих семейств работают с ультразвуковыми частотами, что дает веские доказательства того, что они работают, чтобы обнаруживать присутствие летучих мышей с помощью их ультразвуковой эхолокации. Хотя жуки составляют крупный отряд и живут в различных ниши, примеры слуха у видов на удивление отсутствуют, хотя, вероятно, большинство из них просто не обнаружено.[8]
Dermaptera
В нейроэндокринная система типичен для насекомых. Есть мозг, под пищеводом ганглий, три грудных ганглия и шесть брюшных ганглиев. Сильные нейронные связи соединяют нейрогемные тела cardiaca к головному мозгу и лобному ганглию, где тесно связаны срединное тело производит ювенильный гормон III в непосредственной близости от нейрогемной дорсальной аорты. Пищеварительная система уховерток, как и у всех других насекомых, состоит из передней, средней и задней кишки, но у уховерток отсутствуют желудочные слепые железы, которые приспособлены для пищеварения у многих видов насекомых. Длинный, стройный (экстрафор) мальпигиевы канальцы можно найти между соединением средней и задней кишки.[8]
Репродуктивная система самок состоит из парных яичники, боковые яйцеводы, сперматека, а генитальная камера. В боковых протоках яйцеклетки покидают тело, а в сперматеке хранится сперма. В отличие от других насекомых, гонопора, или генитальное отверстие находится за седьмым брюшным сегментом. Яичники примитивны в том смысле, что они политрофны ( камеры медсестры и ооциты чередовать по длине овариола ). У некоторых видов эти длинные овариолы ответвляются от бокового протока, тогда как у других вокруг протока появляются короткие овариолы.[8]
Двукрылые
Гениталии самок мух повернуты в разной степени по сравнению с положением других насекомых. У некоторых мух это временное вращение во время спаривания, но у других - постоянное скручивание органов, которое происходит на стадии куколки. Это скручивание может привести к анус располагаются ниже гениталий или, в случае перекручивания на 360 °, семявыносящего протока, охватывающего кишку, несмотря на то, что внешние органы находятся в своем обычном положении. Когда мухи спариваются, самец сначала летит поверх самки, смотря в том же направлении, но затем разворачивается, чтобы смотреть в противоположном направлении. Это заставляет самца лечь на спину, чтобы его гениталии оставались связанными с гениталиями самки, или перекручивание гениталий самца позволяет самцу спариваться, оставаясь в вертикальном положении. Это приводит к тому, что мухи обладают большей способностью к размножению, чем большинство насекомых, и гораздо быстрее. Мухи появляются большими популяциями из-за их способности эффективно и быстро спариваться, особенно в период спаривания.[69]
Самка откладывает яйца как можно ближе к источнику пищи, а развитие происходит очень быстро, что позволяет личинке съесть как можно больше пищи за короткий период времени, прежде чем превратиться во взрослую особь. Яйца вылупляются сразу после кладки, или мухи яйцекладущие, с личинкой, вылупившейся внутри матери.[69] Личинки мухи, или личинки, у них нет настоящих ног и небольшая демаркация между грудной клеткой и брюшная полость; у более производных видов голова четко не отличается от остального тела. Личинки без конечностей или имеют небольшие бездельники. Глаза и усики уменьшены или отсутствуют, а на брюшке также отсутствуют такие придатки, как Cerci. Это отсутствие функций является адаптацией к среде, богатой продуктами питания, например, в гниющих органических веществах или в качестве эндопаразит.[69] В куколки принимают различные формы, а в некоторых случаях развиваются внутри шелкового кокона. После выхода из куколки взрослая муха редко живет дольше нескольких дней и служит в основном для размножения и расселения в поисках новых источников пищи.
Чешуекрылые
В репродуктивная система бабочек и мотыльков, самец гениталии сложны и неясны. У самок различают три типа гениталий на основе соответствующих таксонов: монотризий, экзопорианский и дитрезий. У монотризного типа есть отверстия на сросшихся сегментах 9 и 10 грудины, которые служат для осеменения и откладки яиц. В экзопорианском типе (в Гепиалоидея и Mnesarchaeoidea ) есть два отдельных места для осеменения и откладки яиц, причем оба происходят на той же груди, что и монотризный тип, 9/10. У большинства видов гениталии окружены двумя мягкими лопастями, хотя у некоторых видов они могут быть специализированы и склеротизированы для откладывания яиц в таких областях, как щели и внутренняя ткань растений.[67] Гормоны и железы, которые их вырабатывают, управляют развитием бабочек и мотыльков по мере их жизненного цикла, называемого эндокринная система. Первый гормон насекомых PTTH (Проторацикотропный гормон) управляет жизненным циклом и диапаузой вида (см. раздел относится ).[70] Этот гормон вырабатывается Корпуса аллата и Корпорация Кардиака, где он также хранится. Некоторые железы специализируются на выполнении определенных задач, таких как производство шелка или слюноотделение в пальпах.[1]:65, 75 В то время как сердечные тела производят ПТТГ, аллатные тела также производят эванильные гормоны, а проторозные железы производят гормоны линьки.
в пищеварительная система, передняя часть передней кишки была изменена, чтобы сформировать глоточный всасывающий насос, поскольку он нужен им для еды, которую они едят, которая по большей части является жидкостью. An пищевод следует и ведет к задней части глотки и у некоторых видов образует форму урожая. Средняя кишка короткая и прямая, а задняя длиннее и извилистая.[67] Предки чешуекрылых, происходящие от Перепончатокрылые, имел слепые кишки средней кишки, хотя они теряются у современных бабочек и мотыльков. Вместо этого все пищеварительные ферменты, кроме начального пищеварения, иммобилизуются на поверхности клеток средней кишки. У личинок длинношеие и черешковые бокаловидные клетки находятся в переднем и заднем отделах средней кишки соответственно. У насекомых бокаловидные клетки выделяют положительный калий ионы, которые поглощаются листьями, попадающими в организм личинок. Большинство бабочек и мотыльков демонстрируют обычный пищеварительный цикл, однако виды, которые придерживаются другой диеты, требуют адаптации для удовлетворения этих новых требований.[8]:279
в сердечно-сосудистая система, гемолимфа, или кровь насекомых, используется для циркуляции тепла в форме терморегуляция, где сокращение мышц производит тепло, которое передается остальному телу в неблагоприятных условиях.[71] У чешуекрылых гемолимфа циркулирует по венам на крыльях посредством пульсирующего органа в той или иной форме, либо через сердце, либо за счет поступления воздуха в дыхательные пути. трахея.[1]:69 Воздух всасывается через дыхальца по бокам брюшной полости и грудной клетки, снабжая трахею кислородом, когда он проходит через чешуекрылые. дыхательная система. Существует три различных трахеи, снабжающих кислородом, распространяющим кислород по всему телу вида: дорсальная, вентральная и висцеральная. Дорсальные трахеи снабжают кислородом спинную мускулатуру и сосуды, в то время как брюшные трахеи снабжают вентральную мускулатуру и нервный шнур, а висцеральные трахеи снабжают кишечник, жировые тела и гонады.[1]:71, 72
Смотрите также
- Морфология (биология)
- Физиология насекомых
- Внешняя морфология Lepidoptera
- Внешняя морфология Odonata
- Экология насекомых
- Полет насекомых
- Беспозвоночные
- Насекомое
- Энтомология
- Доисторическое насекомое
Рекомендации
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах ай эй ак Gullan, P.J .; P.S. Крэнстон (2005). Насекомые: очерк энтомологии (3-е изд.). Оксфорд: издательство Blackwell Publishing. ISBN 1-4051-1113-5.
- ^ "Зоопарк насекомых им. О. Оркина". Отделение энтомологии Университета Небраски. Архивировано из оригинал на 2009-06-02. Получено 2009-05-03.
- ^ Реш, Винсент Х .; Карде, Ринг Т. (2009). Энциклопедия насекомых (2-е изд.). Сан-Диего, Калифорния: Academic Press. п. 12.
- ^ Кэмпбелл, Н. А. (1996) Биология (4-е издание) Бенджамин Каммингс, Новая работа. п. 69 ISBN 0-8053-1957-3
- ^ Джин Крицкий. (2002). Обзор энтомологии. iUniverse. ISBN 978-0-595-22143-1.
- ^ а б «внешняя морфология насекомых» (PDF). Архивировано из оригинал (PDF) на 2011-07-19. Получено 2011-03-20.
- ^ а б Richards, O.W .; Дэвис, Р. (1977). Общий учебник энтомологии Иммса: Том 1: Структура, физиология и развитие Том 2: Классификация и биология. Берлин: Springer. ISBN 0-412-61390-5.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Реш, Винсент Х .; Ринг Т. Карде (1 июля 2009 г.). Энциклопедия насекомых (2-е изд.). США: Academic Press. ISBN 978-0-12-374144-8.
- ^ Смит, Джон Бернхард, Объяснение терминов, используемых в энтомологии, Издательство: Бруклинское энтомологическое общество, 1906 г. (можно загрузить с: https://archive.org/details/explanationofter00smit )
- ^ Фокс, Ричард (6 октября 2006 г.). «Внешняя анатомия». Лендерский университет. Архивировано из оригинал на 2011-03-14. Получено 2011-03-20.
- ^ «Архивная копия» (PDF). Архивировано из оригинал (PDF) на 2008-10-01. Получено 2011-03-20. Цитировать журнал требует
| журнал =(помощь)CS1 maint: заархивированная копия как заголовок (связь) - ^ Мейер-Рохов, В. (1974). «Строение и функции личиночного глаза личинки пилильщика. Перга". Журнал физиологии насекомых. 20 (8): 1565–1591. Дои:10.1016/0022-1910(74)90087-0. PMID 4854430.
- ^ Völkel, R .; Eisner, M .; Weible, K.J. (2003). «Миниатюрные системы визуализации». Микроэлектронная инженерия. 67–68: 461–472. Дои:10.1016 / S0167-9317 (03) 00102-3. ISSN 0167-9317.
- ^ Гатен, Эдвард (1998). «Оптика и филогения: есть ли понимание? Эволюция суперпозиции глаз у Decapoda (Crustacea)». Вклад в зоологию. 67 (4): 223–236. Дои:10.1163/18759866-06704001.
- ^ Ричи, Александр (1985). "Ainiktozoon loganense Скорфилд, протохордат? из силурия Шотландии ". Алчеринга. 9 (2): 137. Дои:10.1080/03115518508618961.
- ^ Майер, Г. (2006). «Строение и развитие глаз онихофора: каков зрительный орган у членистоногих?». Строение и развитие членистоногих. 35 (4): 231–245. Дои:10.1016 / j.asd.2006.06.003. PMID 18089073.
- ^ а б c d Чепмен, Р.Ф. (1998). Насекомые: строение и функции (4-е изд.). Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 0521570484.
- ^ Krause, A.F .; Винклер, А .; Дюрр В. (2013). «Центральный привод и проприоцептивный контроль движений усиков у насекомых-трости». Журнал физиологии, Париж. 107 (1–2): 116–129. Дои:10.1016 / j.jphysparis.2012.06.001. PMID 22728470. S2CID 11851224.
- ^ Окада, Дж; То, Y (2001). «Периферическое изображение ориентации антенн на волосяной пластине скапала таракана Periplaneta americana». Журнал экспериментальной биологии. 204 (Pt 24): 4301–4309. PMID 11815654.
- ^ Staudacher, E .; Gebhardt, M.J .; Дюрр В. (2005). «Движения антенн и механорецепция: нейробиология активных тактильных датчиков». Достижения в физиологии насекомых. 32: 49–205. Дои:10.1016 / S0065-2806 (05) 32002-9. ISBN 9780120242320.
- ^ Servadei, A .; Zangheri, S .; Масутти, Л. (1972). Entomologia generale ed Applicata. CEDAM. С. 492–530.
- ^ Кэмпбелл, Фрэнк Л .; Пристли, Джун Д. (1970). «Жгутиковые кольца Blattella germanica (Dictyoptera: Blattellidae). - Изменения в их количестве и размерах во время постэмбрионального развития». Анналы энтомологического общества Америки. 63: 81–88. Дои:10.1093 / aesa / 63.1.81.
- ^ «Усики насекомых». Общество энтомологов-любителей. Получено 2011-03-21.
- ^ а б «Морфология насекомых». Университет Минизотта (факультет энтомологии). Архивировано из оригинал на 2011-03-03. Получено 2011-03-21.
- ^ Кирейчук, А.Г. (ноябрь 2002 г.). "Голова". Жуки (Coleoptera) и колеоптеролог. zin.ru. Получено 2011-03-21.
- ^ а б c d е ж грамм час я j k л м Чепмен, Р.Ф. (1998). Насекомые: структура и функции (4-е изд.). Кембридж, Нью-Йорк: Издательство Кембриджского университета. ISBN 0-521-57048-4.
- ^ а б c d е Гиллиотт, Седрик (август 1995). Энтомология (2-е изд.). Springer-Verlag New York, LLC. ISBN 0-306-44967-6.
- ^ Капур, В. С. (январь 1998 г.). Принципы и практика систематики животных. 1 (1-е изд.). Научные издательства. п. 48. ISBN 1-57808-024-X.
- ^ «Комары кусают ротовые органы». allmosquitos.com. 2011 г.. Получено 17 апреля, 2011.
- ^ а б c d Скобл, MJ. (1992). Чешуекрылые: форма, функции и разнообразие. Oxford Univ. Нажмите. ISBN 978-1-4020-6242-1.
- ^ а б Эванс, Артур V .; Беллами, Чарльз (апрель 2000 г.). Чрезмерная любовь к жукам. ISBN 0-520-22323-3.
- ^ Хеппнер, Дж. Б. (2008). «Бабочки и мотыльки». В Capinera, Джон Л. (ред.). Энциклопедия энтомологии. Виртуальная справочная библиотека Gale. 4 (2-е изд.). Ссылка Springer. п. 4345. ISBN 978-1-4020-6242-1.
- ^ Agosta, Salvatore J .; Янзен, Даниэль Х. (2004). «Распределение размеров тела крупных костариканских бабочек сухих лесов и лежащая в основе взаимосвязь между морфологией растений и опылителей». Ойкос. 108 (1): 183–193. Дои:10.1111 / j.0030-1299.2005.13504.x.
- ^ Кунте, Крушнамег (2007). «Аллометрия и функциональные ограничения на длину хоботка у бабочек» (PDF). Функциональная экология. 21 (5): 982–987. Дои:10.1111 / j.1365-2435.2007.01299.x. Архивировано из оригинал (PDF) 29 июня 2011 г.. Получено 26 февраля 2011.
- ^ «Обтирание». Университет Миниссоты. Архивировано из оригинал 27 сентября 2011 г.. Получено 17 апреля, 2011.
- ^ "Fly Mouthparts". Интернет-учебные ресурсы Школы биологических наук. Сиднейский университет. 4 февраля 2010 г.. Получено 17 апреля, 2011.
- ^ Мейер, Джон Р. (5 января 2007 г.). «Внешняя анатомия: КРЫЛЬЯ». Кафедра энтомологии, Государственный университет Северной Каролины. Архивировано из оригинал 16 июля 2011 г.. Получено 2011-03-21.
- ^ а б c d е ж грамм час я j k л м п о п q р s т Снодграсс, Р. Э. (декабрь 1993 г.). Принципы морфологии насекомых. Корнелл Унив Пресс. ISBN 0-8014-8125-2.
- ^ Spieth, HT (1932). «Новый метод изучения жилок крыльев поденок и некоторые его результаты (эфемерида)» (PDF). Энтомологические новости. Архивировано из оригинал (PDF) на 30.09.2011.
- ^ "profemur, profemora - BugGuide.Net". bugguide.net. Получено 10 декабря 2016.
- ^ "protibia - BugGuide.Net". bugguide.net. Получено 10 декабря 2016.
- ^ Шнайдерман, Ховард А. (1960). «Прерывистое дыхание у насекомых: роль дыхалец». Биол. Бык. 119 (3): 494–528. Дои:10.2307/1539265. JSTOR 1539265.
- ^ Eisemann, WK; Jorgensen, W. K .; Merritt, D. J .; Райс, М. Дж .; Cribb, B.W .; Webb, P.D .; Zalucki, M. P .; и другие. (1984). «Насекомые чувствуют боль? - биологический взгляд». Клеточные и молекулярные науки о жизни. 40 (2): 1420–1423. Дои:10.1007 / BF01963580. S2CID 3071.
- ^ Трейси, Дж; Уилсон, Род-Айленд; Laurent, G; Бензер, S; и другие. (18 апреля 2003 г.). «безболезненный, ген дрозофилы, необходимый для ноцицепции». Клетка. 113 (2): 261–273. Дои:10.1016 / S0092-8674 (03) 00272-1. PMID 12705873. S2CID 1424315.
- ^ Сёмме, Л.С. (14 января 2005 г.). «Чувствительность и боль у беспозвоночных». Норвежский научный комитет по безопасности пищевых продуктов. Получено 30 сентября, 2009.[постоянная мертвая ссылка ]
- ^ а б «Общая энтомология - пищеварительная и выделительная система». Государственный университет Северной Каролины. Получено 2009-05-03.
- ^ Terra, Walter R .; Феррейра, Клелия (2009), "Пищеварительная система", Энциклопедия насекомых, Elsevier, стр. 273–281, Дои:10.1016 / b978-0-12-374144-8.00083-7, ISBN 978-0-12-374144-8, получено 2020-09-29
- ^ Terra, Walter R .; Феррейра, Клелия (01.01.2009), «Глава 74 - Пищеварительная система», в Реш, Винсент Х .; Cardé, Ring T. (ред.), Энциклопедия насекомых (второе издание), Сан-Диего: Academic Press, стр. 273–281, Дои:10.1016 / b978-0-12-374144-8.00083-7, ISBN 978-0-12-374144-8, получено 2020-09-29
- ^ а б c Нация, Джеймс Л. (ноябрь 2001 г.). "15". Физиология и биохимия насекомых (1-е изд.). CRC Press. стр. 496с. ISBN 0-8493-1181-0.
- ^ Дункан, Карл Д. (1939). Вклад в биологию веспиновой осы Северной Америки (1-е изд.). Стэнфорд: Издательство Стэнфордского университета. С. 24–29.
- ^ Мейер, Джон Р. (17 февраля 2006 г.). "Сердечно-сосудистая система". Государственный университет Северной Каролины: Департамент энтомологии, Государственный университет Северной Каролины. п. 1. Архивировано из оригинал 27 сентября 2009 г.. Получено 2009-10-11.
- ^ Chown, S.L .; С.В. Николсон (2004). Физиологическая экология насекомых. Нью-Йорк: Издательство Оксфордского университета. ISBN 0-19-851549-9.
- ^ Ричард В. Мерритт, Кеннет В. Камминс и Мартин Б. Берг (редакторы) (2007). Знакомство с водными насекомыми Северной Америки (4-е изд.). Кендалл Хант Издательство. ISBN 978-0-7575-5049-2.CS1 maint: несколько имен: список авторов (связь) CS1 maint: дополнительный текст: список авторов (связь)
- ^ Мерритт, Р.У., К.В. Камминз и М.Б. Берг (2007). Знакомство с водными насекомыми Северной Америки. Кендалл Хант Издательская компания. ISBN 978-0-7575-4128-5.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Макгэвин, Г.С. (2001). Основная энтомология; Заказ по порядку введения. Нью-Йорк: Издательство Оксфордского университета. ISBN 9780198500025.
- ^ а б c Triplehorn, C.A .; Джонсон, Н. Ф. (2005). Введение Боррора и Делонга в изучение насекомых. Брукс / Томсон Коул: Брукс / Томсон Коул.
- ^ а б c Эльзинга, Р.Дж. (2004). Основы энтомологии (6-е изд.). Нью-Джерси, США: Пирсон / Прентис Холл.
- ^ Триплхорн, Чарльз А; Джонсон, Норман Ф (2005). Введение Боррора и Делонга в изучение насекомых (7-е изд.). Австралия: Томсон, Брукс / Коул. ISBN 9780030968358.
- ^ Gullan, P.J .; P.S. Крэнстон (2005). Насекомые: очерк энтомологии (3-е изд.). Оксфорд: издательство Blackwell Publishing. стр.61–65. ISBN 1-4051-1113-5.
- ^ Де Карло; Дж. А. (1983). "Hemipteros acuáticos y semiacuáticos. Estudio en grupos en las partes de igual función de los aparatos genitales masculinos de especies estudiadas". Revista de la Sociedad Entomológica, Аргентина. 42 (1/4): 149–154.
- ^ Андерсен, Н. Мёллер (1991). «Морские насекомые: морфология гениталий, филогения и эволюция морских конькобежцев, род Halobates (Hemiptera: Gerridae)». Зоологический журнал Линнеевского общества. 103 (1): 21–60. Дои:10.1111 / j.1096-3642.1991.tb00896.x.
- ^ Эгглтон, П. (2001). «Термиты и деревья: обзор последних достижений в филогенетике термитов». Насекомые Sociaux. 48 (3): 187–193. Дои:10.1007 / pl00001766. S2CID 20011989.
- ^ Ло, Натан; Клаудио Банди; Хирофуми Ватанабэ; Кристин Налепа; Тициана Бенинат (2003). «Доказательства кокладогенеза между различными ветвями диктиоптеранных и их внутриклеточными эндосимбионтами» (PDF). Молекулярная биология и эволюция. 20 (6): 907–913. Дои:10.1093 / molbev / msg097. PMID 12716997.
- ^ Люн, Чи Чи Люн (22 марта 2007 г.). «Пещера может содержать недостающее звено». theage.com.au. Получено 7 декабря 2013.
- ^ Lo, N; Бенинати, Т; Камень, F; Уокер, Дж; Сакки, Л. (2007). «Тараканы, которым не хватает Blattabacterium эндосимбионты: филогенетически дивергентный род Ноктикола". Письма о биологии. 3 (3): 327–30. Дои:10.1098 / rsbl.2006.0614. ЧВК 2464682. PMID 17376757.
- ^ Кункель, Джозеф Г. "Как дышат тараканы?". Часто задаваемые вопросы о тараканах. UMass Amherst. Получено 7 декабря 2013.
- ^ а б c Гилло, К. (1995). «Бабочки и мотыльки». Энтомология (2-е изд.). С. 246–266. ISBN 978-0-306-44967-3. Получено 14 ноября 2010.
- ^ Шмидт-Нильсен, Кнут (15 января 1997 г.). «Дыхание насекомых». Физиология животных: адаптация и окружающая среда (5 (иллюстрировано) изд.). Издательство Кембриджского университета. п. 55. ISBN 0-521-57098-0. Получено 6 марта 2010.
- ^ а б c Hoell, H.V., Doyen, J.T. И Перселл, A.H. (1998). Введение в биологию и разнообразие насекомых, 2-е изд.. Издательство Оксфордского университета. С. 493–499. ISBN 0-19-510033-6.CS1 maint: несколько имен: список авторов (связь)
- ^ Уильямс, К. М. (1947). «Физиология диапаузы насекомых. II. Взаимодействие куколочного мозга с переднегрудными железами при метаморфозе гигантского тутового шелкопряда» Platysamia cecropia"". Биол. Бык. 92 (2): 89–180. Дои:10.2307/1538279. JSTOR 1538279. PMID 20268135.
- ^ Lighton, J.R.B .; Лавгроув, Б.Г. (1990). «Температурное переключение с диффузионной на конвективную вентиляцию пчелы». Журнал экспериментальной биологии. 154: 509–516.
| Поля |  | |
|---|---|---|
| Бактерии | ||
| Протисты |
| |
| Растения | ||
| Беспозвоночные | ||
| Млекопитающие | ||
| Другие позвоночные | ||
| Другие темы | ||
| ||